հեղինակ՝ Չարլզ Դարվին |
ՏԵՍԱԿՆԵՐԻ ԾԱԳՈՒՄԸ
ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՄԻՋՈՑՈՎ
կամ
ՀԱՐՄԱՐՎԱԾ ՑԵՂԵՐԻ ՊԱՀՊԱՆՈՒՄԸ ԳՈՅՈՒԹՅԱՆ ԿՌՎՈՒՄ
- «Բայց նյութական աշխարհի վերաբերմամբ մենք կարող ենք ընդունել, համենայն դեպս, հետևյալը. մենք կարող ենք տեսնել, որ երևույթներն առաջանում են ոչ թե յուրաքանչյուր առանձին դեպքում իր ազդեցությունը գործող աստվածային ուժի առանձին միջամտություններից, այլ ընդհանուր օրենքների հաստատումից»։
- ՈՒԵՎԵԼԼ։ Բրիջուոտերյան տրակտատ
- ՈՒԵՎԵԼԼ։ Բրիջուոտերյան տրակտատ
- «Բնական» բառի միակ որոշակի նշանակությունն է՝ հաստատված, սահմանված կամ կարգավորված, որովհետև արդյոք բնական չէ՞ այն, ինչ պահանջում կամ ենթադրում է բանականությամբ արդարացվող մի ազդակ, որը նրան անում է այդպիսին, այսինքն՝ նրա կողմից իրագործվում է մշտապես կամ սահմանված ժամանակում, ճիշտ այնպես, ինչպես գերբնականը կամ հրաշագործականը — այն, որ իրագործվում է նրա կողմից միայն մեկ անգամ»։
- ԲԵՏԼԵՐ։ Հայտնության կրոնի անալոգիան
- ԲԵՏԼԵՐ։ Հայտնության կրոնի անալոգիան
- «Ուստի եզրակացնում ենք, որ ոչ մի մարդ, սխալ կերպով գերագնահատելով առողջ դատողությունը կամ ոչ ճիշտ հասկանալով չափավորությունը, չպետք է կարծի կամ պնդի, թե մարդը աստվածային խոսքի գրքի կամ աստծու ստեղծագործության գրքի, աստվածաբանության կամ փիլիսոփայության հետազոտության կամ ուսումնասիրության մեջ կարող է չափազանց խորը թափանցել. բայց մարդիկ թող ավելի շատ ձգտեն անսահման կատարելագործության կամ հաջողությունների թե՛ մեկում և թե՛ մյուսում»։
- ԲԵԿՈՆ։ Գիտության առաջադիմությունը
- «Բայց նյութական աշխարհի վերաբերմամբ մենք կարող ենք ընդունել, համենայն դեպս, հետևյալը. մենք կարող ենք տեսնել, որ երևույթներն առաջանում են ոչ թե յուրաքանչյուր առանձին դեպքում իր ազդեցությունը գործող աստվածային ուժի առանձին միջամտություններից, այլ ընդհանուր օրենքների հաստատումից»։
ՊԱՏՄԱԿԱՆ ԱԿՆԱՐԿ ՏԵՍԱԿՆԵՐԻ ԾԱԳՄԱՆ ՎԵՐԱԲԵՐՄԱՄԲ ԵՂԱԾ ՀԱՅԱՑՔՆԵՐԻ ՄԱՍԻՆ, ՄԻՆՉԵՎ ՍՈՒՅՆ ԱՇԽԱՏՈՒԹՅԱՆ ԱՌԱՋԻՆ ՀՐԱՏԱՐԱԿՈՒԹՅԱՆ ԼՈՒՅՍ ՏԵՍՆԵԼԸ
Այստեղ մի համառոտ ակնարկ կտամ տեսակների ծագման վերաբերմամբ եղած գիտական հայացքի հաջողությունների մասին։ Մինչև վերջին ժամանակներս բնախույզների զգալի մեծամասնությունը համոզված էր, որ տեսակները ներկայացնում են մի ինչ որ անշարժ բան և ստեղծված են մեկը մյուսից անկախ կերպով։ Շնորհալի շատ հեղինակներ վարպետորեն պաշտպանել են այդ տեսակետը։ Մյուս կողմից, սակավաթիվ բնախույզներ կարծում էին, որ տեսակները ենթարկվում են փոփոխության և որ գոյություն ունեցող կենսաձևերը սովորական սաղմնավորման միջոցով սերվել են առաջներում գոյություն ունեցած ձևերից։ Կանգ չառնելով այդ իմաստով արված անորոշ ակնարկների վրա, որոնք հանդիպում են դասական գրողների մոտ[1], պետք է ընդունել, որ նորագույն ժամանակների գրողներից այդ հարցը իսկական գիտական ոգով առաջին անգամ քննության առնողը եղել է Բյուֆոնը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 1] Բայց քանի որ նրա կարծիքները մեծ տատանումների են ենթարկվել և քանի որ նա չի շոշափել այն պատճառները կամ ուղիները, որոնց միջոցով կատարվում էր տեսակների այդ ձևափոխումը, ես այստեղ մանրամասնությունների մեջ չեմ կարող մտնել։
Լամարկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 2] առաջինն էր, որի եզրակացությունները այդ հարցի վերաբերմամբ իրենց վրա ուշադրություն գրավեցին։ Այդ, իրավամբ հռչակավոր գիտնականը առաջին անգամ շարադրեց իր հայացքները 1801 թվականին, նա զգալիորեն ընդարձակեց դրանք 1809 թվականին իր «Philosophie Zoologique» մեջ[2] և ավելի ևս ուշ՝ 1815 թ. իր «Histoire Naturelle des Animaux sans vertébres»-ի առաջաբանում[3]։ Այդ աշխատություններում նա պաշտպանում է այն տեսակետը, որ բոլոր տեսակները, ներառյալ մարդը, ծագում են առել այլ տեսակներից։ Նա մեծ ծառայություն մատուցեց, առաջինը նա սևեռեց ընդհանուրի ուշադրությունը այն ենթադրության հավանականության վրա, որ բոլոր փոփոխությունները օրգանական աշխարհում, ինչպես նաև անօրգանականում, տեղի են ունեցել բնության օրենքների հիման վրա, այլ ոչ թե հրաշագործ միջամտության հետևանքով։ Լամարկը, ըստ երևույթին, տեսակների աստիճանական փոփոխության մասին եզրակացրել է այն դժվարությունների հիման վրա, որոնք զգացվում են տեսակը տարատեսակից զանազանելիս, որոշ խմբերի ներկայացուցիչների միջև գրեթե անզգալի անցումների հիման վրա և ընտանեցված կենդանիների և կուլտուրական բույսերի համանմանության հիման վրա։ Ինչ վերաբերում է փոփոխություններ առաջացնող պատճառներին, ապա նա դրանք վերագրում էր մասամբ ֆիզիկական պայմանների անմիջական ազդեցությանը, մասամբ արդեն գոյություն ունեցող ձևերի միջև խաչաձևմանը, բայց գլխավորապես օրգանների վարժեցման կամ ոչ վարժեցման, այսինքն՝ սովորությանը։ Այս վերջին գործոնին նա, ըստ երևույթին, վերագրում էր բնության մեջ հանդիպող բոլոր հիանալի հարմարումները, ինչպիսին է ծառերի տերևներն ուտող ընձուղտի վիզը։ Բայց նա հավատում էր նաև պրոգրեսիվ զարգացման օրենքի գոյությանը, իսկ որովհետև այդ օրենքի ուժով բոլոր կենդանի էակները ձգտում են կատարելագործման, ապա ժամանակակից դարաշրջանում նաև պարզագույն ձևերի գոյությունը բացատրելու համար նա ընդունում էր նրանց ինքնածնությունը[4]։
Ժոֆֆրուա Սենտ-Իլերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 3] ինչպես երևում է նրա «Կենսագրական»-ից, որ գրել է նրա որդին, դեռ 1795 թ. կարծում էր, որ այսպես կոչված տեսակները միայն մեկ ընդհանուր տիպից զանազան շեղումներ են։ Բայց միայն 1828 թ. նա մամուլում հայտնեց իր համոզմունքը, որ աշխարհի հենց սկզբից ձևերը անփոփոխ չեն մնացել։ Ժոֆֆրուան փոփոխությունների գլխավոր պատճառը տեսնում էր գոյության պայմաններում կամ, ինչպես ինքն է արտահայտվել «le monte ambiant»[5]։ Նա զգույշ էր իր եզրակացություններում և չէր կարծում, որ գոյություն ունեցող տեսակները շարունակում են փոփոխվել, իսկ նրա որդու խոսքերով «c’est donc un problème à reserver entièrement a l’avenir, supposé même que l’avenir doive avoir prise sur lui»[6].
1813 թվականին դոկտոր Վ. Ս. Ուելսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 4] թագավորական ընկերությունում մի հաղորդում կարդաց «Սպիտակ ցեղին պատկանող մի կնոջ մասին, որի մաշկի մի մասը նման էր նեգրերի մաշկին», բայց այդ հոդվածը չտպագրվեց մինչև նրա «Երկու հետազոտություն՝ ցեղի մասին և պարզ տեսողության մասին» հռչակավոր աշխատության լույս տեսնելը, այն է՝ 1818 թվականին։ Այդ հետազոտության մեջ նա որոշակիորեն ընդունում է բնական ընտրության սկզբունքը և այդ սկզբունքը ընդունողներից առաջինն է. բայց Ուելսը այն ընդունում է միայն մարդկային ցեղերի համար՝ այն էլ միայն մի քանի հատկանիշների վերաբերմամբ։ Մատնանշելով, որ նեգրերը, և մուլատները, ըստ երևույթին, արևադարձային որոշ հիվանդությունների չեն ենթարկվում, նա նշում է, որ բոլոր կենդանիները աննշան փոփոխությունների հակում ունեն, և գյուղական տնտեսատերերը իրենց ընտանի անասուններին բարելավում են ընտրության միջոցով, և վերջապես, ավելացնում է. «Այն, ինչ վերջին դեպքում ձեռք է բերվում արհեստականորեն, հավանորեն նույնպիսի հաջողությամբ, թեև ավելի դանդաղ, կատարվում է բնության մեջ, իրենց բնակավայրերին հարմարված մարդկային տարատեսակների առաջացման պրոցեսում։ Մարդու այն պատահական տարատեսակներից, որոնք երևան են եկել միջին Աֆրիկայի նոսր, ցրված բնակչության մեջ, որևէ մեկը գուցե մնացածներից ավելի լավ հարմարված է տեղական հիվանդություններին դիմանալու տեսակետից։
Հետևաբար, այդ ցեղը կբազմանա, մինչդեռ մյուսները կնվազեն ոչ միայն հիվանդություններին դիմադրելու անընդունակության պատճառով, այլև իրենցից ավելի հզոր հարևանների հետ մրցելու հնարավորություն չունենալու հետևանքով։ Այդ ավելի հզոր ցեղի գույնը, ասվածի հիման վրա, կլինի սև։ Բայց քանի որ տարատեսակներ առաջացնելու այդ ձգտումը պահպանվում է, ուստի հետևանքը այն կլինի, որ կառաջանա ավելի ու ավելի մգագույն ցեղ, և որովհետև ամենամուգը ամենից լավ հարմարված կլինի կլիմայական պայմաններին, ապա նա էլ կդառնա գերակշռող, եթե ոչ միակ ցեղը այն երկրում, որտեղ նա երևան է եկել»։ Այնուհետև նա իր հայացքները տարածում է նաև ավելի ցուրտ երկրներում ապրող սպիտակների վրա։ Ես այդ ցուցումների համար պարտական եմ Միացյալ նահանգներում ապրող մ-ր Ռոուլեյին,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 5] որը մ-ր Բրեսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 6] միջոցով իմ ուշադրությունը հրավիրեց Ուելսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 4] աշխատությունների վրա։
Մեծարգո Վ. Հերբերտը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 7] հետագայում Մանչեստերի դեկանը, 1822 թվականին իր «Horticultural Transactions»-ի[7] չորրորդ հատորում և իր «Ամարիլազգիներ» աշխատությունում (1837 թ., էջ 19, 339) հայտնում է իր համոզմունքը, որ «պարտիզագործական փորձերը անհերքելի կերպով հաստատում են այն դրույթը, որ բուսաբանական տեսակները միայն բարձր կարգի, բայց ավելի մնայուն տարատեսակներ են»։ Նա այդ հայացքը տարածում է նաև կենդանիների վրա։ Դեկանը կարծում է, որ յուրաքանչյուր սեռում ստեղծվել է մեկական տեսակ, որը օժտված է եղել ծայրահեղ պլաստիկությամբ, և հետո արդեն այդ տեսակները մասամբ խաչաձևման միջոցով, մասամբ փոփոխության ճանապարհով առաջացրել են բոլոր գոյություն ունեցող տեսակները։
1826 պրոֆ. Գրանտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 8] Spongilla-ի մասին իր հայտնի հետազոտության եզրափակիչ պարագրաֆում (Edinbourgh Philosophical Journal[8], հատոր 14, էջ 283) միանգամայն որոշակիորեն հայտնում է իր համոզմունքը, որ տեսակներն առաջանում են այլ տեսակներից և որ իրենց փոփոխության հետ զուգընթաց նրանք կատարելագործվում են։ Նույն հայացքը նա բերում է իր 55-րդ դասախոսության մեջ, որը տպագրված է «Lancet»-ում 1834 թվականին։
1831 թվականին մ-ր Պատրիկ Մատյուն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 9] հրատարակել է իր «Նավաշինական անտառանյութի և ծառաբուծության մասին» աշխատությունը, որտեղ տեսակների ծագման վերաբերմամբ հայտնում է ճիշտ նույն հայացքը (ինչպես հիմա կտեսնենք), ինչ որ հայտնել է մ-ր Ուոլլեսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] և ես՝ «Linnean Journal»-ում[9] և մանրամասնորեն լուսաբանված է ներկա հատորում։ Դժբախտաբար մ-ր Մատյուն այդ հայացքը հայտնել է շատ համառոտ կերպով, կցկտուր դիտողությունների ձևով՝ բոլորովին այլ հարցի նվիրված մի աշխատության հավելվածում, այնպես, որ նա մնացել էր աննկատ, մինչև, որ մ-ր Մատյուն 1860 թ. ապրիլի 7-ին «Gardener’s Chronicle»-ում[10] նրա վրա ուշադրություն է հրավիրում։ Մ-ր Մատյուի և իմ հայացքների միջև եղած տարբերությունները էական չեն. նա, ըստ երևույթին, կարծում է, որ աշխարհը ժամանակ առ ժամանակ բոլորովին զրկվել է իր բնակչությունից և նորից բնակչությամբ լցվել է և, որպես նոր էակների գոյացման համար հնարավոր եղանակ, ենթադրում է, որ նրանք կարող էին ծնվել «առանց ընդունելու արդեն գոյություն ունեցած ձևերը և չառաջանալով արդեն գոյություն ունեցած սաղմնային ագրեգատներից»։ Ես վստահ չեմ, թե արդյոք լրիվ հասկացե՞լ եմ նրա գրքի որոշ տեղերը, բայց ինձ թվում է, որ նա մեծ նշանակություն է տալիս գոյության պայմանների անմիջական ներգործությանը։ Համենայն դեպս, նա պարզորոշ նկատել է բնական ընտրության սկզբունքի ամբողջ նշանակությունը։
Հռչակավոր երկրաբան և բնախույզ ֆոն-Բուխը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 11] իր «Description phisique des Isles Canaries»[11] հիանալի գրքում (1836 թ. էջ 147) պարզ կերպով հայտնում է իր համոզմունքը, որ տարատեսակները աստիճանաբար դառնում են հաստատուն տեսակներ, որոնք արդեն անընդունակ են միմյանց հետ խաչաձևելու։
Ռաֆինեսկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 12] իր «Հյուսիսային Ամերիկայի նոր բուսականությունը» աշխատությունում, որը լույս է տեսել 1836 թվականին, գրում է (էջ 6). «Բոլոր տեսակները կարող էին մի ժամանակ տարատեսակներ եղած լինել, և շատ տարատեսակներ աստիճանաբար փոխվում են, դառնում տեսակներ, ձեռք բերելով յուրահատուկ և հաստատուն հատկանիշներ», բայց այնուհետև ավելացնում է (էջ 18),— «բացառությամբ յուրաքանչյուր սեռի օրիգինալ տիպերի կամ նախահայրերի»։
1843—1844 թվականներին պրոֆ. Հոլդման[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 13] (Boston Journal of. Nat. Hist.[12] U. States, vol. IV, p. 468) շատ հմտորեն առադրել է տեսակների ձևափոխման և զարգացման հիպոթեզի վերաբերյալ թեր և դեմ փաստարկները, նա ինքը, ըստ երևույթին, հակվում է փոփոխականության կողմը։
1844 թվականին առաջին անգամ լույս են տեսել «Vestiges of Creation»[13]։ Այդ գրքի տասերորդ և զգալի չափով ուղղված հրատարակության մեջ (1853 թ.) անանուն հեղինակն ասում է (էջ 155). «Բազմաթիվ նկատառումների հիման վրա մենք գալիս ենք այն ընդհանուր եզրակացության, որ կենդանի էակների զանազան շարքերը՝ սկսած պարզագույններից ու հնագույններից և վերջացրած բարձրակարգներով ու նորագույններով, աստվածային նախախնամության գործողությամբ հանդիսանում են՝ առաջին, այն ազդակի արդյունքը, որը հաղորդվել է կենդանի էակներին և դրդում է նրանց որոշակի դարաշրջաններում անցնելու կազմվածքի որոշ աստիճաններ՝ հասնելով մինչև երկշաքիլավորները և ողնաշարավորները, ըստ որում այդ աստիճանները սակավաթիվ էին և կազմվածքում բաժանանշվում էին ընդհատումներով, որոնք գործնական դժվարություններ են ներկայացնում ձևերի փոխադարձ նմանությունը որոշելիս. երկրորդ, մի այլ ազդակի արդյունք, որը կապի մեջ է կենսական ուժերի հետ, որոնք ձգտում են սերունդների ընթացքում փոխել կազմվածքը՝ համապատասխանորեն արտաքին պայմաններին, ինչպիսիք են սնունդը, բնակավայրի հատկությունները և օդերևութաբանական պայմանները։ Այս վերջին փոփոխություններն էլ կազմում են այն, որոնք բնական աստվածաբանության մեջ կոչվում են «հարմարումներ»։ Ըստ երևույթին, հեղինակը կարծում է, որ կազմվածքը առաջադիմել է թռիչքներով, բայց գոյության պայմանների ներգործությունը եղել է աստիճանաբար։ Նա բերում է ընդհանուր բնույթ կրող շատ ուժեղ փաստեր հօգուտ այն բանի, որ տեսակները անշարժ ձևեր չեն։ Բայց չեմ պատկերացնում, թե ինչպես նրա ենթադրած երկու «ազդակները» կարող են գիտական բացատրություն տալ բնության մեջ հանդիպող բազմաթիվ հիանալի հարմարումների մասին։ Ես չեմ կարծում, որ այդ ճանապարհով մենք կարող ենք մի քայլ անգամ առաջ գնալ ըմբռնելու գործում, թե ինչպես է, օրինակ, փայտփորիկը ստացել այն բոլոր հարմարումները, որոնք անհրաժեշտ են նրա բացառիկ կենսակերպի համար։ Այդ գիրքը, իր ուժեղ և փայլուն ոճի շնորհիվ և չնայած առաջին հրատարակությունների հաղորդած տեղեկությունների որոշ անճշտությանը և գիտական շրջահայեցության պակասին, հենց սկզբից ձեռք բերեց ընթերցողների լայն շրջան։ Իմ կարծիքով, նա այս երկրում էական օգուտ տվեց, քննարկվող հարցի վրա ուշադրություն գրավելով, վերացնելով արմատացած նախապաշարումները և այսպիսով հող պատրաստելով համանման հայացքների ընդունման համար։
1846 թվականին վետերան երկրաբան Ժ. Օմալիուս դը Ալլուան[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 14] մի փոքրիկ, բայց սքանչելի հոդվածում («Bulletin de l’Acad. Roy. Bruxelles[14] t. XIII, p. 581) կարծիք է հայտնում, որ ուրիշ ձևերից փոփոխվելու միջոցով տեսակների ծագումը ավելի հավանական է, քան նրանց ծագումն առանձին ստեղծագործական գործողությունների միջոցով, հեղինակն այդ կարծիքն առաջին անգամ հայտնել է դեռ 1831 թվականին։
Պրոֆ. Օուենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] 1849 թ. («Nature of Limbs»[15] p. 86) գրել է հետևյալը. «Նախատիպերի գաղափարը մարմնում արտահայտվել է բազմազան փոփոխություններով շատ ավելի առաջ, քան այս մոլորակի վրա երևան են եկել կենդանիների այն տեսակները, որոնց մեջ նա հանդես է գալիս այժմ։ Թե ի՞նչ բնական օրենքների կամ երկրորդային պատճառների վրա են դրված այդ օրգանական երևույթների ճիշտ հաջորդականությունն ու զարգացումը, մեզ հայտնի չէ»։ 1858 թվականին Բրիտանական ասոցիացիայի նիստում իր նախագահական ճառի մեջ նա հիշատակում է (էջ LI) «ստեղծագործական ուժի անընդհատ ազդեցության աքսիոմայի, կամ կենդանի էակների նախասահմանված կենսագործման մասին»։ Այնուհետև (էջ XC), շոշափելով աշխարհագրական տարածման հարցը, նա ավելացնում է. «Այդ երևույթները մեզ ստիպում են կասկածել այն բանում, որ անգլիական կարմիր մայրահավը և նորզելանդական ապտերիքսը ստեղծված են այդ կղզիներից յուրաքանչյուրի համար առանձին և նրանց վրա։ Եվ ընդհանրապես միշտ էլ պետք է հիշել, որ կենդանաբանը, դիմելով «ստեղծված են» արտահայտությանը, դրանով ցույց է տալիս «իրեն անհայտ մի պրոցես»։ Այնուհետև նա մանրամասնորեն զարգացնում է այդ միտքը, ասելով, որ կարմիր մայրահավի նման օրինակներով, որոնք «կենդանաբանը թվարկում է որպես վկայություն այն ենթադրության օգտին, թե թռչուններն առանձին-առանձին ստեղծվել են որոշ կղզիների վրա և այդ կղզիների համար, նա միայն ցանկանում է հայտնել այն միտքը, որ չի հասկանում, թե կարմիր մայրահավը ինչպես է ընկել այնտեղ և բացառապես այնտեղ, որտեղ մենք նրան հանդիպում ենք. նրա անտեղյակությունը դրսևորող արտահայտության այդ եղանակով կենդանաբանը հայտնում է նաև իր համոզվածությունը այն բանում, որ թե՛ թռչունը և թե՛ կղզին իրենց ծագումով պարտական են մեծ ստեղծագործական նախապատճառին»։ Եթե մենք փորձենք մեկը մյուսի օգնությամբ մեկնաբանել միևնույն ճառում հանդիպող այդ երկու նախադասությունները, ապա կգանք այն եզրակացության, որ հռչակավոր գիտնականը 1858 թվականին արդեն համոզված չէր, թե ապտերիքսը կամ կարմիր մայրահավը առաջին անգամ երևացել են իրենց այժմյան գտնված տեղում՝ «անհայտ եղանակով» կամ «իրեն անհայտ պրոցեսի շնորհիվ»։
Այդ ճառն արտասանվել է այն բանից հետո, երբ մ-ր Ուոլլեսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] և իմ զեկուցագրերը, որոնց մասին հիմա կհիշատակենք, կարդացվել էին Լիննեյան ընկերությունում։ Այդ գրքի առաջին հրատարակությունը լույս տեսնելու ժամանակ ես, ուրիշ շատերի հետ միասին, այնպիսի խոր մոլորության մեջ էի ընկել «ստեղծագործական ուժի անընդհատ ազդեցություն» արտահայտությամբ, որ պրոֆ. Օուենին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] այլ հնէաբանների հետ մտցրել էի այն գիտնականների շարքը, որոնք խորապես համոզված են տեսակային ձևերի անշարժության մեջ. բայց բանից դուրս է գալիս, որ դա իմ կողմից հրեշավոր սխալ էր (Anat. of Vertebrates[16] vol. III, p. 796)։ Ներկա աշխատության վերջին հրատարակության մեջ հիմնվելով նրա գրքի այն տեղի վրա, որն սկսվում է «Կասկածի ենթակա չէ, որ տիպիկ ձևը» և այլն բառերով (Ibid., vol. I, p. XXXV), արել եմ ենթադրություն, որն այժմ էլ ինձ ճիշտ է թվում, թե պրոֆեսոր Օուենն ընդունում է, որ բնական ընտրությունը կարող էր որոշ դեր կատարել նոր տեսակների առաջացման գործում, բայց այդ ենթադրությունն էլ դուրս եկավ սխալ և անապացույց (Ibid., vol. III, p. 798)։ Ես բերել եմ քաղվածքներ պրոֆ. Օուենի և «London Review»-ի[17] հրատարակչի միջև եղած նամակագրությունից, որոնցից ակներև դարձավ, ինչպես այդ հրատարակչի, այնպես էլ ինձ համար, որ պրոֆ. Օուենը պաշտպանում էր իր առաջնության իրավունքը բնական ընտրության ուսմունքը ինձնից առաջ տարածելու գործում։ Այդ հայտարարության առթիվ ես արտահայտել եմ իմ զարմանքը և գոհունակությունը, բայց ինչքան կարելի է հասկանալ նորագույն աշխատությունների մի քանի տեղերից (Ibid,, vol-III, p. 798), ես նորից, մասամբ կամ կատարելապես, մոլորության մեջ եմ ընկել։ Կարող եմ միայն այն մտքով մխիթարվել, որ ոչ միայն ես, այլև ուրիշ մարդիկ պրոֆ. Օուենի այդ բանավեճային երկերը համարում են դժվար հասկանալի և միմյանց հետ անհաշտելի։ Իսկ ինչ վերաբերում է բնական ընտրության սկզբունքի սոսկ հայտարարությանը, ապա բոլորովին էական չէ, թե արդյոք պրոֆ. Օուենը ինձ կանխե՞լ է թե ոչ, որովհետև բերված պատմական ակնարկից ակներև է, որ Ուելսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 4] և մ-ր Մատյուն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 9] երկուսիս էլ կանխել են։
Իսիդոր Ժոֆֆրուա Սենտ-Իլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 16] 1850 թվականին կարդացած իր դասախոսություններում (որոնցից քաղվածքը լույս է տեսել «Revue et Mag. de Zoologie»-ում[18] Jan. 1851) համառոտակի բերում է այն հիմքերը, որոնք ստիպեցին ընդունելու, որ տեսակային հատկանիշներր «sont fixés pour chaque espèce, tant qu’elle se perpetue au milieu des mêmes circonstances: ils se modifient, si les circonstances ambìantes viennent à changer». «En résumé l’observation des animaux sauvages démontre déjà la variabilité limitée des espèces. Les expériences sur les animaux sauvages devenus domestiques, et sur les animaux domestiques redevenus sauvages la démontrent plus clairement encore. Ces mêmes, expériences prouvent, de plus, que les differences produites peuvent être de valeur générique»[19]. Իր «Hist. Nat. Génerale»[20], (t. II, p. 480, 1850 թ.) աշխատության մեջ նա մանրամասնորեն զարգացնում է միանգամայն նման եզրակացություններ։
Դոկտոր Ֆրեկի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 17] վերջերս հրատարակած շրջաբերականից երևում է, որ նա 1851 թ. («Dublin Medical Press»[21], p. 322) զարգացնում է այն ուսմունքը, որ բոլոր օրգանական էակները ծագել են մեկ նախասկզբնական ձևից։ Նրա հիմնական համոզմունքները և հարցի հետագա զարգացումը արմատական կերպով տարբերվում են իմիններից, և քանի որ դ-ր Ֆրեկը այժմ (1861 թ.) հրատարակել է իր «Տեսակների ծագումը օրգանական ազգակցության միջոցով» ուրվագիծը, իմ կողմից ավելորդ կլիներ ձեռնարկել նրա գաղափարների շարադրման դժվարին խնդրին։
Հերբերտ Սպենսերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 18] իր ուրվագծում (որը սկզբում լույս է տեսել «Leader»-ում 1852 թ. մարտին ու արտատպվել է նրա «Essays»[22]-ում 1858 թ.) զարմանալի ուժով և հմտությամբ համեմատում է օրգանական էակների ստեղծման և զարգացման տեսությունները։ Ելնելով ընտանի կենդանիների և մշակովի բույսերի անալոգիայից, այն փոփոխություններից, որոնց ենթարկվում են շատ տեսակների սաղմերը, այն դժվարություններից, որ զգացվում են տեսակներն ու տարատեսակները տարբերելիս, և ընդհանուր աստիճանականության սկզբունքից, նա եզրակացնում է, որ տեսակները փոփոխվել են, և նրանց փոփոխությունը վերագրում է գոյության պայմանների փոփոխությանը։ Նույն հեղինակը (1855 թ.) գրել է նաև հոգեբանություն, ելնելով այն սկզբունքից, որ բոլոր մտավոր հատկություններն ու ընդունակությունները անխուսափելիորեն ձեռք են բերվում աստիճանաբար։
1852 թվականի նշանավոր բուսաբան Նոդենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 19] տեսակների ծագման մասին իր հիանալի հոդվածում («Revue Horticole.»[23], p. 102), հետագայում մասամբ արտատպված «Novelles Archives de Museum»[24], I, p. 171, հայտնել է իր համոզմունքը, որ տեսակներն առաջանում են այն եղանակով, ինչ եղանակով առաջանում են կուլտուրական տարատեսակները, իսկ վերջին պրոցեսը նա վերագրում է մարդու կողմից գործադրվող արհեստական ընտրությանը։ Բայց նա չի մատնանշում, թե ընտրությունը ինչպես կարող է գործել բնական վիճակում։ Դեկան Հերբերտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 7] նման նա ենթադրում է, որ տեսակներն իրենց ծագման սկզբում ավելի պլաստիկ էին, քան այժմ։ Նա մեծ նշանակություն է տալիս այն բանին, որին անվանում է վախճանական պատճառի սկզբունք. «puissance mystérieuse, indéterminée; fatalité pour les uns; pour les autres volonté providentielle dont l’action incéssante sur les êtres vïvants détermine à toutes les époques de l’existence du monde la forme, le volume, et la durée de chacun d’eux, en raison de sa déstinée dans l’ordre des choses dont il fait partie. C’est cette puissance qui harmonise chaque membre à l’ensemble, en l’appropriant à la fonction qu’il doit remplir dans l’organism général de la nature, fonction, qui est pour lui sa raison d’être»[25]։
1853 թ. նշանավոր երկրաբան կոմս Կեյզերլինգը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 20] («Bulletin de la Soe Geolog»[26] 2-nd Ser., t. X, p, 357) հայտնել է այն միտքը, որ ինչպես որոշ հիվանդություններ, որոնք առաջանում են միազմների միջոցով, նորից ծագում էին ու արագ կերպով տարածվում ամբողջ աշխարհում, այնպես էլ որոշ դարաշրջաններում գոյություն ունեցող տեսակների սաղմերը կարող էին ենթարկվել իրենց շրջապատող որոշ առանձնահատուկ մոլեկուլների քիմիական ներգործությանը և նոր ձևերի սկիզբ տալ։
Նույն 1853 թ. դ-ր Շաֆհաուզենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 21] {Werhandl. des Naturhist. Vereins des Preuss Rheinlands»[27]) հրատարակել է մի հիանալի կշռադատություն, որտեղ նա ապացուցում է օրգանական ձևերի առաջընթաց զարգացումը երկրի վրա։ Նա եզրակացնում է, որ շատ տեսակներ մեծ ժամանակաշրջանների ընթացքում պահպանվել են անփոփոխ, մինչդեռ մյուսները փոփոխվել են։ Տեսակը տարբերելու հնարավորությանը նա բացատրում է միջանկյալ ձևերի անհետացմամբ։ «Այսպիսով, ապրող բույսերն ու կենդանիները անհետացածներից չեն բաժանվում որպես ստեղծագործության առանձին ակտեր և պետք է դիտվեն որպես նրանց հաջորդները, որոնք նրանցից առաջացել են չընդհատվող վերարտադրության միջոցով»։
Ֆրանսիացի նշանավոր բուսաբան Լեկոկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 22] 1854 թվականին գրում է («Etudes sur Géograph. Bot.»[28] t. I, p. 250). «On voit que nos recherches sur la fixité ou la variation de l’espèce nous conduisent directement aux idées émises par deux hommes justement célèbres, Geoffroy Saint-Hilaire et Goethe[29]. Բայց Լեկոկի ընդարձակ աշխատության մեջ ցրված մյուս նախադասությունները կասկած են հարուցում այն մասին, թե նա ինչ չափերով էր ընդունում տեսակների փոփոխականությունը։
Ստեղծագործության փիլիսոփայությունը վարպետությամբ մշակվել է հարգարժան Բադեն Պոուելի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 23] կողմից իր «Փորձ աշխարհների միասնության մասին» գրքում, որը լույս է տեսել 1855 թվականին։ Նա զարմանալի հստակությամբ ապացուցում է, որ նոր տեսակների երևան գալը պետք է դիտվի որպես «օրինաչափ, այլ ոչ թե պատահական երևույթ» կամ արտահայտվելով սըր Ջոն Հերշելի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 24] խոսքերով՝ որպես «բնական երևույթ, որը հակադրվում է հրաշագործականին»։
Լիննեյան ընկերության ամսագրի երրորդ հատորը բովանդակում է այն հոդվածները, որոնք 1858 թ. հուլիսի 1-ին ներկայացված են մ-ր Ուոլլեսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] և իմ կողմից և պարփակում են, ինչպես երևում է ներկա աշխատության ներածությունից, բնական ընտրության տեսությունը, որ զարգացրել է մ-ր Ուոլլեսը զարմանալի պարզությամբ և ուժով։
Ֆոն-Բերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 25] որը կենդանաբանների այնպիսի խոր հարգանքն է վայելում, մոտավորապես 1859 թվականին (տե՛ս պրոֆ. Ռուդոլֆ Վագների[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 26] «Zoologisch—Antropologische Untersuchungen»[30] 1861 թ., էջ 51), հիմնվելով աշխարհագրական տարածման օրենքների վրա, այն համոզմունքն է հայտնում, որ այժմ իրարից խիստ տարբերվող ձևերն առաջացել են մի ընդհանուր նախածնողից։
1859 թվականի հունիսին պրոֆ. Հյոքսլին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 27] թագավորական ինստիտուտում կարդացել է «Կենդանական կյանքի կայուն տիպերի մասին» դասախոսությունը։ Ուշադրություն դարձնելով նման դեպքերին, նա նկատում է. «Դժվար է բացատրել նման փաստերի նշանակությունը, եթե ենթադրենք, թե կենդանիների և բույսերի բոլոր տեսակները կամ կազմվածքի բոլոր հիմնական տիպերը ստեղծվել ու տեղադրվել են մեր մոլորակի մակերեսի վրա երկար ժամանակամիջոցներից հետո՝ շնորհիվ առանձին ստեղծագործական ակտերի։ Եվ չպետք է մոռանալ, որ նման ենթադրությունը նույնպես քիչ է հիմնավորվում ավանդությամբ կամ հայտնությամբ, ինչպես և հակասում բնության տրամադրած ընդհանուր անալոգիաներին։ Մյուս կողմից, եթե մենք դիտենք «կայուն տիպերը» այն հիպոթեզի տեսանկյունից, որի հիման վրա մի որոշ ժամանակում ապրող տեսակներն առաջանում են առաջներում ապրող տեսակներից աստիճանական փոփոխության միջոցով, մի հիպոթեզի, որը թեև դեռ ապացուցված չէ և զգալիորեն այլանդակված է նրան մի քանի կողմնակիցների կողմից, բայց առայժմ դեռ միակն է, որն ունի ֆիզիոլոգիական իմաստ,— ապա այդ տիպերի գոյությունը միայն կապացուրեր, որ երկրաբանական ժամանակաշրջանում կենդանի էակների կրած փոփոխությունները աննշան են ամբողջությամբ վերցրած այն փոփոխությունների համեմատությամբ, որ կրել են նրանք»։
1859 թ. դեկտեմբերին Հուկերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] լույս ընծայեց իր «Ավստրալիական բուսականության ներածություն» աշխատությունը։ Այդ կապիտալ աշխատության առաջին մասում նա ընդունում է փոփոխության միջոցով տեսակների ծագման ուսմունքը և այն հիմնավորում է բազմաթիվ ինքնուրույն դիտումներով։
Ներկա աշխատության առաջին հրատարակությունը լույս է տեսել 1859 թ. նոյեմբերի 24-ին, իսկ երկրորդը՝ 1860 թ. հունվարի 7-ին։
ՆԵՐԱԾՈՒԹՅՈՒՆ
Ճանապարհորդելով «Բիգլ»-ի վրա որպես բնախույզ, ինձ սաստիկ զարմացրել էին մի քանի փաստեր, որոնք վերաբերում էին Հարավային Ամերիկայում օրգանական էակների տեղաբաշխմանը և այդ մայր ցամաքի նախկին և ժամանակակից բնակիչների միջև եղած երկրաբանական հարաբերություններին։ Այդ փաստերը, ինչպես կտեսնենք այս գրքի վերջին գլուխներից, թվում էր, թե որոշ լույս են սփռում տեսակների ծագման, այդ գաղտնիքների գաղտնիքի վրա, ինչպես ասել է մեր մեծագույն գիտնականներից մեկը։ 1837 թ. տուն վերադառնալուց հետո, իմ մեջ միտք հղացավ, որ գուցե կարելի է որևէ բանի հասնել այդ հարցի լուծման իմաստով՝ համբերատար կերպով հավաքելով ու կշռադատելով բոլոր այն փաստերը, որոնք որևէ առնչություն ունեն նրա հետ։ Հինգ տարվա աշխատանքից հետո ես ինձ թույլ տվեցի որոշ ընդհանուր խորհրդածություններ անել այդ խնդրի մասին և դրանք ուրվագրեցի համառոտ նշումների ձևով. 1844 թվականին այդ ուրվագիրս ընդարձակվելով, դարձավ այն եզրակացությունների ընդհանուր ակնարկ, որոնք այն ժամանակ ինձ հնարավոր էին թվում. այդ ժամանակից սկսած մինչև օրս ես համառ կերպով զբաղվում եմ այդ հարցով։ Հույսով եմ, որ զուտ իմ անձնականին վերաբերող այդ մանրամասները կներվեն, որովհետև ես դրանք բերում եմ միայն նրա համար, որպեսզի ցույց տամ, որ իմ եզրակացություններում չեմ շտապել։
Իմ աշխատությունը այժմ (1859 թ.) գրեթե ավարտված է, բայց քանի որ նրա վերջնական մշակման համար դեռ մի քանի տարիներ են պահանջվում, իսկ իմ առողջությունը բոլորովին էլ ծաղկափթիթ վիճարկում չէ, ուստի ինձ համոզեցին հրատարակել այս «Քաղվածքը»։ Այդ անելու առանձնապես ինձ հարկադրում է այն հանգամանքը, որ մ-ր Ուոլլեսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] որն այժմ ուսումնասիրում է Մալայան արշիպելագի բնապատմությունը, տեսակների ծագման հարցի վերաբերմամբ եկել է եզրակացությունների, որոնք միանգամայն նման են իմ հանած եզրակացություններին։ 1858 թ. նա ուղարկել է այդ հարցին նվիրված մի հոդված, խնդրելով այն ուղարկել սըր Չարլզ Լյաելին,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] որը հոդվածը հղել է Լիննեյան ընկերությանը (այն տպագրված է այդ ընկերության հանդեսի երրորդ հատորում)։ Սըր Չարլզ Լյաելը և դ-ր Հուկերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] որոնք գիտեին իմ աշխատության մասին,— վերջինս կարդացել էր իմ ուրվագիծը 1844 թվականին,— պատիվ ցույց տալով ինձ, խորհուրդ տվեցին մ-ր Ուոլլեսի հոդվածի հետ միասին տպագրել նաև համառոտ քաղվածքներ իմ ձեռագրից։
Այժմ հրատարակվող «Քաղվածքը» անխուսափելիորեն անկատար է։ Ես չեմ կարող այստեղ բերել վկայակոչումներ կամ մատնանշել հեղինակություններ հաստատելու համար իմ այս կամ այն դրույթը. հույսով եմ, որ ընթերցողը կվստահի իմ բարեխղճությանը։ Անկասկած, իմ աշխատության մեջ սպրդել են սխալներ, թեև ես շարունակ մտահոգվել եմ այն մասին, որ վստահեմ միայն լավ հեղինակություններին։ Ես կարող եմ այստեղ շարադրել միայն այն ընդհանուր եզրակացությունները, որոնց եկել եմ ես, լուսաբանելով դրանք միայն մի քանի փաստերով. բայց հուսով եմ, որ մեծ մասամբ նրանք բավարար կլինեն։ Ոչ ոք ինձանից ավելի չի գիտակցում իմ եզրակացությունների հիմնավորման համար փաստերն ու վկայակոչումները ամբողջ մանրամասնությամբ հետագայում ներկայացնելու անհրաժեշտությունը, և ես հույս ունեմ, որ այդ կկատարեմ իմ ապագա աշխատության մեջ։ Ես շատ լավ գիտեմ, որ այդ գրքում գրեթե չկա ոչ մի դրույթ, որի վերաբերմամբ կարելի չլիներ ներկայացնել փաստեր, որոնք բերում են իմ հանած եզրակացություններին հավանաբար ուղղակի հակառակ եզրակացությունների։ Ճիշտ եզրակացություն կարելի է հանել միայն այս կամ այն կողմ հակվող փաստերի լրիվ շարադրումից և փաստարկների գնահատումից հետո, իսկ այդ այստեղ, իհարկե, չի կարելի անել։
Ես շատ ցավում եմ, որ տեղի սղությունը ինձ զրկում է բարոյական բավականությունից՝ իմ շնորհակալությունը հայտնելու այն մեծահոգի աջակցության համար, որ շատ բնախույզներ ցույց են տվել ինձ, որոնցից շատերը նույնիսկ անձնապես անծանոթ են։ Սակայն ես չեմ կարող առիթը բաց թողնել, չասելով, թե որքան շատ պարտական եմ դոկտոր Հուկերին,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] որը վերջին տասնհինգ տարիների ընթացքում բոլոր հնարավոր միջոցներով օգնել է ինձ իր ընդարձակ գիտելիքների և հստակ դատողության շնորհիվ։
Ինչ վերաբերում է տեսակների ծագման հարցին, ապա միանգամայն հնարավոր է, որ օրգանական էակների փոխադարձ ազգակցության, նրանց սաղմնաբանական հարաբերությունների, աշխարհագրական տարածման, երկրաբանական հաղորդականության և այլ նման փաստերի մասին խորհող բնախույզը կարող էր գալ այն եզրակացության, որ տեսակները մեկը մյուսից անկախ կերպով չեն ստեղծվել, այլ տարատեսակների նման առաջացել են այլ տեսակներից։ Այնուամենայնիվ, նման եզրակացությունը, թեկուզ լավ հիմնավորված էլ լիներ, անբավարար կլիներ, մինչև որ ցույց չտրվեր, թե ինչու աշխարհում ապրող անհամար տեսակները փոփոխվում էին հենց այնպիսի ձևով, որ ստացվել է կառուցվածքի և հարմարման այն կատարելությունը, որը իրավացիորեն զարմացնում է մեզ։ Բնախույզները շարունակ մատնանշում են արտաքին պայմանների՝ կլիմայի, սննդի և այլն ազդեցությունը, որպես փոփոխականության միակ պատճառ։ Որոշ, սահմանափակ իմաստով, ինչպես ցույց կտրվի հետագայում, դա գուցե և ճիշտ է, բայց պարզապես անհեթեթություն կլիներ միայն արտաքին պայմանների ազդեցությանը վերագրել, օրինակ, փայտփորի կազմվածքը նրա ոտներով, պոչով, կտուցով և լեզվով, որոնք հիանալի հարմարված են ծառերի կեղևի տակից միջատներ որսալու համար։ Նույնը նաև սպիտակ ճագոմի մասին, որը իր սնունդը վերցնում է որոշ ծառերի ցողուններից և ունի որոշ թռչունների միջոցով տարածվող սերմեր ու բաժանասեռ ծաղիկներ, որոնք որոշ միջատների օժանդակության կարիք են զգում ծաղկափոշին մի ծաղկից մյուսի վրա տեղափոխելու համար, նույնպես անհեթեթություն կլիներ այդ մակաբույծ բույսի կառուցվածքը և նրա կապերը օրգանական էակների զանազան խմբերի հետ բացատրել արտաքին պայմանների ազդեցությամբ, հենց այդ բույսի սովորությամբ կամ կամքի ակտով։
Հետևաբար, վերին աստիճանի կարևոր է պարզ պատկերացում ստանալ օրգանիզմների փոփոխության և հարմարման եղանակների մասին։ Իմ դիտումների սկզբում ինձ հավանական էր թվում, որ ընտանի կենդանիների և մշակովի բույսերի մանրազնին ուսումնասիրությունը ամենալավ միջոցը կտար, որպեսզի պարզաբանվեր այդ մթին հարցը։ Եվ ես չսխալվեցի, ինչպես այդ, այնպես էլ բոլոր այլ խճճված դեպքերում ես միշտ գտնում էի, որ ընտանի ցեղերի փոփոխության մասին մեր տեղեկությունները, չնայած իրենց անլրիվությանը, միշտ էլ եղել են լավագույն և ամենաստույգ բանալին։ Կարող եմ այդ առթիվ հայտնել իմ համոզմունքը նման ուսումնասիրության առանձին արժեքավորության մասին, չնայած այն արհամարհանքին, որին նա սովորաբար ենթարկվել է բնախույզների կողմից։
Այս նկատառումների հիման վրա այս «Քաղվածքի» առաջին գլուխը նվիրում եմ ընտանեցված վիճակում առաջացող փոփոխականությանը։ Այսպիսով, մենք կհամոզվենք, որ ժառանգաբար փոխանցվող փոփոխությունները հնարավոր են լայն չափերով, ինչպես նաև կիմանանք,— որը թերևս ավելի էական է,— թե ինչպես մեծ է մարդու զորությունը ընտրության միջոցով հաջորդական թույլ փոփոխությունները կուտակելու տեսակետից։ Այնուհետև ես կանցնեմ բնական վիճակում տեսակների փոփոխականության ուսումնասիրությանը, բայց ափսոս, որ ես ստիպված կլինեմ այդ հարցը շոշափել միայն ամենահամառոտ գծերով, որովհետև նրա հարկ եղածի պես շարադրումը կպահանջեր փաստերի երկար թվարկումներ։ Սակայն մենք ի վիճակի կլինենք քննարկելու, թե ո՞ր պայմաններն առանձնապես բարենպաստ են փոփոխականության համար։ Հետևյալ գլխում քննության կառնվի գոյության կռիվը, որը դրսևորվում է ամբողջ աշխարհում բոլոր օրգանական էակների միջև և անխուսափելիորեն բխում է նրանց բազմացման երկրաչափական պրոգրեսիայից։ Դա Մալթուսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 30] ուսմունքն է՝ տարածված երկու — կենդանական և բուսական թագավորությունների վրա[31]։ Քանի որ յուրաքանչյուր տեսակից ծնվում են շատ ավելի անհատներ, քան դրանցից կարող են ապրել, և քանի որ դրա հիման վրա շարունակ առաջանում է գոյության կռիվ, ապա դրանից հետևում է, որ ամեն մի էակ, որը թեկուզ և աննշան կերպով փոխվում է իրեն համար նպաստավոր ուղղությամբ իր գոյության հաճախ փոփոխվող և բարդ պայմանների նկատմամբ, ավելի շանսեր կունենա պահպանվելու և, այդպիսով, կենթարկվի բնական ընտրության։ Ժառանգականության հզոր սկզբունքի շնորհիվ յուրաքանչյուր տարատեսակ կձգտի բազմացնել իր նոր փոփոխված ձևը։
Այդ հիմնական հարցը՝ բնական ընտրության տեսությունը մանրամասնորեն կզարգացվի չորրորդ գլխում. մենք այն ժամանակ կտեսնենք, թե ինչպես բնական ընտրության գրեթե անխուսափելի հետևանքով կյանքի ավելի պակաս կատարյալ ձևերը մահաջնջվում են և առաջ է գալիս այն, ինչ ես անվանել եմ հատկանիշների տարամիտում։ Հետևյալ գլխում ես քննարկման կենթարկեմ փոփոխականության բարդ և քիչ հայտնի օրենքները։ Հաջորդ հինգ գլուխներում պարզաբանվելու են ամենից ավելի աչքի ընկնող և ամենաէական դժվարությունները, որոնց հանդիպում է տեսությունը, այն է, նախ՝ անցման, այսինքն՝ պարզ էակի կամ պարզ օրգանի բարձր կազմվածքով էակի կամ բարդ կառուցվածքով օրգանի ձևափոխման դժվարինությունը. երկրորդ՝ կենդանիների բնազդի կամ մտավոր ընդունակությունների հարցը, երրորդ՝ հիբրիդիզմը կամ տեսակների խաչաձևման անպտղաբերությունը և տարատեսակների խաչաձևման պտղաբերությունը, չորրորդ՝ երկրաբանական տարեգրության անկատարելությունը։ Դրանց հաջորդող գլխում ես կքննարկեմ օրգանական էակների երկրաբանական հաջորդականությունը ժամանակի ընթացքում. տասներկուերորդ և տասներեքերորդ գլուխներում՝ նրանց աշխարհագրական տեղաբաշխումը տարածության մեջ. տասնչորսերորդում՝ նրանց դասակարգությունը և փոխադարձ ազգակցությունը հասունացած և սաղմնային վիճակում։ Վերջին գլխում ես կտամ ամբողջ աշխատության մեջ շարադրվածի համառոտ կրկնությունը և մի քանի եզրափակիչ դիտողություններ։
Ոչ ոք չպիտի զարմանա այն բանի համար, որ տեսակների ծագման վերաբերող շատ բան դեռ մնում է չբացատրված, եթե միայն հասկանա, թե մենք ինչ խոր անտեղյակության մեջ ենք գտնվում մեզ շրջապատող անհամար կենդանի էակների փոխադարձ կապերի մասին։ Ո՞վ կբացատրի, թե ինչու մի տեսակը լայնորեն տարածված է և ունի բազմաթիվ անհատներ, իսկ մյուսը քիչ է տարածված և հազվագյուտ։ Մինչդեռ այդ հարաբերությունները խիստ կարևոր են, որովհետև նրանք որոշում են այս աշխարհի յուրաքանչյուր բնակչի արդի բարեկեցությունը և, ինչպես ես կարծում եմ, ապագա հաջողությունն ու հետագա փոփոխությունը։ Է՛լ ավելի քիչ գիտենք մեր մոլորակի անհամար բնակիչների փոխհարաբերությունների մասին նրա պատմության անցյալ երկրաբանական դարաշրջանների ընթացքում։ Թեպետև շատ բան դեռ մութ է և երկար ժամանակ մութ կմնա, սակայն, որքան որ ես ընդունակ եմ անելու, ամենամանրազնին հետազոտությունների և անկողմնակալ քննարկման հետևանքով ես երբեք չեմ կասկածում, որ այն հայացքը, որին մինչև վերջին ժամանակներս համամիտ էր բնախույզների մեծ մասը, ինչպես նաև ես, այսինքն՝ որ յուրաքանչյուր տեսակ ստեղծվել է մյուսներից անկախ կերպով, ճիշտ չէ։ Ես միանգամայն համոզված եմ, որ տեսակները փոփոխական են և որ միևնույն սեռին պատկանող բոլոր տեսակները հանդիսանում են մի որևէ, մեծ մասամբ բնաջնջված տեսակի անմիջական սերունդները, ճիշտ այնպես, ինչպես որևէ տեսակի տարատեսակներ ճանաչվածները այդ տեսակի սերունդներն են։ Այնուհետև, ես համոզված եմ, որ բնական ընտրությունը այդ փոփոխության եթե ոչ միակ, ապա ամենակարևոր գործոնն է եղել։
ԳԼՈՒԽ I։ ՓՈՓՈԽԱԿԱՆՈՒԹՅՈՒՆՆ ԸՆՏԱՆԵՑՄԱՆ ՎԻՃԱԿՈՒՄ
ՓՈՓՈԽԱԿԱՆՈՒԹՅԱՆ ՊԱՏՃԱՌՆԵՐԸ։— ՍՈՎՈՐՈՒԹՅԱՆ ԵՎ ՕՐԳԱՆՆԵՐԻ ՎԱՐԺԵՑՄԱՆ ԿԱՄ ՈՉ ՎԱՐԺԵՑՄԱՆ ԱԶԴԵՑՈՒԹՅՈՒՆԸ։ ՓՈՓՈԽԱԿԱՆՈՒԹՅԱՆ ՀԱՐԱԲԵՐԱԿՑՈՒԹՅՈՒՆՆԵՐԸ. ԺԱՌԱՆԳԱԿԱՆՈՒԹՅՈՒՆ։— ԸՆՏԱՆԵՑՐԱԾ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԸՆԴՀԱՆՈՒՐ ԲՆՈՒՅԹԸ. ՏԵՍԱԿՆԵՐԸ ԵՎ ՏԱՐԱՏԵՍԱԿՆԵՐԸ ՏԱՐԲԵՐԵԼՈՒ ԴԺՎԱՐՈՒԹՅՈՒՆՆԵՐԸ. ԸՆՏԱՆԵՑՐԱԾ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԾԱԳՈՒՄԸ ՄԵԿ ԿԱՄ ՄԻ ՔԱՆԻ ՏԵՍԱԿՆԵՐԻՑ։— ԸՆՏԱՆԻ ԱՂԱՎՆԻՆԵՐԸ, ՆՐԱՆՑ ԾԱԳՈՒՄԸ ԵՎ ՏԱՐԲԵՐՈՒԹՅՈՒՆՆԵՐԸ։— ԸՆՏՐՈՒԹՅԱՆ ՍԿԶԲՈՒՆՔԸ ԿԻՐԱՌՎԵԼ Է ԴԵՌ ՀՆՈՒՄ. ՆՐԱ ԿԻՐԱՌՄԱՆ ԱՐԴՅՈՒՆՔՆԵՐԸ։ ՄԵԹՈԴԱԿԱՆ ԵՎ ԱՆԳԻՏԱԿՑԱԿԱՆ ԸՆՏՐՈՒԹՅՈՒՆ։— ՄԵՐ ԸՆՏԱՆԻ ՑԵՂԵՐԻ ԱՆՀԱՅՏ ԾԱԳՈՒՄԸ։— ԸՆՏՐՈՒԹՅԱՆ ԿԻՐԱՌՄԱՆ ԳՈՐԾՈՒՄ ՄԱՐԴՈՒՆ ՆՊԱՍՏՈՂ ՀԱՆԳԱՄԱՆՔՆԵՐԸ։
ՓՈՓՈԽԱԿԱՆՈՒԹՅԱՆ ՊԱՏՃԱՌՆԵՐԸ
Երբ մենք համեմատում ենք մեր հնագույն ընտանի կենդանիներից կամ կուլտուրական բույսերից որևէ մեկի միևնույն տարատեսակին կամ ենթատարատեսակին պատկանող անհատները իրար հետ, մեզ ամենից առաջ զարմացնում է այն հանգամանքը, որ նրանք միմյանցից ավելի շատ են տարբերվում, քան միևնույն տեսակի կամ տարատեսակի անհատները բնական վիճակում։ Եվ երբ մենք խորհում ենք բույսերի և կենդանիների այն հսկայական բազմազանության մասին, որ արհեստականորեն ստեղծվել ու փոփոխվել է դարերի ընթացքում կլիմայի և իրադրության ամենաբազմազան պայմաններում, ապա գալիս ենք այն եզրակացության, որ ընտանեցման այդ արգասիքները առաջացել են այնպիսի կենսապայմաններում, որոնք եղել են անհամեմատ ավելի քիչ միակերպ և որոշ չափով տարբեր, քան այն կենսապայմանները, որոնց մեջ բնական վիճակում գոյություն են ունեցել նրանց ծնող տեսակները։ Նմանապես բավական հավանական է նաև Անդրու Նայտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 31] արտահայտած կարծիքը, որ այդ փոփոխականությունը որոշ չափով կապ ունի սննդի առատության հետ։ Ակներև է միայն այն, որ օրգանական էակները պետք է մի քանի սերունդների ընթացքում ենթարկվեին նոր պայմանների ազդեցությանը, որպեսզի նրանց մեջ զգալի փոփոխականություն դրսևորվեր. ակներև է նաև այն, որ փոփոխականության ձգտումը դրսևորող օրգանիզմները շարունակում են փոփոխվել բազմաթիվ սերունդների ընթացքում։ Հայտնի չէ ոչ մի դեպք, որ փոփոխական օրգանիզմը ընտանեցման վիճակում դադարեր փոփոխվելուց։ Մեր հնագույն կուլտուրական բույսերը, ինչպես, օրինակ, ցորենը, շարունակում են տալ նոր տարատեսակներ, մեր հնագույն ընտանի կենդանիները դեռ ևս պահպանում են արագ փոփոխվելու և կատարելագործվելու ընդունակությունը։
Այդ հարցը երկարատև ուսումնասիրելուց հետո, որքան ես կարող եմ եզրակացնել, կասեմ, որ կենսապայմանները, ըստ երևույթին, ներգործում են երկու ձևով՝ անմիջականորեն ամբողջ կազմվածքի կամ նրա միայն մի որոշ մասի վրա և անուղղակի կերպով ազդելով վերարտադրողական սիստեմի վրա։ Անմիջական ներգործության վերաբերմամբ մենք պետք է միշտ նկատի ունենանք այն, որ ամեն մի նման դեպքում, ինչպես վերջին ժամանակներս այդ պնդում է պրոֆ. Վեյսմանը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 32] և ինչպես ես ցույց եմ տվել իմ «Փոփոխությունն ընտանեցված վիճակում» աշխատության մեջ, պետք է տարբերել երկու գործոն՝ օրգանիզմի բնույթը և պայմանների բնույթը[32]։ Առաջինը, ըստ երևույթին, ամենից ավելի էական է, որովհետև բոլորովին նման փոփոխություններ են առաջանում, որքան մենք կարող ենք դատել, բոլորովին տարբեր պայմաններում, իսկ մյուս կողմից, ոչ նման փոփոխությունները առաջանում են, ըստ երևույթին, բոլորովին միանման պայմաններում։ Ազդեցությունները սերնդի վրա կարող են լինել կամ որոշակի կամ անորոշ։ Նրանք կարող են որոշակի համարվել, երբ որոշ պայմանների ենթարկված անհատների ամբողջ կամ համարյա ամբողջ սերունդը փոփոխվում է միանման կերպով։ Շատ դժվար է որոշ եզրակացության գալ այդպիսի որոշակի կերպով առաջացող փոփոխության չափերի մասին։ Բայց կասկածի ենթակա չէ, որ մի շարք ոչ խոր փոփոխություններ առաջանում են այդպիսի եղանակով՝ օրինակ, հասակը կախված է սննդի քանակությունից, գույնը՝ սննդի որակից, մաշկի հաստությունը և մազոտությունը՝ կլիմայից և այլն։ Մեր ընտանի թռչունների անսահման բազմազան գույներից յուրաքանչյուրը պետք է ունենա իր ֆիզիկական պատճառը. և եթե նույն պատճառները միանման ձևով ներգործեին սերունդների երկար շարքի ընթացքում մեծ թվով անհատների վրա, ապա նրանք բոլորը հավանորեն կփոփոխվեին միանման ձևով։ Այնպիսի փաստերը, ինչպես, օրինակ, բարդ և արտասովոր մակաճումների գոյացումը, որը միշտ տեղի է ունենում գխտոր առաջացնող միջատների արտադրած թույնի կաթիլի ներթափանցումից հետո, մեզ ցույց են տալիս, թե ինչպիսի տարօրինակ փոփոխություններ կարող են առաջանալ բույսերի մեջ նրանց հյութերի բաղադրության քիմիական փոփոխության ազդեցության ներքո։
Փոփոխված պայմանների անհամեմատ ավելի տարածված հետևանք է հանդիսանում անորոշ փոփոխականությունը, քան որոշակի փոփոխականությունը և հավանորեն ավելի աչքի ընկնող դեր է կատարել մեր ընտանի ցեղերի առաջացման գործում։ Անորոշ փոփոխականությունը մենք տեսնում ենք այն անսահմանորեն բազմազան թույլ տարբերություններում, որոնցով տարբերվում են միևնույն տեսակին պատկանող անհատները և որոնք չէին կարող ժառանգվել ոչ ծնողներից մեկնումեկից, ոչ էլ ավելի հեռավոր նախնիներից։ Հաճախ նույնիսկ շատ խիստ արտահայտված տարբերություններ են երևում միաժամանակ մի մորից ծնված կենդանիների միջև, կամ միևնույն սերմնատուփից ստացված սերմերից աճեցված բույսերի միջև։ Երկար ժամանակամիջոցների ընթացքում միևնույն երկրում, գրեթե միևնույն սննդով աճեցրած միլիոնավոր անհատների միջև երևան են գալիս կազմվածքի այնքան խիստ արտահայտված շեղումներ, որ ստանում են այլանդակություններ անունը, բայց հնարավորություն չկա որևէ որոշակի սահմանագիծ անցկացնելու այլանդակության և ավելի թույլ փոփոխությունների միջև։ Թույլ կամ խիստ արտահայտված նման բոլոր փոփոխությունները, որոնք երևան են գալիս միասին ապրող անհատների մեջ, կարող են դիտվել որպես գոյության պայմանների անորոշ ներգործություններ ամեն մի անհատ օրգանիզմի վրա այնպես, ինչպես ցրտառությունը անորոշ կերպով է ազդում տարբեր մարդկանց վրա՝ նրանց կազմությանը կամ վիճակին համապատասխանորեն, առաջ բերելով երբեմն հազ և հարբուխ, երբեմն ռևմատիզմ կամ զանազան օրգանների բորբոքում[33]։
Ինչ վերաբերում է այն բանին, որ ես անվանել եմ փոփոխված պայմանների անուղղակի ներգործություն՝ վերարտադրողական սիստեմի վրա նրանց ազդեցության միջոցով, ապա այդպիսի ներգործության գոյության մասին կարելի է եզրակացնել մասամբ այդ սիստեմի առանձնահատուկ դյուրազգացությամբ դեպի պայմանների ամեն մի փոփոխություն, մասամբ էլ Կյոլռեյտերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] և ուրիշների նկատած այն նմանության հիման վրա, որ կա տեսակների խաչաձևումից առաջացող փոփոխականության և այն փոփոխականության միջև, որը նկատվում է այն դեպքում, երբ բույսերը կամ կենդանիները աճեցվում են նոր կամ անբնական պայմաններում։ Բազմաթիվ փաստեր պարզ կերպով ցույց են տալիս, որ վերարտադրողական սիստեմը առանձին զգայունություն է ցուցաբերում շրջապատող պայմաններում կատարվող նույնիսկ ամենաթույլ փոփոխությունների նկատմամբ։ Կենդանուն ընտելացնելուց ավելի հասարակ բան չկա և, ընդհակառակը, չափազանց դժվար է այնպես անել, որ նա ազատ կերպով բազմանա գերության մեջ, նույնիսկ այն ժամանակ, երբ էգերն ու արուները գտնվում են միասին։ Ինչքա՜ն մեծ է այն կենդանիների թիվը, որոնք չեն բազմանում նույնիսկ այն ժամանակ, երբ նրանց պահում են համարյա լիակատար ազատության մեջ և նրանց հայրենիքում։ Դա սովորաբար, և միանգամայն սխալ կերպով, վերագրում են այլասեռված բնազդներին։ Կուլտուրական շատ բույսեր փարթամ կերպով զարգանում են, բայց քիչ սերմ են տալիս կամ նույնիսկ բոլորովին չեն տալիս։ Մի քանի հազվագյուտ դեպքերում նկատված է, որ այնպիսի ոչ էական հանգամանքից, ինչպիսին է բույսի զարգացման որոշ շրջանում նրա ստացած ջրի մեծ կամ փոքր քանակությունը, կախված է բույսի սերմ տալը կամ չտալը։ Ես չեմ կարող բերել այստեղ մանրամասն տվյալներ, որ ես հավաքել ու հրատարակել եմ մի այլ տեղ. բայց ցույց տալու համար, թե ինչքան տարօրինակ են գերության մեջ կենդանիների բազմացումը ղեկավարող օրենքները, ես միայն կհիշատակեմ, որ գիշատիչ կաթնասունները, նույնիսկ արևադարձայինները, համեմատաբար հեշտությամբ են բազմանում մեզ մոտ գերության մեջ, բացառությամբ թաթագնացների կամ արջերի ընտանիքի, որոնք գրեթե երբեք ձագ չեն բերում, մինչդեռ գիշատիչ թռչունները, շատ հազվադեպ բացառությամբ, դժվար թե երբևիցե ճտահանության համար պիտանի ձվեր են ածում։ Էկզոտիկ շատ բույսեր արտադրում են բեղմնավորման համար անպետք ծաղկափոշին ճիշտ այնպես, ինչպես անպտուղ հիբրիդների մեծ մասը։ Մի կողմից մենք հանդիպում ենք թեկուզև թույլ ու վտիտ, բայց ազատ կերպով գերության մեջ բազմացող մշակովի բույսերի և կենդանիների. մյուս կողմից, մենք հանդիպում ենք անհատների, որոնք մատաղ հասակում վերցված են իրենց բնական շրջադրությունից, լիովին ընտանեցված են, առողջ և երկարակյաց են (որի համար ես կարող եմ շատ օրինակներ բերել), բայց ունեն վերարտադրողական սիստեմ, որը մեզ համար անըմբռնելի պատճառով ամուլ է։ Մենք չպետք է զարմանանք, որ գերության մեջ զարգացող վերարտադրողական սիստեմը գործում է միանգամայն անկանոն և արտադրում է ծնողներին չնմանվող սերունդ։ Ես կարող եմ ավելացնել, որ մինչդեռ որոշ օրգանիզմներ ազատ կերպով բազմանում են ամենաանբնական պայմաններում (օրինակ, վանդակաբներում պահվող ճագարները և ժանտաքիսները), դրանով ապացուցելով, որ նրանց վերարտադրողական օրգանները հեշտությամբ չեն ախտահարվում, այլ կենդանիներ ու բույսեր ընտելացման չեն ենթարկվում և շատ քիչ են փոփոխվում, գրեթե այնքան քիչ, ինչքան բնական վիճակում։
Որոշ բնախույզներ պնդում էին, որ ամեն մի փոփոխություն կապված է սեռական վերարտադրության ակտի հետ, բայց դա ակներևորեն սխալ է, որովհետև իմ մի այլ աշխատության մեջ ես բերել եմ, այսպես կոչված «բնության խաղեր» (sporting plants) երևույթների մի երկար ցանկ, այսինքն այնպիսի դեպքեր, երբ բույսերը հանկարծակի առաջ են բերում բոլորովին նոր առանձնահատկություններով մի բողբոջ, որը տարբերվում է նույն բույսի մնացած բոլոր բողբոջներից։ Այդ բողբոջային վարիացիաները, ինչպես նրանց կարելի է անվանել, կարող են բազմացվել պատվաստելով, ընձյուղներով և այլն, իսկ երբեմն էլ նույնիսկ սերմերով։ Բնության մեջ այդ դեպքերը քիչ են, բայց մշակույթի մեջ նրանց չի կարելի դասել հազվադեպ երևույթների շարքում։ Միանման պայմաններում տարեցտարի միևնույն ծառի առաջացրած բազմահազար բողբոջներից մեկը հանկարծակի ստանում է բոլորովին նոր բնույթ, մյուս կողմից պատահում է, որ տարբեր պայմաններում աճած ծառերի վրա երևում են բողբոջներ, որոնք սկիզբ ես տալիս ճիևնույն տարատեսակին, ինչպես այդ եղել է դեղձի ծառերի վրա նեկտարիններ[34] երևալու օրինակում և սովորական վարդենիների վրա բազմաթերթիկ վարդեր երևալու օրինակում։ Ելնելով այս փաստերից, մենք իրավունք ունենք եզրակացնելու, որ պայմանների բնույթը տվյալ յուրաքանչյուր փոփոխության առաջացման մեջ ավելի քիչ նշանակություն ունի, քան օրգանիզմի իր բնույթը, գուցե առաջինը ավելի էական ազդեցություն չի ունենում, քան այն կայծի բնույթը, որը բոցավառում է վառելանյութի զանգվածը, ազդում է բռնկվող բոցի հատկությունների վրա[35]։
ՍՈՎՈՐՈՒԹՅԱՆ ԵՎ ԳՈՐԾԱՐԱՆՆԵՐԻ ՎԱՐԺԵՑՄԱՆ ԿԱՄ ՈՉ ՎԱՐԺԵՑՄԱՆ ԱԶԴԵՑՈՒԹՅՈՒՆԸ. ՀԱՐԱԲԵՐԱԿՑԱԿԱՆ (ԿՈՌԵԼՅԱՏԻՎ) ՓՈՓՈԽԱԿԱՆՈՒԹՅՈՒՆ. ԺԱՌԱՆԳԱԿԱՆՈՒԹՅՈՒՆ
Փոփոխված սովորությունները թողնում են ազդեցություն, որը ժառանգաբար փոխանցնում է, ինչպես, օրինակ, մի կլիմայից մյուսը փոխադրած բույսերի ծաղկման շրջանի փոփոխությունը։ Կենդանիների օրգանների ուժեղ վարժեցումը կամ ոչ վարժեցումը դրսևորվում է ավելի խիստ հետևանքներով, այսպես, ես նկատել եմ, որ ընտանի բադի թևերի ոսկորները ավելի քիչ, իսկ ոտքերի ոսկորները ավելի շատ են կշռում ամբողջ կմախքի համեմատությամբ, քան վայրի բադինը, և այդ բանը վստահորեն կարելի է վերագրել այն հանգամանքին, որ ընտանի բադը ավելի քիչ է թռչում և ավելի շատ է քայլում, քան նրա վայրի նախնիները։ Կովերի և այծերի կրծի զգալի և ժառանգական զարգացումը այն երկրներում, որտեղ այդ կենդանիներին կթում են, համեմատած այլ երկրների այդ կենդանիների նույն օրգանների հետ, ամենայն հավանականությամբ օրգանի վարժեցման հետևանքների մի ուրիշ օրինակ է։ Համարյա չի կարելի ցույց տալ ընտանի անասունների որևէ ցեղ, որ որևէ երկրում չունենար կախ ընկած ականջներ, և հավանական է թվում այդ փաստի բացատրությունը ականջի մկանների վարժության բացակայությամբ, որը հետևանք է այն բանի, որ այդ կենդանիները վախի քիչ են ենթարկվում։
Փոփոխականությունը ղեկավարվում է բազմաթիվ օրենքներով, նրանցից մի քանիսն արդեն աղոտ կերպով պարզաբանվում են և համառոտակի քննարկվելու են հետագայում։ Ես այստեղ կանգ կառնեմ միայն այն բանի վրա, որի կարելի է անվանել փոփոխականության հարաբերակցություն։ Սաղմի կամ թրթուրի մեջ կատարվող էական փոփոխությունները հավանորեն առաջ են բերում փոփոխություններ նաև հասուն կենդանու մեջ։ Բոլորովին տարբեր մասերի միջև հարաբերակցությունները այլանդակություններում շատ հետաքրքրական են, այդ երևույթի բազմաթիվ դեպքեր հանդիպում են Իսիդոր Ժոֆֆրուա Սենտ-Իլերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 16] ընդարձակ աշխատության մեջ, որը նվիրված է այդ հարցին։ Անասնաբույծները համոզված են, որ երկար վերջավորությունները միշտ լինում են երկար գլխի հետ։ Հարաբերակցության որոշ դեպքեր զարմանալիորեն տարօրինակ են, այսպես, օրինակ, կապույտ աչքեր ունեցող սպիտակ կատուները սովորաբար խուլ են լինում. իսկ, ըստ միստր Տետի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 34] դիտողության, այդ առանձնահատկությունը ունենում են միայն արու կատուները։ Գույնը և օրգանական առանձնահատկությունները հաճախ կապված են լինում միմյանց հետ, որի մասին կարելի է շատ հիանալի օրինակներ բերել կենդանիների և բույսերի աշխարհից։ Հոյզինգերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 35] հավաքած փաստերից, ըստ երևույթին, հետևում է, որ որոշ բույսեր վնասակար ներգործություն են ունենում սպիտակ ոչխարների և խոզերի վրա, մինչդեռ սև անհատները վնաս չեն կրում։ Վերջերս պրոֆեսոր Վիմանը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 36] ինձ հաղորդել է նման երևույթի մի հիանալի օրինակի մասին։ Նա հարցրել է վիրգինիական մի քանի ֆերմերների թե ինչո՞ւ նրանց բոլոր խոզերը սև են, և պատասխան է ստացել, որ խոզերն ուտում են մի բույսի (Lachanthes-ի) արմատները, որոնք ներկում են նրանց ոսկորները վարդագույնով, որի հետևանքով նրանց, բացի սև տարատեսակներից, կճղակներն ընկնում են, իսկ վիրգինիական գաղութաբնակներից (տեղական արտահայտությամբ՝ crackers) մեկը ավելացրել է. «Մենք յուրաքանչյուր ծնում ընտրում ենք սև խոճկորներին, որովհետև միայն նրանք են կարողանում ապրել»։ Անմազ շներն ունենում են անկատար ատամներ, երկար և կոպիտ մազ ունեցող կենդանիները, ինչպես պնդում են ոմանք, աչքի են ընկնում երկար կամ ճյուղավորված եղջյուրներով, փետրապատ ոտներով աղավնիները մատների միջև ունենում են թաղանթներ, կարճակտուց աղավնիներն ունենում են կարճ, իսկ երկարակտուց աղավնիները՝ երկար ոտքեր։ Այսպիսով, մարդը, ընտրելով և կուտակելով կառուցվածքի մի որևէ առանձնահատկություն, հարաբերակցության (կոռելյացիայի) այդ խորհրդավոր օրենքի հիման վրա գրեթե միշտ առանց դիտավորության կփոփոխի նաև օրգանիզմի այլ մասերը[36]։
Փոփոխականության զանազան, բոլորովին անհայտ կամ աղոտ կերպով հասկացված օրենքների հետևանքները անսահմանորեն բարդ են ու բազմազան։ Արժե մանրազնին կերպով ուսումնասիրել մանրամասնորեն գրված գիտական շարադրությունները մեր հին մշակովի բույսերի, օրինակ, հիացինտի, կարտոֆիլի, նույնիսկ գեորգինի և այլոց մասին։ Պարզապես զարմանում ես կառուցվածքում և հատկություններում եղած անսահման բազմազանության վրա, որոնցով տարատեսակները և ենթատարատեսակները աննշան չափով տարբերվում են մեկը մյուսից։ Նրանց ամբողջ կազմվածքը կարծես դարձել է պլաստիկ և թույլ չափով շեղվում է ծնողների տիպից։
Ոչ ժառանգական փոփոխությունը մեզ համար էական չէ։ Ֆիզիոլոգիական տեսակետից ինչպես աննշան, այնպես էլ շատ կարևոր ժառանգական շեղումների թիվն ու բազմազանությունը կառուցվածքում անսահման է։ Պրոսպեր Լյուկի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 37] երկու ստվար հատորից բաղկացած երկասիրությունը ամենալավն է ու ամենալրիվը այդ հարցի մասին։ Յուրաքանչյուր սելեկցիոներ գիտե, թե ինչքան ուժեղ է հատկանիշների ժառանգական փոխանցման ձգտումը, որ «նմանն առաջ է բերում նմանին», կազմում է նրա հիմնական համոզմունքը, այդ տեսակետից կասկած են հայտնում միայն տեսաբանները։ Եթե որևէ շեղում կառուցվածքում հաճախակի է երևում և մենք այն տեսնում ենք ծնողների և զավակների մեջ, ապա մենք չենք կարող որոշել, թե այդ շեղումը արդյո՞ք տեղի է ունեցել մեկ ընդհանուր պատճառից, որը ազդել է թե՛ ծնողների և թե՛ զավակների վրա։ Բայց եթե միանման պայմանների, ըստ երևույթին, ենթակա անհատների մեջ բացառիկ հանգամանքների բերմունքով առաջացած հազվագյուտ մի շեղում հայտնվում է ծնողի մեջ, ասենք՝ մի քանի միլիոն անհատներից մեկի մեջ և կրկնվում է զավակների մեջ, ապա արդեն միայն հավանականության տեսությունը համարյա ստիպում է մեզ այդ կրկնությունը վերագրել ժառանգականության։ Յուրաքանչյուր ոք անշուշտ լսել է ալբինիզմի, փշոտ մաշկի, մազոտության դեպքերի մասին, որոնք երևան են գալիս միևնույն ընտանիքի մի քանի ներկայացուցիչների մեջ։ Եթե կառուցվածքում եղած տարօրինակ և հազվագյուտ շեղումները ժառանգվում են, ապա, իհարկե, պետք է ընդունենք, որ ավելի պակաս տարօրինակ և ավելի սովորական առանձնահատկությունները ևս ժառանգական են։ Գուցե այդ հարցի վերաբերմամբ ամենաճիշտ տեսակետը կլիներ այն, որ յուրաքանչյուր որևէ հատկանիշի ժառանգումը համարենք կանոն, իսկ նրա չժառանգելը՝ բացառություն։
Ժառանգականությունը ղեկավարող օրենքները մեծ մասամբ անհայտ են։ Ոչ ոք չի կարող ասել, թե ինչու միևնույն առանձնահատկությունը նույն տեսակի տարբեր անհատների մեջ կամ տարբեր տեսակների ներկայացուցիչների մեջ երբեմն ժառանգվում է, երբեմն էլ ոչ. թե ինչու զավակների մեջ վերարտադրվում են պապի, տատի, կամ ավելի ևս հեռավոր նախնիների հատկանիշները, թե ինչու որևէ առանձնահատկություն մի սեռից փոխանցվում է երկու սեռերին կամ միայն մեկին և ավելի հաճախ բացառապես նույն սեռին։ Մեզ համար բավական կարևոր է այն փաստը, որ մեր ընտանի ցեղերի արուներում արտահայտվող առանձնահատկությունները փոխանցվում են բացառապես կամ գերազանցապես միայն արուներին։ Ավելի ևս կարևոր է այն կանոնը, որին, ինձ թվում է, կարելի է լիովին վստահել, որ կյանքի որևէ շրջանում երևան եկած մի որևէ առանձնահատկություն ձգտում է երևան գալ սերնդի մեջ համապատասխան հասակում, թեև երբեմն էլ մի քիչ ավելի վաղ։ Շատ դեպքերում այլ կերպ էլ չէր կարող լինել, այսպես, օրինակ, մեր տավարի եղջյուրների կառուցվածքում ժառանգված առանձնահատկությունները կարող են երևան գալ միայն համարյա հասունացած կենդանիների մոտ. շերամի որդի առանձնահատկությունները երևան են գալիս թրթուրի կամ բոժոժի հասակում։ Բայց ժառանգական հիվանդությունները և մի քանի այլ փաստեր ինձ հարկադրում են ընդունել, որ այդ կանոնը ավելի լայն կիրառություն ունի, և որ նույնիսկ այն ժամանակ, երբ որոշ հատկանիշի դրսևորման համար որոշակի հասակում ոչ մի հիմք չկա, այնուամենայնիվ այդ ձգտումը դրսևորվում է սերնդի մեջ կյանքի այն շրջանում, ինչ շրջանում որ այն սկզբնապես երևացել է նախնիքի մեջ։ Ես կարծում եմ, որ այդ կանոնը վերին աստիճանի կարևոր է որպես սաղմնաբանության հիմնական օրենքների բացատրություն։ Այս դիտողությունները անշուշտ վերաբերում են առանձնահատկությունների միայն առաջին դրսևորմանը, այլ ոչ թե սկզբնական պատճառին, որը կարող է ազդել կամ ձվի, կամ արական տարրի վրա, այնպես, ինչպես կարճեղջյուր կովից և երկարաղջյուր ցուլից առաջացած հորթի եղջյուրների ավելացած երկարությունը թեև երևան է գալիս ուշ հասակում, բայց ակներև է, որ կախված է արական տարրի ազդեցությունից։
Հիշատակելով այստեղ նախկին հատկանիշներին վերադառնալու մասին, ես գտնում եմ, որ օգտակար է կանգ առնել բնախույզների կողմից հաճախ արվող հայտարարության վրա, այն է, որ մեր ընտանի տարատեսակները վայրենանալիս աստիճանաբար և անպայման վերադառնում են բնական վիճակում գտնվող իրենց նախնիների ունեցած հատկանիշներին։ Դրանից արվել է այն հետևությունը, որ ընտանի տարատեսակների վրա դիտված փաստերի հիման վրա հանված որևէ եզրակացությունն անհնարին է տարածել այն տեսակների վրա, որոնք գտնվում են բնական վիճակում։ Սակայն հակառակ իմ բոլոր ջանքերին, ես չկարողացա գտնել, թե ինչ համոզարար փաստերի վրա է հիմնված այդպես հաճախ և այդպես համարձակորեն հայտարարվող այդ դրույթը։ Մինչև անգամ շատ դժվար կլիներ այդ ապացույցները բերել. մենք կարող ենք վստահ լինել, որ խիստ նկատելի ընտանի տարատեսակների զգալի մեծամասնությունը չէր էլ կարողանա նույնիսկ գոյություն ունենալ վայրի վիճակում։ Շատ դեպքերում մենք չգիտենք այդ բնական նախնիքներին, ուստի և չենք կարող դատել, թե արդյոք լրիվ վերադարձ է կատարվել դեպի նախնիքների հատկանիշները կամ ոչ։ Խաչաձևման ազդեցությունից խուսափելու համար անհրաժեշտ կլիներ վայրենանալու թողնել միայն մեկ տարատեսակ։ Այնուամենայնիվ, քանի որ իրոք մեր տարատեսակները երբեմն իրենց որոշ հատկանիշներում դրսևորում են վերադարձ դեպի նախնիքների ձևերը, ինձ անհավանական էլ չի թվում, որ եթե մեզ հաջողվեր երկար սերունդների ընթացքում նատուրալիզացիայի ենթարկել կամ մշակել զանազան ցեղեր, օրինակ, կաղամբի ձևերը շատ աղքատ հողում (ըստ որում, իհարկե, ստացված հետևանքի մի մասը պիտի վերագրվեր աղքատ հողի որոշակի ներգործությանը), ապա նրանք զգալի չափով, գուցե և լիովին վերադառնային իրենց նախնական բնական ձևերին։ Փորձը կհաջողվեր, թե ոչ, մեր պատճառաբանության համար էապես կարևոր չէ, որովհետև փորձն ինքը կհանգեր կենսապայմանների փոփոխության։ Եթե կարելի լիներ ցույց տալ, որ մեր ընտանի տարատեսակները ուժեղ ձգտում են դրսևորում վերադառնալու դեպի նախնիքների հատկանիշները, այսինքն՝ կորցնելու իրենց ձեռք բերած հատկանիշները, երբ պահվում են միևնույն պայմաններում և մեծ թվով այնպես, որպեսզի հատկանիշների խաչաձևման և միաձուլման միջոցով վերացվեր թույլ շեղումների ամրացման հնարավորությունը,— այդ դեպքում ես կհամաձայնվեի, որ ընտանի տարատեսակների վրա կատարված դիտումների եզրակացությունները չի կարելի տարածել բնական տեսակների վրա։ Բայց նման հայացքի օգտին ապացույցի նշույլ էլ չկա. հաստատել, թե չէինք կարողանա պահպանել անորոշ թվով սերունդներում մեր լծելու և արշավելու ձիերը, երկարաեղջյուր և կարճեղջյուր անասուններին, ընտանի թռչուններին և բանջարեղենները, կնշանակեր հակասել մեր բոլոր փորձերին։
ԸՆՏԱՆԵՑՎԱԾ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԸՆԴՀԱՆՈՒՐ ԲՆՈՒՅԹԸ, ՏԵՍԱԿՆԵՐՆ ՈՒ ՏԱՐԱՏԵՍԱԿՆԵՐԸ ՏԱՐԲԵՐԵԼՈՒ ԴԺՎԱՐՈՒԹՅՈՒՆՆԵՐԸ, ԸՆՏԱՆԵՑՎԱԾ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԾԱԳՈՒՄԸ ՄԵԿ ԿԱՄ ՄԻ ՔԱՆԻ ՏԵՍԱԿՆԵՐԻՑ
Մեր ընտանեցված կենդանիների և մշակովի բույսերի ցեղերը կամ ժառանգական տարատեսակները համեմատելով նրանց մերձավոր տեսակների հետ, սովորաբար մենք նրանց մեջ նկատում ենք, ինչպես արդեն ասված է վերևում, հատկանիշներով ավելի նվազ միանմանություն, քան իսկական տեսակների ներկայացուցիչների մեջ։ Ընտանեցված ցեղերը երբեմն կարծես այլանդակ են թվում, դրանով ես ուզում եմ ասել, որ, զանազանվելով իրարից և տարբերվելով նույն սեռի այլ տեսակներից մի քանի չնչին հատկանիշներով, նրանք որևէ տեսակետից հաճախ տարբերվում են ինչպես մեկը մյուսից, այնպես էլ հատկապես իրենց մերձավոր բնական տեսակներից։ Այդ բացառությամբ (ինչպես նաև բացառությամբ խաչաձևվող տարատեսակների լրիվ պտղաբերության, որի մասին խոսք կլինի հետագա գլուխներից մեկում) միևնույն տեսակի ընտանեցված տարատեսակները միմյանցից տարբերվում են նույնքան, ինչքան որ նույն սեռի մերձավոր տեսակները բնական վիճակում. միայն այդ տարբերությունները շատ դեպքերում այնքան էլ խիստ չեն։ Սա պետք է համարել որպես ճշմարտություն, որովհետև ընտանեցված կենդանիների և մշակովի բույսերի շատ ցեղեր իրազեկ դիտողների կողմից ճանաչվում են որպես տեղական զանազան տեսակների սերունդներ, իսկ ուրիշ, նույնքան իրազեկ դատողների կողմից որպես նույն տեսակի տարատեսակներ։ Եթե ընտանի ցեղի և տեսակի միջև գոյություն ունենար մի որևէ ակնհայտ տարբերություն, ապա նման տարաձայնությունը այդքան հաճախ չէր հանդիպի։ Բազմիցս հայտնվել է այն կարծիքը, որ ընտանեցված ցեղերը երբեք չեն ցուցաբերում սեռային աստիճան ունեցող տարբերություններ։ Կարելի է ապացուցել, որ այդ կարծիքը ճիշտ չէ, բայց բնախույզները միմյանց հետ անհամաձայն են այն հարցի վերաբերմամբ, թե ո՞ր հատկանիշները պետք է համարել սեռային, բոլոր նման գնահատականները մինչև այժմ կրում են բացառապես էմպիրիկ բնույթ։ Երբ մենք կպարզենք, թե ինչպես են առաջանում սեռերը բնական դրության մեջ այն ժամանակ պարզ կլինի, որ մենք հիմք էլ չունենք սպասելու, որպեսզի մեր ընտանի ցեղերը կարողանային հաճախ հանդես բերել տարբերության աստիճաններ, որոնք հավասար են սեռայիններին։
Երբ մենք փորձում ենք որոշել իրար մերձավոր ընտանի ցեղերի տարբերության աստիճանը, մենք իսկույն ընկնում ենք կասկածների մեջ արդյոք նրանք մի՞ տեսակից են առաջացել, թե՞ մի քանի տեսակներից։ Այդ հարցը, եթե կարելի լիներ պարզել, մեծ հետաքրքրություն կներկայացներ. եթե, օրինակ, կարելի լիներ ցույց տալ, որ բարակի, խուզարկուի, տերյերի, սպանյոլների ու բուլդոգի ցեղերը, որոնք, ինչպես հայտնի է, իրենց հատկանիշները ժառանգաբար շատ լավ են փոխանցում, առաջացել են մեկ տեսակից, ապա նման փաստը կստիպեր մեզ կասկածել շատ մերձավոր տեսակների, օրինակ, աշխարհի զանազան մասերում ապրող աղվեսների անփոփոխելիության մասին։ Ես չեմ կարծում, ինչպես կերևա հետագա շարադրանքից, որ շների զանազան ցեղերի միջև նկատվող տարբերությունների ամբողջությունը առաջացել է ընտանեցման շնորհիվ. ընդհակառակը, ես կարծում եմ, որ տարբերությունների որոշ փոքր մասը կախված է այն բանից, որ նրանք առաջացել են առանձին տեսակներից։ Ինչ վերաբերում է մեր մի քանի այլ որոշակի նշաններով ջոկվող ընտանի ցեղերին, ապա կարելի է ենթադրել կամ նույնիսկ համոզված լինել, որ նրանք բոլորը առաջացել են մեկ տեսակից։
Հաճախ այն կարծիքն է հայտնվում, թե մարդը մշակության և ընտանեցման համար ընտրել է այնպիսի բույսեր ու կենդանիներ, որոնք աչքի են ընկել փոփոխվելու բացառիկ բնածին ընդունակությամբ, ինչպես նաև կլիմաների նկատմամբ դիմացկունությամբ։ Չեմ վիճարկի, որ այդ երկու հատկությունները զգալի չափով բարձրացրել են մեր ցեղերի մեծ մասի արժեքը, բայց առաջին անգամ կենդանուն ընտելացնող վայրենին ի՞նչպես կարող էր իմանալ, թե արդյոք այդ կենդանին կփոխվի՞ հաջորդ սերունդներում և հեշտությամբ կդիմանա այլ կլիմաների։ Մի՞թե էշի և սագի թույլ փոփոխականությունը կամ հյուսիսային եղջերուի թույլ աստիճանի դիմացկունությունը տաքության վերաբերմամբ, կամ ուղտինը՝ ցրտության վերաբերմամբ կարող էին խոչընդոտ հանդիսանալ նրանց ընտանեցման համար։ Ես չեմ կասկածում, որ եթե կենդանիների և բույսերի այլ տեսակներ, որոնք վերցված են բնական պայմաններից մեր ընտանի ցեղերին թվով հավասար և նույնչափ էլ բազմազան տարբեր դասերի ու տարբեր երկրների պատկանելիության իմաստով, ենթարկվեին ընտանեցման սերունդների նույնքան երկար շարանների ընթացքում, ապա նրանք միջին հաշվով կփոփոխվեին նույնպիսի չափերով, ինչպես որ մեր ընտանի ցեղերի նախնիներ եղած տեսակները։
Ինչ վերաբերում է մեր ամենահին ընտանեցված կենդանիներին և մշակովի բույսերին, ապա անհնարին է գալ մի որևէ եզրակացության այն մասին, թե նրանք ծագել են մեկ տեսակից, թե մի քանի տեսակներից։ Գլխավոր փաստարկը, որի վրա հիմնվում են մի քանի տեսակներից նրանց ծագման կողմնակիցները, սովորաբար այն է, որ ամենահին ժամանակներում արդեն, եգիպտական հուշարձաններում կամ Շվեցարիայի ցցաշենքերում, մենք հանդիպում ենք շատ բազմազան ցեղերի, ըստ որում նրանցից մի քանիսները շատ նման են ժամանակակից ցեղերին կամ մինչև անգամ նույնն են։ Բայց այդ կշռադատումները միայն հեռացնում են քաղաքակրթության սկիզբը և ցույց են տալիս, որ կենդանիները շատ ավելի վաղ են ընտելացվել, քան մինչև այժմ ենթադրվում էր։ Շվեցարական ցցաշենքերի բնակիչները մշակում էին ցորենի, զարու մի քանի տարատեսակներ, ոլոռ, խաշխաշ ձեթի համար և կտավատ, նրանք արդեն ունեին ընտանի կենդանիների ցեղեր։ Նրանք առևտուր էին անում այլ ժողովուրդների հետ։ Այդ բոլորը պարզ կերպով ապացուցում են, ինչպես նկատում է Հեերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 38] որ այդ վաղ շրջանում արդեն նրանք զգալի հաջողություններ են ունեցել քաղաքակրթության ասպարեզում, իսկ այդ իր հերթին ցույց է տալիս, որ դրանից առաջ եղել է ավելի ցածր քաղաքակրթության երկարատև մի շրջան, որի ընթացքում զանազան տոհմերի պահած ընտանի կենդանիները կարող էին փոփոխվել և մեկը մյուսից տարբերվող ցեղերի սկիզբ տալ։ Աշխարհի շատ մասերում երկրաբանական ավելի ուշ ժամանակի նստվածքներում կայծքարե գործիքներ գտնելուց հետո բոլոր երկրաբանները համոզված են, որ մարդը բարբարոս վիճակում գոյություն է ունեցել շատ հեռավոր ժամանակներում. մյուս կողմից մենք գիտենք, որ գրեթե չկա այնքան բարբարոս մի ցեղ, գոնե շուն ընտելացրած չլինի։
Մեր ընտանի կենդանիների մեծ մասի ծագումը, հավանորեն, ընդմիշտ կմնա անորոշ։ Բայց ես այստեղ կարող եմ հաղորդել, որ, ուշադրություն դարձնելով ամբողջ երկրագնդի շների վրա և մանրազնին առադրելով նրանց մասին հայտնի եղած բոլորը, ես եկել եմ այն եզրակացության, որ ընտելացված են եղել Canis սեռից մի քանի տեսակներ և որ նրանց արյունը, որոշ դեպքերում միմյանց հետ խառը, հոսում է մեր ընտանի ցեղերի երակներում։ Ոչխարների և այծերի մասին ես դժվարանում եմ որոշակի կարծիք հայտնել։ Հնդկական սապատավոր տավարի սովորություններին, ձայնին, կազմությանը և կառուցվածքին վերաբերող այն փաստերի հիման վրա, որ ինձ հաղորդել է միստր Բլիտը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 39] ինձ համարյա ակներև է դառնում, որ նա այլ ծագում ունի, քան եվրոպական տավարը, իսկ մի քանի գիտակ դատողներ այն կարծիքին են, որ վերջինս էլ առաջացել է երկու կամ երեք նախահայր ձևերից, թեպետ անհայտ է, թե կարելի՞ է արդյոք նրանց համարել ինքնուրույն տեսակներ։ Այդ եզրակացությունները, ինչպես նաև սապատավոր և սովորական տավարի միջև եղած տեսակային տարբերության մասին արտահայտված կարծիքը կարելի է ապացուցված համարել պրոֆեսոր Ռյուտիմեյերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 40] կատարած հիանալի հետազոտություններից հետո։ Ձիերի վերաբերմամբ ևս կշռադատումների հիման վրա, որոնք այստեղ բերել չեմ կարող, որոշ տատանումներով հակվում եմ շատ հեղինակների կարծիքին հակառակ եզրակացության, որ նրանց բոլոր ցեղերը պատկանում են մեկ տեսակի։ Ես բուծել եմ համարյա բոլոր անգլիական հավերի ցեղերը, կատարել եմ նրանց միջև խաչաձևումներ և հետազոտել նրանց կմախքները, և այդ բոլոր տեղեկությունների հիման վրա ինձ համար գրեթե ակներև է դառնում, որ նրանք բոլորը ծագել են Հնդկաստանում վայրի վիճակում հանդիպող Gallus bankiva-ից. նույն եզրակացությանն են եկել միստր Բլիտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 39] և ուրիշներ, որոնք ուսումնասիրել են այդ թռչունին Հնդկաստանում։ Ինչ վերաբերում է բադերին և ճագարներին, որոնց մի քանի ցեղերը խիստ տարբերվում են միմյանցից, ապա կասկածի ենթակա չէ, որ նրանք բոլորը ծագել են վայրի բադից և ճագարից։
Այն ուսմունքը, որ մեր ընտանի զանազան ցեղերը առաջացել են մի քանի բնական տեսակներից, մի քանի հեղինակներ հասցրել են անհեթեթության։ Նրանք կարծում են, որ ամեն մի ցեղ, որը ժառանգաբար փոխանցում է իր թեկուզև աննշան տարբերիչ հատկանիշները, առաջացել է մի վայրենի նախատիպից։ Այսպես դատելով, մենք պետք է գանք այն եզրակացության, որ գոյություն են ունեցել տավարի մինչև քսան տեսակներ, նույնքան էլ ոչխարի տեսակներ, այծի մի քանի տեսակներ միայն Եվրոպայում և նույնիսկ մի քանի տեսակներն էլ Մեծ Բրիտանիայի սահմաններում։ Մի հեղինակ ենթադրում է, որ գոյություն են ունեցել միայն Մեծ Բրիտանիային հատուկ ոչխարի տասնմեկ վայրի տեսակներ։ Եթե հիշենք, որ Բրիտանիան չունի բացառապես իրեն հատուկ ոչ մի կաթնասուն, իսկ Ֆրանսիան ունի շատ սակավաթիվ կաթնասուններ, որոնք չեն հանդիպում Գերմանիայում, և որ նույնը ճիշտ է նաև Հունգարիայի, Իսպանիայի և այլոց վերաբերմամբ, եթե միաժամանակ հիշենք, որ այդ երկրներից յուրաքանչյուրն ունի տավարի, ոչխարի և այլն մի քանի ցեղեր, որոնք բացառապես հատուկ են իրեն, ապա պիտի ընդունենք, որ Եվրոպայում ծագում են առել մի քանի ընտանի ցեղեր, որովհետև այլ կերպ նրանք որտեղի՞ց կարող էին հայտնվել։ Այդ ճիշտ է նաև Հնդկաստանի վերաբերմամբ։ Նույնիսկ աշխարհի բոլոր մասերում տարածված շների ցեղերի վերաբերմամբ, որոնք, ինչպես ես ենթադրում եմ, առաջացել են մի քանի վայրի տեսակներից, չի կարող կասկած լինել, որ նրանց մեջ շատ բան պետք է վերագրել ժառանգված փոփոխություններին, որովհետև ո՞վ կհավատա, թե իտալական բարակի, խուզարկու շան, բուլդոգի, մոպսի, բլենհեյմյան սպանյոլի պես կենդանիները, որոնք այնքան տարբերվում են Canis սեռի բոլոր վայրի ներկայացուցիչներից, մի ժամանակ գոյություն են ունեցել բնական դրության մեջ։ Հաճախ նաև ենթադրություն է արվել, որ մեր շների բոլոր ցեղերը առաջացել են մի քանի վայրի տեսակների խաչաձևման միջոցով, բայց այդ միջոցով կարելի է ստանալ միայն այնպիսի ձևեր, որոնք ծնողների ձևերի միջև գրավում են միջակա տեղ. և եթե մենք կամենում ենք այդ ձևով մեզ բացատրել մեր ընտանի ցեղերի ծագումը, ապա պիտի ընդունենք, որ վայրի դրության մեջ նախապես գոյություն են ունեցել ամենածայրահեղ ձևերը, ինչպիսիք են իտալական բարակը, խուզարկուն, բուլդոգը և այլն։ Բացի դրանից, իրարից խիստ տարբերվող ցեղերի առաջացման հնարավորությունը խաչաձևման միջոցով շատ չափազանցված է։ Կան շատ օրինակներ, որոնք ապացուցում են, որ ցեղը երբեմն կարելի է խաչաձևման միջոցով փոփոխել, եթե խնամքով ընտրվեն ցանկալի հատկանիշով օժտված անհատներ, բայց իրարից խիստ տարբերվող երկու ցեղերի միջև միջակա ցեղ առաջացնելը չափազանց դժվարին կլիներ։ Սըր Ջ. Սեբրայտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 41] հատուկ այդ նպատակով փորձեր է արել և անհաջողություն է կրել։ Երկու մաքուր ցեղերի առաջին խաչաձևումից ստացված ձագերը (ինչպես ես համոզվեցի աղավնիների նկատմամբ) իրենց հատկանիշներով բավականաչափ, իսկ երբեմն էլ միանգամայն միանման են, և ամեն ինչ շատ պարզ է թվում. բայց հենց որ սկսում են այդ խառնացեղերը միմյանց հետ խաչաձևել, մի քանի սերունդների ընթացքում չեն ստացվում երկու իրար նման էակներ,— և այն ժամանակ միայն երևում է խնդրի ամբողջ դժվարությունը։
ԸՆՏԱՆԻ ԱՂԱՎՆԻՆԵՐԻ ՑԵՂԵՐԸ, ՆՐԱՆՑ ԾԱԳՈՒՄԸ ԵՎ ՏԱՐԲԵՐՈՒԹՅՈՒՆՆԵՐԸ
Համոզված լինելով, որ ամեն մի հարց ամենից ավելի լավ կարելի է ուսումնասիրել մի որևէ հատուկ խմբի վրա, ես երկար կշռադատելուց հետո կանգ առա ընտանի աղավնիների վրա։ Ես բուծում էի այն բոլոր ցեղերը, որ կարող էի գնել կամ այլ կերպով ձեռք բերել, ինձ շատ սիրալիր կերպով մատակարարել են աղավնիների խրտվիլակներ զանազան երկրներից, հատկապես Հնդկաստանից Ու. Էլլիոտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 42] և Պարսկաստանից Կ. Մուրրեյը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 43] Աղավնիների մասին կան զանազան լեզուներով ամբողջ աշխատություններ, նրանցից մի քանիսը շատ կարևոր են, որովհետև վերաբերում են հին ժամանակներին։ Ես կապ էի պահպանում աղավնիների ցեղերի բուծման շատ հայտնի մասնագետների հետ, իսկ Լոնդոնյան երկու աղավնաբուծական ակումբ ինձ ընդունել էին իրենց անդամ։ Ցեղերի բազմազանությունը իսկապես զարմանալի է։ Համեմատեցեք անգլիական փոստատարը կարճակտուց թուրմանի հետ և ուշադրություն դարձրեք նրանց կտուցների տարբերության և դրանց հետ կապված գանգերի ձևի տարբերության վրա։ Փոստատարները, առանձնապես արուները, աչքի են ընկնում նաև գլխի վրա ունեցած մսոտ ուռուցքների զարմանալի զարգացմամբ, և դրան ուղեկցում են խիստ երկարացած կոպերը, շատ մեծ արտաքին քթանցքները և լայն բացվող բերանը։ Կարճակտուց թուրմանի կտուցը իր գծագրով նմանվում է սերինոսի կտցին, իսկ սովորական թուրմանը աչքի է ընկնում խմբով շատ բարձր թռչելու և դարձդարձիկ շարժումներով իրեն ցած գցելու ժառանգական սովորությամբ։ Իսպանական կամ հռոմեական աղավնին (Runt) շատ խոշոր թռչուն է երկար, հոծ կտուցով և մեծ ոտնթաթերով. այդ թռչունի ենթացեղերից մի քանիսն ունեն շատ երկար պարանոց, մյուսները՝ շատ երկար թևեր ու պոչ, կամ, ընդհակառակը, շատ կարճ պոչ։ Հնդկական կամ լեհական աղավնին (Barb) հատկություններով նման է փոստատարին, բայց երկար կտուցի փոխարեն ունի կարճ ու լայն կտուց։ Փքունը (Pouter) ունի շատ երկարավուն մարմին, թևեր և ոտներ, նրա խիստ զարգացած խպիպը, որը նա հպարտությամբ ուռցնում է, առաջ է բերում զարմանք և նույնիսկ ծիծաղ։ Ճայ-աղավնին (Turbit) ունի կարճ կոնաձև կտուց և մի շարք խճճված փետուրներ, որոնք ձգվում են կրծքի երկարությամբ. նա սովորություն ունի շարունակ թեթևակի ուռցնելու կերակրափողի վերին մասը։ Յակոբինյան աղավնու (Jacobin) պարանոցի ետևի փետուրները այնքան շրջված են, որ մի տեսակ կնգուղ են կազմում, բացի դրանից, նրա մեծության համապատասխանորեն երկարացած են թևերի և պոչի փետուրները։ Փողհարը (Trumpeter) և ծաղրածուն (Laugher), ինչպես ցույց են տալիս անունները, արձակում են այնպիսի ձայներ, որոնք տարբերվում են մյուս ցեղերի արձակած ձայներից։ Սիրամարգանման աղավնին (Fantail) ունի երեսուն կամ քառասուն պոչափետուր փոխանակ տասներկուսի կամ տասներեքի՝ այսինքն փոխանակ այն թվի, որը նորմալ է աղավնիների այդ ընդարձակ ընտանիքի բոլոր ներկայացուցիչների համար. այդ փետուրները միշտ բացված են և ուղղված են դեպի վեր, այնպես որ այդ ցեղի լավ ներկայացուցիչների պոչի փետուրները կպչում են գլխին. ճարպագեղձը բոլորովին ծյուրված է։ Կարելի էր թվել էլի մի քանի ավելի քիչ ցայտուն կերպով արտահայտված ցեղեր։
Զանազան ցեղերի կմախքների մեջ մեծ տարբերություններ կան դիմային ոսկրների երկարության, լայնության և կորության տեսակետից։ Ստորին ծնոտի ճյուղի ձևը, երկարությունը և լայնությունը մեծ չափով են տարբերվում։ Թվով տատանվում են պոչի և սրբոսկրի ողները, ինչպես և կողերը, որոնք տարբերվում են նաև հարաբերական լայնությամբ և ելուստների ներկայությամբ։ Կրծոսկրի անցքերի մեծությունը և ձևը, ինչպես նաև ճնաղի երկու ճյուղերի իրարից հեռացման աստիճանը և հարաբերական մեծությունը չափազանց փոփոխական են։ Բերանի բացվածքի հարաբերական մեծությունը, կոպերի հարաբերական երկարությունը, քթանցքների, լեզուն (որը միշտ չէ, որ համապատասխանում է կտուցի երկարությանը), խպիպի և կերակրափողի վերին մասի մեծությունը, պոչուկի ճարպագեղձի զարգացումը և ատրոֆիայի ենթարկվածությունը, թափափետուրների և ղեկափետուրների թիվը, թևերի և պոչի հարաբերական երկարությունը ինչպես միմյանց, այնպես էլ ամբողջ մարմնի չափերի նկատմամբ, ոտքի և ոտնաթաթի հարաբերական երկարությունը, մատների վրա եղած վահանիկների թիվը և մատների միջև գտնվող մաշկի զարգացումը՝ այս բոլորը կառուցվածքի առանձնահատկություններ են, որոնք ենթակա են փոփոխականության։ Իսկական փետրավորման ժամանակը, ինչպես նաև ձվից դուրս եկող ձագերին ծածկող աղվամազի վիճակը նույնպես փոփոխական են։ Ձվի ձևը և մեծությունը տատանվում են։ Ուշագրավ տարբերություններ կան թռիչքի և մի քանի ցեղերի ձայնի ու բարքերի մեջ։ Վերջապես, մի քանի ցեղերի մեջ արուներն ու էգերը թույլ տարբերություն են դրսևորում։
Ի վերջո կարելի էր հավաքել մոտ քսան զանազան աղավնիներ, որոնց ամեն մի թռչնաբան կհամարեր միանգամայն սահմանազատված տեսակներ, եթե նրան ասեին, թե այդ թռչունները գտնված են վայրի վիճակում։ Բացի ղրանից, ես չեմ կարծում, թե որևէ թռչնաբան անգլիական փոստատարին, կարճակտուց թուրմանին, իսպանական աղավնուն, հնդկական աղավնուն, փքունին և սիրամարգանման աղավնուն կդասեր միևնույն սեռին, մանավանդ որ բոլոր այդ ցեղերում կան միանգամայն ժառանգական հատկանիշներ ունեցող ենթացեղեր, որոնց նա կանվաներ տեսակներ։
Ինչքան էլ մեծ լինեն աղավնիների ցեղերի միջև եղած տարբերությունները, ես միանգամայն համոզված եմ, որ արդարացի է բնագետների մեջ տարածված այն կարծիքը, թե նրանք բոլորը առաջացել են ժայռաղավնուց[37] (Columba livia), հասկանալով այդ տերմինի տակ մի քանի աշխարհագրական ցեղեր կամ ենթատեսակներ, որոնք իրարից տարբերվում են շատ աննշան չափով։ Քանի որ ինձ այդ եզրակացություններին բերող հիմունքները կիրառելի են նաև այլ դեպքերի համար, ուստի համառոտակի կբերեմ դրանք այստեղ։ Եթե մեր զանազան ցեղերը ժայռաղավնուց ծագում առած տարատեսակներ չեն, այդ դեպքում նրանք պետք է սերված լինեն առնվազն յոթ կամ ութ բնական տեսակներից, որովհետև ավելի պակաս թվով ձևեր խաչաձևելու միջոցով անհնարին է ստանալ մեր ընտանի ցեղերը։ Օրինակ, ինչպես կարելի էր փքունը ստանալ խաչաձևման միջոցով, եթե ծնողներից գոնե մեկը չունենար բնորոշ հսկայական խպիպը։ Բոլոր ենթադրյալ վայրի նախահայրերը պետք է լինեին ժայռաղավնիներ, այսինքն չբնակալող և նույնիսկ ծառերին դժկամությամբ նստող թռչուններ։ Բայց բացի Columba livia-ից և նրա աշխարհագրական ենթատեսակներից, հայտնի են ընդամենը երկու կամ երեք տեսակ ժայռաղավնիներ, և նրանք չունեն ընտանի ցեղերի տարբերիչ հատկանիշներից ոչ մեկը։ Այստեղից պետք է եզրակացնել, որ կամ այդ վայրի նախնիները դեռ գոյություն ունեն այն երկրներում, որտեղ նրանք ընտանեցված են եղել, բայց թռչնաբաններին անհայտ են մնացել, որը խիստ անհավանական է, եթե ուշադրության առնենք այդ թռչունների մեծությունը, կենսակերպը և զարմանալի հատկանիշները, կամ թե չէ նրանք բոլորը բնաջնջվել են բնական վիճակում։ Սակայն անդունդների վրա բուն դնող և լավ թռչող թռչուններին ոչնչացնելը այնքան էլ հեշտ չէ, և սովորական ժայռաղավնին, որը մեր ընտանի ցեղերի նման կենսակերպ ունի, դեռ չի ոչնչացվել նույնիսկ Մեծ Բրիտանիայի մի քանի փոքր կղզիներում և Միջերկրական ծովի ափերին։ Այսպիսով, այն ենթադրությունը, թե ժայռաղավնու նման կենսակերպ ունեցող այդքան մեծ թվով տեսակները ոչնչացել են, կլիներ չափազանց անմտածված։ Բացի դրանից, վերևում թված ընտանեցված ցեղերից մի քանիսը տարվել են աշխարհի զանազան կողմերը, և նրանցից ոմանք կարող էին ընկնել նաև իրենց հայրենիքը, բայց նրանցից ոչ մեկը չի վայրենացել, թեև սովորական աղավնին (dovecot pigeon), որը ներկայացնում է միայն թույլ կերպով փոփոխված ժայռաղավնուն, մի քանի տեղերում վայրենացել է։ Վերջապես, այդ գործում մեր ունեցած ժամանակակից ամբողջ փորձը ցույց է տալիս, որ չափազանց դժվար է վայրի կենդանիներին ստիպել բազմանալ ընտանի վիճակում, իսկ եթե հետևենք այն հիպոթեզին, թե աղավնիները ծագել են շատ տեսակներից, ապա պիտի ընդունենք, որ շատ հին ժամանակներում կիսաքաղաքակիրթ մարդիկ առնվազն յոթ կամ ութ տեսակ այն աստիճան են ընտանեցրել, որ նրանք գերության մեջ միանգամայն պտղաբեր են դարձել։
Շատ ուժեղ ապացույց է ծառայում այլ դեպքերում կիրառելի այն փաստը, որ բոլոր հիշված ցեղերը, լինելով վայրի ժայռաղավնուն նման իրենց ընդհանուր կազմությամբ, կենսակերպով, ձայնով, գույնով և կառուցվածքի առանձնահատկությունների մեծ մասով, որոշ տեսակետներից ներկայացնում են բոլորովին աննորմալ շեղումներ, օրինակ, զուր տեղը մենք կորոնեինք Columbidae մեծ ընտանիքում այնպիսի կտուց, ինչպիսին ունի անգլիական փոստատարը, կարճակտուց թուրմանը կամ հնդկականը, այնպիսի խճճված փետուրներ, ինչպիսիք ունի յակոբինյան աղավնին, խպիպ՝ ինչպիսին ունի փքունը, պոչի փետուրներ՝ ինչպիսին ունի սիրամարգանման աղավնին։ Այստեղից պետք կլիներ ընդունել, որ կիսաքաղաքակիրթ մարդուն ոչ միայն լիովին հաջողվել է ընտելացնել մի քանի տեսակներ, այլև նա դիտավորյալ կամ պատահաբար ընտրել է բացառապես աննորմալ տեսակներ, և վերջապես, որ հենց այդ բոլոր տեսակները մահաջնջվել են կամ անհայտ են մնացել։ Հանգամանքների այսպիսի տարօրինակ զուգորդումը վերին աստիճանի անհավատալի է։
Աղավնիների գույնին վերաբերող մի քանի փաստեր նույնպես արժանի են ուշադրության։ Ժայռաղավնին ունի գորշակապույտ գույն, սպիտակ փորիկ, բայց հնդկական ենթատեսակի՝ C. intermedia Ստրիկլենդի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 44] այդ մասը կապույտ գույն ունի։ Պոչն ունի իր ծայրին մուգ գույնի լայնական շերտ, իսկ նրա արտաքին փետուրները դրսից և հիմքի մոտ եզրապատված են սպիտակ գույնով։ Թևերի վրա կան երկու սև շերտեր։ Մի քանի կիսաընտանեցված ցեղեր և մի քանի անտարակուսելի վայրենի ցեղեր, բացի այդ շերտերից, ծածկված են նաև սև բծերով։ Այս բոլոր հատկանիշները ամբողջությամբ միասին մենք չենք հանդիպում այս ընտանիքի մնացած տեսակներից ոչ մեկի վրա։ Մինչդեռ մեր ընտանի ցեղերից ամեն մեկի վրա, եթե իհարկե նրանք պահվում են խնամքով, բոլոր վերոհիշյալ նշանները, չբացառելով պոչի արտաքին փետուրների սպիտակ եզրապատումը, երբեմն երևան են դալիս լիակատար ակներևությամբ։ Բացի դրանից, երբ խաչաձևում ենք երկու կամ ավելի թվով ցեղեր, որոնք չունեն նշված գույնը և նշանները, խառնացեղերը շատ հաճախ հանկարծ երևան են հանում դրանք։ Կբերեմ իմ դիտած մի քանի դեպքերից միայն մեկը։ Ես խաչաձևեցի սպիտակ փողհարներին, որոնք իրենց հատկանիշները փոխանցում են մեծ կայունությամբ, սև հնդկականների (Barb) հետ, որոնց երկնագույն (կապտագույն) տարատեսակները այնքան հազվագյուտ են, որ ինձ այդ գույնով ոչ մի օրինակ հայտնի չէ Անգլիայում։ Ստացվեցին սև, մուգ դարչնագույն և բծավոր հիբրիդներ։ Ես խաչաձևեցի նաև հնդկական աղավնին (Barb) բծավորի (Spot) հետ. այս վերջին թռչունը սպիտակ է, շեկ պոչով և ճակատին շեկ բծով և նույնպես աչքի է ընկնում հիանալի կայունությամբ։ Հիբրիդները մուգ գորշ էին և բծավոր։ Այնուհետև ես միմյանց հետ խաչաձևեցի փողհար-հնդկական հիբրիդները հնդկական-բծավոր հիբրիդների հետ, և ստացվեցին հիանալի երկնագույն (կապտագույն) թռչուններ, որոնց փորիկը սպիտակ էր, թևերը երկու սև շերտով, պոչի շերտավոր և սպիտակ եզր ունեցող փետուրներով, բոլորովին նման ժայռաղավնուն։ Մենք կարող ենք բացատրել այս փաստերը, ելնելով նախածնողների հատկանիշներին վերադարձի հայտնի սկզբունքից, միայն պետք է ընդունենք, որ բոլոր ընտանի ցեղերն առաջացել են ժայռաղավնուց։ Իսկ եթե մենք հրաժարվենք այդ բացատրությունից, այդ դեպքում պետք է դիմենք հետևյալ երկու խիստ անհավանական ենթադրություններից որևէ մեկին, այն է՝ առաջինը, կամ պիտի ընդունենք, որ բոլոր ենթադրյալ բնական տեսակներն ունեցել են ժայռաղավնու գույնը և նշանները, թեպետև գոյություն ունեցող տեսակներից ոչ մեկը չունի դրանք, և այն ժամանակ ամեն մի ցեղ պիտի ձգտում ունենար վերադառնալու նախկին հատկանիշներին, երկրորդ, կամ թե մենք պետք է ընդունենք, որ ամեն մի, նույնիսկ ամենամաքուր ցեղ տասներկու կամ ծայրահեղ դեպքում քսան սերունդ մեր ժամանակից առաջ խաչաձևված է եղել ժայռաղավնու հետ։ Ես ասում եմ տասներկու կամ քսան սերունդ, որովհետև հայտնի չէ մի որևէ դեպք, որ կենդանին վերադառնա ավելի մեծ թվով սերունդներ առաջ եղած օտար արյուն ունեցող նախորդի հատկանիշներին։ Միայն մեկ անգամ խաչաձևման ենթարկված ցեղի մեջ շարունակ ավելի ու ավելի թուլանում է այդ խաչաձևումից ձեռք բերված հատկանիշին վերադառնալու ձգտումը, որովհետև սերնդից սերունդ օտար արյան խառնուրդը պիտի քչանա, բայց եթե ոչ մի խաչաձևում չի եղել, իսկ ցեղի մեջ ձգտում կա վերադառնալու նախարդող որևէ սերնդում կորած հատկանիշին, ապա մենք պատճառ չենք տեսնում, որ այդ ձգտումը առանց թուլանալու չփոխանցվի անսահման թվով սերունդների ընթացքում։ Նախնիների հատկանիշներին վերադառնալու այս երկու միանգամայն տարբեր դեպքերը շատ հաճախ խառնել են այն հեղինակները, որոնք գրել են ժառանգականության մասին։
Վերջապես, աղավնու բազմապիսի ցեղերի հիբրիդները կամ խառնացեղերը միանգամայն պտղաբեր են, ինչպես ես այդ կարող եմ հաստատել իմ սեփական փորձերի հիման վրա, որ ես դիտմամբ այդ նպատակով կատարել եմ միմյանցից ամենից ավելի խիստ տարբերվող ցեղերի վրա։ Սակայն հազիվ թե գտնվի գոնե մի լավ ուսումնասիրված դեպք, որից երևա, որ կենդանիների երկու միմյանցից տարբերվող տեսակների խառնացեղերը լրիվ պտղաբեր են։ Մի քանի հեղինակներ հայտնել են այն կարծիքը, որ երկար ժամանակ ընտանեցված վիճակում գտնվելը նվազեցնում է տեսակների անպտղության հակումը։ Շների և մի քանի այլ ընտանի կենդանիների պատմությունը, ըստ երևույթին, հաստատում է այդ եզրակացությունը միմյանց մոտիկ տեսակների վերաբերմամբ։ Բայց այդ եզրակացության այն աստիճան ընդհանրացումը, որի հիման վրա ենթադրվեր, թե բնական վիճակում գտնվող և այնքան միմյանցից տարբերվող տեսակները, ինչպիսիք են փոստատար աղավնիները, թուրմանները, փքունները և սիրամարգանմանները, կկարողանային փոխադարձ խաչաձևումից առաջ բերել միանգամայն պտղաբեր սերունդ, կլիներ չափազանց չմտածված։
Այս բոլոր նկատառումների հիման վրա, այսինքն՝ անհավանականությունը, որ մարդը կարող էր աղավնու յոթ կամ ութ ենթադրյալ տեսակներ ընտանեցման վիճակում լրիվ պտղաբեր դարձնել. որ այդ ենթադրյալ տեսակները մնացել են բոլորովին անհայտ վայրի վիճակում և ոչ մի տեղ նորից չեն վայրենացել. որ բոլոր այդ տեսակները միանգամայն աննորմալ են Columbidae-ի ընտանիքի մնացած բոլոր ներկայացուցիչների համեմատությամբ և միևնույն ժամանակ շատ կողմերով նման են ժայռաղավնուն. ինչպես նաև այն հիման վրա, որ ժայռաղավնու կապտավուն գույնը և նշանները երբեմն երևում են ինչպես զտարյուն, այնպես էլ խաչաձևվող բոլոր ցեղերի մեջ, և վերջապես, բոլոր ցեղերի խառնացեղերից ստացված սերնդի լրիվ պտղաբերության հիման վրա, այս բոլոր նկատառումները ամբողջությամբ վերցրած հիմք են տալիս եզրակացնելու, որ մեր բոլոր ընտանի աղավնացեղերն առաջացել են ժայռաղավնուց՝ Columba livia-ից և նրա աշխարհագրական ենթատեսակներից։
Ի պաշտպանություն այդ տեսակետի ես կարող եմ բերել, նախ, այն փաստը, որ վայրի Columba livia-ն թե՛ Եվրոպայում և թե՛ Հնդկաստանում ընտանեցման համար ընդունակություն է դրսևորել և նա ինչպես իր կենսակերպով, այնպես էլ կառուցվածքի շատ առանձնահատկություններով նման է մեր ընտանի բոլոր ցեղերին։ Երկրորդ, թեև անգլիական փոստատարը կամ կարճակտուց թուրմանը որոշ կողմերով խիստ տարբերվում են ժաչռաղավնուց, այնուամենայնիվ, համեմատելով այդ ռասաների՝ մանավանդ տարբեր երկրներից ստացվածների զանազան ենթացեղերը, մենք կարող ենք կազմել գրեթե մի անընդհատ շարք, որը նրանց կապում է ժայռաղավնու հետ։ Նույնը հնարավոր է նաև այլ ցեղերի, թեև ոչ բոլորի, վերաբերմամբ։ Երրորդ, յուրաքանչյուր ցեղի համար բնորոշ հատկանիշները աչքի են ընկնում առանձին փոփոխականությամբ. այդպես են, օրինակ, փոստատարի կատարը և կտուցի երկարությունը, թուրմանի կտուցի կարճությունը և սիրամարգանմանի պոչի փետուրների թիվը և այս փաստի բացատրությունը ակներև կդառնա, երբ մենք կխոսենք ընտրության մասին։ Չորրորդ, աղավնիները շատ ժողովուրդների ջանադիր հոգատարության և սիրո առարկան են եղել։ Երկրագնդի զանազան մասերում մեր ժամանակից հազարավոր տարիներ առաջ նրանք արդեն ընտելացված են եղել։ Ամենահին տեղեկությունը աղավնիների գոյության մասին վերաբերում է եգիպտական հինգերորդ դինաստիային, այսինքն մեր թվականությունից մոտավորապես 3000 տարի առաջ, ինչպես ինձ պարզաբանել է պրոֆեսոր Լեպսիուսը.[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 45] բայց միստր Բիրչը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 46] հաղորդեց ինձ, որ նրանք հիշատակվում են մի խոհանոցային հաշվի մեջ, որը վերաբերում է նախորդ դինաստիային։ Ինչպես տեղեկանում ենք Պլինիուսից,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 47] հռոմայեցիները աղավնիների համար հսկայական գումարներ էին վճարում. «Բանն այնտեղ էր հասնում, որ հաշվում էին նրանց տոհմագրությունը և տոհմը»։ Հնդկաստանում մոտ 1600 թվականին Ակբար-Խանը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 48] շատ էր գնահատում աղավնիներին, և ոչ պակաս քան 20000 այդ թռչուններից ուղեկցում էին նրա արքունիքը։ «Իրանի և Թուրանի միապետները ուղարկում էին նրան հազվագյուտ թռչուններ», և «Նորին մեծությունը»,— շարունակում է պալատական պատմաբանը,— «կատարելով ցեղերի միջև խաչաձևում, որ նրանից առաջ ոչ մի ժամանակ չէին արվել, զարմանալի կերպով կատարելագործեց նրանց»։ Նույն ժամանակներին մոտ հոլանդացիներն էլ աղավնիների այդպիսի սիրահարներ էին, ինչպես հին հռոմայեցիները։ Այս նկատառումների կարևոր նշանակությունը աղավնիների կրած խոր փոփոխությունների բացատրության համար մեզ նույնպես պարզ կլինի, երբ խոսք կլինի ընտրության մասին։ Մենք այն ժամանակ կտեսնենք, թե ինչու զանազան ցեղեր այդպես հաճախ ունեն այլանդակությունների բնույթ։ Նոր ցեղերի առաջացման համար մեծ նշանակություն ունի այն փաստը, որ աղավնիների արուներն ու էգերը հեշտությամբ զույգեր են կազմում ամբողջ կյանքի ընթացքում. այդ հանգամանքի շնորհիվ զանազան ցեղեր կարելի է պահել միևնույն թռչնանոցում։
Ես ընտանի աղավնու ցեղերի հավանական ծագման հարցը այստեղ փոքր ինչ ավելի, թեպետև ոչ բավարար չափով մանրամասնորեն պարզաբանեցի այն պատճառով, որ, սկսելով աղավնու ցեղերի բուծումը և գիտենալով, թե ինչքան երանք կայուն են իրենց հատկանիշներում, ես նույնպես նույնքան քիչ էի հակված ընդունելու նրանց ծագումը հենց ընտանեցման սկզբից մեկ ընդհանուր նախահորից, որքան որևէ բնախույզ բնական դրության մեջ ապրող սերինոսների կամ այլ թռչունների բազմաթիվ տեսակների վերաբերմամբ։ Ինձ չափազանց զարմացրեց մի հանգամանք, այսինքն այն, որ համարյա բոլոր անասնաբույծերը և բոլոր բուսաբույծները, որոնց հետ ես պատեհություն եմ ունեցել խոսելու կամ նրանց գրվածքները կարդալու, հաստատապես համոզված են, որ այն ցեղերը, որոնց հետ նրանք գործ են ունեցել, առաջացել են միմյանցից տարբեր համապատասխան թվով բնական տեսակներից։ Հարցրեք, ինչպես ես այդ արել եմ բազմիցս, որևէ հայտնի անասնաբույծի, որը բուծում է հերեֆորդյան տավար, արդյոք չէ՞ր կարող նրա ցեղը առաջացած լինել երկարաեղջյուր (լանգհորն) տավարից կամ երկու ցեղերը մի ընդհանուր ծնողներից, և նա կծիծաղի ձեզ վրա։ Ես դեռ չեմ հանդիպել ոչ մի աղավնի, հավ, բադ կամ ճագար պահել սիրողի, որը խորապես համոզված չլիներ, որ յուրաքանչյուր հիմնական ցեղ ծագում է մի ինքնուրույն տեսակից։ Վան-Մոնսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 49] խնձորների և տանձերի մասին գրած իր աշխատության մեջ լիովին կասկածում է այն բանում, որ զանազան սորտեր, ինչպես, օրինակ, Ribston pipplin-ը կամ Codlin-ը կարող էին երբևիցե առաջանալ միևնույն ծառի սերմերից։ Ես կարող եմ բերել անհամար օրինակներ։ Բացատրությունը, ես կարծում եմ, շատ պարզ է, երկարատև ուսումնասիրության հետևանքով մասնագետները խորապես համակվում են իրենց հետաքրքրող ցեղերի միջև եղած տարբերություններով, և չնայած այն բանին, որ նրանք շատ լավ գիտեն, թե որքան փոփոխական են այդ ցեղերը, փոքրիկ շեղումների սահմաններում, քանի որ հենց իրենք մրցանակներ են ստանում այդ շեղումների ընտրության համար,— հրաժարվում են որևէ ընդհանրացումից, այսինքն մտքով գումարելուց այն տարբերությունները, որոնք կուտակվում են սերունդների երկար շարքերի ընթացքում։ Այն բնախույզները, որոնք ժառանգականության օրենքների մասին անհամեմատ ավելի քիչ գիտեն, քան անասնաբույծները, և նրանց պես նույնքան քիչ գիտեին մեր ընտանի ցեղերի նախնիների երկար շարքը շաղկապող օղակների մասին, բայց և այնպես ընդունում են, որ ցեղերն առաջացել են ընդհանուր ծնողներից, արդյոք նրանք այստեղից զգուշության դաս չպիտի՞ վերցնեն և արդյոք չպիտի՞ դադարեն այսուհետև ծանակել այն գաղափարը, որի համաձայն բնական տեսակները ևս այլ տեսակների միայն անմիջական հետնորդներն են։
ՀՆՈՒՄ ԱՐԴԵՆ ԿԻՐԱՌՎԱԾ ԸՆՏՐՈՒԹՅԱՆ ՍԿԶԲՈՒՆՔԸ, ՆՐԱ ԿԻՐԱՌՄԱՆ ՀԵՏԵՎԱՆՔՆԵՐԸ
Այժմ տեսնենք, թե ինչպես աստիճանաբար առաջացել են մեր ընտանի ցեղերը, միևնույն է՝ մեկ կամ մի քանի մերձավոր ցեղերից։ Այդ հետևանքների մի մասը կարելի է վերագրել արտաքին պայմանների որոշակի և անմիջական ներգործությանը և մասամբ էլ սովորությանը, բայց հանդուգն կլիներ այն մարդը, որը կփորձեր բացառապես այդ գործոններին վերագրել ծանրաքաշ և արշավային ձիերի միջև, բարակ և խուզարկու շների, փոստատար և թուրման աղավնիների միջև եղած տարբերությունները։ Մեր ընտանի կենդանիների ամենազարմանալի առանձնահատկություններից մեկն այն է, որ մենք նրանց մեջ տեսնում ենք աներկբայելի հարմարումներ, իհարկե, ոչ կենդանու կամ բույսի իրեն օգտին, այլ մարդու պահանջմունքների կամ քմահաճույքի օգտին։ Մարդու համար մի քանի օգտակար փոփոխություններ կարող էին առաջանալ հանկարծակի կերպով կամ մի թռիչքով, այսպես, շատ բուսաբաններ կարծում են, որ խավատու կոները, որոնք չի կարելի փոխարինել ոչ մի մեխանիկական հարմարանքներով, Dipsacus բնական տեսակի միայն տարատեսակն են, և այդպիսի սահմաններում շեղումը կարող էր առաջանալ հանկարծակի կերպով սերմնաբույսի մեջ։ Նույն կշռադատությունը հավանորեն կիրառելի է նաև շների և ստույգ կերպով հայտնի է անկոնական ոչխարի վերաբերմամբ։ Բայց երբ մենք համեմատում ենք ծանրաքաշ ձին արշավաձիու հետ, միսապատանի ուղտը երկսապատանիի հետ, մարգագետնային կամ լեռնային արոտներին հարմարված ոչխարների տարբեր ցեղերը, որոնց բուրդը մի դեպքում պիտանի է մի նպատակի, մյուս դեպքում մի այլ նպատակի համար, երբ մենք համեմատում ենք շների տարբեր ցեղերը, որոնք մարդուն օգտակար են ամենաբազմազան ուղղություններով. երբ մենք համեմատում ենք կռվի մեջ այնքան համառ կռվան աքլորին ուրիշ բոլորովին խաղաղասեր ցեղերի հետ, «շարունակ ձու ածող» հավերի հետ, որոնք հրաժարվում են թուխս նստելուց, և փոքրիկ նրբակազմ բանտամների հետ. երբ մենք համեմատում ենք միմյանց հետ դաշտային, բանջարանոցային պտղատու և ծաղկատու բույսերի լեգիոնները, որոնք այնքան օգտակար են մարդուն տարվա զանազան եղանակներին զանազան տեսակետներից կամ զվարճացնում են նրա հայացքը,— մենք ստիպված ենք, և կարծում եմ, դրանց բացատրությունները որոնել մի քիչ ավելի խորը, քան փոփոխականության պարզ փաստն է։ Մենք չենք կարող ընդունել, որ բոլոր ցեղերը հանկարծակի կերպով առաջացել են այդպես կատարյալ ու օգտակար, ինչպես որ մենք այժմ նրանց տեսնում ենք, մանավանդ որ շատ դեպքերում մենք հաստատապես գիտենք, որ նրանց պատմությունը այդպես չի եղել։ Բացատրության բանալին գտնվում է նրանում, որ մարդը կարող է ընտրության միջոցով կուտակել փոփոխությունը. բնությունը մատակարարում է հաջորդական փոփոխություններ, մարդը գումարում է դրանք որոշ, իրեն օգտակար ուղղություններով։ Այդ իմաստով կարելի է ասել, որ նա ինքն է ստեղծել իրեն համար օգտակար ցեղերը։
Ընտրության այդ սկզբունքի հզորությունը հիպոթեզ չէ։ Կասկածի ենթակա չէ, որ մեր աչքի ընկնող անասնաբույծներից շատերը նույնիսկ մի մարդկային կյանքի ընթացքում զգալի չափով փոփոխել են տավարի և ոչխարների ցեղերը։ Որպեսզի մենք մեզ լրիվ հաշիվ տանք, թե նրանք ինչ են ձեռք բերել, դրա համար անհրաժեշտ է կարդալ այդ խնդրին նվիրված մի քանի աշխատություններ և տեսնել այն կենդանիներին, որոնց մասին խոսվում է։ Անասնաբույծները սովորաբար կենդանու կազմվածքի մասին խոսում են, ինչպես մի ինչ-որ պլաստիկ բանի մասին, որը նրանք կարող են կերտել ըստ իրենց ցանկության։ Եթե այստեղ դրա համար տեղ բավականացներ, ես կարող էի բերել քաղվածքներ այդ իմաստով ամենահեղինակավոր գրողներից։ Յուատը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 50] որ հավանորեն ավելի գիտակ է գյուղատնտեսության այդ բնագավառում, քան որևէ մեկը և կենդանիներից լավ հասկացող է, խոսում է ընտրության սկզբունքի մասին որպես մի միջոցի, որը «ոչ միայն ազդում է իր հոտի բնույթի վրա, այլև բոլորովին փոփոխում է այն։ Դա մոգական գավազան է, որի օգնությամբ նա կյանքի է կանչում ուզած ձևերը»։ Լորդ Սոմերվիլը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 51] հիշատակելով այն մասին, թե ինչ արդյունքների են հասել անասնաբույծները ոչխարների վերաբերմամբ, ասում է. «Նրանք կարծես թե պատի վրա կավճով գծել են բոլոր կողմերով կատարյալ մի ձև և հետո կյանք են տվել նրան»։ Սաքսոնիայում մերինոսների ընտրության սկզբունքի կարևորությունը այն աստիճանի հանրաճանաչ է, որ կան մարդիկ, որոնք այդ զբաղմունքը դարձրել են իրենց համար արհեստ. ոչխարներին դնում են սեղանի վրա և ուսումնասիրում, ինչպես իմացողներն ուսումնասիրում են նկարը. այդ կրկնում են մինչև երեք անգամ մի քանի ամսվա ընթացքում, ըստ որում ամեն անգամ ոչխարներին նկատի են առնում և դասակարգում, այնպես որ վերջնականապես միայն ամենալավերին են ընտրում բազմացման համար։
Անգլիական անասնաբույծների ձեռք բերած արդյունքները ամենից ավելի լավ ապացուցվում են այն գներով, որ վճարում են լավ տոհմագրություն ունեցող կենդանիների համար, ինչպես նաև այն փաստով, որ նրանք արտահանվել են աշխարհի բոլոր ծայրերը։ Կատարելագործումը ոչ մի դեպքում ձեռք չի բերվում զանազան ցեղերի խաչաձևման միջոցով. լավագույն անասնաբույծները վճռականապես դեմ են արտահայտվում այդ եղանակին, գործադրելով այն միայն մերձավոր ենթացեղերի միջև։ Իսկ այն դեպքերում, երբ խաչաձևում է կիրառվում, ամենախիստ ընտրությունն ավելի ևս անհրաժեշտ է լինում, քան սովորական դեպքում։ Եթե ընտրությունը միայն ցայտուն կերպով արտահայտված տարատեսակների ջոկելն ու բազմացնելը լիներ, ապա այդ սկզբունքը այնքան ակներև կլիներ, որ հազիվ թե արժանանար ուշադրության։ Բայց նրա նշանակությունը հանգում է այն հսկայական արդյունքներին, որոնք ստացվում են անսովոր աչքի համար բոլորովին աննկատելի տարբերությունների կուտակմամբ հաջորդող սերունդներում, տարբերություններ, որոնք գոնե ես ապարդյուն կերպով փորձում էի նկատել։ Ոչ ավելի քան հագարից մեկը ունենում է աչքի և դատողության ճշտություն, որն անհրաժեշտ է աչքի ընկնող անասնաբույծ դառնալու համար։ Եթե նա օժտված է այդ հատկություններով և տարիներով ուսումնասիրել է իր առարկան, ապա իր ամբողջ կյանքը անհաղթահարելի համառությամբ նվիրելով այդ գործին, նա կարող է հասնել զգալի բարելավումների, եթե նա չունի այդ հատկություններից թեկուզ մեկը, նա հաստատապես անհաջողություն կկրի։ Քչերը կհավատան, թե ինչպիսի բնածին հատկություններ և քանի տարվա գործնական աշխատանք է անհրաժեշտ աղավնիներ բուծելու արվեստը սովորելու համար։
Նույն սկզբունքներն են կիրառում նաև պարտիզագործները. բայց այստեղ փոփոխությունները երբեմն ավելի հանկարծակի են լինում։ Ոչ ոք, իհարկե, չի մտածի, որ մեր լավագույն սորտերն ստացվել են բնական տեսակից մի ուժեղ շեղմամբ։ Մենք ունենք ապացույցներ, որ այդպես չի եղել շատ դեպքերում, որոնց մասին ճիշտ տեղեկություններ են պահպանվել։ Մի աննշան օրինակով սահմանափակվելու համար արժե նշել մեր հաղարջի շարունակ աճող մեծությունը։ Մենք զարմանալի փոփոխություններ ենք տեսնում պարտեզի բազմաթիվ ծաղիկների մեջ, երբ համեմատում ենք միայն քսան կամ երեսուն տարի առաջ պատրաստված նրանց նկարների հետ։ Երբ բույսի ցեղը լավ կայունացել է, պարտիզագործները, որոնք այդ ցեղը սերմերով են բուծում, արդեն ոչ թե ընտրում են լավագույն բույսերը, այլ սահմանափակվում են նրանով, որ մարգերից քաղհանում են «խաբեբաներին» (rogues), նրանք այդպես են անվանում իրենց ընտրած օրինակից շեղվող բոլոր բույսերին։ Կենդանիների վերաբերմամբ այդ եղանակը փաստորեն նույնպես կիրառվում է, որովհետև հազիվ թե որևէ անասնաբույծ այնքան անհոգ գտնվի, որ որպես արտադրող ընտրի իր ամենավատ կենդանիներին։
Բույսերի վերաբերմամբ ընտրության աստիճանաբար կուտակված ներգործության մեջ մեզ համոզելու համար կա մի միջոց ևս. արժե ուշադրություն դարձնել պարտեզի բույսի միևնույն տեսակի տարատեսակների ծաղիկների բազմազանության վրա, բանջարաբուծության մեջ գնահատելի տերևների, պատիճների, պալարների և այլ մասերի բազմազանության վրա՝ համեմատած նույն բույսերի ծաղիկների հետ. վերջապես մեր մրգատու բույսերի միևնույն տեսակի պտուղների բազմազանության վրա՝ համեմատած նույն տարատեսակների տերևների ու ծաղիկների հետ։ Տեսեք, թե ինչքան բազմազան են կաղամբի տերևները և ինչքան զարմանալիորեն նման են նրա ծաղիկները, թե ինչքան բազմազան են եռագույն մանուշակի ծաղիկները և ինչքան նման են տերևները, թե ինչքան խիստ կերպով տարբերվում են հաղարջի զանազան սորտերի պտուղները իրենց մեծությամբ, գույնով, ձևով և մազոտությամբ և ինչքան քիչ է տարբերությունը նրանց ծաղիկների միջև։ Դրանով ես չեմ ուզում ասել, որ որևէ տեսակետից միմյանցից տարբերվող տարատեսակները չպետք է տարբերվեն նաև այլ տեսակետներից. այդ հազիվ թե երբևիցե լինում է, ես կարծում եմ, նույնիսկ երբեք չի լինում, ես այդ ասում եմ իմ կատարած մանրազնին դիտումների հիման վրա։ Փոփոխականության հարաբերակցության (կապակցող) օրենքը, որի նշանակությունը երբեք չպետք է անտեսել, կառաջացնի որոշ տարբերություններ, բայց, որպես ընդհանուր կանոն, կասկածի ենթակա չէ, որ տերևների, ծաղիկների կամ պտուղների թույլ շեղումների երկարատև ընտրությունը սկիզբ կտա ցեղերի, որոնք միմյանցից կտարբերվեն գլխավորապես հենց այդ հատկանիշներով։
Կարելի է առարկել, որ ընտրության սկզբունքը խիստ մեթոդիկ կերպով հազիվ երեք քառորդ դարից ավելի է գործադրվում. վերջին տարիներին, իհարկե, նա իր վրա ավելի շատ ուշադրություն է գրավում. այդ խնդրին նվիրված քիչ աշխատություններ չեն հրատարակվել և դրան համապատասխանորեն ստացվել են արագ և նշանակալի արդյունքներ։ Բայց բոլորովին ճիշտ չէր լինի ենթադրել, որ ընտրության սկզբունքի կիրառումը նորագույն հայտնագործություն է։ Ես կարող իմ բերել քաղվածքներ խոր հնության շրջանին պատկանող գրվածքներից, որոնց մեջ այդ սկզբունքի նշանակությունը լիովին ճանաչվում է։ Անգլիական պատմության դեռ բիրտ բարբարոսական շրջանում ուրիշ երկրներից ընտիր կենդանիներ էին ներմուծվում, ինչպես նաև օրենքներ էին հրատարակվում, որոնք արգելում էին նրանց արտահանումը. կարգադրվում էր որոշ հասակից ցածր ձիերը ոչնչացնել, իսկ այդ եղանակը միանգամայն նմանվում է «խաբեբաների» քաղհանին, որը գործադրվում է պարտիզագործության մեջ։ Ընտրության սկզբունքը հանգամանորեն շարադրված է չինական հին հանրագիտարանում։ Նրա հիմնական կանոնների միանգամայն պարզորոշ շարադրանքը կարելի է գտնել հռոմեական մի քանի կլասիկ հեղինակների մոտ։ Գիրք ծննդոցի մեջ եղած մի քանի կտորներից կարելի է եզրակացնել, որ նույնիսկ այն վաղ ժամանակաշրջանում արդեն ուշադրություն էր դարձվում ընտանի կենդանիների գույնի վրա։ Ներկայումս վայրենիներն իրենց շների ցեղերը կատարելագործելու համար նրանց խաչաձևում են Canis սեռի վայրի տեսակների հետ. Պլինիուսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 47] վկայության հիման վրա պարզ է, որ այդ գործադրվում էր նաև իր ժամանակներում։ Հարավային Աֆրիկայի վայրենիներն ընտրում էին միանման գույնի բանող անասուններ, նույնն են անում էսկիմոսները իրենց շների նկատմամբ։ Լիվինգստոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 52] վկայում է, որ Կենտրոնական Աֆրիկայի նեգրերը, որոնք երբեք չեն շփվել եվրոպացիների հետ, բարձր են գնահատում ընտանի անասունների լավ ցեղերը։ Այս փաստերից մի քանիսն ընտրության անմիջական ապացույցներ չեն, բայց ցույց են տալիս, որ ընտանի ցեղերի բուծումը և բարելավումը իրենց վրա ուշադրություն են գրավել շատ հին ժամանակներում և այժմ գրավում են զարգացման ամենացածր աստիճանների վրա կանգնած վայրենիների ուշադրությունը։ Եվ շատ տարօրինակ կլիներ, եթե նրանք ուշադրություն չդարձնեին իրենց ցեղերի հատկությունների վրա,— չէ՞ որ լավ և վատ հատկությունների ժառանգականության փաստը այդքան ակներև է, այդքան աչքի է զարնում։
ԱՆԳԻՏԱԿՑԱԿԱՆ ԸՆՏՐՈՒԹՅՈՒՆ
Ներկայումս հռչակ վայելող անասնաբույծները միանգամայն որոշակի նպատակով աշխատում են մեթոդական ընտրության միջոցով առաջացնել նոր ենթացեղեր, որոնք իրենց հատկություններով գերազանցում են երկրում մինչև այժմ տվյալ ուղղությամբ ստացված բոլորին։ Բայց մեր տեսակետից ավելի կարևոր է ընտրության այն ձևը, որը կարելի է անվանել անգիտակցական ընտրություն և որը կայանում է նրանում, որ ամեն ոք աշխատում է ունենալ կենդանիների հնարավորին չափ լավագույն անհատներ և պահել նրանց բազմացման համար։ Այսպես, օրինակ, պոյնտեր ունենալ ցանկացող մարդը բնականաբար կմտածի, ինչքան կարող է լավագույն շներ ձեռք բերել և նրանցից ամենալավերի ձագերը կպահի, բայց նա նկատի չունի և նույնիսկ չի սպասում, որ այդ անելով կբարելավի ցեղը։ Այսուամենայնիվ, մենք իրավունք ունենք եզրակացնելու, որ նման պրոցեսը, շարունակվելով դարերի ընթացքում, կարող է փոփոխել և կատարելագործել ամեն մի ցեղ նույնպիսի ձևով, ինչպիսի ձևով, որ Բաքուելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 53] կամ Կոլլինսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 54] կիրառելով նույն պրոցեսը, բայց ավելի մեթոդական եղանակով, նույնիսկ միայն իրենց կյանքի ընթացքում զգալիորեն փոփոխեցին տավարի ձևերն ու որակը։ Դրա նման դանդաղ ու աննշմարելի փոփոխությունները չեն կարող նկատվել, եթե չեն պահպանվում չափումների արդյունքները կամ մանրազնին նկարները, որ կատարված են շատ տարիներ առաջ և կարող են ծառայել համեմատության համար։ Սակայն որոշ դեպքերում նույն ցեղի չփոփոխված կամ քիչ փոփոխված անհատները պահպանվում են ավելի քիչ քաղաքակրթված երկրներում, որտեղ ցեղը դեռ այն աստիճանի չի կատարելագործվել։ Հիմք կա ենթադրելու, որ Չարլզ թագավորի (կինգ չարլզ) սպանյոլը այսպիսի անգիտակցական ճանապարհով զգալի չափով ձևափոխվել է սկսած այդ միապետի ժամանակներից։ Միանգամայն գիտակ հեղինակություններ համոզված են, որ սետտերը ուղղակի առաջացել է սպանյոլից՝ վերջինիս դանդաղ փոփոխության ճանապարհով։ Հայտնի է, որ անգլիական պոյնտերը զգալիորեն փոփոխվել է վերջին դարում և այս անգամ փոփոխությունը գլխավորապես վերագրվում է բարակի (fox hound) հետ խաչաձևմանը. բայց մեզ այս դեպքում հետաքրքրում է այն փաստը, որ այդ փոփոխությունը իրագործվել է աստիճանաբար և անգիտակցաբար և, այնուամենայնիվ, այնքան իրական է, որ թեև ոչ մի կասկած չկա հին իսպանական պոյնտերի Իսպանիայից մեզ մոտ գալու մասին, ներկայումս, ինչպես հաղորդում է միստր Բորրոուն,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 55] ոչ մի տեղական իսպանական շուն չի նմանվում պոյնտերին։
Ընտրության և տքնաջան մարզեցման նույնպիսի պրոցեսի շնորհիվ անգլիական արշավաձիերը ինչպես արագությամբ, այնպես էլ մեծությամբ գերազանցել են իրենց արաբական նախնիներին, այնպես որ Գուդվուդի ձիարշավների կանոնները վերջիններին որոշ արտոնություն են տալիս հեծյալի կշռի տեսակետից։ Լորդ Սպենսերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 56] և ուրիշները ցույց են տվել, թե քաշի և վաղահասության իմաստով ինչքան են կատարելագործվել անասունների ժամանակակից ցեղերը Անգլիայում այն նախնական ցեղերի համեմատությամբ, որոնցից առաջացել են նրանք։ Համեմատելով հին աշխատություններում եղած վկայությունները փոստատար և թուրման աղավնիների մասին այժմյան գրության հետ Բրիտանիայում, Հնդկաստանում և Պարսկաստանում, մենք կարող ենք հետամուտ լինել այն բոլոր միջանկյալ ստադիաներին, որոնցով անցել են այդ ցեղերը, մինչև որ հասել են արդի վիճակին, որով նրանք այնպես խիստ տարբերվում են ժայռաղավնուց։
Յուատը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 50] բերում է մի սքանչելի օրինակ, որը կարելի է համարել անգիտակցական ընտրության մի դեպք այն իմաստով, որ անասնաբույծներն իրենք չէին սպասում և չէին ցանկանում իրենց ստացած արդյունքը, այսինքն՝ երկու տարբեր ցեղերի առաջացումը։ Լեյստերյան ոչխարների երկու հոտեր, որոնց պահում էին միստր Բեքլեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 57] և միստր Բերգեսսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 58] «երկուսն էլ առաջացել են միստր Բաքուելի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 53] սկզբնական ցեղից, հիսուն տարուց ավելի պահվում էին հնարավորին չափ զտարյուն վիճակում։ Ամենափոքր կասկած անգամ չի կարող լինել, որ երկու ոչխարատերերը գոնե որքան և իցե փոխեին բաքուելյան հոտի մաքուր արյունը, բայց և այնպես այդ երկու հոտերի միջև եղած տարբերությունը այնքան մեծ է, որ կարելի էր նրանց համարել երկու տարբեր տարատեսակների պատկանող հոտեր»։
Եթե նույնիսկ գոյություն ունենային այնքան տգետ վայրենիներ, որ չնկատեին իրենց ընտանի կենդանիների հատկությունների ժառանգականությունը, ապա մինչև անգամ այդ դեպքում էլ որևէ պատճառով առանձնապես օգտակար կենդանիներին նրանք խնամքով կպահպանեին սովի կամ այլ պատահարների ժամանակ, որոնց այնքան ենթակա է վայրենիների կյանքը, և այդ ընտիր կենդանիները կթողնեին, ընդհանրապես ասած, ավելի շատ սերունդ, քան նվազ կատարելության հասածները, այնպես որ այստեղ ևս կդրսևորվեր անգիտակցական ընտրության մի ձևը։ Թե ինչպես գնահատում են իրենց կենդանիներին նույնիսկ Հրո երկրի բնակիչները, կարող ենք դատել այն փաստի հիման վրա, որ սովի ժամանակ նրանք սպանում և ուտում են իրենց պառավ կանանց, շներից ավելի քիչ գնահատելով նրանց։
Բոլորովին նման պրոցես կարելի է նկատել բույսերի վերաբերմամբ. լավագույն անհատները, որոնք թեև մյուսներից այնքան չեն տարբերվում, որպեսզի կարելի լիներ նրանց առաջին անգամ երևալուց համարել որպես տարատեսակ, կարող են պատահմամբ գերադասորեն ավելի պահպանվել, քան մյուսները, և այդպիսով ցեղերի կամ տեսակների միջև տեղի ունեցող խաչաձևման մասնակցությամբ կամ առանց խաչաձևման ստացվում է աստիճանական կատարելագործման պրոցես, որն արտահայտվում է մեծության և գեղեցկության մեջ, որոնք նկատվում են, օրինակ, եռագույն մանուշակի, վարդի, պելարգոնիայի, գեորգինի և այլ բույսերի տարատեսակներում, երբ նրանց համեմատում ենք հին տարատեսակների կամ նրանց բնական նախահայրերի հետ։ Ոչ ոքի մտքով չի անցնի, որ օրինակելի գեորգինը կամ եռագույն մանուշակը կարելի է ստանալ վայրի տեսակի սերմերից։ Ոչ ոք չի սպասի տանձի սերմերից ստանալ օրինակելի հյութալի տանձ, թեև պատահմամբ այն կարող էր ստացվել այգու տանձի վայրենացած սերմնաբույսից։ Դեռ հին ժամանակներում տանձենին բուծվել է այգիներում, բայց ըստ Պլինիոսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 47] նկարագրության, շատ քիչ արժեքավոր էր։ Այգեգործական գրվածքներում ինձ հանդիպել են զարմանքի արտահայտումներ այգեգործների զարմանալի վարպետության վերաբերմամբ, որոնք կարողացել են այդքան ողորմելի նյութից ստանալ այդպիսի փայլուն արդյունքներ, սակայն այդ վարպետությունը շատ պարզ էր և ստացված վերջնական արդյունքի նկատմամբ գործադրվել է գրեթե անգիտակցաբար։ Այն արտահայտվել է նրանում, որ բուծել են հայտնի տարատեսակներից ամենալավը, ցանել են նրանից ստացված սերմերը, և եթե երևան է եկել մի քիչ ավելի լավ ձև, ընտրել են այն. նույն բանը կրկնել են յուրաքանչյուր նոր սերնդում։ Սակայն հին ժամանակների այգեգործները, որոնք բուծել են իրենց համար մատչելի տանձենիների լավագույն սորտերը, անշուշտ չէին մտածում այնպիսի սքանչելի պտուղների մասին, որոնց մենք ուտում ենք այժմ, թեև մենք այդ պտուղների համար որոշ չափով պարտական ենք այն խնամքին, որ նրանք տարել են իրենց համար մատչելի լավագույն սորտերն ընտրելու և պահպանելու ուղղությամբ։
Այդպիսի դանդաղ և անգիտակցական ճանապարհով ձեռք բերված փոփոխականության զգալի աստիճանը բացատրում է, իմ կարծիքով, այն հանրահայտ փաստը, որ շատ դեպքերում մենք ի վիճակի չենք իմանալու, և հետևապես նաև չգիտենք մեր այգիներում ու բանջարանոցներում ամենից ավելի երկար մշակվող բույսերի վայրի նախնիներին։ Եթե դարեր կամ հազարամյակներ են պահանջվել նրա համար, որպեսզի մեր բույսերի մեծամասնությունը հասներ օգտակարության այն աստիճանին, որով նրանք աչքի են ընկնում այժմ, ապա մեզ հասկանալի է դառնում, թե ինչու ոչ Ավստրալիան, ոչ Բարեհուսո հրվանդանը, ոչ էլ որևէ այլ երկիր, որտեղ ապրում են բոլորովին ոչքաղաքակիրթ ցեղեր, մեզ չեն տվել ոչ մի բույս, որը արժենար մշակել։ Դրա պատճառը ոչ թե նրանումն է, որ տեսակներով այդքան հարուստ այդ երկրները ինչ-որ տարօրինակ պատահականությամբ չեն ունեցել այնպիսի բույսեր, որոնք կարող էին տալ օգտակար տեսակներ, այլ նրանումն է, որ տևական անընդհատ ընտրության միջոցով տեղական բույսերը չեն հասցվել այն աստիճան կատարելության, որին հասցվել են բույսերը հին քաղաքակրթություն ունեցող երկրներում։
Ոչքաղաքակիրթ մարդու կողմից բուծվող ընտանի կենդանիների վերաբերմամբ աչքաթող չպետք է անել այն, որ նրանք համարյա միշտ, գոնե տարվա որոշ եղանակներին, ստիպված են իրենց սննդի համար պայքար մղել։ Իսկ երկու միմյանցից զանազան պայմաններով տարբերվող երկրներում միևնույն տեսակի անհատները, որոնք միմյանցից թեթևակի տարբերվում են կառուցվածքով և կազմվածքով, կարող են ավելի հաջողությամբ զարգանալ այս կամ այն երկրում և այդպիսով «բնական ընտրության» պրոցեսի շնորհիվ, ինչպես ավելի մանրամասնորեն կբացատրվի ստորև, սկիզբ կտան երկու ենթացեղերի։ Այդ հանգամանքը թերևս մասամբ բացատրում է շատ հեղինակների կողմից նկատված այն փաստը, որ վայրենիների բուծած տարատեսակները ավելի նման են բնական տեսակներին, քան քաղաքակրթված երկրներում հանդիպող տարատեսակները։
Մարդու կողմից գործադրվող ընտրության դերի կարևորության վերաբերմամբ շարադրված տեսակետի հիման վրա միանգամայն պարզ է դառնում, թե ինչու մեր ընտանի ցեղերն իրենց կառուցվածքում և սովորություններում կրում են մարդու պահանջմունքների և քմահաճույքի դրոշմը։ Մեզ հասկանալի է դառնում մեր ընտանի ցեղերի մեջ այնքան հաճախ դրսևորվող միանգամայն աննորմալ բնույթը, ինչպես նաև ներքին մասերի և օրգանների համեմատաբար չնչին փոփոխության դեպքում արտաքին հատկանիշների զգալի աստիճանի փոփոխությունը։ Մարդը գրեթե ի վիճակի չէ ընտրելու կամ միայն մեծ դժվարությամբ կարող է առաջ բերել կառուցվածքում այնպիսի շեղումներ, որոնք արտաքուստ չեն արտահայտվում որևէ բանով. ասենք՝ առհասարակ միայն հազվագյուտ դեպքերում նա մտածում է ներքին կառուցվածքի մասին։ Նա կարող է ընտրության միջոցով ազդել միայն բնության ընձեռած թույլ շեղումների վրա։ Երբեք, իհարկե, նրա մտքով էլ չէր անցնի ստանալ սիրամարգանման աղավնի, մինչև որ նա չտեսներ աննորմալ զարգացած պոչով թռչուն, կամ ստանալ փքուն աղավնի, եթե նրան չհանդիպեր խիստ զարգացած խպիպ ունեցող թռչուն, և ինչքան արտասովոր և աննորմալ էին այդ առանձնահատկությունները, նրանք այնքան ավելի շուտ կարող էին բևեռել իրենց վրա մարդու ուշադրությունը։ «Սիրամարգանման աղավնի ստանալու փորձը» և այլ նման արտահայտությունները, իմ կարծիքով, բոլորովին սխալ են։ Առաջին անգամ մի քիչ ավելի լայն պոչ ունեցող աղավնի ընտրող մարդը անշուշտ չէր էլ կարող երազել, թե ինչի նման կլինեն այդ թռչունի սերունդները մասամբ գիտակցական, մասամբ մեթոդական տևական ընտրության շնորհիվ։ Կարող է պատահել, որ բոլոր սիրամարգանման աղավնիների նախածնողը ունեցել է ընդամենը տասնչորս թեթևակի լռված պոչափետուրներ, ինչպես այժմյան յավայան սիրամարգանման աղավնու կամ զանազան այլ ցեղերի մի քանի անհատների մոտ, որոնց մեջ պատահում են մինչև տասնյոթ պոչափետուր ունեցողներ։ Գուցե առաջին փքունը իր խպիպը ավելի չի ուռցրել, քան այդ անում է ճայ-աղավնին (Turbit) այժմ, որի վրա աղավնիներ ունեցողները ուշադրություն չեն էլ դարձնում, որովհետև դա այդ ցեղերի բնորոշ առանձնահատկությունը չէ։
Եվ թող չկարծեն, թե շեղումները պետք է շատ խիստ լինեն, որպեսզի սիրողի ուշադրությունը իրենց վրա գրավեն, նա նկատում է գրեթե աննշմարելի չնչին տարբերությունները, իսկ մարդու բնավորությունն է՝ գնահատել ամեն մի, թեկուզ և աննշան, նորություն, եթե այն պատկանում է իրեն։ Եվ չպետք է աննշան անհատական շեղումների մինչ այժմ տրվող նշանակության մասին դատել ըստ այն պահանջների, որոնք առաջադրվում են այժմ, երբ գոյություն ունեն մի քանի միանգամայն կայունացած ցեղեր։ Հայտնի է, որ հիմա էլ թեթև շեղումները պատահականորեն երևան են գալիս աղավնիների կայունացած տիպերում, բայց նրանք դեն են շպրտվում որպես սխալներ կամ տվյալ ցեղի համար ընդունված տիպարից խոտորումներ։ Սովորական սագը, ինչպես հայտնի է, ոչ մի ցայտուն տարատեսակներ չունի, դրա հետևանքով տուլուզական սագը և մեր սովորական սագը, որոնք իրարից տարբերվում են միայն գույնով, այդ ամենաանկայուն հատկանիշով, հանդես են գալիս մեր ընտանի թռչունների ցուցահանդեսներում որպես ինքնուրույն ցեղեր։
Հենց նոր զարգացված հայացքները բացատրում են վաղուց նկատված մի փաստ, այն է, որ մեզ գրեթե ոչինչ հայտնի չէ մեր ընտանի ցեղերի ծագման կամ պատմության մասին։ Իրականում ցեղը, ինչպես որևէ լեզվի մի բարբառ, հազիվ թե երբևիցե ունենում է իր որոշակի սկիզբը կամ ծագման մոմենտը։ Մարդը պահում է և բուծում է թույլ շեղումներ ցուցաբերող անհատների սերունդը կամ առանձնապես հոգ է տանում լավագույն արտադրողների բնակության մասին և այսպիսով կատարելագործում է իր կենդանիներին, որոնք այնուհետև տարածվում են մոտակա հարևան տեղերում։ Բայց նրանք դեռ դժվար թե առանձին անուն կունենան, և որովհետև նրանց դեռ բավարար չափով չեն էլ գնահատի, ուստի նրանց պատմությանը ուշադրություն չեն դարձնի։ Նույն դանդաղ և աստիճանական պրոցեսի միջոցով էլ ավելի կատարելագործվելով, նրանք է՛լ ավելի լայն տարածում կստանան, կճանաչվեն որպես ինքնուրույն և արժեքավոր մի բան և այն ժամանակ միայն, կարող է պատահել, կստանան տեղական որևէ անուն։ Կիսաքաղաքակրթված երկրներում, որտեղ հաղորդակցության և փոխանակության միջոցները շատ աննշան են, նոր ենթացեղի տարածման պրոցեսը պետք է չափազանց դանդաղ կատարվի։ Հենց որ օգտակար առանձնահատկությունները կգնահատվեն ըստ արժանիքների, երևան կգա իմ անվանած անգիտակցական ընտրության ներգործությունը և նա տեղի կունենա որոշ դարաշրջանում ավելի արագ, քան մյուսում՝ նայած այդ ցեղի վերաբերմամբ եղած մոդայի աճմանը կամ անկմանը, մի տեղում ավելի ուժեղ, քան մյուսում՝ նայած բնակիչների կուլտուրականության աստիճանին, բայց դանդաղ ու անփոփոխ կերպով միևնույն ուղղությամբ կուտակելով ցեղի բնորոշ հատկանիշները, ինչպիսիք էլ որ նրանք լինեն։ Բայց շատ քիչ հավանական է, որ որևիցե տեղեկություններ կպահպանվեին փոփոխության այդպիսի դանդաղ, տատանվող և աննկատելի պրոցեսի մասին։
ԸՆՏՐՈՒԹՅԱՆ ԿԻՐԱՌՄԱՆ ԳՈՐԾՈՒՄ ՄԱՐԴՈՒՆ ՆՊԱՍՏՈՂ ՀԱՆԳԱՄԱՆՔՆԵՐԸ
Այժմ մի քանի խոսք կասեմ այն հանգամանքների մասին, որոնք նպաստում են մարդու կողմից ընտրություն կիրառելուն կամ ազդում են հակառակ իմաստով։ Փոփոխականության զգալի աստիճանը ակներևորեն բարենպաստ է, որովհետև առատ նյութ է ընձեռում ընտրության համար, բայց չպետք է կարծել, որ մանրազնին կերպով ընտրված միայն անհատական տարբերությունները բավական չեն համարյա ամեն մի ցանկալի ուղղությամբ և մեծ չափերով կուտակման համար։ Բայց քանի որ մարդուն ակնհայտ օգտակար կամ հաճելի փոփոխությունները կարող են միայն պատահականորեն առաջանալ, ուստի հասկանալի է, որ նրանց երևան գալու հավանականությունը պետք է աճի պահվող անհատների թվի հետ միասին։ Այստեղից հետևում է, որ թիվը մեծ չափով ազդում է հաջողության վրա։ Այդ հիման վրա Մարշալը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 59] Յորկշիրի մի քանի մասերում պահվող ոչխարների մասին հայտնում է հետևյալ կարծիքը. «Նրանք երբեք չեն կատարելագործվի, որովհետև պատկանում են աղքատ մարդկանց և պահվում են փոքր խմբերով»։ Մյուս կողմից, առևտրական այգեգործությամբ պարապողները, որոնք միևնույն բույսից մշակում են մեծ քանակությամբ, սիրողներից ավելի հաջող կերպով են ստանում նոր ու արժեքավոր տարատեսակներ։ Մեծ քանակությամբ կենդանիներ և բույսեր պահելը կամ մշակելը հնարավոր է միայն այն պայմաններում, որոնք բարենպաստ են նրանց բազմացման համար։ Երբ անհատները սակավաթիվ են, նրանք բոլորը, ինչպես էլ որ լինեն նրանց հատկությունները, կպահվեն բազմացման համար, իսկ այդ փաստորեն կվերացնի ընտրությունը։ Սակայն ամենայն հավանականությամբ հաջողության գլխավոր պայմանը պետք է համարել մարդու համար կենդանու կամ բույսի այնքան բարձր արժեքավորությունը, որպեսզի նա ամենամանրազնին ուշադրությունը դարձներ նրանց կառուցվածքում կամ հատկություններում կատարվող ամենափոքր շեղումների վրա։ Առանց այդպիսի ուշադրության ոչինչ չի ստացվի։ Ես առիթ եմ ունեցել լսելու միանգամայն լուրջ արտահայտված մի դիտողություն, թե ինչ բախտավորություն, որ ելագն սկսել է փոփոխվել հենց այն ժամանակից, երբ պարտիզպաններն սկսել են ուշադրություն դարձնել նրա վրա։ Կասկած չկա, որ ելագը միշտ էլ փոփոխվել է, հենց այն ժամանակից, ինչ սկսել են նրան մշակել, բայց այդ թեթև փոփոխությունների վրա ուշադրություն չի դարձվել։ Սակայն, հենց որ պարտիզպաններն սկսեցին ընտրել ավելի խոշոր, ավելի վաղահաս կամ համեղ պտուղներ ունեցող անհատներ, սկսեցին ցանել նրանց սերմերը և ստացված սերմնաբույսերից ընտրել ամենալավերը (զանազան տեսակների միջև խաչաձևման որոշ աջակցությամբ), այդ ժամանակից էլ երևան եկան այն զարմանալի տարատեսակները, որոնք ստեղծվել են վերջին հիսնամյակում։
Կենդանիների վերաբերմամբ կարևոր դեր է կատարում խաչաձևումների վերացման հնարավորությունը,— դա համենայն դեպս շատ էական հանգամանք է այն երկրների համար, որտեղ արդեն կան շատ ցեղեր։ Այդ տեսակետից մեծ նշանակություն ունի հողամասերի ցանկապատումը։ Թափառակյաց վայրենիները կամ բաց հարթավայրի բնակիչները հազվագյուտ դեպքում են ունենում կենդանու նույն տեսակի մեկ ցեղից ավելի։ Աղավնիների կազմած զույգերը կարող են ամբողջ կյանքում մնալ, և դա մեծ հարմարություն է այդ թռչունը պահել սիրողների համար, որովհետև մի քանի ցեղ կարելի է բարելավել միաժամանակ, պահպանելով նրանց զտարյուն վիճակում մի թռչնանոցի մեջ։ Ես կարող եմ ավելացնել նաև այն, որ աղավնիները շատ արագ են բազմանում և նրանց թիվը արագորեն ավելանում է, իսկ չբավարարող թռչունները հեշտությամբ վերացվում են, որովհետև նրանց գործ են ածում սննդի համար։ Մյուս կողմից, կատուները իրենց գիշերային թափառական կենսակերպի հետևանքով հեշտությամբ չեն ենթարկվում զույգընտրության և, թեպետ բարձր են գնահատվում կանանց ու երեխաների կողմից, շատ հազվագյուտ դեպքում են տալիս որոշակի ցեղեր, իսկ եթե երբեմն հանդիպում են այդպիսի ցեղեր, ապա դրանք գրեթե միշտ էլ ուրիշ երկրներից են բերված։ Թեև ես չեմ կասկածում այն բանում, որ որոշ ընտանի կենդանիներ ավելի քիչ են փոփոխվում, քան մյուսները, այնուամենայնիվ կատվի, էշի, սիրամարգի, սագի և այլոց որոշակի ցեղերի հազվադեպությունը կամ լիակատար բացակայությունը կարող են վերագրվել ընտրության բացակայությանը. կատուներինը՝ արտադրողների զույգընտրության դժվարության հետևանքով. էշերինը այն պատճառով, որ նրանց սովորաբար միայն աղքատ մարդիկ են պահում փոքր թվով և նրանց կատարելագործման վրա ուշադրություն չեն դարձնում, և միայն վերջին ժամանակներս Իսպանիայում և Միացյալ Նահանգներում այդ կենդանին զարմանալիորեն փոփոխվել ու կատարելագործվել է խնամքով ընտրության միջոցով. սիրամարգերինը՝ այն հոգսերի հետևանքով, որոնց հետ կապված է նրանց բուծումը, ինչպես նաև պահվող անհատների սովորաբար սահմանափակ քանակության հետևանքով. սագերինը այն պատճառով, որ նրանք գնահատվում են իրենց տված փետուրների և սննդամթեր քի համար միայն, իսկ գլխավորապես նաև այն պատճառով, որ զանազան ցեղեր բուծելը բավականություն չի պատճառել. բայց սագը իր ընտանի պահվածքի սովորական պայմաններում, ըստ երևույթին, աչքի է ընկնում իր կազմվածքի բացառիկ անշարժությամբ, թեև նա էլ աննշան չափով փոփոխական է, ինչպես ես այդ նշել եմ մի այլ տեղում։
Մի քանի հեղինակներ այն կարծիքն են հայտնել, որ մեր ընտանի ցեղերը փոփոխականության սահմանին շատ շուտով կհասնեն և այնուհետև նրանք արդեն այլևս չեն ենթարկվի փոփոխության։ Բայց մի քիչ անմտածված կլիներ պնդել, թև որևէ դեպքում այդ սահմանին հասել ենք, որովհետև գրեթե մեր բոլոր ընտանի կենդանիներն ու մշակովի բույսերը կատարելագործվել են ամենավերջին ժամանակներս, իսկ այդ ակնհայտորեն վկայում է նրանց փոփոխականության մասին։ Անմտածված կլիներ նաև պնդել, թե իրենց որոշակի զարգացման հասած հատկանիշները ամբողջ դարեր կայունությունից հետո չեն կարողանա փոփոխվել կենսապայմանների փոփոխման դեպքում։ Բայց անշուշտ, ինչպես շատ ճիշտ նկատել է միստր Ուոլլեսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] երբևիցե այդ սահմանը պիտի հասնի։ Այսպես, օրինակ, որևէ ցամաքային կենդանու վազքի արագության սահման պիտի լինի, որը որոշվում է հաղթահարելի շփմամբ, տեղափոխելի մարմնի կշռով և մկանների կծկման ուժով։ Բայց ներկա դեպքում մեզ հետաքրքրում է միայն այն փաստը, որ միևնույն տեսակի ընտանի տարատեսակները միմյանցից տարբերվում են համարյա այն բոլոր ուղղություններով, որոնց վրա մարդը ուշադրություն է դարձրել կամ իր ընտրության առարկան է դարձրել ավելի մեծ չափով, քան նույն սեռի բնական տեսակները։ Իսիդոր Ժոֆֆրուա Սենտ-Իլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 16] այդ ապացուցել է մեծության վերաբերմամբ, և հավանորեն նույնը կարելի է ասել գույնի և, թերևս, մազի երկարության վերաբերմամբ։ Ինչ վերաբերում է արագությանը, որ կախված է միասին վերցրած շատ հատկություններից, ապա Էկլիպսը անկասկած ավելի արագավազ էր, նույնպես էլ մեր ծանրաքաշները անհամեմատ ուժեղ են, քան նույն սեռի որևէ բնական տեսակ։ Նույնն է նաև բույսերի վերաբերմամբ. բակլայի և եգիպտացորենի զանազան տարատեսակների սերմերը իրենց մեծությամբ հավանորեն անհամեմատ ավելի շատ են տարբերվում իրարից, քան այդ երկու ընտանիքներից որևէ սեռի տեսակների սերմերը։ Նույնը ճիշտ է նաև սալորի զանազան տարատեսակների վերաբերմամբ և առավել ևս սեխի ու համանման շատ դեպքերի վերաբերմամբ։
Ամփոփենք ընտանեցրած կենդանիների ցեղերի և մշակովի բույսերի ծագման վերաբերյալ արդյունքները։ Փոփոխված կիսապայմանները չափազանց կարևոր են, որովհետև առաջ են բերում փոփոխականություն, ազդելով կազմվածքի վրա կամ անմիջականորեն, կամ անուղղակիորեն վերարտադրողական սիստեմի միջոցով։ Անհավանական է, որ փոփոխականությունը անխուսափելիորեն հատուկ է օրգանիզմներին ամեն տեսակի պայմաններում։ Ժառանգականության և նախկին հատկանիշներին վերադառնալու ձգտման ավելի մեծ կամ փոքր ուժը որոշում է փոփոխությունների կայունությունը, ամրությունը։ Փոփոխականությունը ղեկավարվում է շատ անհայտ օրենքներով, որոնցից հարաբերակցական (կապակցական) աճը ամենակարևորն է հանդիսանում։ Երևույթի որոշ մասը, բայց մենք չգիտենք թե որքանը, կարելի է վերագրել կենսապայմանների որոշակի ներգործությանը։ Որոշ, գուցե և զգալի ազդեցություն էլ կարելի է վերագրել օրգանների վարժեցմանը և ոչ վարժեցմանը։ Այսպիսով՝ վերջնական արդյունքը չափազանց բարդ է լինում։ Որոշ դեպքերում սկզբնական զանազան տեսակների խաչաձևումը, ըստ երևույթին, կարևոր դեր է կատարել մեր ընտանի ցեղերի առաջացման գործում։ Եթե տվյալ երկրում արդեն առաջացել են մի քանի ցեղեր, նրանց պատահական, ժամանակ առ ժամանակ կրկնվող խաչաձևումը, կապված ընտրության հետ, անկասկած զգալի չափով նպաստել է նոր ենթացեղերի առաջանալուն. սակայն խաչաձևման նշանակությունը զգալիորեն չափազանցվել է ինչպես կենդանիների, այնպես էլ սերմերով բուծվող բույսերի վերաբերմամբ։ Այն բույսերի վերաբերմամբ, որոնք ժամանակ առ ժամանակ բուծվում են մատներով կամ պատվաստով, խաչաձևման նշանակությունը հսկայական է, որովհետև մշակողը այդ դեպքում կարող է ուշադրություն չդարձնել հիբրիդների և խառնացեղերի ծայրահեղ փոփոխականության և հիբրիդների անպտղության վրա, բայց սերմերով չբազմացող բույսերը մեզ համար հետաքրքրություն չեն ներկայացնում, որովհետև նրանց գոյությունը միայն ժամանակավոր է։ Սակայն փոփոխության ամենակարևոր, գերակշռող պայմանը, ըստ երևույթին, եղել է ընտրության կուտակվող ներգործությունը, որը կիրառվել է մեթոդիկորեն ու արագ կամ անգիտակցական ու դանդաղ, բայց դրա փոխարեն ավելի ներգործող արդյունքներով։
ԳԼՈՒԽ II։ ՓՈՓՈԽԱԿԱՆՈՒԹՅՈՒՆԸ ԲՆԱԿԱՆ ՎԻՃԱԿՈՒՄ
ՓՈՓՈԽԱԿԱՆՈՒԹՅՈՒՆ։ ԱՆՀԱՏԱԿԱՆ ՏԱՐԲԵՐՈՒԹՅՈՒՆՆԵՐ։— ԿԱՍԿԱԾԵԼԻ ՏԵՍԱԿՆԵՐ։— ԼԱՅՆՈՐԵՆ ՏԱՐԱԲՆԱԿՎԱԾ, ՏԱՐԱԾՎԱԾ ԵՎ ՍՈՎՈՐԱԿԱՆ ՏԵՍԱԿՆԵՐՆ ԱՄԵՆԻՑ ԱՎԵԼԻ ՓՈՓՈԽԱԿԱՆ ԵՆ։— ՅՈՒՐԱՔԱՆՉՅՈՒՐ ԵՐԿՐՈՒՄ ՄԵԾ ՍԵՌԵՐԻ ՏԵՍԱԿՆԵՐԸ ԱՎԵԼԻ ՓՈՓՈԽԱԿԱՆ ԵՆ, ՔԱՆ ՓՈՔՐ ՍԵՌԵՐԻ ՏԵՍԱԿՆԵՐԸ։— ՄԵԾ ՍԵՌԵՐԻ ՇԱՏ ՏԵՍԱԿՆԵՐ ՆՄԱՆՎՈՒՄ ԵՆ ՏԱՐԱՏԵՍԱԿՆԵՐԻՆ ԱՅՆ ԲԱՆՈՎ, ՈՐ ՆԵՐԿԱՅԱՑՆՈՒՄ ԵՆ ՇԱՏ ՄՈՏԻԿ, ԲԱՅՑ ՈՉ ՄԻԱՆՄԱՆ ԱԶԳԱԿՑՈՒԹՅՈՒՆ ՄԻՄՅԱՆՑ ՀԵՏ ԵՎ ՈՒՆԵՆ ՍԱՀՄԱՆԱՓԱԿ ՏԱՐԱԾՈՒՄ։— ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ։
Նախքան նախորդ գլխում մշակված ընդհանուր հիմունքները բնական վիճակում գտնվող օրգանիզմների վերաբերմամբ կիրառելը, մենք պետք է դեռ համառոտակի քննարկենք այն հարցը, թե նրանք արդյոք ենթակա՞ են փոփոխականության։ Այդ թեմայի պատշաճ շարադրման համար կպահանջվեր բերել չոր փաստերի մի երկար ցանկ, բայց ես այդ կթողնեմ մի այլ, հետագա աշխատության համար։ Այստեղ ես չեմ քննարկի նաև այն տարբեր սահմանումները, որ առաջարկվել են «տեսակ» տերմինի համար։ Ոչ մի սահմանում չի բավարարել բոլոր բնախույզներին. բայց յուրաքանչյուր բնախույզ աղոտ կերպով ըմբռնում է, թե նա տեսակ ասելով ինչ է հասկանում։ Ընդհանրապես այդ տերմինի տակ հասկացվում է առանձին արարչագործական ակտի մի անհայտ տարր։ «Տարատեսակ» տերմինը նույնպես դժվարությամբ է ենթարկվում սահմանման, բայց այստեղ գրեթե միշտ հասկացվում է ծագման ընդհանուր լինելը, թեև այդ միայն շատ հազվադեպ է ապացուցվում։ Մենք ունենք նաև, այսպես կոչված, այլանդակություններ, բայց նրանք աստիճանաբար անցնում են տարատեսակների։ Ես կարծում եմ, այլանդակություն համարում են կառուցվածքում մի զգալի շեղում, որը սովորաբար վնասակար է կամ անօգուտ տեսակի համար։ Մի քանի հեղինակներ գործածում են «վարիացիա» բառը որպես տեխնիկական տերմին և դրանով հասկանում այնպիսի շեղում, որը առաջանում է կյանքի ֆիզիկական պայմանների ազդեցությունից. այդ իմաստով «վարիացիաները» ժառանգական չեն համարվում, բայց ո՞վ կերաշխավորի, որ Բալթիկ ծովի անուշահամ ջրերում խեցիների գաճաճ ձևերը կամ ալպիական բույսերի նույնպիսի ձևերը, կամ հյուսիսային կենդանիների թանձր մորթին որոշ դեպքերում չեն ժառանգվի գոնե մի քանի սերունդներում։ Իսկ այդ դեպքում, ես կարծում եմ, այդ ձևերը կանվանվեին տարատեսակներ։
Կասկածելի է, որ կառուցվածքում այդպիսի հանկարծական և մեծ շեղումները, որոնք երբեմն նկատվում են մեր ընտանի ցեղերի, մանավանդ բույսերի մեջ, կարողանային մշտապես բազմանալ բնական վիճակում։ Օրգանական էակի համարյա յուրաքանչյուր մաս այնքան հիանալի կերպով է հարմարված կենսապայմանների ամբողջությանը, որ նրա հանկարծական առաջանալը այդքան կատարյալ ձևով նույնքան անհավանական է թվում, որքան անհավանական այն ենթադրությունը, թե մարդը միանգամից հնարել է ամենակատարյալ ձևի որևէ բարդ մեխանիզմ։ Ընտանի կենդանիների մեջ երբեմն առաջանում են այլանդակություններ, որոնք նմանվում են իրենց հետ բոլորովին նմանություն չունեցող այլ կենդանիների նորմալ կառուցվածքին։ Այսպես, երբեմն ծնվում են խոզեր, որոնք ունենում են կնճիթի նման մի բան, և եթե նույն սեռի որևէ բնական տեսակ ունենար կնճիթ, ապա կարելի էր եզրակացնել, որ այդ կնճիթը սկզբնապես երևացել է որպես այլանդակություն, բայց մինչև այժմ, չնայած իմ մանրազնին որոնումներին, ինձ դեռ չի հաջողվել գտնել այնպիսի այլանդակությունների առաջացման մի դեպք, որոնք նման են մերձավոր ձևերի նորմալ կառուցվածքին, մինչդեռ միայն այդպիսի դեպքերը կարող էին ուղղակի առնչություն ունենալ քննարկվող հարցի հետ։ Եթե նման այլանդակությունները բնական վիճակում առաջանային էլ և նույնիսկ կարողանային վերարտադրվել (որը ամենևին էլ միշտ չի պատահում), ապա քանի որ նրանք հազվագյուտ և բոլորովին մեկուսացած դեպքեր են, նրանց պահպանումը հնարավոր կլիներ միայն բացառապես բարենպաստ հանգամանքներում։ Բացի դրանից, առաջին և հաջորդ սերունդներում նրանք կխաչաձևվեին սովորական ձևի հետ, և նրանց աննորմալ բնույթը համարյա անխուսափելիորեն կչքանար։ Սակայն հաջորդ գլուխներից մեկում ես կվերադառնամ պատահական եզակի փոփոխության պահպանման և ամրացման փաստերի քննարկությանը։
ԱՆՀԱՏԱԿԱՆ ՏԱՐԲԵՐՈՒԹՅՈՒՆՆԵՐ
Այն բազմաթիվ և աննշան տարբերությունները, որոնք երևան են գալիս անկասկած կամ հավանորեն ընդհանուր ծնողներից առաջացած սերնդում և դիտվում են միևնույն սահմանափակ վայրում ապրող ու միևնույն տեսակին պատկանող անհատների մոտ, կարող են անվանվել անհատական։ Ոչ ոք, իհարկե, չի կարող պնդել, թե միևնույն տեսակի բոլոր անհատները ձուլված են, կարծես թե, մի կաղապարով։ Այդ անհատական տարբերությունները մեզ համար չափազանց կարևոր են, որովհետև նրանք հաճախ ժառանգական են, որի մեջ անշուշտ ամեն ոք առիթ է ունեցել համոզվելու. նրանք բնական ընտրությանը նյութ են ընձեռում ներգործելու և կուտակելու համար, այնպես, ինչպես որ մարդը իր ընտանի արտադրանքներում կուտակում է անհատական տարբերությունները ուզած ուղղությամբ։ Այդ անհատական տարբերությունները սովորաբար վերաբերում են այն մասերին, որոնք բնախույզների կողմից էական չեն համարվում. բայց ես կարող էի բերել փաստերի մի երկար շարան ապացուցելու համար այն, որ այն մասերը, որոնք պետք է էական համարել, միևնույնն է ֆիզիոլոգիական կամ կարգաբանական տեսակետից, նույնպես երբեմն փոփոխվում են միևնույն տեսակի անհատների մոտ։ Ես համոզված եմ, որ ամենափորձված բնախույզը կզարմանար կառուցվածքի նույնիսկ ամենաէական մասերի փոփոխականության դեպքերի բազմության վրա, դեպքեր, որոնք հաստատված են լավ հեղինակությունների կողմից և որոնք ինձ հաջողվել է հավաքել երկար տարիների ընթացքում։ Չպետք է մոռանալ, որ սիստեմատիկայի մասնագետները առանձին հաճությամբ չեն դիմավորում էական հատկանիշներում երևացած փոփոխականության օրինակներին և որ քիչ մարդիկ կգտնվեն, որոնք մանրազնին կերպով կուսումնասիրեն և կհամեմատեն ներքին և էական օրգանները միևնույն տեսակի բազմաթիվ անհատների վրա։ Ոչ ոք, իհարկե, չէր սպասում, որ միջատի գլխավոր նյարդերի ճյուղավորումը կենտրոնական մեծ հանգույցից հենց դուրս գալու տեղում կարող էր փոփոխություններ տալ միևնույն տեսակի ներկայացուցիչների մոտ։ Կարելի էր ենթադրել, որ այդ կարգի տարբերությունները կատարվում են միայն դանդաղորեն և աստիճանաբար, բայց և այնպես սըր. Դ. Լեբբոկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 60] Coccus-ի մոտ այդ գլխավոր նյարդերի փոփոխականությունը ցույց է տվել այնպիսի չափերով, որոնք նմանվում են ծառերի ճյուղավորության պատահական ձևերին։ Կարելի է նաև ավելացնել, որ այդ փիլիսոփա բնախույզը ցույց է տվել, որ մի քանի միջատների թրթուրների մկանները ևս բնավ աչքի չեն ընկնում միաձևությամբ։ Հեղինակները երբեմն պտտվում են սխալ շրջանում, պնդելով՝ թե էական օրգանները երբեք չեն փոփոխվում որովհետև իրենք էական են համարում (ինչպես մի քանի բնախույզեր ազնվաբար խոստովանում են) այն օրգանները, որոնք չեն փոփոխվում, կանգնելով նման տեսակետի վրա, իհարկե անհնարին է գտնել էական մասի փոփոխության որևէ դեպք, բայց ամեն մի այլ տեսակետից մոտենալով՝ դրա համար շատ օրինակներ կգտնվեն։
Կա մի հանգամանք, որը կապի մեջ է գտնվում անհատական տարբերությունների հետ և չափազանց առեղծվածային է. ես նկատի ունեմ, այսպես կոչված, «պոլիմորֆ» սեռերի կամ «պրոտեյներ» սեռերի գոյությունը, որոնց մեջ տեսակները անսահման չափերով փոփոխական են։ Հազիվ թե գտնվեն գեթ երկու բնախույզներ, որոնք այդ ձևերից շատերի վերաբերմամբ կարողանային համաձայնության գալ, թե արդյոք դրանք պետք է համարել տեսակնե՞ր, թե տարատեսակներ։ Այդպիսի ձևերի թվում կարելի է դասել բույսերից Rubus-ը, Rosa-ն և Hieracium-ը և միջատների ու ձեռնաոտանի կակղամորթների (Brachiopoda) մի քանի սեռերը։ Պոլիմորֆ սեռերի մեծամասնության մեջ որոշ տեսակներ ունեն կայուն և որոշակի հատկանիշներ։ Այն սեռերը, որոնք պոլիմորֆ են մի երկրում, փոքրիկ բացառությամբ պոլիմորֆ են նաև այլ երկրներում. դատելով ըստ Brachiopoda-ի խեցիների՝ նույնը վերաբերում է նաև նախորդ դարաշրջանների բնակչությանը։ Այս փաստերը չափազանց առեղծվածային են, որովհետև հավանորեն ստիպում են ենթադրելու, որ այդպիսի փոփոխականությունը կախված չէ գոյության պայմաններից։ Ես համամիտ եմ ենթադրելու, որ այլ պոլիմորֆ սեռերից գոնե մի քանիսում մենք ունենք այնպիսի փոփոխականության օրինակներ, որը տեսակի համար ոչ օգտակար է և ոչ էլ վնասակար և այդ պատճառով բնական ընտրության կողմից աջակցություն չի ստացել և չի ամրացվել, ինչպես այդ կպարզվի հետագա շարադրանքից։
Միևնույն տեսակին պատկանող անհատները, ինչպես հայտնի է ամենքին, հաճախ փոփոխականությունից անկախ մեծ տարբերություններ են ունենում իրենց կառուցվածքում, ինչպես օրինակ, զանազան կենդանիների երկու սեռերի մոտ, միջատների ամուլ էգերի և աշխատավորների երկու կամ երեք կաստաների մոտ և ցածրակարգ կենդանիներից շատերի մոտ անզարգացած կամ թրթուրային վիճակում։ Դիմորֆիզմի և տրիմորֆիզմի դեպքեր հայտնի են նաև կենդանիների և բույսերի մոտ։ Այսպես, օրինակ, միստր Ուոլլեսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] վերջերս ուշադրություն է դարձրել այդ երևույթի վրա և ցույց է տվել, որ Մալայան արշիպելագի թիթեռների որոշ տեսակների իգական անհատները անփոփոխ կանոնավորությամբ հանդես են գալիս երկու կամ երեք խիստ տարբերվող ձևերի տեսքով, որոնք միմյանց հետ չեն շաղկապված միջանկյալ տարատեսակներով։ Ֆրից Մյուլլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] նկարագրել է միանգամայն նման, բայց ավելի ևս զարմանալի դեպքեր բրազիլիական մի քանի խեցեմորթների արուների վերաբերմամբ. այսպես, օրինակ, տանաիսի արուները միշտ հանդիպում են երկու միմյանցից խիստ տարբերվող ձևերով. մեկն ունի ուժեղ և զանազան ձևի չանչեր, մյուսի շոշափուկները ավելի առատորեն ծածկված են հոտառական մազիկներով։ Թեև այս դեպքերի մեծամասնության մեջ բույսերի կամ կենդանիների տվյալ երկու կամ երեք ձևերն այժմ միմյանց հետ շաղկապված չեն միջանկյալ աստիճաններով, բայց և այնպես նրանք մի ժամանակ հավանորեն վերջինների միջոցով կապված են եղել։ Այսպես, օրինակ, միստր Ուոլլեսը նկարագրում է թիթեռի մի հայտնի տեսակ, որը մի կղզու վրա ներկայացնում է մի ամբողջ շարք տարատեսակներ, որոնք միջանկյալ օղակներով կապակցված են իրար հետ, իսկ այդ շարքի ծայրային անդամները չափազանց նման են Մալայան արշիպելագի այլ մասում ապրող մի մերձավոր դիմորֆ տեսակի երկու ձևերին։ Ճիշտ այդպես էլ մրջյունների մոտ աշխատավորների մի քանի կաստաներ սովորաբար բոլորովին տարբերվում են միմյանցից, բայց որոշ դեպքերում, ինչպես մենք կտեսնենք հետագայում, կաստաները իրար հետ կապված են շատ աստիճանական անցումներով։ Նույնը նկատել եմ նաև ես մի քանի դիմորֆ բույսերի մոտ։ Իհարկե, առաջին հայացքից չի կարելի չզարմանալ այն փաստի վրա, որ միևնույն էգ թիթեռը արտադրում է երեք տարբեր իգական և մեկ արական ձև, կամ թե մեկ հերմաֆրոդիտ բույսը միևնույն տուփիկում տալիս է երեք տարբեր իգական և երեք կամ նույնիսկ վեց տարբեր արական ձևի սերմեր։ Այնուամենայնիվ սրանք ամենասովորական երևույթի ավելի ցայտուն դեպքերն են, այսինքն այն երևույթի, որի դեպքում մի էգ ծնում է երկու սեռի անհատներ, որոնք երբեմն ամենազարմանալի կերպով տարբերվում են իրարից։
ԿԱՍԿԱԾԵԼԻ ՏԵՍԱԿՆԵՐ
Այն ձևերը, որոնք մեծ չափով օժտված են տեսակների հատկություններով, բայց այնքան նման են այլ ձևերի կամ այնպես սերտորեն են կապված նրանց հետ միջանկյալ օղակներով, որ բնախույզները չեն սիրում նրանց ճանաչել որպես ինքնուրույն տեսակներ, մեզ համար առանձնապես կարևորեն շատ տեսակետներից։ Մենք ունենք բոլոր հիմքերը մտածելու, որ այդ կասկածելի և միմյանց հետ սերտ կերպով կապակցված ձևերից շատերը երկար ժամանակի ընթացքում պահպանում են իրենց հատկանիշների անփոփոխությունը, նույնքան երկար ժամանակ, որքան մենք կարող ենք դատել, ինչպես այդ տեղի ունի լավ, իսկական տեսակների մոտ։ Գործնականում ամեն անգամ, երբ բնախույզը ի վիճակի է լինում որևէ երկու ձև իրար հետ կապակցելու միջանկյալ օղակներով, նա դրանցից մեկը ճանաչում է որպես մյուսի տարատեսակ, համարելով ամենից ավելի սովորականը, իսկ երբեմն էլ միայն ավելի առաջ նկարագրվածը որպես տեսակ, իսկ մյուսը՝ որպես տարատեսակ։ Բայց կան դեպքեր, որոնք ես այստեղ չեմ թվի, երբ մեծ դժվարություն է առաջանում որոշելու համար այն հարցը, թե արդյոք մենք իրավունք ունե՞նք մի որևէ ձև համարել մյուսի տարատեսակը, թեկուզ և նրանք սերտ կերպով կապակցված լինեն միջանկյալ օղակներով. նույնիսկ միշտ չէ, որ հաջողվում է այդ դժվարություններն անցնել սովորական եղանակով՝ ընդունելով միջանկյալ օղակները որպես խառնացեղեր։ Բայց բազմաթիվ դեպքերում մի ձև մյուսի տարատեսակն է համարվում ոչ այն պատճառով, որ միջանկյալ օղակները իրոք գտնված են, այլ այն պատճառով, որ դիտողը անալոգիայի հիման վրա եզրակացնում է, որ նրանք որևէ տեղ գոյություն ունեն կամ կարող էին մի ժամանակ գոյություն ունենալ,— այստեղ անշուշտ լայն ասպարեզ է բացվում կասկածների և կռահումների համար։
Այստեղից հետևում է, որ որևէ ձև որպես տեսակ կամ տարատեսակ ընդունելու հարցը որոշելիս միակ ղեկավարող սկզբունքը հանդիսանում է ճիշտ դատողություն և մեծ փորձառություն ունեցող բնախույզների կարծիքը։ Այնուամենայնիվ շատ դեպքում հարցը որոշվում է բնախույզների ձայների մեծամասնությամբ, որովհետև քիչ կճարվեն ցայտուն կերպով արտահայտված և լավ հայտնի տարատեսակներ, որոնք գոնե մի քանի իրազեկ դատավորների կողմից ընդունված չլինեին որպես ինքնուրույն տեսակներ։
Որ այդպիսի կասկածելի տարատեսակները այնքան էլ սակավաթիվ չեն, հազիվ թե կարելի է վիճարկել։ Համեմատեցեք Մեծ Բրիտանիայի, Ֆրանսիայի կամ Միացյալ Նահանգների բուսականության մասին զանազան բուսաբանների կազմած աշխատությունները և դուք կզարմանաք այն ձևերի թվի վրա, որոնք մի բուսաբան համարում է որպես լավ տեսակներ, իսկ մյուսը միայն որպես տարատեսակ։ Միստր Հ. Կ. Ուոտսոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 62] որին ես շատ երախտապարտ եմ զանազան տեսակետներից ինձ ցույց տված օգնության համար, ինձ համար նշել է բրիտանական 182 բույսեր, որոնք սովորաբար համարվում են տարատեսակներ, բայց նրանք բոլորը շատ բուսաբանների կողմից համարվել են տեսակներ։ Այդ ցուցակը կազմելիս նրա մեջ նա չէր մտցրել շատ տարատեսակներ, որոնք այնուամենայնիվ մի քանի բուսաբանների կողմից ճանաչված են որպես տեսակներ և բոլորովին բաց էր թողել մի քանի չափազանց պոլիմորֆ սեռեր։ Բաբինգտոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 63] ամենապոլիմորֆ ձևեր պարունակող սեռերում դասում է 251 տեսակ, իսկ Բենտամը՝[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 64] ընդամենը 112,— ուրեմն տարբերությունն է 139 կասկածելի ձև։ Յուրաքանչյուր ձագածնության համար զուգավորվող և զգալիորեն շարժուն կենդանիների մեջ կասկածելի ձևերը, որոնք մի կենդանաբանի կողմից համարվում են տեսակներ, իսկ մյուսի կողից տարատեսակներ, հազվադեպ են հանդիպում միևնույն երկրի սահմաններում, բայց հազվագյուտ չեն մեկը մյուսից անջատված շրջաններում։ Ինչքան շատ թռչուններ և միջատներ, որոնք հանդիպում են Հյուսիսային Ամերիկայում և Եվրոպայում ու քիչ են տարբերվում միմյանցից, մի ականավոր բնախույզի կողմից համարվել են անկասկածելի տեսակներ, իսկ մյուսի կողմից՝ տարատեսակներ, կամ, ինչպես հաճախ գերադասում են ասել, աշխարհագրական ցեղեր։ Միստր Ուոլլեսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] մի քանի արժեքավոր հետազոտություններում, որոնք նվիրված են Մեծ Մալայան արշիպելագում ապրող կենդանիներին և հատկապես թիթեռներին, նշում է, որ նրանց կարելի է դասավորել չորս անվան տակ, այն է՝ փոփոխվող ձևեր, տեղական ձևեր, աշխարհագրական ցեղեր, կամ ենթատեսակներ և իսկական փոխարինող տեսակներ։ Առաջինները կամ փոփոխվող ձևերը շատ փոփոխական են միևնույն կղզու սահմաններում։ Տեղական ձևերը համեմատաբար կայուն են և տարբեր են յուրաքանչյուր առանձին կղզու համար, բայց եթե համեմատենք բոլոր առանձին կղզիներում բնակվողները իրար հետ, ապա տարբերություններն այնքան փոքր են լինում և աստիճանական, որ անհնարին է նրանց որոշել ու նկարագրել, թեև միևնույն ժամանակ ծայրային ձևերը բավականաչափ տարբերվում են միմյանցից։ Աշխարհագրական ցեղերը, կամ ենթատեսակները, ներկայացնում են լիակատար որոշված ու մեկուսացած տեղական ձևեր, բայց որովհետև նրանք իրարից չեն տարբերվում ցայտուն և կարևոր հատկանիշներով, «ուստի հարցը լուծելու համար, թե նրանցից որոնք պետք է համարել տեսակներ և որոնք տարատեսակներ, ուրիշ չափանիշ չկա, բացի անձնական կարծիքից»։ Վերջապես, փոխարինող տեսակները յուրաքանչյուր կղզու բնության տնտեսության մեջ գրավում են նույն տեղերը, ինչ որ տեղական ձևերը, կամ ենթատեսակները, բայց քանի որ նրանց միջև եղած տարբերության աստիճանը գերազանցում է տեղական ձևերի կամ ենթատեսակների միջև եղած տարբերության աստիճանին, ուստի բնախույզները նրանց համարյա միշտ համարում են իսկական տեսակներ։ Այնուամենայնիվ, անհնարին է առաջարկել մի ճշգրիտ չափանիշ փոփոխվող ձևերը, տեղական ձևերը, ենթատեսակները և փոխարինող տեսակները միմյանցից տարբերելու համար։
Շատ տարիներ առաջ, համեմատելով կամ դիտելով, ինչպես ուրիշները համեմատել էին, Գալապագոս արշիպելագի մեկը մյուսից մոտիկ գտնվող կղզիների թռչունները միմյանց հետ և Ամերիկայի մայր ցամաքի թռչունների հետ, ես չափազանց զարմացել էի, թե ինչքան անորոշ ու կամայական է տեսակի և տարատեսակի միջև տարբերությունը։ Մադեյրա կղզու փոքրիկ կղզյակների խմբի վրա կան շատ միջատներ, որոնք Ուոլլաստոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 65] հիանալի աշխատության մեջ նշվում են որպես տարատեսակներ, բայց որոնց շատ միջատաբաններ կհամարեին ինքնուրույն տեսակներ։ Նույնիսկ Իռլանդիան ունի մի քանի կենդանիներ, որոնք այժմ բոլորի կողմից ճանաչվում են որպես տարատեսակներ, բայց նրանք շատ կենդանաբանների կողմից համարվել են տեսակներ։ Մի քանի փորձված թռչնաբաններ մեծ բրիտանական կարմիր մայրահավը համարում են նորվեգիական մի տեսակի խիստ արտահայտված ցեղը, մինչդեռ մեծամասնությունը նրան համարում է անկասկածելի տեսակ, որը հատուկ է միայն Մեծ Բրիտանիային։ Երկու կասկածելի ձևերի հայրենիքների տեղի միջև եղած մեծ հեռավորությունը շատ բնախույզների դրդում է համարելու դրանք որպես ինքնուրույն տեսակներ, բայց հաճախ հարց է ծագել, թե ինչպիսի՞ հեռավորությունը պետք է համարել բավարար, եթե հեռավորությունը Ամերիկայի և Եվրոպայի միջև բավարար է, այդ դեպքում կարելի՞ է արդյոք այդպիսին համարել Եվրոպայից մինչև Ազորյան կղզիները, մինչև Մադեյրա կամ մինչև Կանարյան կղզիները, կամ այդ փոքրիկ արշիպելագների զանազան կղզյակների միջև եղած հեռավորությունը։
Միստր Բ. Դ. Ուելշը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 66] որը հայտնի միջատաբան է Միացյալ Նահանգներում, նկարագրել է այն, ինչ նա անվանում է ֆիտոֆագ տեսակներ և ֆիտոֆագ տարատեսակներ։ Բույսերով սնվող միջատների մեծամասնությունը ապրում է բացառապես մեկ բուսատեսակի կամ բուսախմբի վրա, մյուսները սնվում են առանց խտրության բազմաթիվ բուսատեսակներով, սակայն չդրսևորելով դրա հետևանքով առաջացող փոփոխություններ։ Բայց և այնպես մի քանի դեպքերում զանազան բույսերի վրա հանդիպող միջատները, ըստ մ-ր Ուելշի դիտողությունների կամ թրթուրի, կամ հասուն վիճակում, կամ թե՛ մեկ և թե՛ մյուս վիճակում միասին ցուցաբերում են գույնի, մեծության կամ իրենց արտաթորումների հատկությունների աննշան, բայց հաստատուն տարբերություններ։ Որոշ դեպքերում միայն արուները, այլ դեպքերում թե արուները և թե էգերը ցուցաբերել են այդ աննշան տարբերությունները։ Երբ տարբերությունները շատ թե քիչ խիստ կերպով են արտահայտվում և ընդգրկում են երկու սեռերը և բոլոր հասակները, այդ դեպքում բոլոր միջատաբաններն այդ ձևերը համարում են լավ տեսակներ։ Բայց ոչ մի դիտող հանձն չի առնի որոշել ուրիշի համար, նույնիսկ այն դեպքում, եթե ինքն իրեն համար նա կարողանար այդ անել, թե այդ ֆիտոֆագ ձևերից որոնք պիտի համարել տեսակներ, որոնք՝ տարատեսակներ։ Միստր Ուելշը տարատեսակներ է համարում այն ձևերը, որոնք կարելի է ենթադրել, պահպանել են ազատ կերպով խաչաձևվելու ընդունակությունը, իսկ նրանք, որոնք ըստ երևույթին կորցրել են այդ ընդունակությունը, համարում է տեսակներ։ Քանի որ տարբերությունները կախված են նրանից, որ միջատները երկար ժամանակ սնվել են տարբեր բույսերով, ապա հազիվ թե կարելի է սպասել, որ այժմ կարելի լիներ գտնել նրանց կապակցող ձևերը։ Այսպիսով բնախույզը զրկվում է ինքնուրույն ձևը տեսակ կամ տարատեսակ համարելու իր լավագույն չափանիշից։ Անպայման նույնն է պատահում նաև իրար մոտիկ նմանություն ունեցող օրգանիզմների հետ, որոնք ապրում են տարբեր մայր ցամաքների կամ կղզիների վրա։ Իսկ երբ, ընդհակառակը, կենդանին կամ բույսը տարածված են միևնույն մայր ցամաքի վրա կամ ապրում են նույն արշիպելագի կղզիների վրա և զանազան շրջաններում ներկայացնում են տարբեր ձևեր, ապա միշտ ավելի հավանական է, որ կգտնվեն այն միջանկյալ օղակները, որոնք միմյանց հետ կկապեն սահմանային ձևերը, և վերջիններս կդառնան տարատեսակներ։
Մի քանի, թեև սակավաթիվ բնախույզներ պնդում են, թե կենդանիները երբեք չեն ներկայացնում տարատեսակներ, բայց դրա փոխարեն այդ նույն բնախույզներն ամենափոքր տարբերությանը տալիս են տեսակային նշանակություն, իսկ երբ միևնույն ձևը հանդիպում է երկու մեկը մյուսից հեռավոր երկրներում կամ երկրաբանական տարբեր ֆորմացիաներում, նրանք հետևեցնում են, որ ընդհանուր արտաքինի տակ թաքնված են երկու տեսակներ։ Ուրեմն՝ «տեսակ» տերմինը և դառնում է անօգուտ վերացական հասկացողություն, որը նշում է կամ նկատի է ունենում առանձին ստեղծագործական մի գործողություն։ Կասկածից դուրս է, որ շատ ձևեր, որոնք բարձր հեղինակություն ունեցող դատավորների կողմից ճանաչվում են որպես տարատեսակներ, այն աստիճան նման են տեսակներին, որ ոչ պակաս հեղինակավոր դատավորների կողմից համարվել են տեսակներ։ Բայց այդ հարցի քննարկությունը, թե արդյոք պետք է նրանց տեսակներ կամ տարատեսակներ անվանել, քանի դեռ գոյություն չունի այդ տերմինների համար մի հանրաճանաչ սահմանում, դա կնշանակե իզուր տեղը շաղակրատել։
Այդ խիստ արտահայտված տարատեսակներից կամ կասկածելի տեսակներից շատերը արժանի են ուշադրության, որովհետև աշխարհագրական տարածման համանման վարիացիաների, հիբրիդիզմի և այլ բնագավառներից վերցված հետաքրքրական կշռադատումների շարաններ են առաջադրվել նրանց տաքսոնոմիկ կարգը սահմանելու փորձերում, սակայն տեղի սղությունը թույլ չի տալիս ինձ երկար խոսել այդ նյութի մասին։ Ավելի ուշադիր հետազոտությունը շատ դեպքերում, անկասկած, բնախույզներին կբերի համաձայնության այն հարցում, թե ինչ տեղ պիտի հատկացվի կասկածելի ձևերին։ Սակայն պետք է նկատել, որ հենց ամենալավ ուսումնասիրված երկրներում նրանք ամենամեծ քանակությամբ են հանդիպում։ Ինձ շարունակ զարմացրել է այն փաստը, որ եթե բնական վիճակում որևէ կենդանի կամ բույս շատ օգտակար է մարդուն կամ որևիցե պատճառով գրավում է նրա հատուկ ուշադրությունը, ապա գրեթե անպայման հանդիպում են նշումներ նրանց տարատեսակների մասին։ Բացի դրանից, մի քանի գիտնականներ այդ տարատեսակները պիտի տարատեսակներ համարեն։ Նայեցեք կաղնուն, թե ինչքան նա մանրազնին կերպով է ուսումնասիրվել. բայց և այնպես մի գերմանացի բուսաբան դյուժինից ավելի տեսակներ է հորինել այն ձևերից, որոնք համարյա բոլոր մյուս բուսաբանների կողմից ճանաչվում են որպես տարատեսակներ, իսկ Անգլիայում կարելի է բերել շատ հեղինակավոր բուսաբանների և գործնականորեն իրազեկ մարդկանց վկայությունը ինչպես հօգուտ այն բանի, որ կաղնու երկու ձևերը (Quercus sessiliflora և pedunculata) ինքնուրույն տեսակներ են, այնպես էլ հօգուտ այն բանի, որ նրանք միայն տարատեսակներ են։
Այստեղ ևս կհիշատակեմ Ա. դը-Կանդոլի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] վերջերս լույս տեսած հիանալի աշխատության մասին, որը վերաբերում է ամբողջ աշխարհի կաղնիներին։ Ոչ ոք, իհարկե, երբեք ավելի առատ նյութ չի ունեցել տեսակները սահմանելու համար և մեծ նախանձախնդրությամբ ու խորաթափանցությամբ չի մշակել այն։ Նա ամենից առաջ մանրամասնորեն թվարկում է կառուցվածքի այն գծերը, որոնք զանազան տեսակների մոտ փոփոխվում են և որոշում է թվերով, թե որքան հաճախ են հանդիպում այդ փոփոխությունները։ Նա նշում է դյուժինից ավելի հատկանիշներ, որոնք կարող են փոփոխվել նույնիսկ միևնույն ճյուղի վրա, երբեմն կախված տարիքից կամ զարգացման աստիճանից, երբեմն էլ առանց որևէ նկատելի պատճառի։ Այդ հատկությունները, իհարկե, տեսակային նշանակություն չունեն, բայց նրանք, ինչպես եզրափակում է Ազա-Գրեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 68] այդ մեմուարին նվիրված հոդվածում, սովորաբար մտնում են տեսակային բնութագրերի կազմի մեջ։ Դը-Կանդոլն այնուհետև պարզաբանում է, որ ինքը տեսակներ է անվանում միայն այն ձևերը, որոնք տարբերվում են միմյանցից միևնույն ծառի վրա երբեք չփոփոխվող և միջանկյալ աստիճաններ երբեք չցուցաբերող հատկանիշներով։ Այդքան աշխատանքների արգասիքը հանդիսացող մանրազնին քննարկությունից հետո նա ամփոփումներ է անում այսպիսի խիստ արտահայտություններով։ «Սխալվում են նրանք, ովքեր շարունակում են կրկնել, թե մեր տեսակների մեծամասնությունը հատուկ կերպով սահմանազատված է մեկը մյուսից և որ կասկածելի տեսակները չնչին փոքրամասնություն են կազմում։ Դա կարող էր ճիշտ թվալ, քանի դեռ մի որևիցե սեռ բավականաչափ հայտնի չէր, իսկ նրա տեսակները, որ սահմանվել էին սակավաթիվ անհատների հիման վրա, ունեին, այսպես ասած, ժամանակավոր բնույթ։ Բայց հենց որ մեր տեղեկություններն սկսում են ընդարձակվել, միջանկյալ ձևերը մեկը մյուսի հետևից երևան են գալիս, իսկ դրանց հետ միասին աճում են նաև կասկածները տեսակի սահմանների վերաբերմամբ»։ Նա այնուհետև ավելացնում է, որ հատկապես ամենալավ հայտնի տեսակներն են ունենում մեծ թվով բնական տարատեսակներ և կիսատարատեսակներ։ Այսպես, Quercus robur-ը ունի քսանութ տարատեսակ. նրանք բոլորը բացառությամբ վեցի, խմբավորվում են երեք ենթատեսակների՝ Q. pedunculata-ի, sessiliflora-ի և pubescens-ի շուրջը։ Այդ երեք ենթատեսակները միացնող ձևերը համեմատաբար սակավաթիվ են, և եթե, ինչպես դարձյալ նկատում է Ազա-Գրեյը, այդ կապակցող ձևերը, որոնք այժմ հազվագյուտ են, վերջնականապես անհետանային, ապա այդ երեք ենթատեսակը կընկնեին նույն փոխհարաբերության մեջ, ինչպիսի փոխհարաբերության մեջ են գտնվում այն հինգ կամ վեց ժամանակավորապես սահմանված տեսակները, որ սերտ կերպով խմբված են տիպիկ Quercus robur-ի շուրջը։ Եզրափակման մեջ դը-Կանդոլն ընդունում է, որ այն 300 տեսակներից, որոնք պիտի թվարկվեին իր Prodromus-ի մեջ որպես կաղնու ընտանիքին պատկանողներ, առնվազն երկու երրորդը միայն ժամանակավոր նախնական տեսակներ են, այսինքն դեռ լիովին չեն բավարարում իսկական տեսակի վերոհիշյալ սահմանմանը։ Պետք է ավելացնել, որ դը-Կանդոլն այլևս չի հավատում, որ տեսակները անշարժ են ստեղծված, և գալիս է այն եզրակացության, որ նրանց ծագման միասնականության տեսությունն ավելի բնական է «և ավելի է համապատասխանում հնէաբանության, բույսերի և կենդանիների աշխարհագրության, անատոմիայի և դասակարգության հայտնի փաստերին»։
Երբ երիտասարդ բնախույզը սկսում է ուսումնասիրել իրեն բոլորովին անծանոթ օրգանիզմների մի խումբ, նա սկզբում տարակուսանքի մեջ է գտնվում, թե ինչպիսի տարբերությունները համարի որպես տեսակային, ինչպիսիները՝ տարատեսակային, որովհետև ոչինչ չգիտե այդ խմբին հատուկ փոփոխությունների չափերի և որակի մասին, իսկ դա առնվազն ցույց է տալիս, թե տարատեսակները որքան սովորական են։ Բայց եթե նա սահմանափակել է իր ուշադրությունը միայն մի որևէ դասով, մեկ երկրի սահմաններում, այդ դեպքում նա շուտով կգա որոշ եզրակացության կասկածելի ձևերի մեծամասնության մասին։ Սկզբում նա կհակվի բազմաթիվ տեսակներ սահմանելու կողմը, որովհետև աղավնիների և հավերի վերևում հիշված սիրահարների նման նա կզարմանա հետազոտվող ձևերի փոփոխականության չափերի վրա, չունենալով դեռ բավարար տեղեկություններ այլ խմբերում և այլ երկրներում երևան եկող համանման փոփոխությունների մասին, տեղեկություններ, որոնք կարող էին ուղղել նրա առաջին տպավորությունները։ Ընդարձակելով իր դիտումների սահմանները, նա կհանդիպի մեծ թվով դժվարին դեպքերի, որովհետև կհանդիպի ավելի բազմաթիվ միմյանց հետ մոտիկ ձևերի։ Եթե նրա դիտողությունները ավելի ևս կընդարձակվեն, վերջապես նա կվարժվի այդ կասկածելի դեպքերից գլուխ հանել, բայց այդ հետևանքին կհասնի նա միայն այն գնով, երբ կընդունի, որ ձևերը մեծ չափերով փոփոխական են, իսկ այդ եզրակացության արդարացիությունն իր հերթին կվիճարկվի այլ բնախույզների կողմից։ Իսկ եթե նա առիթ կունենա ուսումնասիրելու մերձավոր ձևեր, որոնք ստացված են ներկայումս իրար ոչ սահմանակից երկրներից, ընդ որում նա չի կարող նույնիսկ սպասել, որ կգտնի միջանկյալ օղակները, նա ստիպված կլինի հիմնվել համարյա ամբողջապես անալոգիաների վրա, և նրա դժվարությունները կհասնեն իրենց գագաթնակետին։
Անշուշտ մինչև այժմ դեռ չի հաջողվել որոշակի սահմանագիծ անցկացնել տեսակների և ենթատեսակների, այսինքն՝ այնպիսի ձևերի միջև, որոնք մի քանի բնախույզների կարծիքով մոտենում են տեսակներին, բայց լիովին չեն հասնում այդ աստիճանին, կամ ենթատեսակների և ակնհայտ տարատեսակների միջև, կամ վերջապես, թույլ տարատեսակների և անհատական տարբերությունների միջև։ Այդ տարբերությունները հարակցվում են մեկը մյուսին, աննշմարելի կերպով միաձուլվելով՝ կազմում են մի անընդհատ շարք, իսկ ամեն մի շարք մեր մտքի վրա իսկական անցման տպավորություն է թողնում։
Դրա հիման վրա անհատական տարբերությունները, որոնք թեև քիչ հետաքրքիր են սիստեմատիկայի համար, ես համարում եմ մեզ համար չափազանց կարևոր՝ որպես տարատեսակներ առաջանալու առաջին քայլեր, որոնք այնքան աննշան են, որ նրանց մասին, ինչպես սովորաբար կարծում են, չարժե անգամ հիշատակել բնապատմական գրվածքներում։ Այն տարատեսակները, որոնք ավելի արտահայտված են ու հաստատուն, ես համարում եմ որպես քայլեր դեպի ավելի խիստ արտահայտված ու հաստատուն տարատեսակները, իսկ այս վերջինները՝ որպես քայլեր դեպի ենթատեսակները և տեսակները։ Տարբերության մեկ աստիճանից մյուսին անցումը շատ դեպքերում տեղի է ունեցել շնորհիվ հենց օրգանիզմի առանձնահատկությունների և զանազան ֆիզիկական պայմանների, որոնց նա երկար ժամանակ ենթարկվել է, բայց հարմարումների բնույթ կրող ամենից ավելի էական հատկանիշների վերաբերմամբ մի աստիճանից մյուսին անցումը կարելի է վստահորեն վերագրել բնական ընտրություն կուտակող ներգործությանը, ինչպես այդ կպարզաբանվի հետագա շարադրանքում, ինչպես նաև օրգանների վարժեցմանը և ոչ վարժեցմանը։ Պարզ կերպով արտահայտված տարատեսակը այդ պատճառով կարող է անվանվել նոր գոյացող տեսակ, թե որքան ճիշտ է այդ հետևությունը, կարելի կլինի դատել միայն այն փաստերի և կշռադատումների հիման վրա, որոնք շարադրված են այս ամբողջ աշխատության մեջ։
Անհրաժեշտություն չկա ենթադրելու, որ բոլոր տարատեսակները կամ սկզբնավորվող տեսակները հասնում են տեսակների աստիճանի։ Նրանք կարող են մահաջնջվել կամ կարող են պահպանվել շատ երկար ժամանակաշրջանների ընթացքում տարատեսակի աստիճանի վրա, ինչպես այդ ցույց է տվել Ուոլլաստոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 65] Մադեյրայի վրա անուշահամ ջրերի մի քանի բրածո խեցիների տարատեսակների վերաբերմամբ և Գաստոն դե-Սապորտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 69] բույսերի վերաբերմամբ։ Եթե տարատեսակը այնպիսի ծաղկման աստիճանի հասներ, որ թվով գերազանցեր ծնողական ձևին, այն ժամանակ նա կդառնար տեսակ, իսկ տեսակը կդառնար տարատեսակ, կամ նա կարող էր դուրս մղել և բոլորովին փոխարինել ծնողական տեսակային ձևին, կամ վերջապես, երկուսն էլ կարող էին միաժամանակ գոյություն ունենալ և համարվել ինքնուրույն տեսակներ։ Բայց մենք այդ հարցին կվերադառնանք հետագա շարադրանքում։
Ամբողջ ասվածից պարզ է, որ «տեսակ» տերմինը ես համարում եմ միանգամայն կամայական, հորինված իրար նման ու մերձավոր անհատների մի խումբ նշելու հարմարության համար և էականապես չի տարբերվում «տարատեսակ» տերմինից, որով նշվում են իրենց հատկանիշներով ավելի քիչ տարբերվող և տատանվող ձևերը։ Այդպես էլ «տարատեսակ» տերմինը անհատական տարբերությունների վերաբերմամբ կիրառվում է կամայականորեն և միայն հարմարության համար[38]։
ԼԱՅՆ ՏԱՐԱԲՆԱԿՎԱԾ, ՏԱՐԱԾՎԱԾ Եվ ՍՈՎՈՐԱԿԱՆ ՏԵՍԱԿՆԵՐՆ ԱՄԵՆԻՑ ԱՎԵԼԻ ՓՈՓՈԽԱԿԱՆ ԵՆ
Առաջնորդվելով տեսական նկատառումներով, ես կարծում էի, թե ամենից ավելի փոփոխական տեսակների բնույթին և փոխհարաբերություններին վերաբերող հետաքրքիր տվյալներ կարելի կլինի ստանալ, եթե կազմեի մի քանի լավ մշակված ֆլորաների բոլոր տարատեսակների ցուցակները։ Առաջին հայացքից խնդիրը շատ պարզ էր թվում, բայց Հ. Կ. Ուոտսոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 62] որին ես շատ պարտական եմ այդ գործում տված նրա արժեքավոր ցուցումների և օգնության համար, շուտով համոզեց ինձ, որ դա կապված է մեծ դժվարությունների հետ. նույն իմաստով և ավելի վճռականորեն հետո արտահայտվեց նաև դոկտոր Հուկերը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] Ուստի՝ այդ դժվարությունների քննարկումը և փոփոխական տեսակների հարաբերական թիվը ցույց տվող ցուցակները հետաձգում եմ մինչև հետագա աշխատությունս։ Դոկտոր Հուկերը թույլ է տալիս ինձ ավելացնելու, որ մանրազնին կերպով կարդալով իմ ձեռագիրը և աչքի անցկացնելով աղյուսակները, հետևյալ եզրակացությունները նա համարում է միանգամայն բավարար չափով հիմնավորված։ Բայց այդ ամբողջ հարցը, որ այնտեղ շարադրված է անխուսափելի համառոտությամբ, բավական խրթին է ներկայացվում. բացի դրանից, անկարելի էր խույս տալ «գոյության կռվի», «հատկանիշների տարամիտման» վրա վկայակոչումներ անելուց, որոնց պարզաբանությունը տրվելու է առաջիկայում։
Ալֆոնս դը-Կանդոլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] և ուրիշներ ցույց են տվել, որ լայնորեն տարաբնակված բույսերը սովորաբար ունենում են տարատեսակներ. այդ կարելի է սպասել, որովհետև նրանք ենթարկվում են ֆիզիկական բազմազան պայմանների ներգործությանը և մրցության մեջ են մտնում կենդանի էակների զանազան խմբերի հետ (իսկ այդ հանգամանքը, ինչպես հետո կտեսնենք, ոչ ավելի պակաս, եթե դեռ ոչ ավելի, կարևոր է)։ Բայց իմ աղյուսակները դրանից ավելի են ցույց տալիս. նրանք ցույց են տալիս, որ ամեն մի սահմանափակ մարզում ամենասովորական տեսակները, այսինքն՝ ամենամեծ թվով անհատներ ունեցողները, և այն տեսակները, որոնք ամենալայն տարածում ունեն իրենց մարզում (այս պայմանը բոլորովին տարբեր է լայն տարաբնակությունից, իսկ որոշ իմաստով նաև տեսակի սովորականությունից), ավելի հաճախ առաջացնում են բավականաչափ ցայտուն տարատեսակներ, որոնց կարելի է մտցնել բուսաբանական աշխատությունների մեջ։ Ուրեմն՝ հենց այն տեսակները, որոնք ամենից ավելի ծաղկած են. կամ, ինչպես կարելի է նրանց անվանել, տիրապետող են, նրանք, որոնք լայնորեն տարաբնակված են, ամենից ավելի լայնորեն տարածված են իրենց մարզում և անհատներով ամենից հարուստ են, ամենից ավելի հաճախ սկիզբ են տալիս պարզ արտահայտված տարատեսակների, կամ, իմ տեսակետով, սաղմնային տեսակների։ Եվ այդ գուցե կարելի էր նախատեսել, քանի որ տարատեսակները հաստատվելու համար, ըստ անհրաժեշտության, պետք է դիմանային երկրի մյուս բնակիչների դեմ մղվող կռվին, ուստի այն տեսակները, որոնք արդեն տիրապետող են, ամենից ավելի հավանական է, որ առաջացնեն սերունդ, որը թեև փոքր ինչ տարբեր է, բայց և այնպես ժառանգել է իր նախնիներից այն առավելությունները, որոնց շնորհիվ ապահովվել է նրանց տիրապետությունը իրենց հայրենակիցների վրա։ Խոսելով տիրապետող ձևերի մասին, ես հասկանում եմ միայն այն ձևերը, որոնք փոխադարձ մրցության մեջ են գտնվում, և առանձնապես միևնույն սեռի կամ դասի ներկայացուցիչները, որոնք միանման կենսակերպ են վարում։ Անհատների թվի կամ այն բանի նկատմամբ, որը ես անվանում եմ տեսակի սովորականություն, համեմատությունը վերաբերում է միայն միևնույն խմբի անդամներին։ Որևէ բարձրակարգ բույս կարող է համարվել տիրապետող, եթե նրա անհատները ավելի բազմաթիվ և ավելի տարածված են երկրում, քան նույն երկրում և գրեթե նույն պայմաններում ապրող մյուս բույսերը։ Այդպիսի բույսը ավելի նվազ տիրապետող չի լինի, եթե նրա կողքին ջրում ապրող որևէ խոզաջրիմուռ (conferva) կամ որևէ մակաբույծ սունկ լինում են անհամար մեծ թվով և ավելի ևս լայն տարածված։ Բայց եթե խոզաջրիմուռը կամ մակաբույծ սունկը նշված իմաստով գերազանցում են իրենց մերձավոր ձևերին, այդ դեպքում նրանք տիրապետող կլինեն իրենց դասի սահմաններում։
ՅՈՒՐԱՔԱՆՉՅՈՒՐ ԵՐԿՐՈՒՄ ՄԵԾ ՍԵՌԵՐԻ ՏԵՍԱԿՆԵՐՆ ԱՎԵԼԻ ՇԱՏ ԵՆ ՓՈՓՈԽՎՈՒՄ, ՔԱՆ ՓՈՔՐ ՍԵՌԵՐԻ ՏԵՍԱԿՆԵՐԸ
Եթե որևէ երկրի բուսականության մեջ նկարագրված բույսերը երկու հավասար խմբում այնպես դասավորենք, որ մի խմբի մեջ մտնեն մեծ սեռերի (այսինքն՝ շատ տեսակներ պարունակող սեռերի) ներկայացուցիչները, իսկ մյուսում՝ փոքր սեռերի ներկայացուցիչները, ապա առաջինում կլինեն մեծ թվով սովորական և լայն տարածված կամ տիրապետող տեսակներ։ Դա կարելի էր նախատեսել. հենց այն փաստը, որ մեկ սեռի բազմաթիվ տեսակները ապրում են որոշ երկրում, արդեն ցույց է տալիս, որ այդ երկրի անօրգանական կամ օրգանական պայմաններում կա ինչ-որ բարենպաստ բան այդ սեռի համար, իսկ այստեղից մենք իրավունք ունենք սպասելու, որ մեծ սեռերի տեսակների մեջ կհանդիպենք համեմատաբար ավելի մեծ թվով տիրապետող տեսակների։ Բայց այնքան շատ կան պատճառներ, որոնք կարող են քողարկել այդ հետևանքը, և ինձ զարմացնում է, որ իմ աղյուսակներում մեծամասնությունը, թեև աննշան, դրսևորվել է մեծ սեռերի օգտին։ Կանգ կառնեմ միայն երկու պատճառների վրա, որոնք կարող են քողարկել այդ հետևանքները։ Անուշահամ ջրերի և աղուտների բույսերը սովորաբար լայն կերպով տարաբնակված են և աչքի են ընկնում մեծ տարածվածությամբ իրենց մարզի սահմաններում. ըստ երևույթին դա կապված է նրանց բնակավայրի հետ և քիչ կամ ոչ մի սերտ կապ չունի այն սեռի մեծության հետ, որին նրանք պատկանում են։ Ցածր կազմվածք ունեցող բույսերը նույնպես սովորաբար անհամեմատ ավելի լայնորեն են տարածված, քան բարձր կազմվածքով բույսերը, այստեղ ևս, իհարկե, սերտ կապ գոյություն չունի սեռի մեծության հետ։ Ստորակարգ բույսերի լայն տարաբնակման պատճառը մենք կքննարկենք աշխարհագրական տարածման վերաբերյալ գլխում։
Տեսակները դիտելով որպես միայն ավելի խիստ նկատելի և որոշակի դարձած տարատեսակներ, ես եկա այն ենթադրության, որ յուրաքանչյուր երկրում մեծ սեռերի տեսակները պետք է ավելի հաճախ տան տարատեսակներ, քան փոքր սեռերի տեսակները, որովհետև ամենուրեք, որտեղ արդեն առաջացել են մերձավոր շատ տեսակներ (այսինքն միևնույն սեռի տեսակներ) այնտեղ պետք է, որպես ընդհանուր կանոն, դեռ շարունակվի նոր տարատեսակների կամ սաղմնային տեսակների առաջացումը։ Որտեղ մեծ թվով մեծահասակ ծառեր են աճում, այնտեղ մենք սպասում ենք գտնել նաև մեծ թվով նոր աճող ծառեր։ Որտեղ փոփոխականության միջոցով միևնույն սեռի շատ տեսակներ են առաջացել, այնտեղ հանգամանքները բարենպաստ են եղել և, կարելի է սպասել, շարունակում են նպաստել այդ փոփոխականությանը։ Մյուս կողմից, եթե տեսակների վրա նայենք որպես առանձին արարչագործական ակտերի վրա, այդ դեպքում ոչ մի հիմք չկա սպասելու, որ տարատեսակները ավելի բազմաթիվ լինեն տեսակներով հարուստ խմբում, քան նրանցով աղքատ խմբում։
Այդ ենթադրությունը ստուգելու համար ես տասներկու երկրների բույսեր և երկու մարզի կարծրաթև միջատներ դասավորեցի երկու գրեթե հավասար խմբերում՝ մեծ սեռերի տեսակները մի կողմ, փոքր սեռերի տեսակները մյուս կողմ, և անփոփոխ դուրս եկավ այն կանոնը, որ տարատեսակներ պարունակող տեսակների հարաբերական մեծ թիվը մեծ սեռերի օգտին էր, այլ ոչ թե փոքր սեռերի օգտին։ Բացի դրանից, մեծ սեռերի տեսակները, եթե միայն նրանք տալիս են տարատեսակներ, մշտապես ավելի շատ տարատեսակներ են ներկայացնում, քան փոքր սեռերի տեսակները։ Այս երկու արդյունքներն ստացվում են նաև մի քիչ այլ կերպ խմբավորելիս, այսինքն երբ մեկից մինչև չորս տեսակներ պարունակող ամենափոքր սեռերը ոչ ամբողջապես են մտցված աղյուսակների մեջ։ Այդ փաստերի իմաստը շատ պարզ է, եթե ընդունենք, որ տեսակները միայն խիստ արտահայտված և հաստատուն տարատեսակներ են, ամենուրեք, որտեղ շատ տեսակներ են առաջացել, կամ եթե կարելի է այսպես արտահայտվել, ամենուրեք, որտեղ տեսակների պատրաստումը հաջող է ընթացել, մենք ընդհանրապես պետք է այդ պատրաստումը դեռ տեսնենք գործողության մեջ, մանավանդ որ ունենք բոլոր հիմքերը ենթադրելու, որ նոր տեսակների պատրաստման այդ պրոցեսը պետք է շատ դանդաղ կատարվի։ Եվ այդ կշռադատությունը արդարանում է, եթե միայն տարատեսակները համարենք սաղմնային տեսակներ, որովհետև իմ աղյուսակները պարզորոշ կերպով հաստատում են այն ընդհանուր կանոնը, որ յուրաքանչյուր սեռում, որի մեջ շատ տեսակներ են առաջացել, այդ տեսակները ներկայացնում են միջինից ավելի բարձր թվով տարատեսակներ կամ սաղմնային տեսակներ։ Դրանից չի հետևում, որ բոլոր մեծ սեռերը այժմ շարունակում են խիստ փոփոխվել կամ ոչ մի փոքր սեռ չի փոփոխվում և չի մեծանում. այդպիսի փաստը ճակատագրական կլիներ իմ տեսության համար, որովհետև երկրաբանությունը լիակատար պարզությամբ պատմում է, որ փոքր սեռերը ժամանակի ընթացքում աճել, դարձել են մեծ, իսկ մեծ սեռերը հաճախ հասել են առավելագույն զարգացման և հետո թեքվել են դեպի անկում և անհետացել։ Մեզ հարկավոր է միայն ցույց տալ, որ այնտեղ, որտեղ մեկ սեռի սահմաններում առաջացել են շատ տեսակներ, միջին հաշվով վերցրած, նրանք դեռ շարունակում են առաջանալ, իսկ դա, իհարկե, ճիշտ է դուրս գալիս։
ՄԵԾ ՍԵՌԵՐԻ ՇԱՏ ՏԵՍԱԿՆԵՐ ՆՄԱՆ ԵՆ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԱՅՆ ԲԱՆՈՎ, ՈՐ ՆԵՐԿԱՅԱՑՆՈՒՄ ԵՆ ՇԱՏ ՄՈՏԻԿ, ԲԱՅՑ ՈՉ ՄԻԱՆՄԱՆ ԱԶԳԱԿՑՈՒԹՅՈՒՆ ՄԻՄՅԱՆՑ ՀԵՏ ԵՎ ՈՒՆԵՆ ՍԱՀՄԱՆԱՓԱԿ ՏԱՐԱԾՈՒՄ
Կան ուշադրության արժանի այլ հարաբերություններ մեծ սեռերի տեսակների և նրանց տարատեսակների միջև, որոնք մտցված են ցուցակների մեջ։ Մենք տեսանք, որ տեսակը խիստ արտահայտված տարատեսակից տարբերելու համար չկա մի անսխալական չափանիշ։ Երբ երկու կասկածելի ձևերի միջև միջանկյալ օղակներ գտնված չեն, բնախույզներն ստիպված են իրենց եզրակացություններում ղեկավարվել այդ ձևերի միջև եղած տարբերության չափերով և լուծել այն հարցը, թե բավակա՞ն է արդյոք տարբերության այդ չափը նրանցից մեկին կամ երկուսին էլ տեսակի աստիճանի վրա բարձրացնելու համար։ Ուրեմն տարբերության չափերը շատ կարևոր չափանիշ են հանդիսանում լուծելու համար այն հարցը, թե պե՞տք է արդյոք երկու ձևերը համարել տեսակներ կամ տարատեսակներ։ Բայց Ֆրիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 70] բույսերի վերաբերմամբ, իսկ Ուեստվուդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 71] միջատների վերաբերմամբ արդեն նշել են, որ մեծ սեռերի մեջ տեսակների միջև տարբերության չափերը չափազանց փոքր են։ Ես աշխատել եմ նրանց եզրակացությունը ենթարկել թվական ստուգման միջին մեծություններ հանելու միջոցով, և իմ ստացած շատ անկատար արդյունքների սահմաններում այդ կանոնը հաստատվել է։ Ես դիմել եմ նաև մի քանի խոհուն և փորձված դիտողների, և գործը ուշադրությամբ քննարկելուց հետո նրանք համաձայն են այդ եզրակացությանը։ Հետևաբար, այդ տեսակետից մեծ սեռերի տեսակները ավելի նմանվում են տարատեսակներին, քան փոքր սեռերի տեսակները։ Կամ, ուրիշ խոսքով, մեծ սեռերում, որոնց մեջ տարատեսակների կամ սաղմնային տեսակների պատրաստումը արտահայտվում է միջինից ավելի բարձր թվով, արդեն պատրաստված տեսակներից շատերը որոշ չափով դեռ նման են տարատեսակների, որովհետև նրանց միջև եղած տարբերությունների չափերը սովորականներից ավելի փոքր են։
Բացի դրանից, մեծ սեռերի տեսակները մեկը մյուսին հարաբերում են այնպես, ինչպես մի տեսակի տարատեսակները միմյանց։ Ոչ մի բնախույզ չի կարող պնդել, որ մի սեռի բոլոր տեսակները միմյանցից տարբերվում են միանման չափով։ Նրանց սովորաբար կարելի է բաժանել ենթասեռերի, կամ հատվածների, կամ այլ ավելի մանր խմբերի։ Ինչպես միանգամայն ճիշտ է նկատում Ֆրիսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 70] տեսակների փոքր խմբերը սովորաբար արբանյակների պես խմբավորվում են ուրիշ տեսակների շուրջը։ Իսկ ի՞նչ են տարատեսակները, եթե ոչ ձևերի խմբեր, որոնք ոչ միանման մոտիկություն ունեն միմյանց հետ և համախմբված են այլ ձևերի՝ իրենց նախածնող տեսակների շուրջը։ Անտարակույս մի շատ կարևոր տարբերություն գոյություն ունի տարատեսակների և տեսակների միջև, այն է՝ տարատեսակների տարբերության չափերը ինչպես իրար հետ, այնպես էլ նախածնողական տեսակային ձևի հետ համեմատած ավելի քիչ են, քան նույն սեռի տեսակների միջև եղած տարբերությունները, բայց երբ խոսք կլինի այն բանի մասին, որ ես անվանում եմ հատկանիշների տարամիտում, մենք կտեսնենք, թե ինչով է այդ բացատրվում և ինչպես տարատեսակների միջև եղած փոքրիկ տարբերությունները ձգտում են մեծանալ, դառնալ զգալի տարբերություններ տեսակների միջև։
Մի նկատառում ևս արժանի է ուշադրության։ Տարատեսակները սովորաբար ունենում են տարածման շատ սահմանափակ շրջաններ։ Այդ դրույթն ըստ էության ծեծված ճշմարտությունից ավելի չէ, այսինքն ինքնըստինքյան ակնհայտ է, որովհետև այն դեպքում, եթե դուրս գար, որ տարատեսակն ավելի լայն տարածում ունի, քան նրա ենթադրյալ նախածնողական տեսակը, նրանք կփոխանակեին իրենց անունները։ Բայց հիմք կա մտածելու, որ այն տեսակները, որոնք շատ մոտ են այլ տեսակների և դրանով էլ նման են տարատեսակների, հաճախ ունենում են շատ սահմանափակ տարածում։ Այդպես, օրինակ, Հ. Կ. Ուոտսոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 62] հիանալի կազմված բույսերի Լոնդոնյան ցուցակում (4-րդ հրատարակություն) ինձ համար նշել է 63 բույս, որոնք այնտեղ հաշվվում են որպես տեսակներ, բայց նրա կարծիքով, նրանք այնքան մոտ են մյուս տեսակներին, որ կասկած են հարուցում իրենց ինքնուրույնության մասին. այդ կասկածելի 63 տեսակները, միջին թվով, գրավում են 6,9 այն մարզերի, որոնց վրա բաժանել է միստր Ուոտսոնը Մեծ Բրիտանիան։ Սույն ցուցակում բերված են 53 հանրաճանաչ տարատեսակներ. և այդ տարատեսակները տարածված են 7,7 մարզերում, մինչդեռ այն տեսակները, որոնց պատկանում են այդ տարատեսակները, տարածված են 14,3 մարզերում։ Այսպիսով, հանրաճանաչ տարատեսակները միջին հաշվով ունեն համարյա նույնպիսի սահմանափակ տարածում, ինչպիսին ունեն միմյանց հետ մոտիկ ձևերը, որոնց նշել էր Ուոտսոնը ինձ համար որպես կասկածելի տեսակներ, բայց նրանք համարյա բոլոր անգլիական բուսաբանների կողմից ճանաչված են որպես իսկական, լավ տեսակներ։
ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ
Ուրեմն՝ տարատեսակը տեսակներից այլ կերպ չի կարելի զանազանել, եթե ոչ, նախ՝ հայտնաբերելով նրանց շաղկապող միջանկյալ ձևերը և երկրորդ ապացուցելով մի ինչ-որ անորոշ չափի տարբերության առկայությունը, որովհետև միմյանցից քիչ տարբերվող ձևերը սովորաբար համարվում են տարատեսակներ, թեկուզև նրանց չի կարելի անմիջականորեն կապել միմյանց հետ։ Սակայն տարբերությունների չափը, որ անհրաժեշտ է համարվում երկու ձևեր տեսակների աստիճանի բարձրացնելու համար, չի կարելի որոշել։ Այն սեռերը, որոնք պարունակում են տվյալ երկրի համար միջինից բարձր թվով տեսակներ, այդ տեսակներն էլ տալիս են միջինից ավելի բարձր թվով տարատեսակներ։ Մեծ սեռերում տեսակները ներկայացնում են ավելի մերձավոր, բայց ազգակցության անհավասարաչափ աստիճաններ և համախմբված են այլ տեսակների շուրջը։ Այն տեսակները, որոնք կապված են այլ տեսակների հետ շատ մոտ ազգակցությամբ, սովորաբար ունենում են սահմանափակ տարածում։ Այս բոլոր տեսակետներից մեծ սեռերի տեսակները ցուցաբերում են մոտիկ համանմանություն տարատեսակների հետ։ Եվ այդ համանմանությունները միանգամայն հասկանալի են, եթե տեսակները առաջացել են այսպես, որ իրենք առաջ եղել են տարատեսակներ, բայց վերջնականապես անբացատրելի են, եթե տեսակներն իրենցից ներկայացնում են առանձին ստեղծագործական ակտեր։
Մենք տեսանք նաև այն, որ յուրաքանչյուր դասի սահմաններում մեծ սեռերի հենց ամենից ավելի բարգավաճող և տիրապետող տեսակները առաջացնում են միջին հաշվով ամենամեծ քանակությամբ տարատեսակներ, իսկ տարատեսակները, ինչպես կտեսնենք հետագայում, ձգտում են դառնալ նոր առանձին տեսակներ։ Այսպիսով, մեծ սեռերը ձգտում են է՛լ ավելի մեծանալու, և ամբողջ բնության մեջ նկատվում է, որ կյանքի այժմյան տիրապետող ձևերը ձգտում են է՛լ ավելի տիրապետող դառնալ, իրենցից հետո, թողնելով բազմաթիվ փոփոխված ու տիրապետող հետնորդներ։ Սակայն խոշոր սեռերը մի ճանապարհով, որը կբացատրվի հետագայում, ձգտում են բաժանվել փոքրերի։ Այս կերպով էլ ամբողջ աշխարհում ամենուրեք կյանքի ձևերը բաժանվում են խմբերի, որոնք ենթակա են այլ խմբերի։
ԳԼՈՒԽ III։ ԳՈՅՈՒԹՅԱՆ ԿՌԻՎ
ՆՐԱ ԱՌՆՉՈՒԹՅՈՒՆԸ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՀԵՏ։— «ԳՈՅՈՒԹՅԱՆ ԿՌԻՎ» ՏԵՐՄԻՆԻ ԳՈՐԾԱԾՈՒՄԸ ԼԱՅՆ ԻՄԱՍՏՈՎ։— ՄՐՑՈՒԹՅԱՆ ԸՆԴՀԱՆՐԱԿԱՆՈՒԹՅՈՒՆԸ։— ԲԱԶՄԱՑՄԱՆ ԵՐԿՐԱՉԱՓԱԿԱՆ ՊՐՈԳՐԵՍԻԱՆ։— ՆԱՏՈՒՐԱԼԻԶԱՑԻԱՅԻ ԵՆԹԱՐԿՎԱԾ ԿԵՆԴԱՆԻՆԵՐԻ ԵՎ ԲՈՒՅՍԵՐԻ ԱՐԱԳ ԲԱԶՄԱՑՈՒՄԸ։— ԲԱԶՄԱՑՈՒՄԸ ԿԱՍԵՑՆՈՂ ԽՈՉԸՆԴՈՏՆԵՐԻ ԲՆՈԻՅԹԸ։— ԿԼԻՄԱՅԻ ԱԶԴԵՑՈՒԹՅՈՒՆԸ։— ԱՆՀԱՏՆԵՐԻ ԲԱԶՄԱՔԱՆԱԿՈՒԹՅՈՒՆԻՑ ԿԱԽՎԱԾ ՊԱՇՏՊԱՆՈՒԹՅՈՒՆԸ։— ԲՆՈՒԹՅԱՆ ՄԵՋ ԱՄԵՆՈՒՐԵՔ ԿԵՆԴԱՆԻՆԵՐԻ ԵՎ ԲՈՒՅՍԵՐԻ ՄԻՋԵՎ ՀԱՐԱԲԵՐԱԿՑՈՒԹՅՈՒՆՆԵՐԻ ՆԿԱՏՎՈՂ ԲԱՐԴՈՒԹՅՈՒՆԸ։ ԳՈՅՈՒԹՅԱՆ ԿՌԻՎԸ ԱՄԵՆԻՑ ԱՎԵԼԻ ՀԱՄԱՌ Է ՄԻԵՎՆՈՒՅՆ ՏԵՍԱԿԻ ԱՆՀԱՏՆԵՐԻ ԵՎ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ՄԻՋԵՎ, ՀԱՃԱԽ ՆԱԵՎ ՄԻԵՎՆՈՒՅՆ ՍԵՌԻ ՏԵՍԱԿՆԵՐԻ ՄԻՋԵՎ։— ՕՐԳԱՆԻԶՄՆԵՐԻ ՄԻՋԵՎ ՓՈԽՀԱՐԱԲԵՐՈՒԹՅՈՒՆՆԵՐԸ ԲՈԼՈՐ ՀԱՐԱԲԵՐՈՒԹՅՈՒՆՆԵՐԻՑ ԱՄԵՆԱԿԱՐԵՎՈՐՆ ԵՆ։
Նախքան այս գլխի բովանդակությունը կազմող նյութի շարադրման անցնելը, ես պետք է մի քանի նախնական դիտողություններ անեմ, որպեսզի ցույց տամ, թե գոյության կռիվը ինչ հարաբերության մեջ է գտնվում բնական ընտրության նկատմամբ։ Նախորդ գլխում մենք տեսանք, որ բնական վիճակում գտնվող օրգանական էակները ցուցաբերում են որոշ աստիճանի փոփոխականություն։ Ճիշտն ասած՝ ես չեմ կարծում, որ այդ դրույթը երբևիցե վիճարկվել է։ Մեզ համար էական չէ, թե տեսակներ, ենթատեսակներ կամ տարատեսակներ կանվանվեն բազմաթիվ կասկածելի ձևերը, ինչպես, օրինակ, բույսերի այն կասկածելի ձևերը, որոնք հաշվվում են բրիտանական բուսականության մեջ, քանի որ հստակ կերպով արտահայտված տարատեսակների գոյությունն ընդունվում է բոլորի կողմից։ Միայն անհատական տարբերությունների և մի քանի խիստ նկատելի տարատեսակների գոյությունը, թեև այդ անհրաժեշտ էլ է որպես մեկնակետային փաստ, բայց շատ քիչ է օգնում մեզ ըմբռնելու այն, թե բնության մեջ տեսակներն ինչպես են առաջանում։ Ի՞նչ կերպով այդպիսի կատարելության են հասել այն զարմանալի հարմարումները, որ ունի օրգանիզմի մի մասը մյուսի կամ ամբողջ օրգանիզմը արտաքին կենսապայմանների կամ, վերջապես, մի օրգանիզմ մյուսի նկատմամբ։ Մենք այդ սքանչելի փոխադարձ հարմարումները լիակատար ակներևությամբ տեսնում ենք փայտփորի կամ ճագոմի վրա և փոքր ինչ ավելի պակաս ակներև այն ողորմելի մակաբույծի մեջ, որը կպած է չորքոտանու մազին կամ թռչունի փետուրին, ջրի մեջ սուզվող բզեզի կառուցվածքում, թեթև քամուց թռչող սերմի հարմարանքներում,— մի խոսքով՝ մենք սքանչելի հարմարումներ ենք տեսնում ամեն տեղ և օրգանական աշխարհի ամեն մի մասում։
Այնուհետև կարելի է հարցնել, թե ինչպես է, որ այդ տարատեսակները, որոնց ես անվանել եմ սաղմնային տեսակներ, վերջ ի վերջո դարձել են լավ և ուրույն տեսակներ, որոնք մեծ մասամբ ակներևորեն անհամեմատ ավելի են տարբերվում միմյանցից, քան մի տեսակի պատկանող տարատեսակները։ Ինչպե՞ս են առաջանում տեսակների այդ խմբերը, որ առաջացնում են այն, ինչ մենք անվանում ենք սեռեր և որոնք միմյանցից անհամեմատ ավելի շատ են տարբերվում, քան միևնույն սեռի տեսակները։ Այս բոլոր հետևանքները, ինչպես մենք ավելի հանգամանորեն կպարզենք հաջորդ գլխում, ուղղակի բխում են գոյության կռվից։ Այդ կռվի շնորհիվ փոփոխությունները, ինչքան էլ նրանք աննշան լինեն և ինչ պատճառից էլ կախված լինեն, եթե միայն նրանք օգտակար են տվյալ տեսակի անհատների համար նրանց ունեցած անսահման բարդ հարաբերություններում այլ էակների և ֆիզիկական կենսապայմանների հետ, կնպաստեն այդ անհատների պահպանմանը և ժառանգաբար կանցնեն նրանց ամենամերձավոր հետնորդներին։ Այդ հետնորդները իրենց հերթին պահպանվելու ավելի շատ հավանականություններ կունենան, որովհետև որևէ տեսակի բազմաթիվ անհատներից, որ պարբերաբար ծնվում են, միայն աննշան քանակությունն է կենդանի մնում։ Այդ սկզբունքը, որի հետևանքով յուրաքանչյուր աննշան փոփոխություն, եթե միայն նա օգտակար է, պահպանվում է, ես անվանել եմ բնական ընտրություն նրա համար, որպեսզի ցույց տամ նրա առնչությունը այն ընտրության հետ, որ գործադրում է մարդը։ Սակայն Հերբերտ Սպենսերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 18] կողմից հաճախ գործածվող արտահայտությունը՝ «ամենահարմարվածի գերապրումը»,— ավելի ճիշտ է և երբեմն էլ նույնքան հարմար։ Մենք տեսանք, որ մարդն ընտրության միջոցով մեծ հետևանքների է հասնում և կարողանում է օրգանիզմը հարմարեցնել իր պահանջմունքներին՝ շնորհիվ այն թույլ, բայց օգտակար փոփոխությունների, որ նրան ընձեռում է բնությունը։ Բայց բնական ընտրությունը, ինչպես մենք կտեսնենք հետագայում, մի սկզբունք է, որը մշտապես պատրաստ է ներգործելու և նույնքան անչափորեն գերազանցում է մարդու թույլ ջանքերին, ինչքան որ բնության արտադրանքները գերազանցում են արվեստի արտադրանքներին։
Մենք այժմ մի քիչ ավելի մանրամասնորեն կզբաղվենք գոյության կռվով։ Իմ ապագա աշխատության մեջ այդ խնդիրը զարգացվելու է ավելի մանրամասնորեն, որին և նա արժանի է։ Դը-Կանդոլ ավագը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] և Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] հանգամանորեն և խորիմաստ կերպով մատնանշել են այն, որ բոլոր օրգանական էակները ենթարկվում են դաժան մրցության։ Բույսերի վերաբերմամբ այդ խնդիրը ոչ ոք այնպիսի եռանդով ու հմտությամբ չի քննարկել, ինչպես Հերբերտը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 7] Մանչեստրի դեկանը, հավանորեն իր պարտիզագործական ընդարձակ գիտելիքների շնորհիվ։ Ոչ մի բան չի կարող ավելի հեշտ լինել, որքան խոսքով ընդունել գոյության այդ համընդհանուր կռվի ճշտությունը, և ոչ մի բան չի կարող ավելի դժվար լինել, ես այդ փորձել եմ ինձ վրա, որքան այդ եզրակացությունը երբեք աչքաթող չանելը։ Սակայն, մինչև որ այդ եզրակացությունը չարմատավորվի մեր մտքում, բնության ամբողջ տնտեսությունը սրան վերաբերող տարածման սակավության, առատության, մահաջնջման և փոփոխականության բոլոր երևույթներով հանդերձ մեզ կթվան մշուշում կամ մեր կողմից բոլորովին սխալ կերպով կհասկացվեն։ Բնությունը մենք պատկերացնում ենք ցնծալի, մենք հաճախ տեսնում ենք սննդի առատություն. մենք չենք տեսնում կամ մոռանում ենք, որ մեր շուրջը անհոգ կերպով դայլայլող թռչունները մեծ մասամբ սնվում են միջատներով ու սերմերով և այսպիսով շարունակ կյանք են ոչնչացնում, մենք մոռանում ենք, թե ինչպես այդ երգեցիկները կամ նրանց ձվերը իրենց հերթին կերակուր են դառնում գիշատիչ թռչուններին ու գազաններին։ Մենք ոչ միշտ ենք ուշադրության առնում այն, որ եթե որոշ պահին սնունդն առատությամբ է գտնվում, ապա այդ չի կարելի ասել ամեն տարվա և տարվա յուրաքանչյուր եղանակի մասին։
«ԳՈՅՈՒԹՅԱՆ ԿՌԻՎ» ՏԵՐՄԻՆԻ ԳՈՐԾԱԾՈՒՄՆ ԼԱՅՆ ԻՄԱՍՏՈՎ
Ես պետք է նախազգուշացնեմ, որ այդ արտահայտությունը գործ եմ ածում լայն և փոխաբերական իմաստով, սրա մեջ ներառելով մի էակի կախումը մյուսից, ինչպես նաև հասկանալով (որը ավելի ևս կարևոր է) ոչ միայն մի անհատի կյանքը, այլև նրա հաջողությունը հաջորդներով իրեն ապահովելու տեսակետից։ Canis սեռին պատկանող երկու կենդանիների մասին կարելի է միանգամայն ստույգ կերպով ասել, որ սովածության շրջանում նրանք պայքարում են իրար դեմ սննդի և ապրուստի համար։ Անապատի եզրին բուսող բույսի մասին ևս ասում են, որ նա պայքարում է չորության դեմ, թեև ավելի ճիշտ կլիներ ասել, որ նա կախված է խոնավությունից։ Այն բույսի մասին, որ ամեն տարի տալիս է հազարավոր սերմեր, որոնցից միջին հաշվով միայն մեկն է աճում, դառնում հասուն բույս, իսկապես կարելի է ասել, որ նա պայքարում է իրեն նմանների դեմ և ուրիշ բույսերի դեմ, որոնք արդեն ծածկել են գետնի նույն տեղամասը։ Ճագոմի կյանքը կախված է խնձորենուց և էլի մի քանի ծառատեսակներից, բայց բռնազբոսիկ բան կլիներ նրանց դեմ ճագոմի պայքարի մասին խոսելը միայն այն պատճառով, որ եթե այդ մակաբույծներից չափազանց շատ աճեն մի ծառի վրա, ծառը կտկարանա և կմեռնի։ Բայց այդ ճագոմի մի քանի սերմնաբույսերի մասին, որոնք աճում են միևնույն ճյուղի վրա, կարելի է միանգամայն ստույգ ասել, որ նրանք պայքարում են իրար դեմ։ Քանի որ ճագոմը տարածվում է թռչունների միջոցով, ապա նրա գոյությունը կախված է նրանցից և, արտահայտվելով փոխաբերական իմաստով, կարելի է ասել, նա պայքարում է ուրիշ պտղատու բույսերի դեմ նրանով, որ հրապուրում է թռչուններին ուտելու իր պտուղները և տարածելու իր սերմերը։ Այս բոլոր առումներով, որոնք փոխանցվում են մեկը մյուսի, ես հարմարության համար դիմում եմ «գոյության կռիվ» արտահայտությանը։
ԲԱԶՄԱՑՄԱՆ ԵՐԿՐԱՉԱՓԱԿԱՆ ՊՐՈԳՐԵՍԻԱՆ
Գոյության կռիվը անխուսափելիորեն բխում է այն արագ պրոգրեսիայից, որով բոլոր օրգանական էակները ձգտում են բազմանալ։ Յուրաքանչյուր էակ, որն իր կյանքի ընթացքում արտադրում է մի քանի ձու կամ սերմ, անխուսափելի կերպով պետք է ոչնչացման ենթարկվի իր կյանքի որևէ հասակում, տարվա որևէ եղանակին կամ, վերջապես, որևէ պատահական տարիներում, այլապես բազմացման երկրաչափական պրոգրեսիայի սկզբունքի հետևանքով, նրա թիվը կհասներ այնպիսի չափերի, որ աշխարհում ոչ մի երկիր չէր կարողանա կերակրել կամ տեղավորել նրա սերունդներին։ Ուրեմն՝ քանի որ ավելի շատ անհատներ են արտադրվում, քան կարող են ապրել, պետք է յուրաքանչյուր դեպքում տեղի ունենա գոյության կռիվ կամ միևնույն տեսակի անհատների միջև, կամ տարբեր տեսակների անհատների միջև, կամ կյանքի ֆիզիկական պայմանների դեմ։ Դա Մալթուսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 30] ուսմունքն է[39], որն ավելի ևս մեծ ուժով կիրառելի է ամբողջ կենդանական և բուսական աշխարհի վերաբերմամբ, որովհետև այստեղ չի կարող լինել ոչ սննդի արհեստական ավելացում, ոչ էլ ամուսնանալուց խոհեմաբար հրաժարում։ Եթե, կարող է պատահել, ներկայումս որոշ տեսակներ բազմանում են շատ թե քիչ արագ կերպով, ապա բոլոր տեսակները չեն կարող հասնել այդ բանին, որովհետև երկիրը նրանց չէր կարող պարփակել։
Չկա ոչ մի բացառություն այն կանոնից, որի համաձայն ամեն մի օրգանական էակ բնականորեն բազմանում է այնպիսի պրոգրեսիայով, որ եթե նա չենթարկվեր ոչնչացման, ապա մեկ զույգի սերունդները կծածկեին ամբողջ երկիրը։ Նույնիսկ դանդաղորեն բազմացող մարդը քսան և հինգ տարվա ընթացքում թվով կրկնապատկվում է, և այդպիսի պրոգրեսիայով ավելի պակաս քան հազար տարի հետո նրա սերունդների համար բառացիորեն կանգնելու տեղ չէր լինի։ Լիննեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 72] հաշվել է, որ եթե որևէ միամյա բույս առաջացներ միայն երկու սերմ (իսկ այդպիսի թույլ արտադրողականությամբ բույս գոյություն չունի), ապա քսան տարի հետո նրա սերունդները կհասնեին միլիոնի։ Համարվում է, որ մեզ հայտնի կենդանիների մեջ փիղը ամենից ավելի դանդաղ է բազմանում, և ես աշխատեցի հաշվել նրա բազմացման նվազագույն չափերը։ Նա սկսում է բազմանալ, ամենից ավելի հավանական է, երեսուն տարեկանից ոչ վաղ հասակում և մինչև իննսուն տարեկան հասակը տալիս է ոչ ավելի քան վեց ձագ, իսկ ապրում է մինչև հարյուր տարի. ընդունելով այդ թվերը, կստանանք այն, որ 740—750 տարվա ընթացքում մեկ զույգից կստացվեր տասնևիննը միլիոն փիղ։
Բայց մենք ունենք ավելի համոզեցուցիչ վկայություններ, քան միմիայն տեսական հաշիվները, այն է՝ որոշ կենդանիների զարմանալի արագ բազմացման բազմաթիվ դեպքերը բնական վիճակում, երբ պայմանները նրանց համար ինչ-որ պատճառով առանձնապես բարենպաստ են եղել իրար հաջորդող երկու կամ երեք տարիների ընթացքում։ Ավելի ևս զարմանալի են այն փաստերը, որոնք վերաբերում են աշխարհի զանազան երկրներում վայրենացած մեր ընտանի մի քանի կենդանիներին. այնպես դանդաղ բազմացող եղջերավոր անասունների և ձիերի Հարավային Ամերիկայում և ավելի ուշ Ավստրալիայում արագ բազմանալուն վերաբերող մատնանշումները եթե չհենվեին ամենաարժանահավատ վկայությունների վրա, ապա նրանք պարզապես անհավատալի կթվային։ Այդպես էլ բույսերի նկատմամբ. կարելի է օրինակներ բերել, երբ բերված բույսերը տասը տարուց ավելի քիչ ժամանակամիջոցում գրավել են ամբողջ կղզիների տարածությունը։ Մի քանի բույսեր, ինչպես, օրինակ, կարդոնները և մի բարձրահասակ տատասկափուշ, որոնք այժմ շատ սովորական են Լա-Պլատայի ընդարձակ հարթություններում և ծածկում են ամբողջ քառակուսի մղոններ, գրեթե դուրս մղելով այլ բույսերին, բերված են Եվրոպայից։ Իսկ այլ բույսեր, որոնք, ինչպես ինձ հաղորդել է դոկտոր Ֆոկոները,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 73] տարաբնակված են Հնդկաստանում Կամորինի Հրվանդանից մինչև Հիմալայան լեռները, բերված են Ամերիկայից՝ նրա հայտնագործումից հետո։ Այսպիսի դեպքերում, իսկ դրանց անվերջ ցուցակը կարելի էր ներկայացնել, ոչ ոք չի ենթադրի, որ կենդանիների կամ բույսերի պտղաբերությունը միայն ժամանակավոր և հանկարծակի կերպով այդքան զգալի աստիճանի է բարձրացել։ Ակնհայտ բացատրությունն այն է, որ կենսապայմանները եղել են չափազանց բարենպաստ, որի հետևանքով ծեր և մատղաշ անհատները ավելի քիչ են ոչնչացման ենթարկվել, և համարյա բոլոր երիտասարդ անհատները կարողացել են անարգել կերպով բազմանալ, նրանց բազմացման երկրաչափական պրոգրեսիան, որի հետևանքները մեզ միշտ զարմացնում են, շատ պարզ կերպով բացատրում է նրանց թվի արագ ավելացումը և լայն տարածումը նոր հայրենիքում։
Իր բնական վիճակում համարյա յուրաքանչյուր չափահաս բույս ամեն տարի տալիս է սերմեր, իսկ կենդանիների մեջ շատ քիչ կգտնվեն այնպիսիները, որոնք ամեն տարի չեն զուգավորվում։ Այստեղից էլ մենք համոզվածությամբ կարող ենք հաստատել, որ բոլոր կենդանիներն ու բույսերը ձգտում են բազմանալ երկրաչափական պրոգրեսիայով, որ նրանք կլցնեին բոլոր այն տեղերը, որտեղ միայն կկարողանան ապրել և որ երկրաչափական պրոգրեսիայով բազմանալու այդ ձգտումը պետք է սահմանափակման ենթարկվի միայն ոչնչացման միջոցով կյանքի որևէ շրջանում։ Ընտանի կենդանիների վրա կատարված սովորական դիտողությունները, ես կարծում եմ, հեշտությամբ կարող են մեզ թյուրիմացության մեջ գցել, որովհետև մենք չենք տեսնում, որ նրանք զգալի ոչնչացման ենթարկվեին, բայց միևնույն ժամանակ մոռանում ենք այն հազարավորների մասին, որոնք ամեն տարի մորթվում են մեր սննդի համար, և որ բնական դրության մեջ ևս այս կամ այլ կերպ նույն քանակով անխուսափելիորեն պիտի կոտորվեին։
Յուրաքանչյուր տարի հազարավոր ձվեր կամ սերմեր արտադրող և շատ քիչ դրանք արտադրող օրգանիզմների միջև միակ տարբերությունը այն է, որ վերջինները կյանքի նպաստավոր պայմաններում ամբողջ մի երկիր, որքան էլ կուզե նա մեծ լինի, լցնելու համար կպահանջեին միայն մի քանի լրացուցիչ տարիներ։ Կոնդորը երկու ձու է ածում, իսկ ջայլամը՝ քսան, բայց չնայած դրան, միևնույն երկրում նրանցից կոնդորը կարող է ավելի բազմաթիվ լինել։ Մրրկահավը ածում է ընդամենը մեկ ձու, բայց և այնպես կարծում են, որ նա ամենատարածված թռչունն է երկրի վրա։ Մի ճանճ հարյուրավոր ձվեր է ածում, իսկ մյուսը, ինչպես օրինակ, Hippobosca-ն՝ միայն մի հատ, բայց այս տարբերությունը չի որոշում յուրաքանչյուր տեսակի անհատների այն քանակը, որը կարող է իր մեջ տեղավորել երկիրը։ Ձվերի բազմաքանակությունը նշանակություն ունի այն տեսակների համար, որոնք կախում ունեն սննդի տատանվող քանակությունից, որովհետև դա է թույլ տալիս նրանց թվով ավելանալու։ Բայց ձվերի կամ սերմերի բազմաքանակության իսկական նշանակությունն այն է, որ կարողանա ծածկել նրանց զգալի պակասումը, որն առաջացնում է ոչնչացումը նրանց կյանքի որևէ շրջանում, իսկ այդ շրջանը մեծ մասամբ լինում է շատ վաղ հասակում։ Եթե կենդանին որևէ կերպով կարող է պաշտպանել իր ածած ձվերը կամ ձագերը, ապա նույնիսկ ծնվածների փոքրաթիվ լինելու դեպքում էլ կարող է պահպանվել միջին քանակությունը. բայց եթե մեծ քանակությամբ ձվեր կամ ձագեր են ենթարկվում ոչնչացման, այդ դեպքում պետք է շատ ծնվեն, այլապես այդ տեսակը կբնաջնջվի։ Միջին հաշվով հազար տարի ապրող որևէ ծառատեսակի թիվը կարող էր պահպանվել առանց փոփոխության, եթե հազար տարվա ընթացքում նա առաջ բերեր մեկական սերմ, միայն թե այդ սերմը երբեք չենթարկվեր ոչնչացման և նրա համար ապահովված լիներ ծլումը մի հարմար տեղում։ Նշանակում է, կենդանիների կամ բույսերի միջին թիվը բոլոր դեպքերում միայն անուղղակիորեն կախված է ձվերի կամ սերմերի թվից։
Բնությունը լավ դիտելիս մենք երբեք չպետք է աչքաթող անենք վերոհիշյալ նկատառումները, մենք չպետք է մոռանանք, որ յուրաքանչյուր օրգանական էակ, կարելի է ասել, լարում է իր ամբողջ ուժերը, որպեսզի ավելացնի իր թիվը, որ նրանցից ամեն մեկն ապրում է, միայն դիմանալով պայքարին իր կյանքի որևէ շրջանում, որ անողոք բնաջնջումը անխուսափելիորեն տարածվում է ծերի կամ երիտասարդի վրա ամեն մի սերնդում կամ կրկնվող ընդմիջումներով։ Նվազեցրեք խոչընդոտները, պակասեցրեք ոչնչացումը, թեկուզ և ամենափոքր չափերով, և տեսակի թիվը գրեթե վայրկենապես կաճի որևէ չափով։
ԲԱԶՄԱՑՈՒՄԸ ԿԱՍԵՑՆՈՂ ԽՈՉԸՆԴՈՏՆԵՐԻ ԲՆՈՒՅԹԸ
Այն պատճառները, որոնք կասեցնում են յուրաքանչյուր տեսակի բազմացման բնական ձգտումը, չափազանց մութն են։ Նայեցեք ամենահզոր տեսակին, ինչքան նա իր բազմաթիվ ներկայացուցիչներով լեփ-լեցուն է, այնքան նա ձգտում է ավելի ևս շատ բազմանալու։ Ոչ մի դեպքի վերաբերմամբ մեզ հայտնի չէ, թե ըստ էության ինչումն են կայանում բազմացման խոչընդոտները։ Եվ դա բնավ զարմանալի չէ, եթե մենք հիշենք, թե մեզ այդ ուղղությամբ ինչքան քիչ բան է հայտնի նույնիսկ մարդու վերաբերմամբ, որին մենք անհամեմատ ավելի լավ գիտենք, քան որևէ այլ կենդանու։ Բազմացման խոչընդոտների հարցը մեծ հմտությամբ մշակել են մի քանի հեղինակներ, և ես հույս ունեմ իմ ապագա մի աշխատության մեջ հանգամանորեն քննարկել այն՝ նկատի ունենալով գլխավորապես Հարավային Ամերիկայի գիշատիչ կենդանիներին։ Այստեղ ես կբավարարվեմ մի քանի դիտողություններով միայն նրա համար, որպեսզի ընթերցողների ուշադրությունը սևեռեմ հիմնական կետերից մի քանիսի վրա։ Ձվերը կամ շատ մատղաշ կենդանիները սովորաբար ամենից շատ են տուժում. բայց այդ կանոնը ունի նաև բացառություններ։ Բույսերի սերմերը ոչնչանում են լայն չափերով, բայց իմ կատարած դիտումների հիման վրա պարզվում է, որ ծլած սերմերն ամենից ավելի հաճախ ոչնչանում են նրանից, որ ծլել են ուրիշ բույսերով արդեն խիտ ծածկված հողի վրա։ Մեծ քանակությամբ ծիլեր ոչնչացվում են նաև զանազան թշնամիների կողմից։ Այսպես, լավ փխրեցրած և մաքրված երեք ոտնաչափ երկարությամբ և երկու ոտնաչափ լայնությամբ հողակտորի վրա, որտեղ նորածիլ բույսերին չէին կարող խեղդել մյուս բույսերը, ես հաշվեցի մեր տեղական մոլախոտերի ծիլերը, և դուրս եկավ, որ 357 ծլածներից 295-ը ոչնչացրել են գլխավորապես կակղամորթները և միջատները։ Եթե թողնենք աճելու այն մարգագետինը, որը երկար հնձվել է կամ չորքոտանիների կողմից տակից է արածացվել, այդ դեպքում ավելի ուժեղ բույսերը հետզհետե կոչնչացնեն ավելի թույլ, թեև միանգամայն զարգացած բույսերին, այսպես օրինակ, շարունակ հնձվող գազոնի մի փոքրիկ (երեք ոտնաչափ լայնքը, չորս՝ երկայնքը) հողաբաժնյակի վրա աճած քսան տեսակներից իննը տեսակը ոչնչացան միայն այն պատճառով, որ մյուսներին հնարավորություն էր տրվել ազատ աճելու։
Յուրաքանչյուր տեսակի համար անհրաժեշտ սննդի քանակը անշուշտ որոշում է նրա բազմացման ծայր աստիճանը. բայց հաճախ տեսակի միջին թիվը կախված է ոչ թե նրա հայթայթած կերից, այլ նրանից, որ տեսակն ինքը այլ կենդանիների համար կերակուր է ծառայում։ Այսպես, հազիվ կասկածի ենթակա է այն, որ կաքավների, մայրահավերի և նապաստակների քանակությունը որևէ մեծ կալվածքի սահմաններում կախված է գլխավորապես նրանց այն ոչնչացումից, որ պատճառում են այդտեղի մանր գիշատիչները։ Եթե քսան տարի Անգլիայում չսպանվեր ոչ մի որսակենդանի, բայց և միևնույն ժամանակում չոչնչացվեին նաև մանր գիշատիչները, ապա ամենայն հավանականությամբ հանրագումարում կստացվեր ավելի քիչ որսակենդանի, քան այժմ կա, չնայած այն բանին, որ այժմ ամեն տարի հազարավոր որսակենդանիներ են սպանվում։ Մյուս կողմից, որոշ դեպքերում, ինչպես, օրինակ, փղերի վերաբերմամբ, ոչ մի անհատ չի ոչնչանում գիշատիչների կողմից, որովհետև Հնդկաստանում նույնիսկ վագրը միայն շատ հազվագյուտ դեպքում համարձակվում է հարձակվել երիտասարդ փղի վրա, որին պաշտպանում է նրա մայրը։
Տեսակների միջին թվի որոշման գործում կարևոր դեր են կատարում կլիման և շատ ցած ջերմաստիճանի կամ երաշտի ժամանակաշրջանները, որոնք, ըստ երևույթին, բազմացման ամենաներգործող արգելակներն են։ Ես մոտավորապես հաշվել եմ (գլխավորապես գարնանը թռչնաբների թվի նվազման վրա հիմնվելով), որ իմ կալվածքում 1854/55 թ. ձմեռը ոչնչացրեց թռչունների չորս հինգերորդ մասը և դա իսկապես սոսկալի ոչնչացում է, եթե միայն հիշենք, որ մարդուն այցելող համաճարակների վերաբերմամբ 10% մահացությունը անսովոր բարձր է համարվում։ Առաջին հայացքից կլիմայի ազդեցությունը կարող է մեզ թվալ գոյության կռվից միանգամայն անկախ, բայց այն պատճառով, որ կլիման գլխավորապես ազդում է սննդի կրճատման վրա, նա առաջ է բերում ամենադաժան պայքար թե մի տեսակի և թե տարբեր տեսակների անհատների միջև, որոնք սնվում են միևնույն սննդով։ Նույնիսկ այն դեպքերում էլ, երբ կլիմայական պայմանները, ինչպես, օրինակ, խիստ ցուրտը, ազդում է անմիջականորեն, ամենից ավելի տուժում են ամենաթույլ անհատները կամ նրանք, որոնք ձմեռվա ընթացքում ավելի քիչ սնունդ են հայթայթել։ Երբ ճանապարհորդելիս տեղափոխվում ենք հարավից դեպի հյուսիս կամ խոնավ երկրից դեպի չորային երկիր, մենք անպայման նկատում ենք, որ որոշ տեսակներ հետզհետե սակավանում և, վերջապես, անհետանում են, իսկ քանի որ կլիմայի փոփոխությունը աչքի է զարնում, ուստի մենք հակվում ենք այդ ամենը վերագրելու կլիմայի անմիջական ներգործությանը։ Սակայն այդ տեսակետը բոլորովին սխալ է. մենք մոռանում ենք, որ յուրաքանչյուր տեսակ նույնիսկ այնտեղ, որտեղ նա տիրապետում է, իր գոյության որևէ շրջանում շարունակում է ենթարկվել ահագին ոչնչացման թշնամիների կողմից կամ նույն տեղի ու սննդի համար իր դեմ մրցողների կողմից, իսկ եթե կլիմայում մի աննշան տարբերություն նպաստավոր կլինի այդ թշնամիների կամ մրցակիցների համար, այդ դեպքում նրանք կսկսեն բազմանալ, բայց քանի որ ամեն մի վայր արդեն բնակիչներով լեփ-լեցուն է, ուրեմն մյուս տեսակների թիվը պիտի պակասի։ Ճանապարհորդելով դեպի հարավ և տեսնելով, որ որևէ տեսակ թվով պակասում է, մենք կարող ենք վստահ լինել, որ դրա պատճառը նույնքան նրանումն է, որ պայմանները բարենպաստ են մյուս տեսակների համար, որքան նաև նրանումն է, որ նրանք վնասում են պակասող տեսակին։ Նույնն է նկատվում նաև այն ժամանակ, երբ մենք գնում ենք դեպի հյուսիս, թեև մի քիչ ավելի փոքր չափով, որովհետև տեսակների թիվն ընդհանրապես և հետևաբար նաև մրցության մեջ գտնվողների թիվը, հյուսիսային ուղղությամբ գնալով պակասում է։ Այդ պատճառով, երբ շարժվում ենք դեպի հյուսիս կամ բարձրանում ենք լեռները, մենք ավելի հաճախ ենք հանդիպում վտիտ ու գետնակիպ ձևերի, որոնք առաջացել են կլիմայի վնասակար ազդեցությունից անմիջականորեն, քան երբ գնում ենք դեպի հարավ կամ իջնում ենք սարերից։ Իսկ երբ հասնում ենք բևեռային երկրներին կամ ձյունապատ լեռնակատարներին, կամ կատարյալ անապատի, մենք հանդիպում ենք բացառապես տարերքի դեմ մղվող գոյության կռվի։
Որ կլիման ներգործում է գլխավորապես անուղղակի կերպով՝ նպաստելով մյուս տեսակներին, մենք պարզ կերպով տեսնում ենք այն փաստից, որ հսկայական թվով բույսեր, որոնք հիանալի դիմանում են կլիմային, մեր պարտեզներում նատուրալիզացիայի չեն ենթարկվում, որովհետև չեն կարող մրցել մեր տեղական բույսերի հետ կամ դիմանալ տեղական կենդանիների առաջացած բնաջնջմանը։
Երբ որևէ տեսակ առանձնապես բարենպաստ հանգամանքների շնորհիվ անչափորեն բազմանում է մի սահմանափակ վայրում, ապա շատ հաճախ երևան են գալիս համաճարակներ, համենայն դեպս այդ բանը սովորաբար պատահում է որսակենդանիների հետ մեր անտառներում, այստեղ մենք ունենք բազմացման համար մի խոչընդոտ, որն անկախ է գոյության կռվից։ Սակայն այսպես կոչված համաճարակներից մի քանիսն, ըստ երևույթին, կախված են մակաբույծ որդերից, որոնց բազմացմանը նպաստել է մի որևէ պատճառ, մասամբ թերևս կուտակված կենդանիների մեջ ավելի հեշտ կերպով տարածվելը. այսպիսով, այստեղ էլ նկատվում է մակաբույծի և նրա զոհի միջև կռվի նման բան։
Մյուս կողմից շատ դեպքերում միևնույն տեսակին պատկանող անհատների թվի մեծությունը, համեմատած թշնամիների թվի հետ, անհրաժեշտ պայման է հանդիսանում տեսակի պահպանման համար։ Այսպես, մեզ հաջողվում է մեր դաշտերից առատությամբ հավաքել հացահատիկների սերմեր, հլածուկի կամ այլ բույսերի սերմեր այն պատճառով, որ նրանցով սնվող թռչունների թվի համեմատությամբ նրանք լինում են հսկայական առատությամբ. իսկ մյուս կողմից, թռչուններն էլ, թեև տարվա այդ եղանակին առատ սնունդ են գտնում, չեն կարող սերմերի եղած քանակության համեմատ բազմանալ, որովհետև նրանց բազմացմանը արգելք է լինում ձմեռը։ Բայց ամեն ոք, ով փորձել է, գիտե այն, թե ինչքան դժվար է ցորենի կամ պարտեզում բուսնող որևէ այլ բույսի մի քանի օրինակներից սերմեր հավաքելը. ինձ վիճակվել է այդպիսի դեպքերում կորցնել մինչև վերջինը։ Տեսակի պահպանության համար անհատների թվի մեծության այդ անհրաժեշտության տեսակետից, իմ կարծիքով, հասկանալի են դառնում շատ երևույթներ բնության մեջ, ինչպես օրինակ, սակավաթիվ վայրերում հազվագյուտ բույսերի առատությամբ գտնվելու փաստը, կամ այն փաստը, որ սոցիալական բույսերը մնում են սոցիալական, այսինքն անհատներով առատ են լինում նույնիսկ իրենց տարաբնակման ծայրային սահմաններում։ Այդպիսի դեպքերում մենք պետք է ընդունենք, որ բույսը կարող էր պահպանվել միայն այնտեղ, որտեղ պայմաններն այնքան բարենպաստ էին, որ թույլ էին տալիս բազմաթիվ անհատների համատեղ գոյություն և դրանով իսկ կարող էին տեսակը փրկել վերջնական մահաջնջումից։ Ես պետք է ավելացնեմ, որ խաչաձևման բարեբար ազդեցությունը և ազգակցության մոտիկ աստիճաններով բազմանալու վնասակար հետևանքները նույնպես դեր են խաղացել նման շատ դեպքերում. բայց այստեղ այդ խնդրի վրա երկար չեմ խոսի։
ԲՈԼՈՐ ԿԵՆԴԱՆԻՆԵՐԻ ԵՎ ԲՈՒՅՍԵՐԻ ՄԻՋԵՎ ԲԱՐԴ ՀԱՐԱԲԵՐՈՒԹՅՈՒՆՆԵՐԸ ԳՈՅՈՒԹՅԱՆ ԿՌՎՈՒՄ
Հայտնի են շատ դեպքեր, որոնք ցույց են տալիս, թե ինչքան բարդ ու անակնկալ են խոչընդոտները և փոխհարաբերությունները օրգանական էակների միջև, որոնք գոյության կռիվ են մղում միևնույն երկրում։ Կբերեմ միայն մեկ օրինակ, որը պարզ է, բայց ինձ խիստ հետաքրքրել է։ Ստաֆֆորդշիրում, իմ մի ազգականի կալվածքում, որտեղ ես հետազոտություններ կատարելու համար ունեի բոլոր հարմարությունները, գտնվում էր հավամրգու մի ընդարձակ և չափազանց անբերրի հարթություն, որին մարդկային ձեռք երբեք չէր դիպել, բայց բոլորովին նույնպիսի հարթությունից մի քանի հարյուր ակր տարածություն քսան և հինգ տարի առաջ ցանկապատել էին ու այնտեղ տնկել շոտլանդական սոճիներ։ Տնկիներ ունեցող մասի բնական բուսականության մեջ կատարված փոփոխությունը զարմանալի էր և գերազանցում էր այն տարբերությանը, որը սովորաբար նկատվում է մի հողատեսակից մի բոլորովին այլ հողատեսակի անցնելիս, ոչ միայն բոլորովին փոխվել էր հավամրգային ֆորմացիան բնորոշող զանազան տեսակներին պատկանող բույսերի հարաբերակցությունը, այլև երևան էին եկել տասներկու նոր տեսակներ (չհաշված հացազգիները և բոշխերը), որոնք չէին հանդիպում հավամրգուտի մնացած մասում։ Այդ փոփոխությունը միջատների վրա ավելի էական ազդեցություն էր գործել, որովհետև սոճու տնկարկում երևացել էին վեց տեսակի միջատակեր թռչուններ, որոնք չեն հանդիպում հարթության մնացած մասում, որտեղ լինում էին միջատակեր թռչունների երկու կամ երեք տեսակներ։ Մենք տեսնում ենք, թե որքան հզոր է միայն մեկ ծառատեսակի տնկելու ազդեցությունը, քանի որ ուրիշ ոչինչ չէր արված, բացի նրանից, որ այդ տեղը ցանկապատվել էր անասունների արածավերումից պաշտպանելու համար։ Բայց թե ցանկապատումն ինչպիսի կարևոր դեր կարող է խաղալ, ես դրանում պարզորոշ կերպով համոզվեցի Ֆարնեմի մոտերքը՝ Սորրեյում։ Այնտեղ հանդիպում են ընդարձակ հավամրգուտներ՝ շոտլանդական սոճիների փոքրաթիվ խմբերով, որոնք ցրված են բլուրների գագաթների վրա. վերջին տասնամյակում մեծ տարածություններ ցանկապատվել են, և ինքնացան սոճիներն այնքան խիտ են բուսել, որ իրենք իրենց խեղդում են։ Երբ ես ստույգ իմացա, որ ոչ ցանք է եղել, ոչ էլ ծառեր են տնկվել, այնպես զարմացել էի նրանց բազմաքանակության վրա, որ բարձրացա մի քանի բարձրադիր կետերի վրա, որտեղից կարող էի տեսնել չցանկապատված հարթության հարյուրավոր ակրերը, և բառացի կերպով չտեսա ոչ մի ծառ, բացի այն սոճիներից, որոնք տնկված էին բլուրների վրա։ Բայց մտիկ տալով, հավամրգու ցողունների միջև ես գտա բազմաթիվ սերմնատունկեր և մատղաշ ծառեր, որոնց կրծոտել էին անասունները։ Հին ծառերի խմբից մոտավորապես հարյուր յարդ հեռավորության վրա մեկ քառակուսի յարդում ես հաշվեցի երեսուներկու ծառ. նրանցից մեկն ուներ քսան վեց տարեկան շերտեր, այդ երկար տարիների ընթացքում նա ապարդյուն կերպով աշխատել է իր գլուխը շրջակա հավամրգուց վեր բարձրացնել։ Զարմանալի չէ, որ հողակտորը ցանկապատելուն պես նա ծածկվել է խիտ բուսած մատղաշ սոճիներով։ Բայց այդ հավամրգուտները այնքան ընդարձակ էին և այնքան անբերրի, որ ոչ ոքի մտքով չէր անցնի, թե անասունները կարող էին այդքան ջանադիր կերպով ու մինչև տակը արածավերել նրանց։
Այստեղ մենք տեսանք, որ շոտլանդական սոճու գոյությունը կախված էր անասուններից. բայց աշխարհի որոշ մասերում անասունների գոյությունը կախված է միջատների ներկայությունից։ Թերևս այդ տեսակետից ապշեցուցիչ օրինակ է ներկայացնում Պարագվան. այնտեղ չեն վայրենացել ոչ ձիերը, ոչ տավարը, ոչ էլ շները, թեև նրանից ավելի հարավ և հյուսիս նրանք վխտում են վայրենացած վիճակում։ Թե Ազարան[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 74] և թե Ռենգերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 75] ցույց են տվել, որ այդ կախված է Պարագվայում մեծ քանակությամբ հանդիպող որոշ տեսակի ճանճից, որն իր ձվերը դնում է նորածին կենդանիների պորտում։ Այդ ճանճը, ինչքան էլ որ նա բազմաքանակ է, չի կարողանում էլ ավելի բազմանալ գուցե մի խոչընդոտի, հավանորեն այլ մակաբույծ միջատների պատճառով։ Հետևաբար, եթե Պարագվայում միջատակեր թռչունների թիվը պակասեր, այդ դեպքում մակաբույծ միջատները կբազմանային, իսկ այդ կպակասեցներ պորտի մեջ ձու դնող ճանճերի թիվը. այն ժամանակ տավարը և ձիերը կվայրենանային, իսկ դա զգալի լափով կփոխեր (ինչպես ես նկատել եմ Հարավային Ամերիկայի մի քանի մասերում) բուսականության բնույթը, որը դարձյալ կանդրադառնար միջատների վրա, իսկ վերջիններս իրենց հերթին, ինչպես մենք հենց նոր տեսանք Ստաֆֆորդշիրում, կազդեին միջատակեր թռչունների վրա և այսպես գնալով ավելի հեռուները ավելի ու ավելի ընդարձակվող բարդ կերպով խառնահյուսվող շրջաններով։ Սակայն չպետք է կարծել, որ բնության մեջ էակների փոխհարաբերությունները երբևիցե այդքան պարզ են եղել, ինչպես այս օրինակում։ Մարտերն իրար հաջորդում են շարունակ փոփոխական հաջողությամբ և այնուամենայնիվ ստացված երկար հանրագումարում ուժերն այնպես նուրբ կերպով են հավասարակշռված, որ բնության արտաքին տեսքը երկար ժամանակաշրջանների ընթացքում մնում է անփոփոխ, թեև ամենաչնչին հանգամանքը կարող է պայմանավորել մի օրգանիզմի հաղթանակը մյուսի վրա։ Բայց և այնպես մեր անգիտությունը այնքան խորն է, իսկ մեր ինքնավստահությունը այնքան մեծ, որ մենք զարմանում ենք, երբ լսում ենք որևէ օրգանական էակի մահաջնջման մասին և, չհասկանալով դրա պատճառը, մոգոնում ենք ինչ որ կատակլիզմներ, որոնք ամայացնում են ամբողջ աշխարհը, կամ հորինում ենք ինչ որ օրենքներ, որոնք սահմանափակում են օրգանական էակների երկարատևությանը։ Չեմ կարող զսպել գայթակղությունս բերելու էլի մի օրինակ, որը ցույց է տալիս, թե ինչպես բույսերն ու կենդանիները, որոնք տեղավորված են օրգանական սանդուխքի իրարից հեռու գտնվող աստիճանների վրա, սերտորեն շրջահյուսված են լինում բարդ փոխհարաբերությունների ցանցով։ Ես առիթ կունենամ ցույց տալու, որ էկզոտիկ Lobelia fulgens-ին իմ պարտեզում միջատները երբեք չեն այցելում և այդ պատճառով, նրա հատուկ կառուցվածքի շնորհիվ նա երբեք սերմեր չի տալիս։ Գրեթե մեր բոլոր խոլորձիազգի բույսերն անպայման այն միջատների այցելության կարիք ունեն, որոնք տեղափոխում են նրանց ծաղկափոշին և այսպիսով նրանց փոշոտում են։ Իմ կատարած փորձերի հիման վրա ես համոզվել եմ, որ եռագույն մանուշակի (Viola tricolor) բեղմնավորման համար իշամեղուները գրեթե անհրաժեշտ են, որովհետև մյուս մեղուները այդ ծաղիկներին չեն այցելում։ Ես գտել եմ նաև, որ մեղուների այցելությունը անհրաժեշտ է երեքնուկի մի քանի տեսակների բեղմնավորման համար. այսպես, օրինակ, սպիտակ երեքնուկի (Trifolium repens) քսան գլխիկները տվել են 2290 սերմ, մինչդեռ մեղուների այցելությունից պաշտպանված մյուս քսան գլխիկները չեն տվել ոչ մի հատ սերմ։ Մի ուրիշ փորձում կարմիր երեքնուկի (T. pratense) հարյուր գլխիկը ավել է 2700 սերմ, իսկ մեղուների այցելությունից պաշտպանված նույնքան գլխիկները՝ ոչ մի հատ։ Միայն իշամեղուներն են այցելում կարմիր երեքնուկին, որովհետև մյուս մեղուները չեն կարողանում հասնել նրա նեկտարին։ Ենթադրություն է արվել, թե երեքնուկը կարող է բեղմնավորվել ցեցազգի միջատների միջոցով, բայց ես կասկածում եմ այն բանում, որ նրանք կարողանային այդ անել կարմիր երեքնուկի վերաբերմամբ,— նրանց մարմնի կշիռը չի բավականանա, որպեսզի նրանք կարողանային ճնշել այդ ծաղկի կողքային պսակաթերթիկները, այսպես կոչված, թևերը։ Այստեղից մենք իրավունք ունենք եզրակացնելով որ եթե իշամեղուների ամբողջ ցեղը մահաջնջվի կամ շատ հազվագյուտ դառնա Անգլիայում, այդ դեպքում եռագույն մանուշակն ու կարմիր երեքնուկը նույնպես կդառնային հազվագյուտ կամ իսպառ կանհետանային։ Իշամեղուների թիվը մի երկրում զգալի չափով կախված է դաշտային մկների քանակությունից, որոնք ոչնչացնում են նրանց բները և խորիսխները. գնդապետ Նյումանը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 76] որ երկար ժամանակ ուսումնասիրել է իշամեղուների կյանքը, կարծում է, որ Անգլիայում նրանց երկու երրորդ մասը ոչնչանում է այդ ձևով։ Բայց մկների թիվը, ինչպես ամեն ոք գիտե, մեծ չափով կախված է կատուների թվից, և գնդապետ Նյումանը ասում է. «Գյուղերի և փոքրիկ քաղաքների մոտակայքում ես ավելի մեծ քանակությամբ եմ հանդիպել իշամեղուների բների, քան այլ տեղերում, և այդ վերագրում եմ կատուների ներկայությանը, որոնք մկներին որսում են»։ Այստեղից միանգամայն հավանական է դառնում, որ մեծ քանակությամբ կատվացեղ կենդանիների ներկայությունը մի որոշ վայրում որոշում է նախ, մկների, և ապա իշամեղուների միջոցով բույսերի որոշ ծաղիկների առատությունը այդ վայրում։
Յուրաքանչյուր տեսակի վրա հավանորեն անդրադառնում է ամենաբազմազան խոչընդոտների ազդեցությունը, որոնք ներգործում են կյանքի տարբեր շրջաններում, տարվա տարբեր եղանակներին կամ տարբեր տարիներում, և թեև նրանցից մեկը կամ մի քանիսը կարող են ավելի զորավոր լինել, այնուամենայնիվ տեսակի միջին թիվը կամ նույնիսկ գոյությունը կախված կլինի նրանց միացած ներգործությունից։ Որոշ դեպքերում կարող է թվալ, թե միմյանց բոլորովին չնմանվող խոչընդոտները ազդում են միևնույն տեսակի բազմացման վրա տարբեր երկրներում։ Երբ մենք նայում ենք խիտ բուսակալած ջրափին խռնված խոտերի և թփուտների վրա, մենք այդ բուսականության կազմը և նրա ներկայացուցիչների հարաբերական թիվը հակամիտ ենք վերագրելու, այսպես ասած, պատահականության։ Բայց որքա՜ն թյուր է այդ կարծիքը։ Ամեն ոք, իհարկե, լսել է, որ երբ Ամերիկայում կտրատում են անտառը, նրան փոխարինում է բոլորովին այլ բուսականություն, բայց նկատված է, որ Հյուսիսային Ամերիկայի Հարավային Նահանգներում հին հնդկական ավերակները, որոնց վրա միաժամանակ, իհարկե, ոչ մի ծառ չի բուսել, այժմ ծածկված են այնպիսի բուսականությամբ, որը ներկայացնում է նույն սքանչելի բազմազանությունը և նույն թվական հարաբերությունը տեսակների միջև, ինչ որ նկատվում է շրջապատող կուսական անտառում։ Ինչպիսի կռիվ պիտի շարունակվեր դարերի ընթացքում զանազան ծառատեսակների միջև, որոնք հազարներով շաղ էին տալիս իրենց սերմերը, ինչպիսի կռիվ պիտի մոլեգներ միջատների, խխունջների և այլ կենդանիների միջև, թռչունների և գիշատիչ գազանների միջև, որոնք միանման կերպով ձգտում են բազմանալ, սնվում են մեկը մյուսի հաշվին կամ ծառերի, նրանց սերմերի ու ծիլերի կամ այն մյուս բույսերի հաշվին, որոնք սկզբում ծածկել էին հողը և խեղդում ծառերի աճեցողությունը։ Վեր նետեք մի բուռ փետուրներ օդի մեջ, և նրանցից ամեն մեկը գետին կընկնի միանգամայն որոշակի օրենքների համաձայն. բայց ինչքան հասարակ խնդիր է որոշել, թե ուր կընկնի յուրաքանչյուր փետուրիկը՝ համեմատած անհամար բույսերի և կենդանիների բոլոր այն ազդեցություններին և հակազդեցություններին հետամտնելու խնդրի հետ, որոնք դարերի ընթացքում որոշել են բուսականության կազմը և ծառատեսակների հարաբերական թիվը հին հնդկական ավերակների վրա։
Մի օրգանիզմի կախումը մյուսից, օրինակ, մակաբույծի կախումը իր զոհից, սովորաբար կապում է միմյանց հետ այնպիսի էակներ, որոնք օրգանական սանդուխքի միմյանցից հեռու աստիճանների վրա են գտնվում։ Նույնը կարելի է ասել երբեմն նաև այն օրգանիզմների մասին, որոնք խիստ իմաստով պայքարում են գոյության համար, ինչպես, օրինակ, մորեխի և խոտակեր չորքոտանի կենդանիների միջև եղած մրցակցությունը։ Բայց կռիվը անպայման ամենից ավելի կատաղի կլինի նույն տեսակի ներկայացուցիչների միջև, որովհետև նրանք ապրում են միևնույն վայրում, միևնույն սննդի կարիք ունեն և ենթարկվում են միևնույն վտանգներին։ Միևնույն տեսակի տարատեսակների միջև կռիվը կլինի համարյա նույնքան սրված, և մենք երբեմն տեսնում ենք, որ նրա վախճանը որոշվում է շատ արագ կերպով. այսպես, օրինակ, եթե ցորենի մի քանի տարատեսակներ ցանենք միասին և ստացված խառը սերմերը նորից ցանենք, այդ դեպքում որոշ տարատեսակներ, որոնք ավելի համապատասխանում են կլիմային ու հողին կամ ավելի պտղաբեր են, կտան ավելի շատ սերմ և, այսպիսով, մի քանի տարում դուրս կմղեն մնացած տարատեսակներին։ Խառնուրդը կայուն հարաբերության մեջ պահպանելու համար, երբ խառնուրդը կազմված է նույնիսկ այնպիսի միմյանց մոտիկ տարատեսակներից, ինչպիսիք են զանազան երանգի հոտավետ ոլոռները, անհրաժեշտ է սերմերը հավաքել առանձին-առանձին և խառնել հարկ եղած հարաբերությամբ, այլապես ավելի թույլ տարատեսակների թիվը հետզհետե կպակասի և, վերջապես, նրանք իսպառ կանհետանան։ Նույնն է նաև ոչխարի տարատեսակների նկատմամբ. պնդում են, որ ոչխարների որոշ լեռնային տարատեսակներ դուրս են մղում մյուսներին, այնպես որ վերջիններս սոված են մնում, և նրանց չի կարելի միասին պահել։ Նույնպիսի հետևանք է ստացվել բժշկական տզրուկների մի քանի տարատեսակները միասին բազմացնելիս։ Կարելի է մինչև անգամ կասկածել, թե արդյոք մեր ընտանի կենդանիների և մշակովի բույսերի տարատեսակները օժտված են այնքան միանման ուժով, սովորություններով և կացույթով, որպեսզի նրանց միջև եղած սկզբնական հարաբերությունը (իհարկե խաչաձևման բացակայության դեպքում) կարողանար պահպանվել կես դյուժին սերունդների ընթացքում, եթե նրանց թողնվեր նույնպիսի կռիվ վարելու միմյանց դեմ, ինչպիսին վարում են էակները բնական վիճակում, և եթե նրանց սերմերը կամ ձագերը չպահպանվեին ամեն տարի հարկ եղած թվական հարաբերության մեջ։
ԳՈՅՈՒԹՅԱՆ ԿՌԻՎԸ ԱՌԱՆՁՆԱՊԵՍ ՀԱՄԱՌ է, ԵՐԲ ԱՅՆ ՏԵՂԻ է ՈՒՆԵՆՈՒՄ ՄԻԵՎՆՈՒՅՆ ՏԵՍԱԿԻ ԱՆՀԱՏՆԵՐԻ ԵՎ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ՄԻՋԵՎ
Քանի որ միևնույն սեռի տեսակները սովորաբար, թեպետև բնավ ոչ միշտ, իրենց սովորություններով ու կացույթով նման են և միշտ նման են իրենց կառուցվածքով, ուստի, ընդհանուր առմամբ, կռիվը նրանց միջև, եթե միայն նրանք մտնում են մրցության մեջ, սովորաբար լինում է ավելի դաժան, քան տարբեր սեռերի տեսակների միջև։ Այդ տեսնում ենք, օրինակ, Միացյալ Նահանգների որոշ մասերում վերջերս ծիծեռնակի մի տեսակի տարածվելուց, որին ուղեկցել է ծիծեռնակի մի այլ տեսակի չքացումը։ Վերջերս Շոտլանդիայի մի քանի մասերում կեռնեխի մի տեսակի (missel-thrush) բազմանալը առաջ է բերել մի այլ տեսակի՝ երգեցիկ կեռնեխի (song-thrush) թվի նվազում։ Ինչքան հաճախ առիթ է լինում լսելու, որ մեծամկան մի տեսակ դուրս է մղում մյուսին ամենաբազմազան կլիմայական պայմաններում։ Ռուսաստանում աշխետ ուտիճը ամենուրեք դուրս է մղում խոշոր սև ուտիճին։ Ավստրալիայում ներմուծված սովորական մեղուն երկրում արագ կերպով դուրս է մղում տեղական փոքրիկ, խայթոցից զուրկ մեղվին։ Դաշտային մանանեխի մի տեսակը դուրս է մղում մյուսին և այլն։ Մենք աղոտ կերպով ենք ըմբռնում, թե ինչու մրցությունը պետք է ամենից ավելի դաժան լինի այնպիսի մերձավոր ձևերի միջև, որոնք գրեթե միևնույն տեղն են գրավում բնության տնտեսության մեջ. բայց ամենայն հավանականությամբ մենք ոչ մի դեպքում չէինք կարողանա ճշտությամբ որոշել, թե ինչու հատկապես մի տեսակը մյուսի նկատմամբ հաղթող է դուրս գալիս մեծ կենսակռվում։
Բերված դիտողություններից կարելի է շատ կարևոր եզրակացություն անել, այն է, որ յուրաքանչյուր օրգանական էակի կառուցվածքը ամենաէական ձևով, թեպետ երբեմն էլ թաքուն ձևով, կապված է մյուս բոլոր օրգանական էակների հետ, որոնց հետ նա մրցության մեջ է մտնում սննդի կամ բնակատեղի համար, որոնցով նա սնվում է, կամ որոնցից ինքն է փրկություն որոնում։ Դա ակներևորեն արտահայտվում է ինչպես վագրի ատամների և ճիրանների մեջ, այնպես էլ այդ վագրի մազերին կպչող մակաբույծի կառչուն վերջավորությունների մեջ։ Բայց խտուտիկի հիանալի մազապատված թռչող սերմի վրա և ջրաբզեզի տափակ ու եզրազարդ ոտի վրա առաջին հայացքից նկատվում է միայն նրանց առնչությունը օդային կամ ջրային միջավայրի հետ։ Սակայն թռչող սերմերի առավելությունը ակներևորեն հարընչության մեջ է գտնվում այլ բույսերով տվյալ երկրի բնակվածության խտության հետ. թռիչքաթևերի շնորհիվ սերմերը կարող են հեռու տարածվել և ընկնել դեռ չզբաղեցված հողի վրա։ Ջրաբզեզի ոտները, որ հարմարված են սուզվելու համար, նրան թույլ են տալիս մրցելու ջրային այլ միջատների հետ, հետապնդելու իր որսին և ինքն էլ որս չդառնա այլ կենդանիների համար։
Շատ բույսերի սերմերում եղած սննդապաշարը առաջին հայացքից կարծես առնչություն չունի այլ բույսերի հետ։ Բայց ոլոռի կամ բակլայի նման և բարձր խոտերի մեջ ցանված սերմերից առաջացած մատղաշ բույսերի արագ աճեցողության հիման վրա կարելի է ենթադրել, որ սերմի մեջ եղած այդ սննդապաշարների գլխավոր նշանակությունը նրանումն է, որպեսզի նպաստեն սերմնաբույսերի աճեցողությանը, քանի դեռ նրանք ստիպված են պայքարել իրենց շրջապատող ու արագ զարգացող բուսականության դեմ։
Դիտեցեք բույսը նրա տարածման բնական բնագավառում, թե ինչու նա չի կրկնապատկում, չի քառապատկում իր անհատների թիվը։ Մենք գիտենք, որ նա կարող է դիմանալ քիչ ավելի տաքության կամ ցրտության, չորության կամ խոնավության, որովհետև իր տարածման սահմանագծերում նա մտնում է ավելի տաք կամ ցուրտ, ավելի չոր կամ խոնավ շրջաններ։ Այդ օրինակի վրա մենք պարզ տեսնում ենք, որ եթե մենք մտովի ցանկանում ենք բույսին բազմանալու հնարավորություն տալ, մենք պետք է օժտենք նրան որոշ առավելություններով՝ նրա դեմ մրցող ախոյանների համեմատությամբ կամ նրան ուտող կենդանու համեմատությամբ։ Աշխարհագրական տարածման սահմանագծերում կլիմայի վերաբերմամբ բույսի ընդհանուր կազմվածքի մեջ կատարված փոփոխությունը կլիներ ակնհայտ առավելություն մեր բույսի համար. բայց մենք հիմք ունենք ենթադրելու, որ շատ քիչ բույսեր կամ կենդանիներ են այդքան լայնորեն տարաբնակվում, որ իրենց տարածման սահմանագծերի վրա ոչնչանում են բացառապես կլիմայական պայմաններից։ Միայն կյանքի ծայրային սահմաններում՝ կամ բևեռային երկրներում, կամ թե անապատի եզրին ամեն ձևի մրցումը դադարում է։ Երկիրը կարող է լինել խիստ ցուրտ կամ անջուր, և այնուամենայնիվ այնտեղ ապրող սակավաթիվ տեսակների, կամ միևնույն տեսակի անհատների միջև տեղի է ունենալու կռիվ ամենատաք, ամենախոնավ տեղերի համար։
Այստեղից մենք նկատում ենք, որ եթե բույսը կամ կենդանին տեղափոխենք մի նոր երկիր, այդ դեպքում նոր ախոյանների մեջ նրա կենսապայմանները էականորեն կփոխվեն, թեկուզև կլիման բոլորովին նման լիներ նրա հայրենիքի կլիմային։ Նոր հայրենիքում նրա միջին թիվը բարձրացնելու համար մենք պետք է նրան փոփոխենք բոլորովին ոչ այնպես, ինչպես կփոփոխեինք նրան իր հայրենիքում, որովհետև պետք է որևէ գերեզանցություն տանք նրան բոլորովին այլ մրցակիցների կամ ոսոխների հանդեպ։
Օգտակար է փորձել երևակայությամբ օժտել որևէ տեսակ մի այլ տեսակի հանդեպ որևէ առավելությամբ։ Ամենայն հավանականությամբ ոչ մի դեպքում մենք չենք կարողանա այդ անել։ Դա կբերեր մեզ այն համոզման, որ մենք կատարելապես անտեղյակ ենք բոլոր օրգանական էակների միջև գոյություն ունեցող փոխհարաբերությունների վերաբերմամբ, մի համոզման, որ ինչքան անհրաժեշտ է, նույնքան էլ դժվար է ձեռք բերել։ Այդ ամենը, ինչ կարող ենք անել, դա միշտ հիշելի է, որ օրգանական էակները ձգտում են բազմանալ երկրաչափական պրոգրեսիայով, որ նրանցից ամեն մեկը որևէ հասակում, տարվա որևէ եղանակի, յուրաքանչյուր սերնդում կամ ընդմիջումներով, ստիպված է գոյության կռիվ մղել և ենթարկվել զգալի բնաջնջման։ Խորհելով այդ կռվի մասին, մենք կարող ենք մեզ մխիթարել այն մտածողությամբ, որ այդ պատերազմը բնության մեջ ունի իր ընդմիջումները, որ նրա ժամանակ ոչ մի երկյուղ չի զգացվում, որ մահը սովորաբար արագ է հարվածում և որ ուժեղները, առողջները և երջանիկներն ապրում են և բազմանում։
ԳԼՈՒԽ IV։ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅՈՒՆ ԿԱՄ ԱՄԵՆԱՀԱՐՄԱՐՎԱԾՆԵՐԻ ԳԵՐԱՊՐՈՒՄԸ
ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅՈՒՆ. ՆՐԱ ՀԶՈՐՈՒԹՅՈՒՆԸ ՄԱՐԴՈՒ ԿՈՂՄԻՑ ԿԻՐԱՌՎՈՂ ԸՆՏՐՈՒԹՅԱՆ ՀԱՄԵՄԱՏՈՒԹՅԱՄԲ. ՆՐԱ ՆԵՐԳՈՐԾՈՒԹՅՈՒՆՆ ԱՄԵՆԱՉՆՉԻՆ ՀԱՏԿԱՆԻՇՆԵՐԻ ՎՐԱ. ՆՐԱ ՀԶՈՐՈՒԹՅՈՒՆԸ ԲՈԼՈՐ ՀԱՍԱԿՆԵՐԻ ԵՎ ԵՐԿՈՒ ՍԵՌԵՐԻ ՀԱՄԱՐ։— ՍԵՌԱԿԱՆ ԸՆՏՐՈՒԹՅՈՒՆ։— ԽԱՉԱՁԵՎՄԱՆ ԸՆԴՀԱՆՐՈՒԹՅՈՒՆԸ ՄԻԵՎՆՈՒՅՆ ՏԵՍԱԿԻՆ ՊԱՏԿԱՆՈՂ ԱՆՀԱՏՆԵՐԻ ՄԻՋԵՎ։— ՀԱՆԳԱՄԱՆՔՆԵՐ, ՈՐՈՆՔ ՆՊԱՍՏՈՒՄ ԵՆ ԵՎ ՉԵՆ ՆՊԱՍՏՈՒՄ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՀԵՏԵՎԱՆՔՆԵՐԻ ԱՐՏԱՀԱՅՏՄԱՆԸ, ԱՅՆ Է՝ ԽԱՉԱՁԵՎՈՒՄԸ, ՄԵԿՈՒՍԱՑՈՒՄԸ ԵՎ ԱՆՀԱՏՆԵՐԻ ԹԻՎԸ։ ՆՐԱ ՆԵՐԳՈՐԾՈՒԹՅԱՆ ԴԱՆԴԱՂՈՒԹՅՈՒՆԸ։ ՄԱՀԱՋՆՋՈՒՄԸ ՈՐՊԵՍ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՀԵՏԵՎԱՆՔ։— ՀԱՏԿԱՆԻՇՆԵՐԻ ՏԱՐԱՄԻՏՈՒՄԸ. ՆՐԱ ԱՌՆՉՈՒԹՅՈՒՆԸ ՍԱՀՄԱՆԱՓԱԿ ՏԱՐԱԾՈՒԹՅԱՆ ՎՐԱ ԱՊՐՈՂ ԷԱԿՆԵՐԻ ԲԱԶՄԱԶԱՆՈՒԹՅԱՆ ԵՎ ՆԱՏՈՒՐԱԼԻԶԱՑԻԱՅԻ ՀԵՏ։— ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ԱԶԴԵՑՈՒԹՅՈՒՆՆ ԸՆԴՀԱՆՈՒՐ ԾՆՈՂՆԵՐԻՑ ԱՌԱՋԱՑԱԾ ՍԵՐՆԴԻ ՎՐԱ ՀԱՏԿԱՆԻՇՆԵՐԻ ՏԱՐԱՄԻՏՄԱՆ ԵՎ ՄԱՀԱՋՆՋՄԱՆ ՄԻՋՈՑՈՎ։— ՆԱ ԲԱՑԱՏՐՈՒՄ է ԲՈԼՈՐ ՕՐԳԱՆԱԿԱՆ ԷԱԿՆԵՐԻ ԽՄԲԱՎՈՐՈՒՄԸ։— ԿԱԶՄՎԱԾՔԻ ԲԱՐՁՐԱՆԱԼԸ։— ՍՏՈՐԻՆ ՁԵՎԵՐԻ ՊԱՀՊԱՆՎԵԼԸ։— ՀԱՏԿԱՆԻՇՆԵՐԻ ՀԱՄԱՄԵՏՈՒԹՅՈՒՆԸ (ԿՈՆՎԵՐԳԵՆՑԻԱ) ՏԵՍԱԿՆԵՐԻ ԱՆՍԱՀՄԱՆ ԲԱԶՄԱՑՈՒՄԸ։— ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ։
Գոյության կռիվը, որ մենք նախորդ գլխում թռուցիկ կերպով քննարկեցինք, ինչպե՞ս է ազդում փոփոխականության վրա։ Կարո՞ղ է արդյոք ընտրության սկզբունքը, որն այնքան զորավոր է մարդու ձեռքում, կիրառվել բնության նկատմամբ։ Ես կարծում եմ, մենք կհամոզվենք, որ կարող է և այն էլ շատ ներգործող ձևով։ Հիշենք անհամար, թեթև շեղումները և անհատական տարբերությունները, որոնք առաջանում են մեր ընտանի ցեղերի մեջ և ավելի նվազ չափովի բնական պայմաններում ապրող օրգանիզմների մեջ, ինչպես նաև հիշենք ժառանգական փոխանցման հզորությունը։ Ընտանի վիճակում ամբողջ կազմվածքը դառնում է որոշ չափով պլաստիկ։ Բայց այդ փոփոխականությունը, որ համարյա միշտ հատուկ է մեր ընտանի ցեղերին, ինչպես արդարացիորեն նկատում են Հուկերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] և Ազա-Գրեյը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 68] անմիջականորեն մարդը չի ստեղծել, նա ինչպես որ անզոր է նոր տարատեսակ առաջ բերելու, այնպես էլ անզոր է արգելակելու նրա առաջացումը. նա կարող է միայն պահպանել և կուտակել այն փոփոխությունները, որոնք երևան են գալիս իրենք իրենց։ Առանց կանխամտածության նա օրգանիզմները դնում է նոր և փոփոխվող կենսապայմանների մեջ և առաջ է բերում փոփոխականություն. բայց պայմանների այդպիսի փոփոխություններ կարող են երևան գալ և իրոք երևան են գալիս նաև բնության մեջ։ Չպետք է աչքաթող անել նաև այն, թե ինչքան անսահմանորեն բարդ են և ինչքան սերտորեն շաղկապված են օրգանիզմների փոխհարաբերությունները միմյանց հետ և շրջապատող ֆիզիկական պայմանների հետ, և այստեղից հասկանալի է, թե ինչքան անսահմանորեն բազմազան են կառուցվածքում եղած այն տարբերությունները, որոնք կարող են օգտակար լինել ամեն մի էակի համար շարունակաբար փոփոխվող կենսապայմանների դեպքում։ Կարելի՞ է արդյոք, մարդու համար օգտակար բազմաթիվ փոփոխությունները նկատի ունենալով, անհավանական համարել, որ մյուս փոփոխությունները, որոնք որևէ տեսակետից օգտակար են կենդանի էակների համար նրանց բարդ և կենսական մեծ մարտում, երևան են եկել սերունդների երկար շարանների ընթացքում։ Բայց եթե այդպիսի փոփոխությունները երևան են գալիս, ապա մենք կարո՞ղ ենք արդյոք կասկածել (հիշելով, որ անհամեմատ ավելի շատ անհատներ են ծնվում, քան կարող են ապրել), որ այն անհատները, որոնք օժտված են թեկուզ և ամենաչնչին առավելությամբ մնացածների հանդեպ, ավելի շատ շանսեր կունենան պահպանվելու և իրենց սերունդները բազմացնելու։ Մյուս կողմից, մենք կարող ենք հավատացած լինել, որ ամեն մի որքան և իցե վնասակար փոփոխություն անպայման կենթարկվի ոչնչացման։ Օգտակար անհատական տարբերությունների կամ փոփոխությունների պահպանումը և վնասակարների ոչնչացումը ես անվանել եմ բնական ընտրություն կամ ամենահարմարվածների գերապրում։ Անօգուտ և անվնաս փոփոխությունները չեն ենթարկվի ընտրության ազդեցությանը, այլ կներկայացներ անկայուն տատանվող մի էլեմենտ, որը մենք թերևս նկատում ենք որոշ պոլիֆորմ տեսակների մեջ, կամ էլ վերջ ի վերջո կամրանան՝ շնորհիվ օրգանիզմի բնույթի և շրջապատող պայմանների հատկությունների։
Մի քանի հեղինակներ կամ թյուր կերպով են հասկացել «բնական ընտրություն» արտահայտությունը, կամ ուղղակի առարկել են նրա դեմ։ Ոմանք նույնիսկ պատկերացրել են, թե բնական ընտրությունը առաջացնում է փոփոխականություն, մինչդեռ այդ արտահայտությամբ հասկացվում է միայն այնպիսի փոփոխությունների պահպանումը, որոնք առաջանում են և օգտակար են լինում ձեռք բերող էակներին տվյալ կենսապայմանների մեջ։ Ոչ ոք չի առարկում, երբ խոսքը մարդու կողմից գործադրվող ընտրության հզոր ներգործության մասին է, բայց այդ դեպքում էլ անհատական տարբերությունները որ մարդն այս կամ այն նպատակով ընտրում է, պիտի նախապես երևային։ Մյուսներն առարկում էին, թե «ընտրություն» բառը ենթադրում է գիտակցական ընտրություն փոփոխություն կրող կենդանիների կողմից. հասնում էին նույնիսկ մինչև այնտեղ, որ ժխտում էին ընտրության կիրառելիությունը բույսերի վերաբերմամբ, որովհետև նրանք կամքից զուրկ են։ «Բնական ընտրություն» արտահայտության բառացի իմաստով կիրառումը անշուշտ սխալ կլիներ. բայց ո՞վ է երբևիցե առարկել քիմիկոսների գործածած «ընտրողական ազգակցություն» արտահայտության դեմ։ Եվ այնուամենայնիվ չի կարելի, խիստ ասած, ընդունել, որ թթուն ընտրում է այն հիմքը, որի հետ գերադասորեն միանում է։ Ասվել է նաև, որ իբրև թե ես վկայակոչում եմ բնական ընտրությունը, որպես մի ինչ որ գործոն սկզբունք կամ աստվածություն, բայց ո՞վ է երբևիցե կշտամբել հեղինակներին այնպիսի արտահայտությունների համար, ինչպիսին է «տիեզերական ձգողականությունը ղեկավարում է մոլորակների շարժումը»։ Ամեն ոք գիտե, թե այդ փոխաբերական արտահայտությունների տակ ինչ է հասկացվում, և նրանք խոսքի հակիրճության համար համարյա անխուսափելի են։ Ճիշտ այդպես էլ «բնություն» բառը անձնավորելուց դժվար է խուսափել, բայց «բնություն» բառի տակ ես հասկանում եմ միայն բազմաթիվ բնական օրենքների համատեղ գործողությունը և արգասիքը, իսկ «օրենքներ» բառի տակ՝ երևույթների ապացուցված հաջորդականությունը։ Խնդրի հետ ավելի մոտ ծանոթանալիս այդպիսի մակերեսային առարկությունները կմոռացվեն։
Մենք մեզ համար բնական ընտրության հավանական ուղղությունը ամենից ավելի լավ կպարզենք, եթե վերցնենք դեպք, երբ մի երկրում որևէ ֆիզիկական պայման, օրինակ՝ կլիման, աննշան չափով է փոփոխվում։ Նրա բնակիչների թվական հարաբերությունը անմիջապես կենթարկվի փոփոխության, իսկ որոշ տեսակներ ամենայն հավանականությամբ վերջնականապես կմահաջնջվեն։ Հիմնվելով նրա վրա, ինչ որ մեզ հայտնի է ամեն մի երկրի բնակիչների սերտ և բարդ փոխադարձ կախման մասին, մենք իրավունք ունենք եզրակացնելու, որ բնակիչների մի մասի հարաբերական թվի ամեն մի փոփոխություն խորապես կազդի մյուսների վրա՝ անկախ բուն կլիմայի փոփոխությունից։ Եթե մի երկրի սահմանները բաց են, այդ դեպքում նոր ձևերն անշուշտ այնտեղ կներթափանցեն դրսից, իսկ այդ հանգամանքը նույնպես լուրջ կերպով կազդի նախկին բնակիչների փոխհարաբերությունների վրա։ Հիշենք, թե ինչքան զորավոր է լինում որևէ երկիր դրսից ներմուծված միմիայն մի ծառատեսակի կամ կաթնասունի ազդեցությունը։ Բայց եթե նկատի է առնվում մի կղզի կամ երկիր, որը գեթ մասամբ անջրպետներով սահմանազատված է և, հետևաբար, անմատչելի է նոր և ավելի լավ հարմարված ձևերի ներխուժման համար, ապա բնության տնտեսության մեջ կգտնվեին ազատ վայրեր, որոնք միշտ լավագույն կերպով կլցվեին, եթե տեղական բնակիչներից մի քանիսը կփոփոխվեին որևէ ուղղությամբ, որովհետև եթե այդ երկիրը բաց լիներ ներգաղթի համար, ապա այդ վայրերը կգրավեին նոր եկվորները։ Այդպիսի դեպքերում այն աննշան փոփոխությունները, որոնք օգտակար են որևէ տեսակի անհատների համար երկրի փոփոխված պայմաններին նրանց ավելի լավ հարմարվելու տեսակետից, պահպանվելու հակում կունենային, և բնական ընտրությունը լիակատար ազատություն կունենար իր ամբողջ բարելավող ներգործությունը ցուցաբերելու համար։
Մենք լիակատար հիմք ունենք մտածելու, ինչպես այդ ցույց է տրված առաջին գլխում, որ կենսապայմաններում տեղի ունեցած փոփոխությունները ուժեղացած փոփոխականության դրդապատճառ են լինում, բերված օրինակներում պայմանները փոփոխվում էին, և դա հավանորեն պետք է նպաստեր ընտրությանը, մեծացնելով օգտակար փոփոխությունների երևան գալու շանսերը։ Նրանց բացակայության դեպքում բնական ընտրությունն անզոր է որևէ բան անելու։ Չպետք է մոռանալ, որ «փոփոխություններ» բառով հասկացվում են բացառապես անհատական փոփոխությունները։ Ինչպես որ մարդը զգալի հետևանքների է հասել իր ընտանի կենդանիների և մշակովի բույսերի վերաբերմամբ, կուտակելով անհատական տարբերությունները որևէ տվյալ ուղղությամբ, նույնպես էլ կարող էր դրան հասնել նաև բնական ընտրությունը, բայց անհամեմատ ավելի հեշտությամբ, որովհետև գործում էր անհամեմատ ավելի երկարատև ժամանակաշրջանների ընթացքում։ Եվ ես չեմ կարծում, որ ֆիզիկական պայմանների, ինչպես օրինակ, կլիմայի շատ մեծ փոփոխություն, կամ ներգաղթից պաշտպանվելու համար շատ խիստ մեկուսացում պահանջվեր, որպեսզի բացվեին նոր չգրավված վայրեր, որ հետո բնական ընտրությունը լցներ այդ վայրերը՝ կատարելագործելով փոփոխության ենթարկված բնակիչներից մի քանիսին։ Որովհետև, նկատի առնելով այն, որ երկրի բոլոր բնակիչները կռվում են նրբորեն հավասարակշռված ուժերով, մի տեսակի կառուցվածքում կամ սովորություններում տեղի ունեցող աննշան փոփոխությունները նրան առավելություն կտային մյուսների նկատմամբ. իսկ այդպիսի հատկության հետագա փոփոխությունները ավելի ևս կմեծացնեին նրա առավելությունները, և դա կշարունակվեր այնքան ժամանակ, քանի դեռ այդ տեսակը կմնար միևնույն կենսապայմաններում և կօգտվեր գոյության և պաշտպանության միանման միջոցներով։ Չի կարելի անվանել որևէ երկիր, որի տեղական բնակիչները այնպիսի կատարելությամբ հարմարված լինեին միմյանց և իրենց կյանքի ֆիզիկական պայմաններին, որ նրանցից ոչ մի էակ չկարողանար ավելի ևս կատարելագործվել, ավելի ևս լավ հարմարվել, որովհետև բոլոր երկրներում բնիկները հաղթվել են նատուրալիզացիայի ենթարկված օրգանիզմների կողմից այնքան, որ թույլ են տվել եկվորներին հաստատուն ոտք դնելու իրենց հողի վրա։ Քանի որ օտարականները ամեն մի երկրում տեղացիներից ոմանց ոչնչացրել են, ապա մենք լիովին կարող ենք եզրակացնել, որ տեղացիներն էլ կարող էին իրենց օգտին փոփոխվել այնքան, որպեսզի կարողանային հակահարված տալ այդ եկվորներին։
Եթե մարդը սիստեմատիկ կամ անգիտակցական ընտրություն կիրառելով կարող է հասնել և իրոք հասել է մեծ արդյունքների, ապա ինչե՞ր ասես չի կարող իրագործել բնական ընտրությունը։ Մարդը կարող է ազդել միայն արտաքին և տեսանելի հատկանիշների վրա։ Բնությունը,— եթե ինձ թույլ կտրվի այդ տերմինով մարմնավորել օրգանիզմների բնական պահպանումը կամ ամենահարմարվածների գերապրումը,— արտաքինի մասին հոգ է տանում լոկ այնքան, որքան այդ արտաքինը օգտակար է որևէ էակի։ Նա կարող է ազդել ամեն մի ներքին օրգանի վրա, ընդհանուր մարմնակազմության յուրաքանչյուր նրբերանգի վրա, ամբողջ կենսական մեխանիզմի վրա։ Մարդն ընտրություն է կատարում իր օգտի համար, բնությունը՝ պահպանվող օրգանիզմի օգտի համար։ Կառուցվածքի յուրաքանչյուր առանձնահատկություն, որը ենթարկվում է ընտրության, օգտագործվում է լիովին, դա բխում է հենց ընտրության փաստից։ Մարդը միևնույն երկրում պահում է զանազան կլիմաների ծնունդ առածներ. նա ընտրված հատկանիշը հազվագյուտ դեպքում է ենթարկում համապատասխանող վարժության ազդեցության. նա կերակրում է թե կարճակտուց և թե երկարակտուց աղավնիներին միևնույն կերակրով. նա առանձին վարժություններ չի հնարում երկար մեջք կամ կարճ ոտներ ունեցող չորքոտանիների համար, նա կարճաբուրդ և երկարաբուրդ ոչխարներին ենթարկում է միևնույն կլիմայի ազդեցությանը։ Նա չի թողնում, որ ամենաուժեղ արուն խլի էգին ամենաթույլերից։ Նա անողորմ ոչնչացման չի ենթարկում անբավարար կենդանիներին, այլ ընդհակառակը պաշտպանում է, որքան նա այդ կարող է անել, իր բոլոր արգասիքները։ Հաճախ նրան ելակետային ձև են ծառայում կիսաայլանդակները, կամ, առնվազն, բավականաչափ ցայտուն շեղումները, որ նրա ուշադրությունը կգրավեն կամ նրան կզարմացնեն իրենց օգտակարությամբ։ Բնական վիճակում կառուցվածքի կամ ընդհանուր կերտվածքի մեջ առաջացած ամենափոքր տարբերությունը կարող դեպի իրեն հակել նրբորեն հավասարակշռված կշեռքի նժարը, գոյության կռվում ստանալ առավելություն և դրա շնորհիվ պահպանվել։ Ինչքա՜ն վաղանցուկ են մարդու ցանկություններն ու ջանքերը։ Ինչքա՜ն կարճատև են նրա օրերը։ Հետևապես և ինչքա՜ն խղճուկ են նրա ստացած արդյունքները՝ համեմատած այն արդյունքների հետ, որ կուտակել է բնությունը ամբողջ երկրաբանական ժամանակաշրջանների ընթացքում։ Դրանից հետո արդյոք մենք կարո՞ղ ենք զարմանալ, որ բնության արգասիքները մարդու արգասիքների համեմատությամբ աչքի են ընկնում ավելի «ճիշտ» հատկանիշներով, որ նրանք անհամեմատ ավելի լավ են հարմարված կյանքի անսահմանորեն բարդ պայմաններին և պարզ կերպով իրենց վրա կրում են ավելի բարձր կերտվածքի կնիք։
Փոխաբերական ձևով արտահայտվելով՝ կարելի է ասել, որ բնական ընտրությունը, որ ամեն օր, ամեն ժամ ամբողջ աշխարհում հետաքննում է ամենափոքրիկ փոփոխությունները, դեն գցելով վատերը, պահում և գումարում է լավերը, անձայն, անտեսանելի կերպով, ամենուրեք և ամեն ժամանակ, հենց որ դրա համար առիթ է ներկայանում, աշխատում է յուրաքանչյուր էակի կատարելագործման վրա՝ հարմարեցնելով նրան կյանքի օրգանական և անօրգանական պայմաններին։ Մենք չենք նկատում այդ դանդաղ կատարվող փոփոխությունների բուն ընթացքը, մինչև որ ժամանակի ձեռքը չի նշում անցյալ դարերը, բայց այդ դեպքում էլ այնքան անկատար է մեր առջև բացվող երկրաբանական անցյալի պատկերը, որ մենք նկատում ենք միայն, որ կյանքի գոյություն ունեցող ձևերը նման չեն մի ժամանակ գոյություն ունեցածներին։
Տեսակի մեջ զգալի փոփոխություններ իրականացնելու համար մի անգամ առաջացած տարատեսակը պետք է գուցե երկար ժամանակամիջոց անցնելուց հետո նորից փոփոխվի կամ ի հայտ բերի միևնույն ամենանպաստավոր ուղղությամբ անհատական շեղումներ. այդ փոփոխությունները, իրենց հերթին, պետք է պահպանվեն և այդպես շարունակ, քայլ առ քայլ։ Դիտելով իրար նման անհատական շեղումների մշտական կրկնությունը, մենք հազիվ թե իրավունք ունենանք հենց նոր ասածը համարել պարզ ենթադրություն, որի ստուգության մասին չի կարելի երաշխավորել։ Բայց նա համապատասխանում է ճշմարտության, թե ոչ, այդ մասին կարող ենք դատել միայն որոշելով, թե այդ հիպոթեզը որքան համապատասխանում է բնության ընդհանուր երևույթներին և բացատրում դրանք։ Մյուս կողմից, այն տարածված համոզմունքը, որ հնարավոր փոփոխականության չափերը խիստ սահմանափակ են, ճիշտ նույնպես էլ միայն պարզ ենթադրություն է։
Թեև բնական ընտրությունը կարող է գործել միայն տվյալ օրգանիզմի օգտին և միայն այդ օգտի շնորհիվ, այնուամենայնիվ այն հատկանիշները և կառուցվածքները, որոնք մենք համարում ենք խիստ անէական, կարող են մտնել նրա ներգործության շրջանակի մեջ։ Երբ մենք նկատում ենք, որ տերևներով սնվող միջատները կանաչ գույն ունեն, իսկ կեղևով սնվողները՝ բծավոր-գորշ, որ ալպիական կաքավը ձմեռը սպիտակ է, իսկ շոտլանդական կարմիր մայրահավը հավամրգու գույն ունի, մենք պետք է ընդունենք, որ այդ գույները օգուտ են բերում այդ թռչուններին և միջատներին, պաշտպանելով նրանց վտանգներից։ Եթե մայրահավերին չոչնչացնեին նրանց կյանքի որոշ շրջանում, ապա նրանք կբազմանային անհամար քանակությամբ. հայտնի է, որ գիշատիչ թռչունները դաժան կերպով հալածում են նրանց. մյուս կողմից, հայտնի է, որ ճուռակները ղեկավարվում են տեսողությամբ, այդ պատճառով մայր ցամաքի շատ վայրերում նախազգուշացնում են չպահել սպիտակ աղավնիներ, որոնց գիշատիչներն ամենից ավելի շատ են ոչնչացնում։ Ուրեմն՝ բնական ընտրության ներգործությունը կարող է արտահայտվել ինչպես մայրահավերի զանազան տեսակների համապատասխան գույն ձեռք բերելու մեջ, այնպես էլ արդեն ձեռք բերված այդ հատկանիշը հաստատուն պահելու մեջ։ Եվ չպետք է կարծել, որ մի առանձին ձևով գունավորված կենդանու պատահական ոչնչացումը ոչ մի էական բան չի ներկայացնում. հիշենք, թե ինչքան կարևոր է սպիտակ ոչխարների հոտի մեջ թեկուզ աննշան սև բծեր ունեցող գառների ոչնչացնելը։ Մենք տեսանք, թե ինչպես Վիրգինիայում «ներկի արմատով» կերակրվող խոզերի գույնից է կախված նրանց ապրելը կամ ոչնչանալը։ Բուսաբանները պտուղների մակերեսի վրա եղած մազմզուկները կամ նրանց պտղամսի գույնի հատկանիշները խիստ անէական են համարում. սակայն փորձված այգեգործ Դոունինգը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 77] հաղորդում է, որ Միացյալ Նահանգներում հարթ մաշկով պտուղները Curculio սեռի մի բզեզից անհամեմատ ավելի շատ են վնասվում, քան մազմզուկներով պատած պտուղները, որ կապույտ սալորները, որոշ հիվանդություններից ավելի շատ են վնասվում, քան դեղինները. ընդհակառակը, դեղին պտղամիս ունեցող դեղձերն ավելի շուտ են ենթարկվում մի այլ հիվանդության, քան ուրիշ գույնի դեղձերը։ Եթե վարպետության ամբողջ օժանդակության դեպքում էլ հատկություններում եղած այդ չնչին տարբերությունները ուղեկցվում են այդպիսի մեծ տարբերությամբ բույսերից ստացվող արդյունքներում, ապա անկասկած է, որ բնական վիճակում, որտեղ ծառերը ստիպված են պայքարել այլ ծառերի դեմ և թշնամիների մեծ խմբերի դեմ, այդպիսի տարբերությունները, ինչպիսիք են հարթ կամ մազապատ մաշկը, դեղին կամ կարմիր պտղամիսը, շատ էականորեն պետք է ազդեն տարատեսակների միջև մղվող կռվի վախճանի վրա։
Կանգ առնելով տեսակների միջև եղած բազմաթիվ փոքրիկ տարբերությունների վրա, որոնք, ինչքան մեր անտեղյակությունը թույլ է տալիս դատելու, մեզ թվում են ոչ էական, մենք չպետք է մոռանանք, որ կլիման, սնունդը և այլն անտարակույս նրանց վրա որոշ անմիջական ազդեցություն են ունեցել։ Անհրաժեշտ է նաև միշտ նկատի ունենալ, որ զարգացման հարաբերակցության օրենքի շնորհիվ, երբ փոփոխվում է որևէ մաս և բնական ընտրության միջոցով փոփոխությունները կուտակվում են, այդ դեպքում առաջանում են նաև այլ փոփոխություններ հաճախ ամենաանսպասելի հատկությամբ։
Եթե մենք տեսնում ենք, որ մեր ընտանի ցեղերի կյանքի որոշ շրջանում երևան եկող փոփոխությունները ձգտում են արտահայտվել սերունդների կյանքի միևնույն շրջանում,— սրանց են վերաբերում զանազան բանջարանոցային և դաշտային բույսերի սերմերի ձևերը, մեծությունը և համը, շերամի զանազան սորտերի բոժոժի և թրթուրի հատկությունները. մեր հավերի ձվերը և նրանց ճուտերի աղվամազի գույնը, ոչխարների և տավարի գրեթե չափահաս անհատների եղջյուրները, ապա բնական վիճակում բնական ընտրությունը պիտի ներգործի օրգանիզմների վրա ամեն մի հասակում, այդ հասակի համար ձեռնտու փոփոխությունների կուտակման միջոցով և համապատասխան հասակում դրանք ժառանգելու միջոցով։ Եթե բույսի համար ձեռնտու է քամու օժանդակությամբ իր սերմերը լայն կերպով ցրել, ապա ես չեմ հասկանում, թե ինչու դրա իրագործումը բնական ընտրության միջոցով ավելի դժվար է, քան սելեկցիայի միջոցով բամբակագործին հաջողվում է բամբակենու սերմերի վրա գտնվող թելիկները երկարացնել ու կատարելագործել։ Բնական ընտրությունը կարող է միջատի թրթուրը փոփոխել և հարմարեցնել բազմաթիվ կենսապայմաններին, որոնք բոլորովին տարբերվում են այն պայմաններից, որոնց մեջ ապրում է չափահաս միջատը. իսկ այդ փոփոխությունները հարաբերակցության օրենքի ուժով կարող են ներգործություն ունենալ նաև չափահաս ձևի վրա։ Այդպիսի ճանապարհով էլ հակառակն է լինում՝ չափահաս միջատների մեջ առաջացած փոփոխությունները կարող են անդրադառնալ նաև թրթուրների կառուցվածքի վրա, բայց համենայն դեպս բնական ընտրությունը կապահովի նրանց անվնասակարությունը, որովհետև հակառակ դեպքում դրանցով օժտված տեսակը կենթարկվի մահաջնջման։
Բնական ընտրությունը փոփոխում է ձագերի կառուցվածքը ծնողների համեմատությամբ և ծնողներինը՝ ձագերի համեմատությամբ։ Սոցիալական կյանք վարող կենդանիների մոտ նա հարմարեցնում է յուրաքանչյուր անհատի կառուցվածքը ամբողջ համայնքի պահանջներին, եթե միայն համայնքը կարող է մի օգուտ քաղել ընտրության ենթարկվող անհատի փոփոխությունից։ Բնական ընտրությունը չի կարող անել մի բան՝ դա որևէ տեսակի կառուցվածքի այնպիսի փոփոխությունն է, որից այդ տեսակը ոչ մի օգուտ չունի, այլ օգտվում է մի ուրիշ տեսակ, և թեև բնապատմական աշխատություններում կան վկայություններ, որոնք խոսում են իբր թե այդ փաստի օգտին, բայց ես չկարողացա գտնել ոչ մի դեպք, որ քննադատության դիմանար։ Այն կառուցվածքը, որը օրգանիզմի կյանքում միայն մի անգամ է ծառայում, եթե միայն նա ունի շատ էական նշանակություն, կարող է ընտրության միջոցով հասնել ուզած աստիճանի կատարելագործության. այդպես են, օրինակ, մեծ ծնոտները, որ միջատները գործածում են բացառապես բոժոժը բացելու համար, կամ կտուցի ծայրը, որ գործ են ածում ձվից դեռ դուրս չեկած ձագերը ձվի կեղևը կոտրելու համար։ Ապացուցված է, որ լավագույն կարճակտուր թուրմանների մեծ մասը ոչնչանում է ձվում, ի վիճակի չլինելով կոտրել այն, այդ պատճառով շատ աղավնաբույծներ նրանց օգնում են ձվից դուրս գալու ժամանակ։ Եթե բնության մեջ պետք լիներ աղավնուն, իր շահերի տեսակետից օժտել հասունացած վիճակում շատ կարճ կտուցով, ապա փոփոխության պրոցեսը խիստ դանդաղ կերպով կկատարվեր և դրա հետ զուգընթաց կընթանար այն ձագերի ընտրությունը, որոնք ձվի մեջ կունենային ամենազորեղ և պինդ կտուցներ, որովհետև բոլոր թույլ կտուցավորները անխուսափելիորեն կոչնչանային, կամ ընդհակառակը, կընտրվեին ամենաբարակ, հեշտ ծակվող կեղևները, որովհետև հայտնի է, որ կեղևի հաստությունը կառուցվածքի մյուս գծերի նման տատանվում է։
Գուցե այստեղ տեղին է նկատել, որ բոլոր էակները զգալի չափով ենթարկվում են նաև զուտ պատահական ոչնչացման, որը քիչ կամ բոլորովին չի ազդում բնական ընտրության ընթացքի վրա։ Այսպես, օրինակ, ամեն տարի ահագին քանակությամբ ձվեր և սերմեր ճարակ են դառնում, և նրանք կարող էին բնական ընտրությամբ փոփոխվել միայն այն դեպքում, եթե առաջացած փոփոխություններն այս կամ այն կերպ կարողանային նրանց ծառայել թշնամիներից պաշտպանվելու համար։ Մինչդեռ այդ ձվերից կամ սերմերից շատերը, եթե նրանք չոչնչացվեին, գուցե առաջացնեին մնացած կենսապայմաններին ավելի լավ հարմարված անհատներ, քան նրանք, որոնք պատահաբար ողջ են մնացել։ Ճիշտ այդպես էլ ահագին թվով չափահաս կենդանիներ ու բույսեր, անկախ նրանից, թե նրանք լավագույն կերպով են հարմարված իրենց գոյության պայմաններին կամ ոչ, ամեն տարի ոչնչանում են պատահական պատճառներից, և այդ պատճառների ներգործությունը բնավ չի կարող թուլանալ կառուցվածքի կամ ընդհանուր կազմության որևէ փոփոխություններից, որոնք այլ հանգամանքներում կարող են օգտակար լինել։ Չափահասների ոչնչացումը ինչքան կուզե թող դաժան լինի, միայն թե անհատների այն քանակությունը, որ կարող է տվյալ մարզում գոյություն ունենալ, այդ պատճառների ազդեցությամբ վերջնականապես չնվազի, կամ թող ձվերի և սերմերի ոչնչացումն էլ այնքան մեծ լինի, որ միայն նրանց հարյուրերորդական կամ հազարերորդական մասը զարգանար, այնուամենայնիվ ապրողներից ամենահարմարված անհատները, դարձյալ այն ենթադրությամբ, որ նպաստավոր ուղղությամբ շեղումներ գոյություն ունեն, կձգտեն բազմանալ ավելի մեծ թվով, քան ավելի քիչ հարմարված անհատները։ Իսկ եթե անհատների թիվը բոլորովին կնվազի հենց նոր նշված պատճառներից, որը, իհարկե, հաճախ կպատահի, այն ժամանակ բնական ընտրությունը անզոր կլինի իր ներգործությունը ցուցաբերելու որոշ բարենպաստ ուղղություններով. սակայն այդ չի կարող որպես առարկություն ներկայացվել նրա ներգործության դեմ մի այլ ժամանակում կամ այլ ուղղությամբ, որովհետև մենք ոչ մի առիթ չունենք ենթադրելու, որ երբևիցե միևնույն ժամանակում և միևնույն վայրում մեծ թվով տեսակներ ենթարկվել են փոփոխությունների ու կատարելագործությունների։
ՍԵՌԱԿԱՆ ԸՆՏՐՈՒԹՅՈՒՆ
Քանի որ ընտանի ցեղերի մոտ հաճախ երևան են գալիս առանձնահատկություններ, որոնք հատուկ են միայն մեկ սեռի և ժառանգաբար փոխանցվում են մյուս սեռին, ուստի կասկած չկա, որ նույնը պետք է լինի նաև բնության մեջ։ Ուրեմն՝ հնարավորություն կա, որ բնական ընտրության միջոցով երկու սեռերը կենսակերպի տարբերության կապակցությամբ փոփոխության ենթարկվեն, որը երբեմն էլ լինում է, կամ փոփոխվի մի սեռը մյուսի համեմատությամբ, որը սովորական է։ Դա ինձ ստիպում է մի քանի խոսք ասել այն մասին, որ ես անվանել եմ սեռական ընտրություն։ Ընտրության այս ձևը կախված է ոչ թե այլ էակների կամ արտաքին պայմանների հետ առնչություն ունեցող գոյության կռվից, այլ մի սեռի, առավելապես արու անհատների միջև մղվող կռվից, որը տեղի է ունենում մյուս սեռի անհատներին տիրանալու համար։ Դրա հետևանքով ստացվում է ոչ թե անհաջող կռվողի մահ, այլ նրա սերունդների սահմանափակում կամ լիակատար բացակայություն։ Ուրեմն՝ սեռական ընտրությունը այնպես դաժան չէ, ինչպես բնական ընտրությանը։ Սովորաբար ամենաուժեղ արուները, որոնք ամենից ավելի հարմարված են բնության մեջ իրենց գրաված վայրերին, թողնում են ավելի բազմաքանակ սերունդ։ Սակայն շատ դեպքերում հաղթանակը կախված է ոչ այնքան ընղհանուր ուժեղությունից, որքան կռվի առանձնահատուկ միջոցներ ունենալուց, որոնք բացառապես հատուկ են արուներին։ Անեղջյուր եղջերուն կամ բիտ չունեցող աքլորը իրենցից հետո բազմաթիվ սերունդ թողնելու շատ վատ շանսեր կունենային։ Սեռական ընտրությունը, որ հաղթողին միշտ հնարավորություն է տալիս շարունակելու իր սերունդը, կարող է, իհարկե, զարգացնել արուների մեջ անսաստելի քաջություն, բիտերի երկարություն և թևերի ուժեղություն, համարյա այնպես, ինչպես աքլորակռվի կոպիտ սիրահարը հասնում է նույն հետևանքին՝ խնամքով ընտրելով իր լավագույն աքլորներին։ Օրգանական սանդուխքի թե ո՞ր ամենացածր աստիճանի վրա դադարում է այդ օրենքի գործողությունը, որի ուժով արուները մարտնչում են էգերի համար, ես չգիտեմ. արու ալիգատորները, ըստ եղած նկարագրությունների, իրենց էգերին տիրանալու համար կռվում են իրար հետ և այդ ժամանակ մռնչում և պտտվում են, ինչպես հնդիկներն իրենց ռազմական պարի ժամանակ. տեսել են սալմոնների արուներին իրար հետ ամբողջ օրերով կռվելիս. եղջերաբզեզի արուները երբեմն իրենց վրա կրում են մյուս արուների հսկայական ծնոտների հասցրած վերքերի հետքերը. Ֆաբրի՝[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 78] այդ անզուգական բնախույզի վկայությամբ, մի քանի ցանցաթև միջատների արուները իրար հետ կռվում են մի որևէ էգի համար, որը կողմնակի հանդիսատեսի պես նստած է այդտեղ մոտիկ և հետո հեռանում է հաղթողի հետ։ Այդ կռիվները, ըստ երևույթին, ամենից ավելի դաժան են բազմամուսնական կենդանիների արուների միջև, հենց այդ արուները ամենից ավելի հաճախ զինված են կռվի հատուկ միջոցներով։ Գիշակեր կենդանիների արուներն արդեն առանց այդ էլ լավ զինված են, թեև նրանք, ինչպես և մյուս կենդանիները, սեռական ընտրության միջոցով կարող են ձեռք բերել նաև պաշտպանության առանձին միջոցներ, ինչպիսիք են, օրինակ, առյուծի բաշը և արու սալմոնների կեռաձև ծնոտը. չէ որ վահանը նույնպես կարող է հաղթանակ ապահովել, ինչպես սուրը կամ նիզակը։
Թռչունների մեջ այդ մրցությունը հաճախ ավելի խաղաղ բնույթ ունի. բոլոր հետազոտողները, որոնք ուշադրություն են դարձրել այդ խնդրի վրա, համաձայն են այն բանին, որ շատ թռչնատեսակների արուների միջև կա ուժեղ մրցակցություն, որը հիմնված է էգերին երգով գրավելու վրա։ Գվիանական լեռնային կեռնեխի, դրախտահավի և մի քանի այլ թռչունների արուներն ու էգերը հավաքվում են մի տեղ, ընդ որում արուները էգերի առջև խնամքով բաց են անում իրենց վառ գունավորված փետուրները ցույց տալու համար և կատարում են օտարոտի մարմնաշարժումներ, իսկ էգերը մնում են որպես անտարբեր հանդիսատեսներ, մինչև որ ընտրում են իրենց համար ամենագրավիչ արուներին։ Նրանք, ովքեր առիթ են ունեցել մանրազնին կերպով դիտելու գերության մեջ գտնվող թռչունների բարքերը, շատ լավ գիտեն, որ նրանք հաճախ դրսևորում են անհատական համակրանք և հակակրանք. այսպես, սըր Ռ. Հիրոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 79] հաղորդում է մի բծավոր արու սիրամարգի մասին, որն առանձնապես գրավիչ է եղել թռչնանոցում ապրող բոլոր էգ սիրամարգերի համար։ Ես այստեղ չեմ կարող մտնել անհրաժեշտ մանրամասնությունների մեջ, բայց ինձ թվում է, որ եթե մարդը կարճ ժամանակում կարող է որևէ բանտամյան հավերի փոփոխել գեղեցկության մասին իր ունեցած իդեալի համեմատ, ապա ես պատճառ չեմ տեսնում, թե ինչու նաև թռչունների էգերը, հազարավոր սերունդների ընթացքում ընտրելով ամենաքաղցրալուր երգող և ամենագեղեցիկ արուներին, չէին կարող հասնել ակնհայտ արդյունքների իրենց գեղեցկության իդեալի տեսակետից։ Մի քանի լավ հայտնի օրենքներ, որոնք վերաբերում են չափահաս արուների և էգերի փետրավորմանը՝ համեմատած ձագերի փետրավորման հետ, կարող են մասամբ բացատրվել նրանով, որ սեռական ընտրությունն ազդում է այն փոփոխությունների վրա, որոնք արտահայտվում են որոշ հասակում և փոխանցվում են կամ միայն արուներին, կամ երկու սեռերին նրանց համապատասխան հասակներում, բայց այստեղ տեղը չէր բավականանա այդ խնդրի լայն լուսաբանման համար։
Այսպես, օրինակ, ինձ թվում է, որ այն դեպքերում, երբ որևէ կենդանու արուներն ու էգերը, ունենալով միևնույն կացութաձևը, տարբերվում են իրենց կառուցվածքով, գույնով կամ առանձին զարդարանքներով, ապա այդ տարբերությունները առաջացել են գլխավորապես սեռական ընտրության միջոցով, այսինքն մի շարք սերունդների ընթացքում առանձին արուներ մյուսների համեմատությամբ ունեցել են որոշ թեթև առավելություններ կամ զինման եղանակով, կամ պաշտպանության միջոցներով, կամ թե առանձին հրապույրներով և այդ առավելությունները փոխանցել են բացառապես միայն իրենց արական սեռի հետնորդներին։ Սակայն ես համաձայն չեմ առանց բացառության բոլոր սեռական տարբերությունները վերագրել միայն այդ դրդապատճառի ազդեցությանը. մենք տեսնում ենք, որ մեր ընտանի կենդանիների մեջ առաջացել են բացառապես միայն արական սեռի անհատներին փոխանցվող առանձնահատկություններ, որոնք, ըստ երևույթին, չեն ուժեղացել արհեստական ընտրությամբ։ Վայրի հնդկական աքլորի կրծքի վրայի մազափունջը ոչ մի օգուտ չի կարող տալ, և հազիվ թե նա կարող է հնդուհավերի աչքում նաև որպես զարդարանք ծառայել, եթե նա առաջանար ընտանի թռչունի վրա, այն կհամարեին այլանդակություն։
ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՆԵՐԳՈՐԾՈՒԹՅԱՆ ԿԱՄ ԱՄԵՆԱՀԱՐՄԱՐՎԱԾՆԵՐԻ ԳԵՐԱՊՐԵԼՈՒ ՕՐԻՆԱԿՆԵՐ
Բնական ընտրության ներգործության եղանակը պարզելու համար, ինչպես ես ըմբռնում եմ այն, ես թույլատվություն կխնդրեմ ներկայացնել մի քանի երևակայական օրինակներ։ Կանգ առնենք գայլի օրինակի վրա, որը սնվում է զանազան կենդանիներով և հաղթում է մեկին խորամանկությամբ, մյուսին ուժով, երրորդին իր ոտքերի արագավազությամբ. այնուհետև ենթադրենք, թե ամենաարագավազ զոհերը, օրինակ եղջերուները, երկրում տեղի ունեցած այս կամ այն փոփոխությունների հետևանքով թվով շատացել են, կամ ընդհակառակը, նրա մի այլ որսը թվով քչացել է տարվա հենց այն ժամանակը, երբ գայլերը սննդի ամենից ավելի պակաս են զգում։ Այդպիսի պայմաններում ամենաարագավազ և ամենաճարպիկ գայլերն ամենից ավելի շանսեր կունենան գերապրելու և այսպիսով կպահպանվեն, կընտրվեն, իհարկե այն պայմանով, եթե նրանք չեն կորցնի այն ուժը, որն անհրաժեշտ է, որպեսզի հաղթահարեն իրենց որսակենդանիներին տարվա այս կամ այն ժամանակներում, երբ ստիպված են բավարարվել այլ զոհերով։ Ես ոչ մի հիմք չեմ գտնում այդ հետևանքը ավելի կասկածելի համարելու, քան այն, որ մարդը կկարողանար կատարելագործել իր բարակների արագավազությունը մանրազնին մեթոդիկ ընտրության կամ անգիտակցական ընտրության միջոցով, որը հետևանք է այն բանի, որ նա հոգ է տանում լավագույն շներ պահել, առանց մտածելու ցեղն ազնվացնելու մասին։ Կարող եմ ավելացնել, որ, ըստ Պիրսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 80] վկայության, Միացյալ Նահանգներում, Կատսկիլյան լեռներում հանդիպում են գայլի երկու տարատեսակներ՝ մեկը թեթև, որ նման է մեր բարակ շանը և հետապնդում է եղջերուներին, իսկ մյուսը ավելի հաստամարմին է, ավելի կարճաոտ, որը հաճախ հարձակվում է ոչխարների հոտերի վրա։
Պետք է նկատենք, որ նախորդ օրինակում ես խոսում եմ գայլերի ամենադյուրաշարժ անհատների մասին, այլ ոչ թե մի որևէ ցայտուն կերպով արտահայտված տարատեսակի մասին։ Նախորդ հրատարակություններում ես երբեմն այնպես եմ արտահայտվել, որ իբր թե այդ վերջին պատահականությունը հաճախակի է հանդիպում։ Ես նկատել եմ անհատական տարբերությունների կարևոր նշանակությունը, և այդ ինձ հանգեցրեց հանգամանորեն քննարկելու այն հետևանքները, որ ունենում է մարդու կողմից կատարվող անգիտակցական ընտրությունը, որը կախված է բոլոր շատ թե քիչ արժեքավոր անհատները պահելուց և վատերը ոչնչացնելուց։ Ինձ համար նաև պարզ էր, որ կառուցվածքում տեղի ունեցող որևէ պատահական շեղում, ինչպես օրինակ, այլանդակությունները, բնական դրության մեջ կարող է հանդիպել միայն շատ հազվագյուտ դեպքերում և որ սկզբում պահպանվելու դեպքում էլ սովորական անհատների հետ խաչաձևվելու հետևանքով նա սովորաբար կորչում է։ Այնուամենայնիվ, քանի դեռ չէի կարդացել «North British Review»[40] (1867 թ.) հանդեսում շատ հմուտ և արժեքավոր հողվածը, ես լիովին չէի գնահատում, թե ինչքան հազվադեպ կարող են սերունդների մեջ պահպանվել եզակի փոփոխությունները, միևնույնն է՝ թույլ են արտահայտված, թե խիստ։ Հեղինակը քննարկում է մի դեպք, երբ կենդանիների մի զույգ իր կյանքի ընթացքում առաջ է բերում երկու հարյուր ձագեր, որոնցից զանազան պատճառներից ոչնչանալու հետևանքով միջին հաշվով մնում են երկուսը և իրենցից հետո թողնում սերունդ։ Այդ գնահատականը շատ բարձր է բարձրակարգ կենդանիների վերաբերմամբ, բայց բնավ այդպիսին չէ ստորին կարգի կենդանիներից շատերի վերաբերմամբ։ Այնուհետև նա ցույց է տալիս, որ եթե ծնվեր այնպես փոփոխված մի անհատ, որ նրա ապրելու շանսերը կրկնապատկվեին մյուս անհատների ունեցած շանսերի համեմատությամբ, ապա նույնիսկ այդ դեպքում էլ նրա ապրելու հավանականությունները քիչ կլինեին։ Բայց ենթադրենք, թե նա ապրում և թողնում է սերունդ, որի կեսը ժառանգում է բարենպաստ փոփոխությունը. այնուամենայնիվ շարունակում է հոդվածի հեղինակը, այդ մատաղ սերունդը միայն մի քիչ ավելի շանսեր կունենա ապրելու և ցեղը պահպանելու համար, և այդ շանսերը յուրաքանչյուր նոր սերնդի մեջ գնալով պիտի պակասեն։ Այս դիտողությունների ճշտությունը հազիվ թե կարող է կասկածի ենթարկվել։ Եթե, օրինակ, որևէ թռչուն կեռանման կտուցով ավելի հաջողությամբ կարողանա իր սնունդը ճարել և եթե ծնվի խիստ կորացած կտուցով մի թռչուն, որը և փարթամորեն աճի, ապա, այնուամենայնիվ, շատ քիչ շանսեր կան, որ այդ մեկ թռչունի սերունդները այնքան բազմանան, որ ամբողջովին դուրս մղեն հիմնական ձևը. բայց հազիվ թե որևէ կասկած կարող է լինել, դատելով նրանով, ինչ տեղի է ունենում մեր աչքերի առաջ մեր ընտանի ցեղերի մեջ, որ այդ հետևանքը կստացվեր, եթե քիչ թե շատ խիստ կեռացած կտուց ունեցող անհատները պահպանվեին բազմաթիվ սերունդների ընթացքում, իսկ ամենաուղիղ կտուց ունեցող ավելի ևս մեծ թվով անհատներ ենթարկվեին ոչնչացման։
Սակայն չպետք է աչքաթող անել, որ մի քանի բավական ուժեղ արտահայտված փոփոխություններ, որոնք ոչ ոք չէր կարող համարել պարզ անհատական տարբերություններ, հաճախ են կրկնվում այն պատճառով, որ նման կազմվածքը կրում է նաև միջավայրի նման ներգործությունները,— այս փաստը կարող է հաստատվել բազմաթիվ օրինակներով, որ տալիս են մեր ընտանի ցեղերը։ Այդպիսի դեպքերում եթե փոփոխվող անհատը իրականում չփոխանցեր իր հետնորդներին նոր ձեռք բերված հատկանիշը, ապա անշուշտ, քանի որ արտաքին պայմանները կմնային անփոփոխ, նա իր հետնորդներին կփոխանցեր միևնույն ուղղությամբ փոփոխվելու ավելի ևս ուժեղ արտահայտված ձգտում։ Նմանապես կասկած չի կարող լինել, որ նույն ուղղությամբ փոփոխվելու ձգտումը երբեմն արտահայտվել է այնպիսի ուժով, որ միևնույն տեսակի բոլոր անհատները փոփոխվել են նման ձևով, առանց որևէ ընտրության մասնակցության։ Կամ այդպիսի փոփոխության ենթարկվել է բոլոր անհատների մի երրորդական, մի հինգերորդական կամ մի տասերորդական մասը, որի վերաբերյալ կարելի է բերել մի քանի օրինակներ։ Այսպես, Գրաբան[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 81] գտնում է, որ Ֆարերյան կղզիների վրա բոլոր կայրաների մոտավորապես մի հինգերորդը ներկայացնում է այն աստիճանի ցայտուն կերպով նշմարելի տարատեսակ, որ առաջ նրան նույնիսկ համարում էին առանձին տեսակ Uria lacrimans անունով։ Նման դեպքերում, եթե փոփոխությունը օգտակար է, ապա ամենահարմարվածի գերապրելու հետևանքով փոփոխված ձևը արագ կերպով դուրս կմղի նախասկզբնական ձևին։
Խաչաձևման հետևանքներին, ամեն մի փոփոխության վերացման իմաստով, ես առիթ կունենամ վերադառնալու, բայց այստեղ էլ արդեն կարելի է նկատել, որ կենդանիների և բույսերի մեծ մասը կառչում է իր բնակավայրին և առանց կարիքի չի թողնում այն. մենք այդ նկատում ենք նույնիսկ չվող թռչունների վերաբերմամբ, որոնք միշտ վերադառնում են միևնույն տեղը։ Սրանից հետևում է, որ ամեն մի նոր առաջացած տարատեսակ սկզբում համարյա միշտ կլինի տեղական, ինչպես այդ թվում է, թե կարելի է ընդունել որպես կանոն բնության մեջ հանդիպող տարատեսակների վերաբերմամբ, այսպիսով, այն անհատները, որոնք փոփոխվել են միանման ուղղությամբ, շուտով համախմբված կլինեն մի խմբում և կբազմանան միասին։ Եթե նոր տարատեսակը հաջողություն ունենա գոյության կռվում, այդ դեպքում նա կսկսի դանդաղ կերպով տարածվել իր կենտրոնական շրջանից, մրցության մեջ մտնելով չփոփոխված անհատների հետ և հաղթելով նրանց իր շարունակ ընդարձակվող բնակավայրի եզրագծերում։
Գուցե անօգուտ չի լինի բերել էլի մեկ և ավելի բարդ օրինակ, որը պարզաբանում է բնական ընտրության ներգործության եղանակը։ Մի քանի բույսեր արտադրում են քաղցր հեղուկ, ըստ երևույթին նրա համար, որ հեռացնեն իրենց հյութերից ինչ-որ վնասակար բան. այդ կատարվում է, օրինակ, գեղձերի օգնությամբ, որոնք տեղավորված են տերևակիցների հիմքի մոտ, ինչպես, օրինակ, մի քանի լոբազգի բույսերինն է, կամ տերևի ստորին մակերեսի վրա, ինչպես դափնունն է։ Այդ հյութը, թեև քանակությամբ չնչին է, ագահությամբ կլանում են միջատները, բայց նրանք իրենց այցելություններով բույսին ոչ մի օգուտ չեն տալիս։ Այժմ պատկերացնենք, թե այդ հյութը կամ նեկտարը կարտազատվի որևէ տեսակի որոշ ծաղիկների ներսում։ Միջատները նեկտար որոնելիս հաճախ կծածկվեն ծաղկափոշով և շատ հաճախ կտեղափոխեն այն մեկ ծաղկից մյուսի վրա։ Այսպիսով տեղի կունենա խաչաձևում այնպիսի ծաղիկների միջև, որոնք պատկանում են երկու տարբեր անհատների, իսկ խաչաձևման պրոցեսը, ինչպես այդ կարելի է ապացուցել ամենաանկասկածելի ձևով, սկիզբ կտա ավելի հզոր սերմնաբույսերի, որոնք, հետևաբար, մեծագույն շանսեր կունենան բարգավաճելու և գերապրելու համար։ Այն բույսերի, որոնք առաջացնում են ամենամեծ քանակությամբ նեկտար արտազատող ամենամեծ նեկտարանոցներ ունեցող ծաղիկներ, ավելի հաճախ կենթարկվեն միջատների այցելությունների և ավելի հաճախ կենթարկվեն խաչաձևման և, վերջ ի վերջո, կհաղթահարեն իրենց մրցակիցներին, առաջացնելով տեղական տարատեսակ։ Ճիշտ այդպես և այն ծաղիկները, որոնց առէջքները և վարսանդները տեղադրված են իրենց հաճախակի այցելող միջատների մեծության և շարժուձևին համապատասխանորեն և այդպիսով գոնե մի քիչ հեշտացնում են ծաղկափոշու տեղափոխությունը, նույնպես կլինեին ընտրյալներ։ Մենք կարող ենք պատկերացնել նա և մի այլ դեպք, այն է՝ նեկտարի փոխարեն ծաղկափոշի հավաքող միջատներ։ Քանի որ ծաղկափոշին բացառապես ծառայում է բեղմնավորման համար, ապա նրա ոչնչացումը, թվում է, թե պիտի միայն վնաս հասցներ բույսին, այնուամենայնիվ, եթե գեթ մի քիչ ծաղկափոշի, սկզբում պատահաբար, իսկ հետո մշտապես, ծաղկափոշի լափող միջատների կողմից տեղափոխվի մի ծաղկից մյուսի վրա և դրանով իսկ ստացվի խաչաձևում, այդ դեպքում թեկուզ ամբողջ ծաղկափոշու ինը տասերորդական մասն էլ ոչնչանա, բույսը մեծ օգուտի մեջ կլիներ, իսկ այն անհատները, որոնք շարունակ ավելի ու ավելի շատ ծաղկափոշի կարտադրեն և օժտված կլինեն ավելի ու ավելի խոշոր առէջքներով, կենթարկվեն ընտրության։
Երբ երկար ժամանակ կրկնվող այդպիսի պրոցեսի հետևանքով մեր բույսը կդառնա միջատների համար խիստ գրավիչ, նրանք առանց որևէ դիտավորության կսկսեն կանոնավոր կերպով ծաղկափոշին տեղափոխել ծաղկից ծաղիկ. իսկ որ նրանք այդ էլ անում են իրականում, ես կարող եմ ցույց տալ բազմաթիվ զարմանալի օրինակներով։ Կբերեմ այստեղ միայն մեկը, որը միաժամանակ պարզաբանում է բույսերի սեռերի բաժանման պրոցեսի աստիճաններից մեկը։ Մատեի որոշ ծառեր տալիս են միայն բավական քիչ ծաղկափոշի առաջացնող չորս առէջքներով և սաղմնային վարսանդով արական ծաղիկներ. մյուսները տալիս են միայն իգական ծաղիկներ, որոնք ունենում են միանգամայն զարգացած վարսանդներ և չորս առէջքներ բոլորովին թորշոմած փոշանոթներով, որոնց մեջ ոչ մի հատիկ ծաղկափոշի չի լինում։ Գտնելով մի իգական ծառ արականից վաթսուն յարդ հեռավորության վրա, ես հետազոտեցի տարբեր ճյուղերից վերցրած քսան ծաղիկների սպիները մանրադիտակի տակ և առանց բացառության բոլորի վրա կային ծաղկափոշու հատիկներ, իսկ մի քանիսի վրա նույնիսկ առատությամբ։ Քանի որ քամին մի քանի օրվա ընթացքում փչում էր իգական ծառից դեպի արականը, ուրեմն ծաղկափոշին չէր կարող քամու բերածը լինել։ Եղանակը ցուրտ էր և փոթորկոտ, հետևաբար և անբարենպաստ էր մեղուների համար, բայց և այնպես իմ հետազոտած յուրաքանչյուր վարսանդային ծաղիկ փոշոտված էր մեղուների միջոցով, որոնք նեկտար հավաքելու համար թռել էին ծառից ծառ։ Բայց վերադառնանք մեր երևակայական դեպքին, հենց որ բույսն այնքան գրավիչ դարձավ միջատների համար, որ ծաղկափոշին կանոնավոր կերպով փոխադրվում է ծաղկից ծաղիկ, կարող է դրսևորել մի այլ պրոցես։ Իհարկե, ոչ մի բնախույզ չի կասկածում այսպես կոչված «աշխատանքի ֆիզիոլոգիական բաժանման» սկզբունքի օգտակարության մասին, սրանից մենք կարող ենք հետևցնել, որ բույսի համար օգտակար է եղել առաջացնել միայն առէջքները մեկ ծաղկի մեջ, կամ ամբողջ բույսի վրա, և միայն վարսանդները մի այլ ծաղկի մեջ կամ մի ամբողջ բույսի վրա։ Նոր կենսապայմանների մեջ տեղափոխված մշակովի բույսերի վրա երբեմն արական, իսկ այլ դեպքերում իգական օրգանը ավելի կամ պակաս չափով ամուլ է լինում, եթե մենք ընդունենք, որ նույնը կատարվում է, թեկուզ թույլ չափերով, բնության մեջ, ապա գիտենալով, որ ծաղկափոշին արդեն կանոնավոր կերպով տեղափոխվում է ծաղկից ծաղիկ և որ աշխատանքի բաժանման սկզբունքի հիման վրա սեռերի ավելի լիակատար բաժանումը բույսի համար միայն օգտակար է, մենք կգանք այն եզրակացության, որ այն անհատները, որոնց մոտ այդ ձգտումը ավելի ու ավելի ցայտուն կերպով կարտահայտվի, կենթարկվեն ընտրության, մինչև որ վերջապես կիրագործվի սեռերի լիակատար բաժանումը։ Չափազանց շատ տեղ կպահանջվի ցույց տալու համար, թե ինչպիսի աստիճանական քայլերով, դիմորֆիզմի միջոցով կամ այլ ուղիներով զանազան բույսերի մոտ, ըստ երևույթին, կատարվում է սեռերի բաժանումը։ Բայց ես կարող եմ ավելացնեի որ Հյուսիսային Ամերիկայի մի քանի մատեները, ըստ Ազա-Գրեյի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 68] գտնվում են միջանկյալ վիճակում և, ինչպես ասում է նա, կարող են անվանվել առավել կամ պակաս չափով երկտուն-բազմամուսին բույսեր։
Այժմ դառնանք նեկտարով սնվող միջատներին. մենք կարող ենք ենթադրել, որ այն բույսը, որի նեկտարի պարունակությունը մենք տևական սելեկցիայի միջոցով հետզհետե բարձրացրել ենք, տվյալ երկրում սովորական է և նրա նեկտարը որոշ միջատների հիմնական սնունդն է։ Ես կարող էի բերել շատ փաստեր, որոնք ցույց են տալիս, թե ինչպես մեղուները խնայում են ժամանակը. այսպես է, օրինակ, նրանց վարվելաձևը, երբ կրծելով ծակում են պսակի հիմքը և այդպիսով արտածծում են նեկտարը, որին նրանք կարող են հասնել նաև վերևի մուտքից։ Ուշադրության առնելով այդպիսի փաստերը, մենք կարող ենք ընդունել, որ որոշ պայմաններում կնճիթի կորության կամ երկարության անհատական փոփոխությունները, որոնք այնքան չնչին են, որ մենք դրանք չէինք էլ նկատի, կարող են մեղվի կամ մի այլ միջատի համար օգտակար լինել, այնպես որ որոշ անհատներ ի վիճակի կլինեն իրենց սնունդը հայթայթել ավելի հեշտությամբ, քան մյուսները. այսպիսով, այն համայնքները, որոնց պատկանում են նրանք, կբարգավաճեն և շատ ձագեր կտան, որոնք կժառանգեն նույն հատկությունը։ Սովորական կարմիր երեքնուկի և ինկարնատային երեքնուկի (Trifolium pratense և Tr. incarnatum) պսակի խողովակները մակերեսորեն դիտելիս երկարությամբ չեն տարբերվում միմյանցից, բայց և այնպես սովորական մեղուն կարողանում է հեշտությամբ արտածծել ինկարնատային երեքնուկի նեկտարը, բայց չի կարողանում հասնել սովորական երեքնուկի նեկտարին, որին այցելում են միայն իշամեղուները, այնպես որ կարմիր երեքնուկի ամբողջ դաշտերը իզուր առաջարկում են մեր մեղվին իրենց արժեքավոր նեկտարի առատ պաշարները։ Կասկածի ենթակա չէ, որ մեր մեղուները սիրում են այդ նեկտարը, որովհետև ես բազմիցս նկատել եմ, բայց միայն աշնանը, թե ինչպես մեղուները արտածծում էին նեկտարը ծաղկի հիմքի մոտի անցքերով, որ կրծելով բաց են արել իշամեղուները։ Երեքնուկի երկու տեսակների պսակների երկարության տարբերությունը, որից կախված է մեղուների այցելությունը ծաղիկներին, պիտի որ շատ աննշան լինի, որովհետև ինձ հավատացնում էին, որ կարմիր երեքնուկի առաջին հարից հետո երևացած ծաղիկները մի քիչ ավելի փոքր են առաջին ծաղիկներից, բայց և այնպես նրանց այցելում են բազմաթիվ մեղուներ։ Չգիտեմ, ճի՞շտ է արդյոք այդ ցուցմունքը, ինչպես նաև չգիտեմ, արդյոք կարելի՞ է վստահել մի այլ տպագիր վկայության, որ իբր թե լիգուրիական մեղուն, որը բնախույզների կողմից ճանաչված է որպես սովորական մեղվի մի տարատեսակ, որի հետ նա հեշտությամբ խաչաձևվում է, կարողանում է հասնել նեկտարանոցներին և արտածծել նաև սովորական երեքնուկի նեկտարը։ Ուրեմն՝ այն երկրում, որտեղ առատությամբ աճում է այդ երեքնուկը, մեղուների համար շատ ձեռնտու կլիներ ունենալ մի քիչ ավելի երկար և մի քիչ այլ ձևի կնճիթ։ Մյուս կողմից, քանի որ այդ երեքնուկի պտղաբերությունը անպայման կախված է նրա ծաղիկներին մեղուների այցելությունից, ուստի իշամեղուների թիվը քչանալու դեպքում բույսի համար ձեռնտու կլիներ ձեռք բերել ավելի կարճ և ավելի խոր ճեղքված պսակ, որը թույլ կտար մեղուներին ծծելու նրա ծաղիկների նեկտարը։ Այսպիսով, ես կարողանում եմ հասկանալ, թե ինչպես ծաղիկը և մեղուն աստիճանաբար միաժամանակ կամ հաջորդականորեն կփոփոխվեն և կհարմարվեն մեկը մյուսին ամենակատարյալ ձևով, անընդհատ պահպանելով այն անհատներին, որոնք իրենց կառուցվածքում ունեն աննշան փոխադարձաբար օգտակար շեղումներ։
Ես լիովին գիտակցում եմ, որ բերված օրինակներով պարզաբանված բնական ընտրության ուսմունքը կհանդիպի նույնպիսի առարկությունների, ինչ առարկությունների որ ենթարկվում էին նաև սըր Չարլզ Լյաելի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] մեծ գաղափարները, «Երկրի մակերևույթի ժամանակակից փոփոխությունները, որոնք մեզ բացատրում են երկրաբանական երևույթները» աշխատության առաջին լույս ընծայման ժամանակ, բայց այժմ շատ հազվադեպ կարելի է լսել, որ այն գործոնները, որոնց մենք մինչև այժմ էլ տեսնում ենք գործողության մեջ, ճանաչվեին չնչին և ոչ մի նշանակություն չունեցող, երբ խոսքը վերաբերում է ամենախոր գետահովիտների առաջացման պատճառներին կամ մայր ցամաքների ներսում ձգվող երկար ժայռաշարքերի գտնվելու պատճառների մասին։ Բնական ընտրությունը ներգործում է միայն պահպանելով և կուտակելով փոքրիկ ժառանգական փոփոխությունները, որոնցից յուրաքանչյուրը ձեռնտու է պահպանվող էակի համար։ Ինչպես որ արդի երկրաբանությունը գիտությունից գրեթե արտաքսել է այնպիսի հայացքները, ինչպես, օրինակ, ամբողջ խոր հովտի փորվելը մի հզոր դիլյուվիալ ալիքի միջոցով, այնպես էլ բնական ընտրությունը գիտությունից կարտաքսի այն հավատը, որ նոր օրգանական էակները շարունակ ստեղծագործվում են և նրանց կառուցվածքում որևէ հանկարծական խոր փոփոխություններ են առաջանում։
ԽԱՉԱՁԵՎՈՒՄԸ ԱՆՀԱՏՆԵՐԻ ՄԻՋԵՎ
Ես ստիպված եմ այստեղ մի փոքրիկ շեղում անել։ Բաժանասեռ կենդանիների և բույսերի վերաբերմամբ ինքնին ակնհայտ է, որ յուրաքանչյուր ծննդի համար անհրաժեշտ է երկու անհատների մասնակցությունը (բացառությամբ կուսածնության ուշագրավ և ոչ լիովին հասկանալի դեպքերի), բայց հերմաֆրոդիտ էակների վերաբերմամբ դա այնքան էլ ակնհայտ չէ։ Այնուամենայնիվ հիմք կա ենթադրելու, որ նաև բոլոր հերմաֆրոդիտների մոտ ժամանակ առ ժամանակ կամ միշտ վերարտադրության համար երկու անհատներ միանում են։ Այդ տեսակետը, թեև որոշ կասկածանքով, արտահայտել են Շպրենգելը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 82] Նայտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 31] և Կյոլռեյտերը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] Մենք հիմա կհամոզվենք նրա կարևորության մեջ, բայց ես հարկադրված եմ միայն համառոտակի շոշափել այդ հարցը, թեպետ ունեմ առատ նյութ նրա հիմնավոր քննարկության համար։ Բոլոր ողնաշարավորները, բոլոր միջատները և կենդանիների մի քանի այլ ընդարձակ խմբեր ամեն մի ծննդի համար զուգավորվում են։ Նորագույն հետազոտությունները զգալի չափով կրճատել են կարծեցյալ հերմաֆրոդիտների թիվը, իսկ իսկական հերմաֆրոդիտներից շատերը զուգավորվում են, այսինքն երկու անհատներ միանում են վերարտադրության համար, իսկ դա այն ամենն է, ինչ մեզ հարկավոր է։ Բայց դեռ մնում են մեծ թվով հերմաֆրոդիտ կենդանիներ, որոնք սովորաբար չեն զուգավորվում և բույսերի հսկայական մեծամասնությունը հերմաֆրոդիտ է։ Կարելի է հարցնել իսկ ի՞նչ հիմք կա ենթադրելու, որ այդ դեպքերում երբևիցե երկու անհատներ պիտի մասնակցեն վերարտադրությանը։ Քանի որ այստեղ անհնարին է մտնել մանրամասնությունների մեջ, ուստի ես պետք է հենվեմ միայն մի քանի ընդհանուր կշռադատությունների վրա։
Նախ՝ ես հավաքել եմ շատ փաստեր և ինքս կատարել եմ բազմաթիվ դիտումներ, որոնք ցույց են տալիս, համաձայն անասնաբույծների և այգեգործների ունեցած գրեթե ընդհանուր համոզմունքի, որ կենդանիների և բույսերի մեջ զանազան տարատեսակների կամ նույն տարատեսակի տարբեր ծագում ունեցող անհատների միջև խաչաձևումը ստացվող սերնդին հաղորդում է առանձին ուժ և պտղաբերություն։ Երկրորդ՝ ազգակցության մոտ աստիճաններ ունեցող անհատների միջև խաչաձևումը նվազեցնում է ցեղի ուժը և պտղաբերությունը։ Միայն այս փաստերը բավական էին, որպեսզի ինձ դրդեին ընդունելու որպես բնության մի ընդհանուր օրենք, որ ոչ մի օրգանական էակ սերունդների անսահման շարքում չի սահմանափակվում ինքնաբեղմնավորմամբ, այլ ընդհակառակը, մի անհատի խաչաձևումը մյուսի հետ ժամանակ առ ժամանակ, գուցե երկար ընդմիջումներից հետո, անհրաժեշտություն է։
Ելնելով այն համոզմունքից, որ բնության այդպիսի օրենք գոյություն ունի, մենք, կարծում եմ, ի վիճակի ենք ըմբռնելու փաստերի մի ամբողջ ընդարձակ խումբ, նման այն փաստերին, որոնք կշարադրվեն ստորև և որոնք ոչ մի այլ տեսակետից չեն ենթարկվում բացատրության։ Բույսերի խաչաձևմամբ զբաղվող յուրաքանչյուր այգեգործ գիտե, թե ծաղիկների թրջվելը ինչպես վնասակար կերպով է անդրադառնում բույսերի բեղմնավորման վրա, բայց և այնպես ծաղիկների ինչպիսի՜ բազմություն իր առէջքները և սպին ենթարկում է վատ եղանակի բոլոր պատահականություններին։ Հակառակ սեփական առէջքների և վարսանդի սերտ հարևանության, որը գրեթե ապահովում է ինքնաբեղմնավորում, եթե օգտակար է ժամանակ առ ժամանակ կատարվող խաչաձևումը, ապա կողմնակի ծաղկափոշու համար հնարավորին չափ ազատ մուտք ապահովելը կբացատրի մեզ մթնոլորտային խոնավությունից օրգանների այդ չպաշտպանված լինելու դրությունը։ Շատ ծաղիկներ, ընդհակառակը, ունեն պինդ ծածկված բեղմնավորման օրգաններ, ինչպես, օրինակ, թիթեռնածաղիկների մեծ ընտանիքում, բայց դրա փոխարեն միջատների այցելության համար նրանք համարյա միշտ ունեն ամենահետաքրքրական և հիանալի հարմարումներ։ Թիթեռնածաղիկ բույսերից շատերի համար մեղուների այցելությունը այնքան անհրաժեշտ է, որ նրանց պտղաբերությունը զգալիորեն նվազում է, եթե այդ այցելությունները վերացվում են։ Բայց գրեթե անհնարին է, որ միջատները կարող են ծաղկից ծաղիկ թռչել և ծաղկափոշին չտեղափոխել, բույսերին մեծ օգուտ չտալ։ Միջատները գործում են այնպես, ինչպես այգեգործի վրձինը, բավական է այդ վրձինով միայն դիպչել մի ծաղկի առէջքին, և հետո մյուսի սպիին, որպեսզի ապահովված լինի բեղմնավորումը։ Սակայն չպետք է կարծել, որ մեղուներն այդ ձևով առաջացնում են զանազան տեսակների միջև բազմաթիվ հիբրիդներ, որովհետև եթե սպիի վրա ընկնում է նույն բույսի ծաղկափոշին և մի այլ տեսակի ծաղկափոշին, այդ դեպքում առաջինը այնքան հաղթահարում է երկրորդին, որ բոլորովին վերացնում է նրա ազդեցությունը, ինչպես այդ ապացուցել է Գերտները։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83]
Երբ որևէ ծաղկի մեջ առէջքները հանկարծակի կամ դանդաղորեն մեկը մյուսի հետևից հպվում են վարսանդին, կարելի է կարծել, որ այդ հարմարանքի նպատակը բացառապես ինքնափոշոտումն ապահովելն է, և անկասկած դա այդ տեսակետից օգտակար է։ Բայց, ինչպես ցույց է տվել Կյոլռեյտերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] կծոխորի վերաբերմամբ, առէջքները շարժման մեջ դնելու համար անհրաժեշտ է հատկապես միջատների մասնակցությունը, և, բացի դրանից, հենց այդ սեռի վերաբերմամբ, որն ըստ երևույթին հատուկ հարմարումներ ունի ինքնափոշոտման համար, հայտնի է, որ եթե իրար կողքի տնկենք մերձավոր նմանություն ունեցող ձևերը կամ տարատեսակները, ապա համարյա անհնարին է լինում ստանալ մաքուր սերմնաբույսեր, քանի որ այնքան լայնորեն է տարածված խաչաձևումը նրանց մեջ։ Շատ դեպքերում ծաղկի կառուցվածքը ոչ միայն չի նպաստում ինքնափոշոտմանը, այլև երևան են գալիս հատուկ հարմարանքներ, որոնք փակում են սեփական ծաղկափոշու մուտքը դեպի սպին, որի համար օրինակներ ես կարող էի բերել Շպրենգելի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 82] և այլ հեղինակների աշխատություններից և իմ սեփական դիտողություններից։ Այսպես, օրինակ, Lobelia fulgens-ի մեջ գոյություն ունի իսկապես մի հիանալի և բարդ կառուցվածք, որի շնորհիվ ծաղկափոշու բազմաթիվ հատիկները բոլորը հեռանում են յուրաքանչյուր ծաղկի սերտաճած փոշանոթներից ավելի վաղ, քան նրա սպին պատրաստ կլինի ընդունելու նրանց, իսկ քանի որ այդ ծաղիկներին, գոնե իմ պարտեզում, միջատները երբեք չեն այցելում, ուստի նրանք երբեք էլ սերմ չեն տալիս, թեև ծաղկափոշին մի ծաղկից մյուսի վրա փոխադրելով ես առատությամբ սերմեր էի ստանում։ Լոբելիայի մի այլ տեսակը, որին այցելում են մեղուները, իմ պարտեզում նաև սերմեր է տալիս։ Այլ բազմաթիվ դեպքերում թեև հատուկ հարմարանք չի լինում, որ սպիի վրա սեփական ծաղկափոշու ընկնելուն խոչընդոտ հանդիսանա, այնուամենայնիվ, ինչպես ցույց են տվել Շպրենգելը, վերջերս Հիլդեբրանդն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 84] ու մի քանի այլ հեղինակներ և ինչպես ես ինքս էլ կարող եմ հաստատել, կամ փոշանոթները պայթում են ավելի վաղ, քան սպին պատրաստ է լինում բեղմնավորման համար, կամ սպին ծաղկափոշուց ավելի առաջ է պատրաստ լինում, այնպես որ այդպիսի բույսերը, որոնք դիխոգամ անունն են ստացել, իսկապես բաժանասեռ են և սովորաբար պիտի ենթարկվեն խաչաձևման։ Նույնը իրավացի է նաև դիմորֆ և տրիմորֆ բույսերի վերաբերմամբ, որոնց մասին հիշատակված է վերևում։ Ինչքա՜ն արտասովոր են այս բոլոր փաստերը։ Ինչքա՜ն արտասովոր է, որ միևնույն ծաղկի փոշեհատիկը և սպին, որոնք սերտ հարևանության մեջ են գտնվում կարծես ինքնաբեղմնավորումն ապահովելու համար, այդքան շատ դեպքերում փոխադարձաբար անօգուտ են լինում։ Եվ ինչքան պարզ կերպով են բացատրվում այդ բոլոր փաստերը, եթե միայն ընդունենք, որ այլ անհատների հետ խաչաձևվելը կարող է լինել օգտակար և նույնիսկ անհրաժեշտ։
Եթե թողնենք, որ կաղամբի, ամսական բողկի, սոխի և մի քանի այլ բույսերի տարատեսակներ կողք-կողքի աճեն և ցրեն իրենց սերմերը, ապա սերմնաբույսերը մեծ մասամբ, ինչպես ես կարողացա համոզվել, լինում են խառնացեղեր։ Այսպես, ես մի անգամ ստացել եմ կաղամբի 233 սերմնաբույսեր, որոնք առաջացել էին միասին աճած մի քանի տարատեսակներից և նրանցից միայն 78 պահպանել էին ցեղի հատկանիշները և այն էլ ոչ ամբողջովին մաքուր։ Բայց և յուրաքանչյուր կաղամբածաղկի վարսանդը շրջապատված է ոչ միայն իր վեց առէջքներով, այլև միևնույն բույսի վրա գտնվող մնացած ծաղիկների առէջքներով, և յուրաքանչյուր ծաղկի ծաղկափոշին հեշտությամբ ընկնում է նրա սպիի վրա առանց միջատների աջակցության, որովհետև ես նկատել եմ, որ միջատներից խնամքով պաշտպանված բույսերը տալիս են նորմալ թվով պատիճներ։ Հապա ի՞նչ կերպով այդպիսի մեծ թվով սերմնաբույսեր խառնացեղ են լինում։ Այդ պետք է կախված լինի նրանից, որ այլ տարատեսակի ծաղկափոշին հաղթահարում է տվյալ ծաղկի սեփական ծաղկափոշուն, և դրանում նույնպես արտահայտվում է այն ընդհանուր օրենքը, որ նույն տեսակի այլ անհատների հետ խաչաձևումը օգտակար է։ Երբ խաչաձևվում են տարբեր տեսակներ, բոլորովին հակառակ հետևանք է ստացվում, որովհետև սեփական ծաղկափոշին համարյա միշտ հաղթահարում է օտար ծաղկափոշուն, բայց այդ հարցին մենք կվերադառնանք հաջորդ գլուխներից մեկում։
Եթե օրինակի համար մենք կանգ առնենք անհամար ծաղիկներով պատած մի ծառի վրա, ապա կարող են առարկել, որ ծաղկափոշին միայն հազվադեպ կարող էր մի ծառից մյուսի վրա բերվել, ամենաշատը՝ ծաղկափոշին, կարող էր միևնույն ծառի ծաղկից ծաղիկ տարվել, իսկ միևնույն ծառի ծաղիկները միայն շատ սահմանափակ իմաստով կարելի է ինքնուրույն անհատներ համարել։ Ես կարծում եմ, որ այս առարկությունը բավական ծանրակշիռ է, բայց բնությունը զգալի չափով վերացրել է այն, ծառերին տալով բաժանասեռ ծաղիկներ առաջացնելու ուժեղ ձգտում։ Երբ սեռերը բաժան են, այդ դեպքում, եթե նույնիսկ միևնույն ծառի վրա միաժամանակ կան փոշանոթային և վարսանդային ծաղիկներ, ծաղկափոշին համենայն դեպս պետք է տեղափոխվի ծաղկից ծաղիկ, իսկ դրա շնորհիվ կավելանան ծաղկափոշու ծառից ծառ տեղափոխվելու շանսերը։ Այն կանոնը, որ ամենաբազմազան ընտանիքների պատկանող ծառերին ավելի հատուկ է սեռերի բաժանվածությունը, քան մյուս բույսերին, Անգլիայի համար արդարանում է։ Իմ խնդրանքով դոկտոր Հուկերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] կազմել է Նոր Զելանդիայի բուսականության ծառատեսակների ցուցակը, իսկ Ազա-Գրեյը՝[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 68] Միացյալ Նահանգների բուսականության, և հետևանքը համապատասխանում էր իմ ակնկալություններին։ Մյուս կողմից, Հուկերը տեղեկացնում է ինձ, որ այդ կանոնը Ավստրալիայի վերաբերմամբ չի արդարանում. բայց եթե ավստրալիական ծառերի մեծ մասը դիխոգամ է, ապա հետևանքը նույնը կլինի, ինչպես եթե նրանք ունենային բաժանասեռ ծաղիկներ։ Ես բերեցի այս դիտողությունները ծառերի մասին միայն նրա համար, որպեսզի ուշադրություն հրավիրեմ այդ հարցի վրա։
Անցողակի կանգ առնենք կենդանիների վրա. ցամաքային բազմաթիվ տեսակներ, ինչպես, օրինակ, ցամաքային կակղամորթները և անձրևաորդերը, հերմաֆրոդիտ են, բայց նրանք բոլորն էլ զուգավորվում են։ Ես մինչև այժմ չեմ գտել մի ցամաքային կենդանի, որը կարողանար ինքն իրեն բեղմնավորել։ Այդ զարմանալի փաստը, որը այդպիսի հակադրություն է ներկայացնում ցամաքային բույսերի նկատմամբ, հասկանալի է միայն խաչաձևման անհրաժեշտության տեսակետից, որովհետև բեղմնավորող սկզբնահիմքի յուրահատկության պատճառով չկան այնպիսի եղանակներ, ինչպես միջատների կամ քամու գործողությունն է բույսերի վերաբերմամբ, որոնք կարողանային ժամանակ առ ժամանակ խաչաձևում առաջացնել առանց երկու անհատների անմիջական մերձեցման։ Ջրային կենդանիների մեջ հանդիպում են բազմաթիվ ինքնաբեղմնավորվող հերմաֆրոդիտներ, բայց այստեղ ջրի հոսանքը ակներև միջոց է հանդիսանում խաչաձևման համար այնպես, ինչպես և ծաղիկների վերաբերմամբ, ինձ չի հաջողվել նույնիսկ ամենախոշոր հեղինակություններից մեկի պրոֆեսոր Հյոքսլիի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 27] հետ խորհրդակցելուց հետո գտնել մի հատ հերմաֆրոդիտ կենդանի, որի վերարտադրության օրգանները այն աստիճանի փակ լինեին, որպեսզի հիմք ունենայինք ապացուցելու նրա մեջ դրսից մուտք գործելու կամ կողմնակի անհատի ազդեցության ֆիզիկական անհնարինությունը։ Փնջոտնյա խեցգետինները երկար ժամանակ այդ տեսակետից, ինձ թվում էր, թե մեծ դժվարություն էին ներկայացնում, բայց շնորհիվ մի բախտավոր պատահականության՝ ինձ հաջողվեց ցույց տալ, որ երկու անհատներ, որոնցից յուրաքանչյուրը թեև ներկայացնում է ինքնաբեղմնավորվող հերմաֆրոդիտ, երբեմն միմյանց հետ խաչաձևվում են։
Հավանորեն բնախույզների մեծամասնությանը զարմացրել է այն, կարծես թե, անորմալ փաստը, որ ինչպես կենդանիների, այնպես էլ բույսերի մոտ նույն ընտանիքի և նույնիսկ սեռի սահմաններում իրենց ամբողջ կազմվածքով նման տեսակները լինում են մերթ հերմաֆրոդիտ, մերթ բաժանասեռ։ Բայց եթե իրականում բոլոր հերմաֆրոդիտները ժամանակ առ ժամանակ խաչաձևվում են, ապա նրանց և բաժանասեռ օրգանիզմների միջև եղած տարբերությունը, գոնե ինչքանով որ գործը վերաբերում է նրանց սեռական գործունեությանը, շատ էլ մեծ չէ։
Հիմնվելով այդ զանազան կշռադատումների և բազմաթիվ հատուկ փաստերի վրա, որ ես հավաքել եմ, բայց տեղի սղության պատճառով չեմ կարող այստեղ բերել, կարելի է, ըստ երևույթին, եզրակացնել, որ ինչպես կենդանիների, այնպես էլ բույսերի մոտ առանձին անհատների միջև ժամանակ առ ժամանակ կատարվող խաչաձևումը բնության լայն տարածված, եթե ոչ ընդհանուր օրենքն է։
ՀԱՆԳԱՄԱՆՔՆԵՐ, ՈՐՈՆՔ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՄԻՋՈՑՈՎ ՆՊԱՍՏՈՒՄ ԵՆ ՆՈՐ ՁԵՎԵՐԻ ԱՌԱՋԱՑՄԱՆԸ
Սա մի չափազանց կնճռոտ խնդիր է։ Փոփոխականության մեծ աստիճանը, իսկ փոփոխականություն տերմինի մեջ միշտ ներառվում են անհատական տարբերությունները, ըստ երևույթին, կնպաստի ընտրության ներգործությանը։ Անհատների մեծ թիվը, ավելացնելով տվյալ ժամանակաշրջանում օգտակար փոփոխությունների երևան գալու հավանականությունները, կարող է փոխհատուցել յուրաքանչյուր առանձին անհատի մեջ առաջացող փոփոխականության ավելի նվազ աստիճանը և դա, իմ կարծիքով, հաջողության կարևոր տարրն է։ Թեև բնությունը բնական ընտրության գործունեության արտահայտման համար տրամադրում է երկար ժամանակաշրջաններ, բայց և այնպես նրանք անսահման երկար չեն, որովհետև օրգանիզմների ընդհանուր ձգտումն է իրենց համար տեղ գրավել բնության տնտեսության մեջ, և եթե որևէ տեսակ չի փոփոխվի ու չի կատարելագործվի իր մրցակիցների հետ համահավասար չափով, ապա նա կոչնչանա։ Եթե ծնողների օրգանիզմների օգտակար փոփոխությունները չեն ժառանգվում թեկուզ մի քանի հաջորդների կողմից, ապա բնական ընտրությունն անզոր է որևէ բան անել։ Նախկին հատկանիշներին վերադառնալու ձգտումը կարող է երբեմն արգելակել բնական ընտրությունը կամ խոչընդոտ հանդիսանալ նրա ներգործությանը, բայց եթե այդ հատկությունը չի խանգարել մարդուն արհեստական ընտրության միջոցով առաջացնել բազմաթիվ ցեղեր, ապա ինչո՞ւ նա ուժեղ հակազդեցություն պիտի ցույց տար բնական ընտրությանը։
Մեթոդիկ ընտրության ժամանակ անասնաբույծը միանգամայն որոշակի նպատակով է ընտրում, և եթե թույլ տա ազատ խաչաձևում, ապա նրա աշխատանքը բոլորովին կկորչի։ Եթե մեծ թվով մարդիկ ցեղը փոփոխելու որևէ մտադրություն չունենալով, այլ առաջնորդվելով կատարելագործության ընդհանուր գաղափարով, հոգ տանեն այն մասին, որ ձեռք բերեն ու բազմացնեն լավագույն կենդանիներին, ապա այդ անգիտակցական ընտրության հետևանքով կստացվի ցեղի դանդաղ, բայց հաստատուն կատարելագործություն, չնայած նրան, որ ընտրված անհատների մեկուսացում գոյություն չունի։ Այդպես էլ բնության մեջ է. մի սահմանափակ մարզի սահմաններում, որտեղ բնակչությունը դեռ չի լցրել բոլոր ազատ տեղերը, հարկ եղած ուղղությամբ թեկուզ տարբեր աստիճաններով փոփոխվող բոլոր անհատները կձգտեն պահպանվել։ Բայց եթե մարզը մեծ է, նրա առանձին մասերը համարյա հաստատապես կներկայացնեն տարբեր կենսապայմաններ, եթե տեսակը կենթարկվի փոփոխության զանազան մասերում միաժամանակ, այդ դեպքում առաջացած նոր տարատեսակները կխաչաձևվեն սահմանագծային գոտիներում։ Վեցերորդ գլխում մենք կտեսնենք, որ միջանկյալ տարատեսակները, որոնք իրենց բնակված մարզում գրավում են նաև միջանկյալ դրություն, վերջ ի վերջո դուրս կմղվեն հարևան տարատեսակների կողմից։ Խաչաձևումը ամենից ավելի ազդում է այն կենդանիների վրա, որոնք ամեն մի ծննդի համար զուգավորվում են, թափառական կյանք են վարում և շատ էլ արագ չեն բազմանում։ Այդ պատճառով նման կենսակերպ ունեցող կենդանիների, ինչպես, օրինակ, թռչունների տարատեսակները սովորաբար կտեղաբաշխվեն առանձին անջատ երկրներում. այդ այդպես էլ լինում է իրականում։ Միայն ժամանակ առ ժամանակ խաչաձևվող հերմաֆրոդիտ օրգանիզմների մոտ, ինչպես նաև այն կենդանիների մոտ, որոնք ամեն մի ծննդի համար խաչաձևվում են, բայց թափառական կյանք չեն վարում և արագ կերպով են բազմանում, նոր կատարելագործված տարատեսակը կարող է արագ կերպով առաջանալ ամեն մի վայրում և այնտեղ պահպանվել, իսկ հետո տարածվել, այնպես որ նրա անհատները հաճախակի կխաչաձևվեն միմյանց հետ։ Դրա հիման վրա պարտիզագործները միշտ գերադասում են պահել մեծ խմբերով աճող բույսերից ստացված սերմերը, որովհետև այդ պայմաններում մեծ չափով պակասում են խաչաձևման շանսերը։
Նույնիսկ այն կենդանիների նկատմամբ, որոնք յուրաքանչյուր ծննդի համար զուգավորվում և արագ չեն բազմանում, մենք չպետք է ենթադրենք, որ ազատ խաչաձևումը միշտ կոչնչացնի բնական ընտրության հետևանքները, որովհետև ես կարող եմ ներկայացնել մեծ թվով փաստեր, որոնք ցույց են տալիս, որ միևնույն մարզի սահմաններում միևնույն կենդանու երկու տարատեսակները կարող են երկար ապրել առանց խառնվելու իրար հետ, արդյոք այն պատճառով, որ բազմանում են տարվա տարբեր ժամանակներում, կամ թե այն պատճառով, որ մի տարատեսակի անհատները գերադասում են միմյանց հետ զուգավորվել։
Խաչաձևումը կարևոր դեր է կատարում բնության մեջ, որովհետև պահպանում է միևնույն տեսակի կամ միևնույն տարատեսակի անհատների մեջ հատկանիշների միակերպությունը և կայունությունը։ Նա հավանորեն ավելի ներգործող ազդեցություն կթողնի այն կենդանիների վրա, որոնք ամեն մի ծննդի համար զուգավորվում են, բայց ինչպես արդեն ասված է, մենք լիակատար հիմք ունենք ենթադրելու, որ բոլոր կենդանիները և բոլոր բույսերը ժամանակ առ ժամանակ ենթարկվում են խաչաձևման։ Եթե այդ տեղի ունենա նույնիսկ երկար ժամանակամիջոցներից հետո, այդ դեպքում այդ խաչաձևումից ծագում առած էակներն իրենց ուժով և պտղաբերությամբ այնքան կգերազանցեն երկարատև ինքնաբեղմնավորման արդյունք հանդիսացող էակներին, որ գերապրելու և բազմանալու ավելի շանսեր կստանան։ Այսպիսով, վերջ ի վերջո խաշաձևման ազդեցությունը նույնիսկ երկար ժամանակամիջոցներից հետո շատ մեծ կլինի։ Ինչ վերաբերում է ամենաստորին կարգի օրգանական էակներին, որոնք սեռական ճանապարհով չեն բազմանում և ոչ էլ կոնյուգացիայի են ենթարկվում, հետևաբար և խաչաձևման չեն ենթարկվում, ապա նրանց հատկանիշների միակերպությունը կարող է պահպանվել պայմանների անփոփոխության դեպքում միայն ժառանգականության սկզբունքի շնորհիվ և բնական ընտրության ներգործության շնորհիվ, որը կոչնչացնի բոլոր այն անհատներին, որոնք շեղվում են իսկական տիպից։ Իսկ եթե կենսապայմանները փոփոխվում են և ձևը ենթարկվում է փոփոխության, այդ դեպքում կառուցվածքի միակերպությունը կարող է պահպանվել փոփոխված սերնդի մեջ միայն բնական ընտրության միջոցով, որը պահպանում է նման օգտակար փոփոխությունները։
Մեկուսացումը նույնպես կարևոր տարր է հանդիսանում բնական ընտրության միջոցով տեսակների փոփոխության պրոցեսում։ Սահմանափակված կամ մեկուսացած մի մարզում, եթե նա շատ մեծ չէ, կյանքի օրգանական և անօրգանական պայմաններն ընդհանրապես գրեթե միանման կլինեն, այնպես որ բնական ընտրությունը կձգտի փոփոխել միևնույն տեսակին պատկանող բոլոր ձևափոխվող անհատներին միևնույն ուղղությամբ։ Շրջապատող մարզերի բնակիչների հետ խաչաձևումը նույնպես կվերացվի։ Մորից Վագները[41][ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 85] այդ խնդրի վերաբերմամբ վերջերս հրատարակել է մի հետաքրքիր աշխատություն և ցույց է տվել, որ մեկուսացումը որպես նոր առաջացած տարատեսակների խաչաձևման խոչընդոտ շատ ավելի կարևոր նշանակություն ունի, քան ես ենթադրել եմ։ Սակայն իմ կողմից արդեն մատնանշված պատճառների հիման վրա ես ոչ մի դեպքում չեմ կարող համաձայնվել այդ բնախույզի հետ, որ միգրացիան և մեկուսացումը անհրաժեշտ տարրեր են նոր տեսակների առաջացման պրոցեսում։ Մեկուսացման նշանակությունը նաև մեծ է այն տեսակետից, որ երկրի որևէ ֆիզիկական փոփոխությունների ժամանակ, ինչպիսիք են՝ ցամաքի բարձրացումը, կլիմայի փոփոխությունը և այլն, նա կկանխի ավելի լավ հարմարված օրգանիզմների ներգաղթը և, այսպիսով, երկրի բնական տնտեսության մեջ նոր ազատ վայրեր կմնան, որոնք կլցվեն հին բնակիչների փոփոխված սերունդներով։ Վերջապես մեկուսացումը նոր առաջացած տարատեսակին անհրաժեշտ ժամանակ է տրամադրում դանդաղ, աստիճանական կատարելագործման համար, որը երբեմն կարող է շատ կարևոր լինել։ Եթե մեկուսացած տարածությունը շատ փոքր է, կամ այն պատճառով, որ շրջապատված է խոչընդոտներով,— կամ թե այն պատճառով, որ նա ունի չափազանց բացառիկ ֆիզիկական պայմաններ,— այդ դեպքում նրա բնակիչների ընդհանուր թիվը փոքր կլինի, և այդ կդանդաղեցնի նոր տեսակների առաջացումը բնական ընտրության միջոցով, որովհետև կնվազեն բարենպաստ փոփոխությունների երևան գալու շանսերը։
Ժամանակի երկարատևությունը ինքնին ոչ օժանդակում և ոչ էլ խոչընդոտ է հանդիսանում բնական ընտրությանը։ Ես այդ հայտարարում եմ այն պատճառով, որ բոլորովին սխալ կերպով պնդել են, իբրև թե ես ժամանակի տարրին տալիս եմ ամենակարող նշանակությունը տեսակների փոփոխության պրոցեսում,— իբրև թե բոլոր կենսական ձևերը ժամանակի ընթացքում անխուսափելիորեն և անընդհատ փոփոխվում են մի ինչ-որ նրանց բնատուր օրենքի շնորհիվ։ Ժամանակի երկարատևությունը նշանակություն ունի և կարևոր նշանակություն միայն այնքան, որքան նա ավելացնում է բարենպաստ փոփոխությունների երևան գալու, նրանց ընտրության, կուտակման և ամրացման հավանականությունները։ Ժամանակի տևականության հետ միասին աճում է նաև կյանքի ֆիզիկական պայմանների անմիջական ներգործության ազդեցությունը յուրաքանչյուր օրգանիզմի ընդհանուր կերտվածքի վրա:
Եթե այս դիտողությունների արդարացիությունը ստուգելու համար մենք դառնանք բնությանը և կանգ առնենք մի որևէ փոքրիկ մեկուսացած տերիտորիայի վրա, օրինակ՝ որևէ օվկիանոսային կղզու վրա, ապա թեև նրա վրա ապրող տեսակների թիվը, ինչպես մենք կտեսնենք աշխարհագրական բաշխման գլխում, մեծ չէ, այնուամենայնիվ նրանց մեծ մասը էնդեմիկ է, այսինքն առաջացել է այդ տեղում, այլ ոչ թե երկրագնդի որևէ այլ կետում։ Այսպիսով, օվկիանոսային կղզին առաջին հայացքից առանձնապես բարենպաստ կթվա նոր տեսակների առաջացման համար։ Բայց հանելով այդ եզրակացությունը, մենք հեշտությամբ կարող ենք սխալվել, որովհետև լուծելու համար այն հարցը, թե օրգանական նոր ձևերի առաջացման համար ո՛րն է ավելի նպաստավոր՝ արդյոք փոքրիկ մեկուսացած մարզը, թե՞ ամբողջ մայր ցամաքի մի լայնածավալ արձակ տերիտորիան,— մենք պետք է դրանք համեմատենք իրար հետ հավասար ժամանակաշրջանների ընթացքում, իսկ այդ մենք չենք կարող անել։
Թեպետև մեկուսացումը մեծ նշանակություն ունի նոր տեսակների առաջացման գործում, այնուամենայնիվ, ընդհանրապես ես հանգում եմ այն համոզման, որ մարզի ընդարձակությունը է՛լ ավելի մեծ նշանակություն ունի, մանավանդ այնպիսի տեսակների առաջացման պրոցեսում, որոնք կարող կլինեն երկար ժամանակ պահպանվել և լայնորեն տարածվել։ Մեծ և բաց տարածության վրա ոչ միայն ավելանում են բարենպաստ փոփոխությունների երևան գալու շանսերը՝ շնորհիվ միևնույն տեսակի անհատների բազմաքանակության, որոնց կարող է կերակրել տվյալ երկիրը, այլև այն պատճառով, որ գոյության պայմաններն իրենք անհամեմատ ավելի բարդ են արդեն գոյություն ունեցող տեսակների բազմաքանակության հետևանքով, իսկ եթե այդ բազմաթիվ տեսակներից մի քանիսը կփոփոխվեն և կկատարելագործվեն, այդ դեպքում նաև մնացածները պետք է համապատասխանորեն փոփոխվեն, այլապես նրանք կբնաջնջվեն։ Յուրաքանչյուր նոր ձև, հենց որ նա ձեռք է բերում զգալի առավելություններ, ի վիճակի կլինի տարածվելու բաց և հարթ երկրում և այդպիսով մրցության մեջ կմտնի այլ բազմաթիվ ձևերի հետ։ Բացի դրանից, ընդարձակ տարածությունները թեև այժմ անընդմիջվող են լինում, բայց անցյալում կարող էին մակերևույթի տատանումների հետևանքով ենթարկվել մասնատման, այնպես որ նաև մեկուսացման ընձեռած բարենպաստ պայմանները կարող էին որոշ չափով իրենց աջակցությունը ցույց տալ։ Ես գալիս եմ այն եզրակացության, որ թեև փոքրիկ, սահմանափակված մարզերը որոշ տեսակետից ներկայացրել են խիստ բարենպաստ պայմաններ նոր տեսակների առաջացման համար, այնուամենայնիվ լայնածավալ մարզերում փոփոխությունները մեծ մասամբ կատարվել են ավելի արագ և, որն ավելի ևս կարևոր է, մեծ տարածությունների վրա նոր առաջացած և արդեն շատ մրցակիցների հաղթած ձևերը ավելի ընդունակ կլինեն լայնորեն տարաբնակվելու և ամենամեծ թվով տեսակների ու տարատեսակների սկիզբ կտան։ Ուրեմն նրանք ավելի աչքի ընկնող դեր են խաղացել օրգանական աշխարհի փոփոխական պատմության մեջ։
Այդ տեսակետների համաձայն թերևս մեզ հասկանալի կդառնան որոշ փաստեր, որոնց մասին խոսք կլինի աշխարհագրական տեղաբաշխման գլխում, ինչպես, օրինակ, այն փաստը, որ ավելի քիչ ընդարձակ մայր ցամաքի Ավստրալիայի բնակիչները չեն դիմանում իրենց նեղողների, ավելի ընդարձակ Եվրոպա-ասիական մարզից եկվորների ճնշմանը։ Դրանով է բացատրվում նաև այն փաստը, որ մայր ցամաքների բնակիչները այնպիսի հեշտությամբ նատուրալիզացիայի են ենթարկվել ամենուրեք կղզիների վրա։ Փոքրիկ կղզու վրա գոյության համար մղվող մրցությունը ավելի պակաս կատաղի է եղել, փոփոխականությունը և բնաջնջումը ոչ այնքան ուժեղ։ Այստեղից մեզ համար հասկանալի է, թե ինչու Մադեյրա կղզու բուսականությունը, ըստ Օսվալդ Հեերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 38] վկայության, որոշ չափով հիշեցնում է Եվրոպայի բնաջնջված բուսականությունը երրորդական դարաշրջանում։ Բոլոր քաղցրահամ ջրավազանները ի մի առած ավելի փոքր տարածություն են ներկայացնում ծովի կամ ցամաքի համեմատությամբ։ Հետևաբար, քաղցրահամ ջրերի բնակիչների միջև մրցակցությունը մյուսների համեմատությամբ ավելի թույլ է եղել, նոր ձևերը ավելի դանդաղ էին առաջանում, և հին ձևերն էլ ավելի դանդաղ էին ոչնչանում։ Եվ հենց քաղցրահամ ջրերում մենք հանդիպում ենք գանոիդ ձկների յոթ սեռերին՝ երբեմնի տիրապետող կարգի մնացորդներին։ Քաղցրահամ ջրերում էլ մենք հանդիպում ենք երկրի վրա գոյություն ունեցող ձևերից ամենատարօրինակներին՝ բադակտուցին և լեպիդոսիրենին, որոնք բրածո ձևերի նման որոշ չափով շաղկապում են էակների սանդուղքի վրա միմյանցից հեռու կանգնած խմբերը։ Այդ անկանոն ձևերը կարող են անվանվել կենդանի բրածոներ. նրանք պահպանվել են մինչև այժմ այն պատճառով, որ ապրել են մեկուսացած մարզում և ենթարկվել են ավելի քիչ բազմազան և, հետևաբար, ավելի քիչ կատաղի մրցության։
Ամփոփում եմ, այն հանգամանքները, որքան այդ թույլ կտա հարցի չափազանց խրթինությունը, որոնք բնական ընտրության միջոցով նոր տեսակների առաջացման համար բարենպաստ և անբարենպաստ են։ Ես գալիս եմ այն եզրակացության, որ ցամաքային օրգանիզմների համար ցամաքային մեծ մի մարզ, որի մակերևույթը բազմիցս ենթարկվել է տատանումների, պետք է ամենանպաստավոր լիներ բազմաթիվ այնպիսի նոր ձևերի առաջացման համար, որոնք հարմարված են երկարատև գոյության և լայն տարաբնակման։ Քանի դեռ այդ մարզը գոյություն է ունեցել որպես մայր ցամաք, նրա բնակիչները պետք է աչքի ընկնեին անհատների և ձևերի բազմաքանակությամբ և ենթարկվեին կատաղի մրցության։ Երբ ցամաքի իջնելու հետևանքով նա բաժանվել է առանձին կղզիների, նրանցից յուրաքանչյուրի վրա այնուամենայնիվ մնացել են ամեն մի տեսակից զգալի թվով անհատներ. կանխվել է խաչաձևման հնարավորությունը նոր առաջացած տեսակների տարածման սահմաններում. վերացվել է ներգաղթի հնարավորությունը, այնպես որ ֆիզիկական պայմանների որևէ փոփոխության դեպքում յուրաքանչյուր կղզու վրա օրգանական կարգում առաջացած նոր ազատ տեղերը պետք է լցվեին հին բնակիչների փոփոխված սերունդներով. և վերջապես, ապահովվել է ժամանակի անհրաժեշտ տևողությունը, որպեսզի յուրաքանչյուր կղզու տարատեսակ կարողանար բավարար չափով փոփոխվել ու կատարելագործվել։ Երբ մակերևույթի կրկնական բարձրացման հետևանքով կղզիները միանալով՝ նորից կազմել են մայր ցամաք, վերսկսվել է ուժեղ մրցությունը, ամենից ավելի կատարելագործված և հարմարված տարատեսակները հնարավորություն են ստացել լայնորեն տարածվելու. տեղի է ունեցել ավելի նվազ կատարելագործված ձևերի զգալի ոչնչացում, և նոր առաջացած ցամաքի զանազան բնակիչների հարաբերական թիվը կրկին փոփոխվել է. այսպիսով, բնական ընտրության համար նոր տեսակների առաջացման ու հետագա կատարելագործման իմաստով գործունեության նոր ասպարեզ է բացվել։
Ես միանգամայն ընդունում եմ, որ բնական ընտրությունը ներգործում է չափազանց դանդաղ կերպով։ Նա կարող է ներգործել միայն այն ժամանակ, երբ մի երկրի օրգանական կարգում բացվում են ազատ տեղեր, որոնք կարող են հաջողությամբ գրավվել նրա բնակիչների փոփոխված ձևերով։ Այդպիսի ազատ տեղերի երևան գալը հաճախ կախված է ֆիզիկական պայմանների փոփոխությունից, որի համար սովորաբար շատ ժամանակ է հարկավոր, և միաժամանակ ավելի լավ հարմարված ձևերի ներգաղթը կանխելուց։ Հենց որ հին բնակիչներից մի քանիսը կսկսեն փոփոխվել, կխախտվեն նրանց փոխհարաբերությունները մնացածների հետ, իսկ դա կստեղծի նոր տեղեր ավելի լավ հարմարված ձևերի համար. բայց այդ բոլորը տեղի կունենա շատ դանդաղորեն։ Թեև միևնույն տեսակի բոլոր անհատները փոքր ինչ տարբերվում են մեկը մյուսից, այնուամենայնիվ հավանորեն շատ ժամանակ կանցնի, մինչև որ երևան կգան օրգանիզմի զանազան մասերի նրանց համար ձեռնտու փոփոխությունները։ Այդ հետևանքի հայտնվելը հաճախ մեծ չափով կդանդաղի ազատ խաչաձևման պատճառով։ Շատերը կբացականչեն, որ այդ բոլոր պատճառները միանգամայն բավական են, որպեսզի չեզոքացնեն բնական ընտրության ուժը։ Ես այդպես չեմ մտածում։ Բայց ես կարծում եմ, որ բնական ընտրությունը կներգործի շատ դանդաղ կերպով միայն ժամանակի երկար ընդմիջումներից հետո և միայն տվյալ երկրի բնակիչներից քչերի վրա միաժամանակ։ Այնուհետև ես կարծում եմ, որ ընտրության այդ դանդաղ ընդմիջվող ներգործությունը լավ համապատասխանում է այն բանին, ինչ որ ուսուցանում է մեզ երկրաբանությունը երկրագնդի նախկին բնակչության փոփոխության տևողության և եղանակի վերաբերմամբ։
Ինչքան էլ ուզում է դանդաղորեն կատարվեր ընտրության պրոցեսը, եթե թույլ մարդը կարողացել է արհեստական ընտրության միջոցով հասնել այնպիսի մեծ արդյունքների, ապա ես չեմ տեսնում վերջնակետը այն փոփոխականության համար, օրգանիզմների մեկը մյուսի և կյանքի ֆիզիկական պայմանների վերաբերմամբ ունեցած հարմարումների այն գեղեցկության և բարդության համար, որոնք կարող էին իրագործվել երկար ժամանակի ընթացքում բնական ընտրության շնորհիվ, այսինքն՝ ամենից ավելի հարմարվածների գերապրելու շնորհիվ։
ՄԱՀԱՋՆՋՈՒՄԸ, ՈՐ ՊԱՏՃԱՌՈՒՄ է ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅՈՒՆԸ
Այս խնդիրը մենք ավելի մանրամասն քննարկման կենթարկենք երկրաբանությանը նվիրված գլխում, բայց նրա մասին անհրաժեշտ է հիշատակել նաև այստեղ բնական ընտրության հետ նրա ունեցած սերտ կապի պատճառով։ Բնական ընտրությունը ներգործում է միայն պահպանելով այն փոփոխությունները, որոնք որևէ տեսակետից օգտակար են և, հետևաբար, արմատանում են։ Բոլոր օրգանական էակների երկրաչափական պրոգրեսիայով արագ կերպով բազմանալու հետևանքով յուրաքանչյուր մարզ արդեն ծայր աստիճան լցված է բնակիչներով, իսկ այստեղից բխում է, որ որքան հարմարված ձևերը թվով կշատանան, այնքան ավելի քիչ հարմարվածները սովորաբար թվով կնվազեն և կդառնան հազվագյուտ։ Հազվադեպությունը, ինչպես ուսուցանում է մեզ երկրաբանությունը, մահաջնջման նախագուշակն է։ Մեղ համար պարզ է, որ սակավաթիվ անհատներ ունեցող յուրաքանչյուր ձև կլիմայական զգալի տատանումների ժամանակ կամ նրա թշնամիների ժամանակավոր բազմացման հետևանքով վերջնականապես անհետանալու մեծ շանսեր ունի։ Բայց մենք կարող ենք ավելի հեռու գնալ, նոր ձևերի առաջացման հետ զուգընթաց, եթե միայն չընդունենք, որ տեսակային ձևերի թիվը կարող է անսահմանորեն մեծանալ, հին ձևերից շատերը պետք է մահաջնջվեն։ Բայց որ տեսակային ձևերի թիվը իրականում անսահմանորեն չի աճել, դրանում մեզ պարզորոշ համոզում է երկրաբանությունը, և մենք այժմ կփորձենք բացատրել, թե ինչու աշխարհում գոյություն ունեցող տեսակների թիվը անչափ չի մեծացել։
Մենք տեսանք, որ անհատների թվով ամենահարուստ տեսակները ամենաշատ շանսեր ունեն տվյալ ժամանակաշրջանում բարենպաստ փոփոխություններ դրսևորելու համար։ Դրան ապացույց կարող են ծառայել այն փաստերը, որոնք բերված են երկրորդ գլխում և ցույց են տալիս, որ ամենից ավելի սովորական, լայնորեն տարաբնակված և տիրապետող տեսակները տալիս են տարատեսակների ամենամեծ թիվը։ Ուրեմն՝ հազվագյուտ տեսակները յուրաքանչյուր տվյալ ժամանակաշրջանում կփոփոխվեն ու կկատարելագործվեն ավելի դանդաղ կերպով և գոյության կռվում կպարտվեն ավելի սովորական տեսակների փոփոխված ու կատարելագործված հաջորդներից։
Այս զանազան կշռադատումներից, ես կարծում եմ, անխուսափելիորեն բխում է այն, որ ժամանակի ընթացքում բնական ընտրության գործունեությամբ ինչպես որ առաջանում են նոր տեսակներ, այնպես էլ մյուս տեսակները կսակավանան և, վերջապես, կանհետանան։ Այն ձևերը, որոնք ամենամոտ մրցության մեջ են մտնում փոփոխվող ու կատարելագործվող ձևերի հետ, անշուշտ ավելի շատ կտուժեն։ Գոյության կռվին նվիրված գլխում մենք տեսանք, որ ամենից ավելի կատաղի մրցություն պետք է տեղի ունենա ամենից ավելի մերձավոր ձևերի միջև՝ միևնույն տեսակի տարատեսակների, կամ միևնույն սեռի տեսակների, կամ թե միմյանց մոտիկ սեռերի միջև, որովհետև այդ ձևերը կունենան գրեթե միանման կառուցվածք, ընդհանուր կացութաձև և սովորություններ։ Այստեղից բխում է, որ յուրաքանչյուր նոր առաջացող տարատեսակ կամ յուրաքանչյուր նոր տեսակ պետք է ավելի ուժեղ ճնշում գործի իր մերձավոր ազգակիցների վրա և ձգտի նրանց ոչնչացնել։ Այդ ոչնչացման պրոցեսը մենք նկատում ենք նաև ընտանի ցեղերի վերաբերմամբ՝ ամենակատարելագործված ձևերի ընտրության հետևանքով։ Կարելի է բերել հետաքրքրական շատ դեպքեր հաստատելու համար այն, թե ինչպես տավարի, ոչխարների և այլ կենդանիների նոր ցեղերը կամ ծաղիկների նոր սորտերը արագ կերպով դուրս են մղել ավելի հին և ավելի քիչ կատարելագործված ձևերը։ Օրինակ, պատմականորեն հայտնի է, որ հնուց Յորկշիրում ապրող սև տավարը փոխարինվել է «լոնգհորներով», իսկ այս վերջիններս «բնաջնջվել են շորտհորների կողմից (ես բերում եմ մի գյուղատնտեսական հեղինակի իսկական խոսքերը) ասես թե ինչ-որ ժանտախտից»։
ՀԱՏԿԱՆԻՇՆԵՐԻ ՏԱՐԱՄԻՏՈՒՄԸ
Այն սկզբունքը, որ ես նշում եմ այս տերմինով, խիստ կարևոր է և, ինչպես ինձ թվում է, բացատրում է մի քանի էական փաստեր։ Նախ, նույնիսկ ցայտուն կերպով արտահայտված տարատեսակները թեև տեսակների հետ ընդհանուր հատկանիշներ ունեն,— ինչպես այդ հետևում է այն դժվարություններից, որ հաճախ ծագում են նրանց գնահատական տալիս,— ի հարկե միմյանցից անհամեմատ ավելի քիչ են տարբերվում, քան լավ, ցայտուն կերպով արտահայտված տեսակները իրարից։ Եվ, այնուամենայնիվ, ըստ իմ հայեցակետի, տարատեսակները միայն տեսակներ են, որոնք գտնվում են գոյացման պրոցեսում, կամ ինչպես ես անվանել եմ նրանց՝ սկզբնավորվող տեսակներ։ Բայց ինչպե՞ս տարատեսակների միջև եղած թույլ տարբերությունը հասնում է մինչև տեսակային տարբերության չափերին։ Որ այդ ձևափոխումը տեղի է ունենում իրականում, մենք պետք է եզրակացնենք այն փաստից, որ բնության մեջ ցրված անհամար տեսակների մեծ մասն ունի լավ արտահայտված տարբերություններ, մինչդեռ տարատեսակները, այդ ապագա, լավ արտահայտված տեսակների ենթադրելի նախատիպերը և նախահայրերը ներկայացնում են միայն թույլ և աղոտ կերպով նշմարելի տարբերությունները։ Մի պարզ դեպք, ինչպես սովորաբար ասում են, անշուշտ կարող է պատճառ լինել, որ մի որոշ տարատեսակ շեղվի իր նախահայրերի հատկանիշներից, իսկ նրա սերունդն էլ իր հերթին նույն ուղղությամբ և ավելի ևս մեծ չափով շեղվի իր ծնողներից, բայց միայն այդ պրոցեսը անբավարար կլիներ բացատրելու համար այն սովորական, խոր տարբերությունը, որ ներկայացնում են միևնույն սեռին պատկանող տեսակները[42]։
Այս դեպքում ևս, ինչպես և միշտ, ես աշխատել եմ այդ հարցը լուսաբանել ելնելով այն փաստերից, որոնք վերաբերում են մեր ընտանի ցեղերին։ Մենք այստեղ էլ կգտնենք համանմանություն։ Յուրաքանչյուր ոք, ի հարկե, կհամաձայնվի, որ այնպես խիստ կերպով միմյանցից տարբերվող ցեղեր, ինչպիսիք են շորտհորները և հերեֆորդյան տավարը, արշավաձին և ծանրաքաշ ձին, աղավնու զանազան ցեղերը և այլն ստանալը չէր կարող հաջորդական սերունդների երկար շարանների ընթացքում միանման փոփոխությունների միայն պատահական կուտակման արդյունք լինել։ Եվ իրոք, գործնականում աղավնաբուծության մի սիրահար ուշադրություն է դարձնում սովորականից մի քիչ ավելի կարճ կտուցով աղավնու վրա, իսկ մյուսը, ընդհակառակը, սովորականից մի քիչ ավելի երկար կտուցով աղավնու վրա, և երկուսն էլ այն հայտնի կանոնի հիման վրա, «որ սիրահարները չեն գնահատում և չեն ցանկանում գնահատել միջին ձևերը և պահանջում են միայն ծայրահեղություններ», կշարունակեն ընտրել ու բազմացնել միայն ավելի ու ավելի կարճ կամ ավելի ու ավելի երկար կտուց ունեցող աղավնիներ։ Իրականության մեջ նման մի բան տեղի է ունեցել թուրմանի ենթացեղերի հետ։ Մենք կարող ենք նաև ընդունել, որ պատմության շատ վաղ ժամանակաշրջանում մի տոհմի պատկանող կամ որոշ տեղի մարդիկ կարիք են ունեցել արագավազ ձիերի, իսկ մյուսները կամ մի այլ տեղի մարդիկ ավելի ուժեղ և ծանրամարմին կենդանիների։ Սկզբնական տարբերությունը կարող էր շատ փոքր լինել, բայց ժամանակի ընթացքում, մի կողմից ամենից ավելի արագավազ, իսկ մյուս կողմից ամենաուժեղ կենդանիների ընտրության հետևանքով տարբերությունը նրանց միջև կարող էր մեծանալ և սկիզբ տալ երկու ենթացեղերի։ Վերջապես, դարերի ընթացքում այդ ենթացեղերը կերպափոխվել, դարձել են երկու լավ կայունացած և իրարից բոլորովին տարբերվող ցեղեր։ Այդ տարբերության մեծացման հետ զուգընթաց միջանկյալ հատկանիշներ ունեցող, ոչ այնքան արագավազ և ոչ այնքան ուժեղ, վատ ձիերը արդեն չեն թողնվել բազմացման համար և հետզհետե անհետացել են։ Այստեղ, մարդկային գործունեության կիրառության մեջ մենք տեսնում ենք այն բանի արտահայտումը, որ կարելի է անվանել հատկանիշների տարամիտման սկզբունք, որը ցեղերի հատկանիշներում առաջ է բերում սկզբում հազիվ նկատելի տարբերությունների մշտական մեծացում ինչպես միմյանց, այնպես էլ իրենց ընդհանուր նախահոր հատկանիշների նկատմամբ։
Բայց հավանորեն կհարցնեն, թե կարո՞ղ է արդյոք նման որևէ սկզբունք կիրառվել բնության նկատմամբ։ Ես կարծում եմ, որ կարող է և շատ ներգործոն ձևով (թեպետև շատ ժամանակ անցավ, մինչև որ ես իմացա, թե հատկապես ինչպես)։ Նրա կիրառելիությունը բխում է այն ամենապարզ հանգամանքից, որ որքան մի որևէ տեսակի սերունդները ավելի կտարբերվեն միմյանցից իրենց կառուցվածքով, ընդհանուր կերտվածքով ու սովորություններով, այնքան նրանք ավելի հեշտությամբ ի վիճակի կլինեն գրավելու բնության տնտեսության մեջ ավելի բազմաթիվ և ավելի բազմազան վայրեր, հետևաբար և այնքան ավելի հեշտությամբ նրանք կբազմանան։
Դրանում մենք կարող ենք հեշտությամբ համոզվել պարզ սովորություններ ունեցող կենդանիների օրինակի վրա։ Կանգ առնենք մի գիշատիչ չորքոտանու տեսակի վրա, որի թիվը վաղուց հասել է այն միջին սահմանին, որպիսին կարող է տեղավորել տվյալ երկիրն իր մեջ։ Եթե նրա բազմանալու բնական ձգտման գործողությանը ազատություն տրվի, ապա այդ բազմացումը կարող է իսկապես իրագործվել (ենթադրելով, որ երկրի ֆիզիկական պայմանները մնում են նույնը), եթե միայն փոփոխված սերունդները կկարողանան գրավել մյուս կենդանիների գրաված վայրերը, իսկ դրան կարելի է հասնել կամ նրանով, որ նրանք հնարավորություն ձեռք կբերեն սնվելու նոր տեսակի կենդանի կամ մեռած որսով, կամ նրանով, որ նրանք կսովորեն ապրել նոր պայմաններում՝ մագլցել ծառերի վրա, ապրել ջրում, կամ թե նրանով, վերջապես, որ նրանք կդառնան ավելի պակաս գիշակեր։ Ինչքան մեր գիշատչի այդ սերունդներն իրենց սովորություններով և կառուցվածքով ավելի բազմազան լինեն, այնքան ավելի մեծ թվով վայրեր կարող են գրավել։ Ինչ որ կիրառվում է մի կենդանու վերաբերմամբ, նույնը կիրառելի է նաև բոլորի վերաբերմամբ և բոլոր ժամանակներում, հասկանալի է՝ միայն մի պայմանով, որ նրանք փոփոխվում են, առանց որի բնական ընտրությունը, իհարկե, չի կարող դրսևորել իր ներգործությունը։ Նույնը ճիշտ է նաև բույսերի վերաբերմամբ։ Փորձով ապացուցված է, որ եթե մի հողամասի վրա ցանում ենք խոտի մի տեսակ, մի այլ նման հողամասի վրա՝ մի քանի տարբեր սեռերի պատկանող խոտեր, ապա երկրորդ դեպքում կստացվեն ավելի մեծ թվով բույսեր և ավելի մեծ քանակությամբ խոտ, քան առաջինում։ Նույնը ճիշտ է եղել, երբ ցորենի մեկ կամ մի քանի տարատեսակներ ցանել են հավասար մեծությամբ հողամասերում։ Հետևաբար, եթե խոտի մի որևէ տեսակ սկսի փոփոխվել, և շարունակ ընտրվեն այն տարատեսակները, որոնք միմյանցից զանազանվում են թեպետև փոքր չափով, բայց միևնույն ուղղությամբ, ինչպես մեր խոտերի տեսակները և սեռերն են զանազանվում, ապա հետևանքն այն կլինի, որ նույն հողամասի վրա կտեղավորվեն այդ տեսակին պատկանող մեծ թվով անհատներ, այստեղ ներառյալ նաև նրա փոփոխված սերունդները։ Իսկ մենք գիտենք, որ խոտի յուրաքանչյուր տեսակ կամ յուրաքանչյուր տարատեսակ ամեն տարի ցրում է գրեթե անթիվ սերմեր և, այսպես ասած, լարում է իր բոլոր ուժերը, որպեսզի ավելացնի իր թիվը։ Հետևաբար, բազմահազար սերունդների ընթացքում խոտի որևէ տեսակի ամենախիստ կերպով տարբերվող տարատեսակները կունենան հաջողության և բազմացման համար ամենամեծ շանսերը ու դուրս կմղեն ավելի քիչ տարբերություններ ունեցող տարատեսակներին, իսկ երբ տարատեսակները շատ խիստ տարբերվում են մեկը մյուսից, այդ դեպքում նրանց բարձրացնում են տեսակի աստիճանի։
Այս դրույթի ճշտությունը, որ կյանքի խոշորագույն գումարը իրականանում է կառուցվածքի առավելագույն բազմազանությամբ, արդարանում է բնական պայմաններում շատ դեպքերում։ Շատ փոքրիկ, հատկապես ներգաղթի համար բաց տարածություններում, որտեղ անհատների միջև մրցակցությունը պետք է չափազանց կատաղի լինի, մենք միշտ բնակիչների մեջ նկատում ենք մեծ բազմազանություն։ Այսպես, օրինակ, ես գտել եմ, որ չորս ոտնաչափ երկարությամբ և երեք ոտնաչափ լայնությամբ ճմուտի մի հողակտոր, որը երկար տարիներ գտնվել է միանման պայմաններում, բովանդակում էր քսան տեսակ բույսեր, որոնք պատկանում էին տասնևութ սեռերի և ութ ընտանիքների, որը ապացուցում է, թե որքան խիստ կերպով էին տարբերվում նրանք միմյանցից։ Նույնը ճիշտ է փոքրիկ միապաղաղ կղզիների վրա կամ քաղցրահամ լճակներում ապրող բույսերի և միջատների վերաբերմամբ։ Երկրագործները լավ գիտեն, որ իրենք կարող են ամենամեծ քանակությամբ սննդանյութեր հավաքել ամենատարբեր ընտանիքներին պատկանող բույսերի ցանքաշրջանառությւսն միջոցով. բնությունը դիմում է, եթե կարելի է այսպես ասել, միաժամանակյա պտղափոխության։ Մի որևէ հողակտոր շրջապատող բույսերի և կենդանիների մեծամասնությունը կարող էր ապրել նաև հողակտորի վրա (ենթադրելով, որ նրա բնությունը բացառիկ ոչինչ չի ներկայացնում) և նույնիսկ, կարելի է ասել, լարում է բոլոր ուժերը, որպեսզի տիրապետի նրան, և ահա մենք տեսնում ենք, որ ամենուրեք, որտեղ այդ բույսերն ու կենդանիները անմիջական շփման մեջ են մտնում միմյանց հետ, կաոուցվածքի բազմազանության և նրա հետ կապված կազմվածքի ու կենսաձևի տարբերության օգտակարությունը այն հետևանքն է ունենում, որ անմիջական գոտեմարտում միմյանց դեմ դուրս եկող օրգանիզմները, որպես ընդհանուր կանոն, պատկանում են նրանց, որոնց անվանում ենք տարբեր սեռեր և ընտանիքներ։
Նույն սկզբունքն արտահայտվում է նաև այն ժամանակ, երբ բույսերը մարդու օժանդակությամբ նատուրալիզացիայի են ենթարկվում իրենց համար օտար երկրներում։ Կարելի էր սպասել, որ այն բույսերը, որոնք հաջողությամբ նատուրալիզացիայի են ենթարկվում որևէ երկրում, ամենից ավելի մոտ կլինեն տեղականներին, որովհետև սովորաբար կարծում են, թե վերջիններս հատկապես ստեղծված ու հարմարված են իրենց բնակավայրին։ Նմանապես կարելի էր սպասել, որ նատուրալիզացված բույսերը կպատկանեն այն փոքրաթիվ խմբերին, որոնք իրենց նոր հայրենիքում առանձնապես հարմարված են որոշ բնակավայրերի։ Սակայն իրականության մեջ բոլորովին հակառակն է լինում, և Ալֆոնս դը Կանդոլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] հաջող կերպով է արտահայտվել իր ընդարձակ և հիանալի աշխատության մեջ, որ բուսականությունը նատուրալիզացիայի միջոցով տեղական սեռերի և տեսակների թվի համեմատությամբ ձեռք է բերում ավելի շատ սեռեր, քան տեսակներ։ Բերեմ մի օրինակ. «Հյուսիսային Ամերիկայի Միացյալ Նահանգների բուսականությունը» վերջին հրատարակության մեջ Ազա-Գրեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 68] թվում է 260 նատուրալիզացված տեսակներ, և նրանք պատկանում են 162 սեռերի։ Սրանից մենք տեսնում ենք, որ նատուրալիզացիայի ենթարկված այդ բույսերը չափազանց բազմազան են։ Բացի դրանից, նրանք զգալիորեն տարբերվում են տեղականներից, որովհետև նատուրալիզացիայի ենթարկված 162 սեռերից ոչ պակաս քան 100-ը չեն պատկանում տեղական բույսերին, հետևաբար, ստացվել է Միացյալ Նահանգներում արդեն գոյություն ունեցող սեռերի հարաբերական մեծ ավելացում։
Ուսումնասիրելով այն բույսերի և կենդանիների բնույթը, որոնք հաջողությամբ դիմացել են տեղականների դեմ մղված կռվին և կարողացել են նատուրալիզացիայի ենթարկվել, մենք կարող ենք մոտավոր գաղափար կազմել այն մասին, թե ինչ ուղղությամբ պիտի փոփոխվեին բնիկներից մի քանիսը, որպեսզի ձեռք բերեին առավելություններ իրենց հայրենակիցների հանդեպ, և համենայն դեպս մենք իրավունք ունենք եզրակացնելու, որ կառուցվածքի մեջ սեռային տարբերություններին համազոր տարբերություններ ձեռք բերելը պետք է նրանց համար ձեռնտու լիներ։
Այն առավելությունները, որ ստանում են միևնույն երկրի բնակիչները իրենց կառուցվածքի բազմազանության շնորհիվ, ըստ էության նույնն են, ինչ որ անհատական օրգանիզմը ձեռք է բերում իր տարբեր օրգանների միջև աշխատանքի բաժանման շնորհիվ, մի հարց, որ հիանալի կերպով պարզաբանել է Միլն Էդվարդսը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 86] Ոչ մի ֆիզիոլոգ չի կասկածում այն բանում, որ բացառապես միս կամ բացառապես բուսական նյութեր մարսելու հարմարված ստամոքսը ամենամեծ քանակությամբ սննդանյութեր է դուրս քաշում ընդունած կերակրից։ Այդպես էլ որևէ երկրի ընդհանուր տնտեսության մեջ. ինչքան ավելի մեծ է, ինչքան ավելի լիակատար է բույսերի և կենդանիների բազմազանությունը իրենց կենսաձևի տեսակետից, այնքան ավելի մեծ թվով անհատներ կկարողանան պահպանել իրենց գոյությունը։ Կենդանիների մի խումբ, որոնց կազմվածքը քիչ բազմազանություն ունի, դժվար թե դիմանա ավելի բազմազան կազմվածք ունեցող մի այլ խմբի մրցությանը։ Կարելի է կասկածել, օրինակ, կարող էին արդյոք ավստրալիական պարկավորները, որոնք ստորաբաժանվում են մեկը մյուսից քիչ չափով տարբերվող խմբերի և որոնք, ըստ Ուոտերհաուզի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 87] և այլ կենդանաբանների կարծիքի, համապատասխանում են մեր գիշատիչներին, որոճողներին և կրծողներին, կարո՞ղ էին արդյոք այդ պարկավորները հաջողությամբ մրցել մեր լավ արտահայտված կարգերի հետ։ Ավստրալիական կաթնասուններին մենք գտնում ենք բաժանման այդ պրոցեսի վաղ և անկատար աստիճանի վրա։
ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ԱԶԴԵՑՈՒԹՅԱՆ ՀԱՎԱՆԱԿԱՆ ՀԵՏԵՎԱՆՔՆԵՐԸ ՄԻ ԸՆԴՀԱՆՈՒՐ ՆԱԽԱՀՈՐ ՀԱՋՈՐԴՆԵՐԻ ՎՐԱ ՀԱՏԿԱՆԻՇՆԵՐԻ ՏԱՐԱՄԻՏՄԱՆ ԵՎ ՄԱՀԱՋՆՋՄԱՆ ՄԻՋՈՑՈՎ[43]
Վերևում համառոտակի կերպով շարադրված կշռադատումների հիման վրա մենք կարող ենք հետևցնել, որ մի որևէ տեսակի փոփոխված հաջորդները այնքան ավելի հաջողություն կունենան, ինչքան ավելի բազմազան կլինեն իրենց կառուցվածքով, այդպիսով հնարավորություն կունենան տիրապետելու այլ էակների գրաված տեղերը։ Այժմ տեսնենք, թե հատկանիշների տարամիտման օգտակարության այդ սկզբունքը ինչպես կներգործի բնական ընտրության և մահաջնջման կապակցությամբ։
Կցված աղյուսակը (էջ 205) մեզ կօգնի պարզելու այդ բավական դժվարին հարցը։ Դիցուք՝ A-ից մինչև L տառերը ցույց կտան մի երկրում ապրող ընդարձակ սեռի տեսակներին, ենթադրվում է, որ այդ տեսակները միմյանց հետ հավասար չափով նմանություն չունեն, ինչպես որ բնության մեջ հաճախ է հանդիպում և մեր աղյուսակում արտահայտված է տառերի միջև եղած անհավասար տարածություններով։ Ես ասացի մի ընդարձակ սեռ նրա համար, որովհետև մենք տեսանք երկրորդ գլխում, որ միջին հաշվով փոփոխականությունը ավելի մեծ թվով տեսակներ է ընդգրկում մեծ սեռերում, քան փոքրներում, իսկ մեծ սեռերի փոփոխվող տեսակները հանդես են բերում մեծ թվով տարատեսակներ։ Մենք տեսանք նաև, որ սովորական և լայնորեն տարաբնակված տեսակներն ավելի փոփոխական են, քան հազվագյուտ և սահմանափակ տարածվածություն ունեցող տեսակները։ Թող A-ն լինի իր հայրենիքում մի ընդարձակ սեռի սովորական, լայն կերպով տարածված և փոփոխվող մի տեսակը։ A-ից ճյուղավորվող և իրարից հեռացող տարբեր երկարության կետագծերը պատկերում են նրա փոփոխվող հաջորդներին։ Ենթադրենք, թե փոփոխությունները շատ փոքր են, բայց խիստ բազմազան, նրանք առաջանում են ոչ թե միաժամանակ, այլ հաճախ երկար ժամանակամիջոցներից հետո և պահպանվում են ոչ միահավասար տևողությամբ։ Միայն այն փոփոխությունները, որոնք այսպես թե այնպես օգտակար են, պահպանվում են և ենթարկվում բնական ընտրության։ Այստեղ իր ներգործությունը կդրսևորի հատկանիշների տարամիտման օգտակարության սկզբունքը. այդ սկզբունքի շնորհիվ այն փոփոխությունները, որոնք ամենից շատ են տարբերվում միմյանցից, ամենից շատ հեռանում միմյանցից (պատկերված են արտաքին գծերով), կպահպանվեն և կկուտակվեն բնական ընտրությամբ։ Երբ կետագիծը հասնում է հորիզոնական գծերից մեկին, որտեղ հատման կետը նշված է թվանշան ունեցող փոքրատառով, այն ժամանակ ենթադրվում է, որ փոփոխականությունը հասել է այնպիսի չափի, որը բավական է, որպեսզի այդ ձևը համարվի ցայտուն կերպով արտահայտված այն տարատեսակներից մեկը, որպիսիք տեղ են գտնում սիստեմատիկայի վերաբերյալ աշխատություններում։

Հորիզոնական գծերի միջև եղած տարածությունները կարող են համապատասխանել հազար կամ ավելի ևս մեծ թվով սերունդների։ Ենթադրվում է, թե հազար սերնդից հետո A տեսակն առաջ է բերել երկու պարզորոշ կերպով նկատելի տարատեսակներ, այն է՝ a և m։ Այդ երկու տարատեսակները առաջվա պես կենթարկվեն միևնույն պայմանների ներգործությանը, որոնք առաջացրել են նրանց ծնողների փոփոխությունները, իսկ փոփոխականության ձգտելը ինքնին ժառանգական է, հետևաբար, նրանք նույնպես կհակվեն հետագա փոփոխման և սովորաբար համարյա նույն ուղղությամբ, ինչպես որ նրանց ծնողներինն էր։ Բացի դրանից, քանի որ այդ տարատեսակները այնքան էլ խոր չեն փոփոխվել, ուստի նրանք կձգտեն ժառանգել այն առավելությունները, որոնց շնորհիվ նրանց սկզբնական ձևը (A) դարձել է ավելի բազմաթիվ, քան այդ նույն երկրի բնակիչների մեծամասնությունը։ Նրանք պետք է ունենան նաև այն ավելի ընդհանուր առավելությունները, որոնց շնորհիվ այն սեռը, որին պատկանում է նրանց առաջացնող տեսակը, դարձել է այդ երկրում խոշոր սեռ։ Իսկ այս բոլոր հանգամանքները նպաստում են նոր տարատեսակների առաջացմանը։
և m։ Այդ երկու տարատեսակները առաջվա պես կենթարկվեն միևնույն պայմանների ներգործությանը, որոնք առաջացրել են նրանց ծնողների փոփոխությունները, իսկ փոփոխականության ձգտելը ինքնին ժառանգական է, հետևաբար, նրանք նույնպես կհակվեն հետագա փոփոխման և սովորաբար համարյա նույն ուղղությամբ, ինչպես որ նրանց ծնողներինն էր։ Բացի դրանից, քանի որ այդ տարատեսակները այնքան էլ խոր չեն փոփոխվել, ուստի նրանք կձգտեն ժառանգել այն առավելությունները, որոնց շնորհիվ նրանց սկզբնական ձևը (A) դարձել է ավելի բազմաթիվ, քան այդ նույն երկրի բնակիչների մեծամասնությունը։ Նրանք պետք է ունենան նաև այն ավելի ընդհանուր առավելությունները, որոնց շնորհիվ այն սեռը, որին պատկանում է նրանց առաջացնող տեսակը, դարձել է այդ երկրում խոշոր սեռ։ Իսկ այս բոլոր հանգամանքները նպաստում են նոր տարատեսակների առաջացմանը։
Այնուհետև եթե այդ երկու տարատեսակները փոփոխական կլինեն, ապա նրանց փոփոխակներից նորից ամենից ավելի միմյանցից հեռացողները սովորաբար կպահպանվեն հաջորդ հազար սերունդների ընթացքում։ Այդ ժամանակաշրջանն անցնելուց հետո ենթադրվում է, որ a տարատեսակը, ինչպես երևում է աղյուսակից, առաջացրել է a տարատեսակը, որը տարամիտման սկզբունքի շնորհիվ A-ից ավելի շատ է տարբերվում, քան՝ a տարատեսակը։ Ենթադրվում է, որ m տարատեսակը այդ ժամանակամիջոցում առաջացրել է երկու տարատեսակ՝ m և s, որոնք տարբերվում են մեկը մյուսից և առավել ևս A նախահորից։ Այդ պրոցեսը նման փուլերով կարող է անորոշ ժամանակով երկար շարունակվել, տարատեսակներից մի քանիսը հազար սերնդից հետո կառաջացնեն միայն մեկական, շարունակ ավելի ու ավելի շեղվող տարատեսակ, մյուսները կառաջացնեն երկուական կամ երեքական և, վերջապես երրորդները ոչինչ չեն առաջացնի։ Այսպիսով, տարատեսակները կամ A ընդհանուր նախածնողական ձևի փոփոխված հաջորդները իրենց թիվը կավելացնեն և կդառնան հատկանիշներով բազմազան։ Աղյուսակի վրա այդ պրոցեսը հասցրած է մինչև տասհազարերորդ սերունդը, իսկ սեղմ և պարզացրած ձևով՝ մինչև տասնչորս հազարերորդ սերունդը։
տարատեսակը, որը տարամիտման սկզբունքի շնորհիվ A-ից ավելի շատ է տարբերվում, քան՝ a տարատեսակը։ Ենթադրվում է, որ m տարատեսակը այդ ժամանակամիջոցում առաջացրել է երկու տարատեսակ՝ m և s, որոնք տարբերվում են մեկը մյուսից և առավել ևս A նախահորից։ Այդ պրոցեսը նման փուլերով կարող է անորոշ ժամանակով երկար շարունակվել, տարատեսակներից մի քանիսը հազար սերնդից հետո կառաջացնեն միայն մեկական, շարունակ ավելի ու ավելի շեղվող տարատեսակ, մյուսները կառաջացնեն երկուական կամ երեքական և, վերջապես երրորդները ոչինչ չեն առաջացնի։ Այսպիսով, տարատեսակները կամ A ընդհանուր նախածնողական ձևի փոփոխված հաջորդները իրենց թիվը կավելացնեն և կդառնան հատկանիշներով բազմազան։ Աղյուսակի վրա այդ պրոցեսը հասցրած է մինչև տասհազարերորդ սերունդը, իսկ սեղմ և պարզացրած ձևով՝ մինչև տասնչորս հազարերորդ սերունդը։
Սակայն ես պետք է վերապահում անեմ, որ ես չեմ կարծում, թե այդ պրոցեսը երբևէ այդպիսի կանոնավորությամբ է ընթացել, ինչպես ցույց է տրված աղյուսակի վրա, թեև նրա մեջ ընդունված են որոշ անկանոնություններ. ես չեմ կարծում նաև, թե այդ պրոցեսը անընդհատ է եղել. շատ ավելի հավանական է, որ յուրաքանչյուր ձև երկար ժամանակաշրջանների ընթացքում մնում է անփոփոխ և հետո նորից ենթարկվում է փոփոխության։ Ես չեմ կարծում նույնպես, թե իրարից շատ հեռացող տարատեսակներն անպայման միշտ պահպանվում էին, երբեմն միջին մի ձև կարող է ողջ մնալ երկար ժամանակ, չառաջացնելով, կամ կարող է պատահել, և առաջացնելով մի քանի փոփոխված հաջորդներ, որովհետև բնական ընտրությունը միշտ ներգործում է այլ էակների կողմից բոլորովին կամ մասամբ չգրավված տեղերի բնության հետ համապատասխան կերպով, իսկ դա կախված է անսահմանորեն բարդ հարաբերություններից։ Բայց, որպես ընդհանուր կանոն, որքան բազմազան է դառնում որևէ տեսակի հաջորդների կառուցվածքր, այնքան ավելի մեծ կլինի նրանց տիրապետած տեղերի թիվը բնության մեջ և այնքան ավելի կծավալվեն նրանց փոփոխված հաջորդները։
Մեր աղյուսակի վրա տոհմաբանական գծերը որոշակի տարածությունների վրա ընդհատվում են թվանշան ունեցող փոքրատառերով, որոնցով նշված են բավականաչափ ցայտուն տարբերությունների հասած հաջորդական ձևերը, որոնք կարող են ցուցակներում գրանցվել որպես տարատեսակներ։ Սակայն այդ րնդհատումներր երևակայական են. նրանց կարելի էր տեղավորել ժամանակամիջոցները ցույց տվող տարածությունների վրա ցանկացած տեղում, որոնք պետք է բավականաչափ երկար լինեն զգալի լափով փոփոխականություն և տարամիտում կուտակելու համար։
Քանի որ մեծ սեռի պատկանող սովորական և լայնորեն տարածված տեսակի բոլոր փոփոխված հաջորդները հակում կունենան պահպանելու այն առավելությունները, որոնք ապահովել են նրանց նախածնողական ձևի կենսական հաջողությունը, ուստի նրանք թվով կշատանան և իրենց հատկանիշներով կտարամիտվեն, որը ցույց է տրված աղյուսակի վրա մի քանի իրարից հեռացող ճյուղերով, որ դուրս են գալիս A-ից։ Հետագա և ավելի կատարելագործված տոհմաբանական ճյուղերի փոփոխված հետնորդները, շատ հավանական է, կգրավեն ավելի վաղ առաջացած և ավելի քիչ կատարելագործված ճյուղերի տեղերը, հետևաբար և կոչնչացնեն նրանց։ Աղյուսակի վրա այդ արտահայտված է նրանով, որ ստորին ճյուղերից մի քանիսը չեն հասնում ավելի բարձր հորիզոնական գծերին։ Որոշ դեպքերում, անկասկած, փոփոխականության պրոցեսը կսահմանափակվի միայն մի տոհմաբանական գծով, և փոփոխված հաջորդների թիվը չի աճի, թեև փոփոխության և հատկանիշների տարամիտման աստիճանը կմեծանա։ Այդպիսի դեպք կարելի էր արտահայտել աղյուսակի վրա, եթե ոչնչացնենք A-ից դուրս եկող բոլոր գծերը, բացի այն գծից, որը ձգվում է a-ից մինչև a -ը։ Ուրեմն՝ անգլիական արշավաձին և անգլիական պոյնտերը, ըստ երևույթին, իրենց նախածնողական ձևերից հատկանիշներով ավելի հեռացել են, ոչ մեկ և ոչ մյուս դեպքում չառաջացնելով նոր ճյուղեր կամ ցեղեր։
-ը։ Ուրեմն՝ անգլիական արշավաձին և անգլիական պոյնտերը, ըստ երևույթին, իրենց նախածնողական ձևերից հատկանիշներով ավելի հեռացել են, ոչ մեկ և ոչ մյուս դեպքում չառաջացնելով նոր ճյուղեր կամ ցեղեր։
Ենթադրենք, թե A տեսակը տասը հազար սերունդ տալուց հետո առաջացրել է երեք ձև՝ a, f և m, որոնք հատկանիշների տարամիտման հետևանքով հետագա սերունդներում դրսևորում են խոր, թեև, կարող է պատահել, անհավասար տարբերություններ ինչպես իրարից, այնպես էլ նախածնողական ձևից։ Եթե մենք ենթադրենք, որ երկու հորիզոնական գծերի միջև ընկած արանքում ձեռք բերված տարբերության աստիճանը չափազանց փոքր է, ապա այդ երեք ձևերը կարող են հանդես բերել միայն երեք պարզորոշ արտահայտված տարատեսակներ. բայց բավական է ընդունել, որ տարբերության այդ աստիճանները ավելի բազմաթիվ կամ ավելի խիստ կլինեն, որպեսզի այդ երեք ձևերը դառնային կասկածելի կամ նույնիսկ ցայտուն կերպով արտահայտված տեսակներ։ Այդ ձևով աղյուսակն արտահայտում է այն աստիճանները, որոնցով տարատեսակների միջև եղած փոքրիկ տարբերությունները դառնում են տեսակային խոշոր տարբերություններ։ Շարունակվելով է՛լ ավելի մեծ թվով սերունդների ընթացքում (ինչպես աղյուսակի վրա ցույց է տրված սեղմ, պարզացրած ձևով), այդ պրոցեսը կտա ութ տեսակներ, որոնք նշանակված են a -ից մինչև m տառերով և բոլորն էլ առաջացել են A-ից։ Ես կարծում եմ, այս ճանապարհով բազմապատկվում է տեսակների թիվը և առաջանում են սեռերը։
-ից մինչև m տառերով և բոլորն էլ առաջացել են A-ից։ Ես կարծում եմ, այս ճանապարհով բազմապատկվում է տեսակների թիվը և առաջանում են սեռերը։
Տեսակներով հարուստ սեռի մեջ, ամենայն հավանականությամբ, կփոփոխվի ոչ թե մեկ տեսակ, այլ մի քանի տեսակներ։ Աղյուսակի վրա ընդունված է, որ մյուս տեսակը (I) ևս փոփոխվելով նույնպիսի հաջորդական աստիճաններով, տասը հազար սերունդներից հետո առաջ է բերել կամ երկու լավ արտահայտված տարատեսակներ (w և z), կամ երկու տեսակներ, նայած այն բանին, թե տարբերության ինչպիսի աստիճան կներկայացնի հորիզոնական գծերի միջև ընկած տարածությունը։ Այնուհետև կարելի է ենթադրել, որ տասնչորս հազար սերունդ անցնելուց հետո առաջանում են վեց նոր տեսակներ, որոնք նշանակված են n-ից մինչև z տառերով։ Յուրաքանչյուր սեռում այն տեսակները, որոնք արդեն ամենից շատ են տարբերվում միմյանցից, սովորաբար կհակվեն առաջացնելու ամենամեծ թվով փոփոխված հաջորդներ, որովհետև վերջիններս ավելի շատ շանսեր կունենան տիրանալու բնության տնտեսության մեջ նոր և ամենաբազմազան վայրերի։ Այդ կշռադատության հիման վրա էլ ես ընտրել եմ ծայրամասային A տեսակը և գրեթե ծայրամասային I տեսակը որպես ամենից ավելի փոփոխված ձևերի ներկայացուցիչներ, որոնք սկիզբ են տվել նոր տարատեսակների և տեսակների։ Մեր նախասկզբնական սեռի մնացած ինը տեսակները (որ նշանակված են մեծատառերով) կարող են չափազանց երկար, բայց անհավասար ժամանակաշրջաններում իրենց հաջորդներին փոխանցել գրեթե անփոփոխ հատկանիշներ, դա աղյուսակի վրա արտահայտված է անհավասար երկարությամբ վերընթաց կետագծերով։
Սակայն փոփոխության այդ պրոցեսի ընթացքում շատ էական դեր կխաղա նաև մեր սահմանած սկզբունքներից մյուսը՝ մահաջնջման սկզբունքը։ Քանի որ ամեն մի երկրում, որը ծայր աստիճան լցված է բնակիչներով, բնական ընտրությունը ներգործում է միայն շնորհիվ ինչ-որ հատկությունների, որոնք ընտրյալ ձևերին գոյության կռվում տալիս են առավելություններ մնացածների հանդեպ, ապա փոփոխության յուրաքանչյուր ստադիայում կատարելագործված հաջորդների մեջ կարտահայտվի իրենց նախնիներին և առաջին նախահորը փոխարինելու և ոչնչացնելու ձգտումը, որովհետև չպետք է մոռանալ, որ մրցությունն ամենից ավելի կատաղի կլինի այն ձևերի միջև, որոնք կառուցվածքով, կերտվածքով և կենսակերպով ամենից ավելի մոտ են միմյանց։ Դրա հետևանքով բոլոր միջանկյալ այն ձևերը, որոնք գտնվում են սկզբնական և ավելի հետագա ձևերի միջև, կամ այդ միևնույնն է թե, նույն տեսակի ավելի նվազ կատարելագործված և ավելի կատարելագործված ձևերի միջև, ինչպես նաև նախածնողական տեսակային ձևը, կդրսևորեն մահաջնջվելու ձգտում։ Հավանորեն այդ նույնը կդրսևորվի նաև կողմնակի շատ գծերի մեջ, որոնց կհաղթվեն հետագա և ավելի կատարելագործված գծերի կողմից։ Սակայն եթե որևէ տեսակի փոփոխված սերունդները կընկնեն բոլորովին մի նոր երկիր, կամ արագ կերպով կհարմարվեն բոլորովին նոր բնակավայրին, որտեղ հաջորդն ու նախահայրը չեն մրցակցի, այդ դեպքում երկուսն էլ կարող են պահպանվել։
Իսկ եթե մեր աղյուսակը պատկերում է փոփոխականությունն ավելի լայն չափերով, այդ դեպքում A տեսակը և նրա բոլոր վաղ առաջացած տարատեսակները մահաջնջված կլինեն և կփոխարինվեն ութ նոր տեսակներով (a-ից մինչև m), նույնպես էլ I տեսակը կփոխարինվի վեց նոր տեսակներով (n-ից մինչև z)։
Բայց մենք կարող ենք ավելի հեռու գնալ։ Մեր սեռի նախասկզբնական տեսակները, ինչպես մենք արդեն պայմանավորվել ենք, միմյանց հետ շատ տարբեր աստիճանի նմանություններ ունեն. այդ այդպես էլ իրոք լինում է բնության մեջ. A տեսակն ավելի մոտ է B, C և D տեսակներին, քան մյուսներին, իսկ I տեսակն ավելի մոտ է G, H, K, L-ին, քան մյուս տեսակներին։ Մենք ենթադրել ենք նաև, որ այդ երկու՝ A և I տեսակները պատկանում են շատ սովորական և լայնորեն տարածված տեսակների թվին, այնպես որ սկզբից նրանք պետք է ունենային նույն սեռի մյուս տեսակների հանդեպ որևէ էական առավելություններ։ Նրանց տասնչորս փոփոխված հաջորդները տասնևչորս հազար սերունդների ընթացքում հավանորեն ժառանգել են այդ առավելությունների մի մասը, նրանք զարգացման յուրաքանչյուր ստադիայում էլ փոփոխվել են ու կատարելագործվել նաև զանազան ուղղություններով, այնպես որ կարողացել են հարմարվել տվյալ երկրի բնության տնտեսության մեջ շատ վայրերի։ Հետևապես, շատ հավանական է, որ նրանք տեղ են գրավել և, այսպիսով, ոչնչացրել են ոչ միայն իրենց A և I նախահայրերին, այլ նաև նախասկզբնական տեսակներից մի քանիսին, որոնք ամենից ավելի մոտ են եղել այղ նախահայր ձևերին։ Այսպիսով, շատ քիչ թվով տեսակներ կպահպանվեն մինչև տասնչորս հազարերորդ սերունդը։ Մենք կարող ենք ենթադրել, որ միայն մի (F) կամ երկու (F և E) տեսակները, որ ամենից քիչ մոտիկ են մնացած ինը սկզբնական տեսակներին, հաջորդներ կունենան քննարկվող պրոցեսի այդ վերջին ստադիայում։
Սկզբնական տասնմեկ տեսակներից առաջացած նոր տեսակների թիվը մեր աղյուսակի վրա կլինի տասնևհինգ։ Շնորհիվ տարամիտման, որը միշտ ընտրության հետևանքն է, a և z տեսակների միջև սահմանային տարբերությունը զգալիորեն կգերազանցի սկզբնական տասնևմեկ տեսակներից ամենածայրի տեսակների միջև եղած տարբերությանը։ Բացի դրանից, նոր տեսակների միջև փոխադարձ կապը բոլորովին այլ կլինի։ A-ի ութ հաջորդներից երեքը, որ նշված են a, g և p տառերով, միմյանց հետ շատ մոտիկ ազգակցության մեջ կլինեն, որովհետև ներկայացնում են ոչ շատ վաղուց a-ից առաջացած ճյուղեր, b և f-ը, որ ավելի վաղ շրջանում են բաժանվել a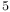 -ից, որոշ չափով կտարբերվեն առաջին երեք տեսակներից, և վերջապես o, e և m-ը մոտ ազգակցության մեջ կլինեն միմյանց հետ, բայց քանի որ նրանք մնացածներից հեռացել են փոփոխության պրոցեսի հենց սկզբում, ուստի խորապես կտարբերվեն մնացած հինգ տեսակներից և կկազմեն ենթասեռ կամ ինքնուրույն սեռ։
-ից, որոշ չափով կտարբերվեն առաջին երեք տեսակներից, և վերջապես o, e և m-ը մոտ ազգակցության մեջ կլինեն միմյանց հետ, բայց քանի որ նրանք մնացածներից հեռացել են փոփոխության պրոցեսի հենց սկզբում, ուստի խորապես կտարբերվեն մնացած հինգ տեսակներից և կկազմեն ենթասեռ կամ ինքնուրույն սեռ։
I-ի վեց հաջորդները կազմում են երկու ենթասեռ կամ երկու առանձին սեռ։ Բայց քանի որ I սկզբնական տեսակը լայն կերպով տարբերվում էր A-ից՝ գտնվելով նրանից սկզբնական սեռի հակադիր ծայրում, ուստի և I տեսակի վեց հաջորդները միայն ժառանգականության շնորհիվ խիստ կտարբերվեն A տեսակի ութ հաջորդներից. բացի դրանից, մենք ենթադրել ենք, որ այդ խմբերը շարունակել են հեռանալ դեպի հակառակ կողմեր։ Միջանկյալ տեսակները (և դա առանձնապես կարևոր նկատառում է), որոնք շաղկապում էին նախասկզբնական տեսակներին (A և I իրար հետ), բոլորը, բացառությամբ F-ի, մահաջնջվել են, չթողնելով հաջորդներ։ Այդ պատճառով էլ I-ից առաջացած վեց և A-ից առաջացած ութ նոր տեսակները պետք է դասել երկու շատ տարբեր սեռերի կամ նույնիսկ առանձին ենթաընտանիքների շարքը։
Այսպիսով, իմ կարծիքով, մի սեռի երկու կամ ավելի տեսակներից փոփոխությամբ ուղեկցվող ազգակցական հաջորդման միջոցով կարող են առաջանալ երկու կամ էլ ավելի մեծ թվով սեռեր։ Իսկ այդ երկու կամ ավելի սկզբնական տեսակները, կարելի է ենթադրել, ծագել են մի տեսակից, որը պատկանել է մի ավելի հնագույն սեռի։ Աղյուսակի վրա այդ արտահայտված է գլխատառերի տակի գծերով, որոնք կազմում են վարընթաց ուղղությամբ մի կետում միացող ճյուղերի փնջեր. այդ կետը ներկայացնում է այն տեսակը, որ եղել է նոր ենթասեռերի և սեռերի ենթադրելի նախահայրը։
Արժե մի րոպե ուշադրություն սևեռել F նոր տեսակի հատկության վրա, որի վերաբերմամբ մենք ենթադրել ենք, որ նա հատկանիշների զգալի տարամիտում չի ցուցաբերել, բայց պահպանել է F-ի ձևը առանց փոփոխության կամ թեթևակի փոփոխված։ Այդ դեպքում նրա ազգակցական կապը մնացած տասնչորս տեսակների հետ կկրի շատ հետաքրքրական և ինչ որ զարտուղի բնույթ։ Ծագում առնելով մի ձևից, որը միջանկյալ է A-ի և I-ի միջև, որոնք ենթադրվում են մահաջնջված ու մեզ անհայտ, նշված ձևը երևան կհանի այնպիսի հատկանիշներ, որոնք որոշ չափով միջանկյալ են այդ երկու տեսակներից առաջացած երկու խմբերի միջև։ Բայց քանի որ այդ երկու խմբերը շարունակել են իրենց հատկանիշներով հեռանալ ու շեղվել իրենց նախածնողական ձևերից, ուստի նոր տեսակը (F-ը) կհանդիսանա անմիջականորեն, միջանկյալ ձև ոչ թե գոյություն ունեցող տեսակների միջև, այլ ավելի շուտ երկու խմբերի տիպերի միջև. յուրաքանչյուր բնախույզ, անկասկած, կհիշի այդպիսի դեպքեր։
Մեր աղյուսակում յուրաքանչյուր հորիզոնական գիծ պայմանականորեն համապատասխանել է հազար սերնդի, բայց նրանցից ամեն մեկը կարող է համապատասխանել միլիոն և ավելի սերունդների, նա կարող է նաև պատկերել երկրի կեղևի հաջորդական շերտավորումների կտրվածքը, որն իր մեջ պարունակում է մահաջնջված օրգանիզմների մնացորդները։ Երկրաբանության նվիրված գլխում մենք կվերադառնանք այդ խնդրին և, ես կարծում եմ, կհամոզվենք, որ այդ աղյուսակը զգալի չափով լուսաբանում է մահաջնջված ձևերի հետ ազգակցությունը, որոնք, թեև սովորաբար կարող են դասվել այժմյան սեռերի, ընտանիքների և կարգերի մեջ, այնուամենայնիվ հաճախ ներկայացնում են ինչ-որ մի միջին բան այժմյան խմբերի միջև։ Եվ մենք հեշտությամբ կարող ենք հասկանալ այդ փաստը, որովհետև մահաջնջված տեսակներն ապրել են հեռավոր դարաշրջաններում, երբ մեր ծննդաբանության ճյուղերը դեռևս չէին կարողացել այնպես հեռանալ, ինչպես այժմ։
Ես հիմք չեմ տեսնում, որ օրգանական ձևերի փոփոխության այստեղ նկարագրված պրոցեսը սահմանափակվի միմիայն սեռերի առաջացմամբ։ Եթե մենք ենթադրենք, որ մեր աղյուսակի վրա միմյանցից հեռացող կետագծերի յուրաքանչյուր խումբը ներկայացնում է շատ զգալի փոփոխություն, այդ դեպքում a-ից մինչև p-ը, ինչպես նաև b-ից մինչև f-ը և o-ից մինչև m տառերով նշանակված ձևերը կհամապատասխանեն երեք միմյանցից խիստ տարբերվող սեռերի։ Մենք կստանանք նաև երկու տարբեր սեռեր, որոնք ծագում են առել I-ից և խիստ տարբերվում են A-ի հաջորդներից։ Այդ երկու սեռախմբերը կներկայացնեն արդեն երկու տարբեր ընտանիքներ կամ կարգեր, նայած այն բանին, թև ինչպիսին կլինի աղյուսակում փոփոխականության և տարամիտման մեր ընդունած աստիճանը։ Իսկ այդ երկու ընտանիքը կամ կարգը առաջացել են նախասկզբնական սեռի երկու տեսակներից, որոնք իրենց հերթին հանդիսանում են ավելի ևս հին և մեզ անհայտ մի ձևի հետնորդները։
Մենք տեսանք, որ յուրաքանչյուր երկրում խոշոր սեռերի պատկանող տեսակներն ամենից ավելի հաճախ են տալիս տարատեսակներ կամ սկզբնավորվող տեսակներ։ Այդ էլ պետք էր սպասել, քանի որ բնական ընտրությունը ներգործում է այն հանգամանքի շնորհիվ, որ գոյության կռվում որևէ ձև մյուսների հանդեպ գերազանցություն է ունենում, ուստի և նա կպաշտպանի նրանց, որոնք արդեն օժտված են այդ առավելություններով, իսկ որևէ խմբի լայնատարածությունը ապացուցում է, որ նրա տեսակները ժառանգել են ընդհանուր նախահորից նրանց բոլորի համար որևէ առավելություն։ Այսպիսով, նոր փոփոխված հաջորդների առաջացման մեջ մրցությունը տեղի կունենա միայն մեծ խմբերի միջև, որոնք բոլորը ձգտում են ավելացնել իրենց թիվը։ Մի մեծ խումբ դանդաղորեն կհաղթահարի մյուսին, կրճատելով նրա թիվը և դրանով կնվազեցնի նրա փոփոխության և կատարելագործության հետագա շանսերը։ Միևնույն մեծ խմբի սահմաններում ավելի ուշ առաջացած և ավելի կատարելագործված ենթախմբերը, ճյուղավորվելով և բնության տնտեսության մեջ գրավելով նոր տեղեր, կփոխարինեն ու կոչնչացնեն ավելի հին և ավելի քիչ կատարյալ ենթախմբերին։ Փոքրիկ և մասնատված խմբերն ու ենթախմբերը, վերջապես, բոլորովին կանհետանան։ Ապագայի մասին մտածելով, մենք կարող ենք կանխատեսել, որ օրգանական էակների այն խմբերը, որ այժմ լայնատարած են ու տիրապետող և միևնույն ժամանակ ամենից ավելի քիչ ցրված են, այսինքն ամենից ավելի քիչ են տուժել մահաջնջումից, դեռ երկար կմեծանան։ Բայց թե ո՞ր խմբերը վերջնականապես կհաղթեն, ոչ ոք չի կարող նախագուշակել, որովհետև մենք գիտենք, որ շատ խմբեր, որոնք առաջ շատ բազմաթիվ և զարգացած են եղել, այժմ արդեն մահաջնջվել են։ Ավելի ևս հեռավոր ապագայի մասին մտածելով, մենք կարող ենք նախագուշակել, որ մեծ խմբերի անընդհատ և համառ ծավալման շնորհիվ բազմաթիվ մանր խմբեր վերջնականապես կոչնչանան և իրենցից հետո չեն թողնի փոփոխված հաջորդներ, որի հետևանքով որևէ ժամանակաշրջանում ապրող տեսակներից միայն շատ քչերը հեռավոր ապագայում կպահպանեն իրենց սերունդները։ Ես պետք է այս հարցին վերադառնամ դասակարգության նվիրված գլխում, բայց և այնպես ես կավելացնեմ, քանի որ այդ տեսակետի համաձայն հնագույն տեսակների շատ չնչին քանակն է իրենից հետո հաջորդներ թողել մինչև այժմ և քանի որ մի տեսակի բոլոր հաջորդները առաջացնում են մի դաս, ապա և հասկանալի է, թե ինչու դասերի թիվն այնքան սահմանափակ է կենդանական կամ բուսական թագավորության յուրաքանչյուր մեծ բաժնում։ Թեև հնագույն տեսակներից շատ քչերն են իրենցից հետո թողել փոփոխված հաջորդներ, այնուամենայնիվ հեռավոր անցյալի երկրաբանական դարաշրջաններում երկիրը կարող էր համարյա նույնքան խիտ բնակված լինել բազմաթիվ սեռերի, ընտանիքների, կարգերի և դասերի տեսակներով, ինչքան որ այժմ։
ԱՄԵՆԱԲԱՐՁՐ ՍԱՀՄԱՆԻ ՄԱՍԻՆ, ՈՐԻՆ ՁԳՏՈՒՄ է ՀԱՍՆԵԼ ԿԱԶՄՎԱԾՔԸ
Բնական ընտրությունը ներգործում է բացառապես միայն այն փոփոխությունների պահպանման և կուտակման միջոցով, որոնք նպաստավոր են օրգանիզմի համար օրգանական և անօրգանական այն պայմաններում, որոնց նա ենթարկվում է իր կյանքի բոլոր շրջաններում։ Վերջնական հետևանքը արտահայտվում է նրանով, որ յուրաքանչյուր էակ ձգտում է ավելի ու ավելի կատարելագործվել իրեն շրջապատող պայմանների նկատմամբ։ Այդ կատարելագործության անխուսափելի հետևանքը լինում է ամենամեծ թվով կենդանի էակների կազմվածքի աստիճանական զարգացումը ամբողջ աշխարհում։ Բայց այստեղ մենք մտնում ենք մի շատ խճճված հարցի բնագավառ, որովհետև մինչև այժմ բնախույզները չեն առաջարկել բոլորին բավարարող մի սահմանում, թե ինչ պետք է հասկանալ այդ ավելի բարձր կազմվածք հասկացողության տակ։ Ողնաշարավորների նկատմամբ մտավոր ընդունակությունների զարգացումը և մարդու կազմվածքին մոտ լինելու աստիճանը գլխավոր դեր են խաղում։ Կարելի էր կարծել, որ այն փոփոխությունների չափերը, որոնց ենթարկվում են զանազան մարմնամասերը և օրգանները սաղմնային վիճակից միանգամայն հասուն վիճակի անցնելու ժամանակ, կարող էին չափանիշ ծառայել համեմատելու համար. բայց հայտնի են դեպքեր, ինչպես մի քանի մակաբույծ խեցեմորթների մոտ, երբ կազմվածքի մի մասը զարգացման հետ դառնում է ավելի պակաս կատարյալ, այնպես որ միանգամայն զարգացած կենդանին չի կարող անվանվել ավելի բարձր կազմակերպված, քան նրա թրթուրը։ Ֆոն-Բերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 25] առաջարկած չափանիշն ըստ երևույթին ամենից ավելի կիրառելի է և ամենալավն է թվում, այն է՝ օրգանիզմի մասերի դիֆերենցման աստիճանը (ես կավելացնեի՝ հասուն վիճակում) և նրանց մասնագիտացումը զանազան գործունեության վերաբերմամբ, կամ, ինչպես կարտահայտվեր Միլն Էդվարդսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 86] աշխատանքի ֆիզիոլոգիական բաժանման լրիվության աստիճանը։ Բայց մենք կհամոզվենք, թե ինչքան մութն է այդ խնդիրը, եթե դառնանք ձկներին, որոնց մոտ բնախույզներից ոմանք բարձրակարգ են համարում նրանց, որոնք շնաձկների նման ավելի մոտենում են երկկենցաղներին, մինչդեռ այլ բնախույզներ բարձրակարգ են համարում սովորական ոսկրոտ ձկներին, որովհետև նրանց մեջ ամենից ավելի պարզորոշ կերպով է արտահայտված ձկան տիպը և նրանք ամենից ավելի են տարբերվում ողնաշարավոր կենդանիների մյուս դասերից։ Էլ ավելի ակներև է դառնում այդ հարցի խճճվածությունը, եթե մենք դառնանք բույսերին, որոնց վերաբերմամբ մտավոր ընդունակությունների չափանիշը կիրառելի չէ, այստեղ մենք կհանդիպենք բուսաբանների, որոնք բարձրակարգ են համարում այն բույսերը, որոնց բոլոր օրգանները, ինչպես, օրինակ, բաժակաթերթերը, պսակաթերթերը, առէջքները և վարսանդները լիովին զարգացած են ամեն մի ծաղկի մեջ, մինչդեռ այլ բուսաբաններ, և ամենայն հավանականությամբ ավելի մեծ հիմք ունենալով, բարձրակարգ են համարում այն բույսերը, որոնց զանազան օրգանները ամենից ավելի են փոփոխված, իսկ նրանց թիվը կրճատված է։
Եթե բարձր կազմվածքի չափանիշ մենք ընդունում ենք չափահաս օրգանիզմի զանազան օրգանների դիֆերենցման և մասնագիտացման աստիճանը (այստեղ ներառելով նաև ուղեղի զարգացման աստիճանը, որը որոշում է մտավոր ընդունակությունները), ապա բնական ընտրության ներգործությունն այդ ուղղությամբ ակնհայտ կլինի. բոլոր ֆիզիոլոգներն ընդունում են, որ օրգանների մասնագիտացումը օգտակար է դրանցով օժտված էակին, որովհետև այդ դեպքում նրանք ավելի լավ են կատարում իրենց դերը, այստեղից էլ պարզ է, որ մասնագիտացման հանգող փոփոխությունների կուտակումը մտնում է բնական ընտրության ներգործության շրջանի մեջ։ Մյուս կողմից՝ նկատի առնելով երկրաչափական պրոգրեսիայով բազմացումը և բոլոր օրգանական էակների ձգտումը գրավելու բնության տնտեսության մեջ ամեն մի ազատ կամ անբավարար կերպով գրավված տեղ, մենք հեշտությամբ կըմբռնենք, որ բնական ընտրությունը կարող է օրգանիզմները աստիճանաբար հարմարեցնել նաև այնպիսի դրույթների, երբ որոշ օրգաններ ավելորդ կամ անօգուտ կլինեն, և այդպիսի դեպքում նրա ներգործությունը կդրսևորվի հետադարձ շարժումով դեպի կազմվածքի ստորին աստիճանները։ Արդյո՞ք կազմվածքն ամբողջությամբ վերցրած շարժվել է առաջ՝ սկսած ամենահեռավոր երկրաբանական դարաշրջաններից մինչև օրս, ավելի հարմար կլինի քննարկել օրգանիզմների երկրաբանական հաջորդականությանը նվիրված գլխում։
Բայց կարող են առարկել, թե՝ եթե բոլոր օրգանական էակները դրսևորել են կազմվածքի ավելի բարձր աստիճանների բարձրանալու ձգտում, ապա ինչպե՞ս բացատրել, որ ամբողջ աշխարհում գոյություն ունի ստորակարգ ձևերի այդպիսի բազմություն, և ինչպե՞ս է, որ միևնույն դասի սահմաններում կարելի է հանդիպել ձևերի, որոնք անհամեմատ ավելի բարձր զարգացման են հասել, քան մյուսները։ Ինչո՞ւ ավելի բարձր կազմվածքով ձևերը ամենուրեք դուրս չեն մղել և չեն ոչնչացրել ստորակարգ ձևերին։ Լամարկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 2] համոզված լինելով, որ բոլոր օրգանիզմներին հատուկ է կատարելագործման անշեղ ձգտումը, այնքան խիստ ապշել է այդ դժվարությունից, որ եկել է այն ենթադրության, թե ինքնածնության միջոցով շարունակ ծնվում են նոր և ամենապարզ ձևեր։ Ինչ էլ որ հայտնագործելու լինի ապագայի գիտությունը, սակայն մինչև այժմ նա չի հաստատել այդ ենթադրությունը։ Մեր տեսության տեսակետից ստորակարգ օրգանիզմների անընդհատ գոյությունը ոչ մի դժվարություն չի ներկայացնում, որովհետև բնական ընտրությունը, կամ ամենահարմարվածի գերապրումը, չի ենթադրում անպայման առաջադիմական զարգացում՝ նա միայն առավելություն է տալիս այն փոփոխություններին, որոնք բարենպաստ են նրանցով օժտված էակին նրա բարդ կենսապայմաններում։ Բայց հարց է առաջանում, ի՞նչ օգուտ, որքան մենք ի վիճակի ենք այդ մասին դատել, կկարողանային քաղել ավելի բարձր կազմվածքից ինֆուզորիան, ճիճուն կամ նույնիսկ անձրևորդը։ Իսկ եթե դրանից ոչ մի օգուտ չկա, այդ դեպքում բնական ընտրությունը կամ բոլորովին չի կատարելագործի այդ ձևերը, կամ կկատարելագործի նրանց շատ թույլ չափով, այնպես որ նրանք կպահպանվեն անվերջ ժամանակներ իրենց այժմյան կազմվածքի ցածր աստիճանի վրա։ Երկրաբանությունն էլ վկայում է, որ ամենապարզ ձևերից մի քանիսը, ինֆուզորիան և արմատոտանիները, հսկայական ժամանակաշրջանների ընթացքում պահպանվել են այնպես, ինչպես մենք նրանց գիտենք այժմ։ Բայց ենթադրել, թե գոյություն ունեցող ստորակարգ ձևերի մեծամասնությունը ամենևին առաջ չի շարժվել սկսած օրգանական կյանքի հենց արշալույսից, կլիներ չափազանց անշրջահայաց, որովհետև ամեն մի բնախույզ, որ հետազոտել է այդ այսպես կոչված ստորակարգ օրգանիզմներից որևէ մեկին, պետք է զարմացած լինի նրանց իսկապես զարմանալի ու հիանալի կազմվածքով։
Գրեթե նույն դիտողությունները կիրառելի են նաև միևնույն մեծ խմբի սահմաններում եղած կազմվածքի զանազան աստիճանների համեմատության վերաբերմամբ, օրինակ, երբ ողնաշարավորների մեջ մենք հանդիպում ենք միաժամանակ կաթնասուններին և ձկներին, երբ կաթնասունների մեջ հանդիպում ենք մարդուն և բադակտուցին, ձկների մեջ՝ շնաձկանը և նշտարաձկանը (Amphioxus). վերջինս իր կազմվածքի պարզությամբ մոտենում է արդեն անողնաշարավորներին։ Բայց հազիվ թե կան փոխադարձ մրցման դեպքեր կաթնասունների և ձկների միջև, կաթնասունների ամբողջ դասի կամ նրա մի քանի ներկայացուցիչների կողմից կազմվածքի ամենաբարձր սահմանին հասնելը չէր ստիպի նրանց գրավելու ձկների տեղը։ Ֆիզիոլոգները ենթադրում են, որ ուղեղի բարձրագույն գործունեության համար անհրաժեշտ է, որպեսզի նա ողողվի տաք արյունով, և դրա համար, իր հերթին, անհրաժեշտ է օդային շնչառություն, ուրեմն ջրում ապրող տաքարյուն կաթնասունները ստիպված են շնչելու համար շարունակ ջրից դուրս ցցվել, որը վնաս է իրենց։ Ձկների վերաբերմամբ կարելի է ասել, որ շնաձուկը չէր ձգտի փոխարինել նշտարաձկանը, որովհետև վերջինս, ինչպես ինձ հայտնում է Ֆրից Մյուլլերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] Բրազիլիայի անպտուղ ավազուտ ափերում ունի միայն մի ուղեկից և մրցակից՝ մի ինչ որ անոմալ օղակավոր որդ։ Կաթնասունների երեք ամենապարզ կարգերը՝ պարկավորները, թերատամնավորները և կրծողները, ապրում են Հարավային Ամերիկայում բազմաթիվ կապիկների հետ միասին և, ամենայն հավանականությամբ, քիչ են շփվում իրար հետ։ Թեև էակների կազմվածքը ամբողջությամբ վերցրած ամբողջ աշխարհում առաջ է շարժվել և շարունակում է առաջ շարժվել, այնուամենայնիվ օրգանական սանդուղքը միշտ կներկայացնի կատարելագործության տարբեր աստիճաններ, որովհետև ամբողջ դասերի կամ նրանց առանձին ներկայացուցիչների բարձր զարգացումը իր հետ չի բերում այն խմբերի անխուսափելի կերպով մահաջնջում, որոնց հետ նրանք անմիջականորեն չեն մրցում։ Որոշ դեպքերում, ինչպես կտեսնենք հետագայում, ցածր կազմվածքով ձևերը, ըստ երևույթին, պահպանվել են մինչև այժմ այն հանգամանքի շնորհիվ, որ նրանք ապրել են բոլորովին բացառիկ և մեկուսացած բնակավայրերում, որտեղ ենթարկվել են ավելի քիչ անողոք մրցության և որտեղ նրանց փոքրաթիվ լինելը թուլացրել է բարենպաստ փոփոխությունների առաջացման հավանականությունները։
Ի վերջո ես կարծում եմ, որ ցածր կազմվածք ունեցող բազմաթիվ ձևերի գոյությունը ներկայումս բացատրվում է զանազան պատճառներով։ Որոշ դեպքերում կարող էին բոլորովին երևան չգալ բարենպաստ փոփոխություններ կամ անհատական շեղումներ, որոնց վրա կարողանար ներգործել բնական ընտրությունը և կուտակել նրանց։ Ամենայն հավանականությամբ կան նաև այնպիսի դեպքեր, երբ ժամանակ չի եղել հասնելու կատարելության հնարավոր բարձրագույն սահմանին։ Մի քանի հազվագյուտ դեպքերում երևան է եկել այն, որը կարելի է անվանել կազմվածքի հետադիմություն։ Բայց գլխավոր պատճառը այն փաստի մեջ է, որ շատ պարզ կենսապայմաններում բարձր կազմվածքը ոչ մի ծառայություն չէր մատուցի, թերևս նույնիսկ վատ ծառայություն մատուցեր, որովհետև իր քնքշության պատճառով ավելի ենթակա կլիներ վնասվելու և փչանալու։
Եթե մեր հայացքները դարձնենք դեպի կյանքի արշալույսը, երբ, կարելի է կարծել, բոլոր օրգանիզմները ունեցել են ամենապարզ կառուցվածքը, ապա հարց է առաջանում, ինչպե՞ս կարող էին արվել առաջին քայլերը մարմնամասերի կատարելագործման և դիֆերենցման ուղիով։ Հերբերտ Սպենսերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 18] հավանորեն կպատասխաներ, որ հենց որ միաբջիջ օրգանիզմը աճման և բաժանման հետևանքով դառել է բազմաբջիջ կամ ամրացել է այն մակերևույթին, որի վրա նա գտնվում է, ապա իսկույն ևեթ իր ներգործությունն է ցուցաբերել Սպենսերի ձևակերպած օրենքը, որ «ամեն կարգի հոմոլոգ միավորները այնքան ավելի շատ են դիֆերենցվում, որքան ավելի բազմազան են դառնում նրանց առնչությունները իրենց վրա ներգործող ուժերի հետ»։ Բայց քանի որ մենք փաստեր չունենք, որոնցով կարողանայինք ղեկավարվել, ուստի այդ առթիվ մտահայեցողությունը համարյա ապարդյուն կլիներ։ Սակայն սխալ կլիներ ենթադրել, թե ո՛չ գոյության կռիվը, ո՛չ էլ նրանից բխող բնական ընտրությունը չէին կարող դրսևորվել մինչև շատ ձևերի երևան գալը. մեկուսացած վայրում ապրող միայնակ տեսակի մեջ եղած փոփոխությունները կարող էին օգտակար լինել և այդպիսով կարող էր փոփոխվել անհատների ամբողջ բազմությունը կամ կարող էին առաջանալ երկու տարբեր ձևեր։
Բայց, ինչպես ես արդեն նկատել եմ իմ ներածության վերջում, ոչ ոք չպետք է զարմանա, որ տեսակների ծագման հարցում շատ բան դեռ մնում է չպարզաբանված,— բավական է միայն, որ ի նկատի առնենք մեր տգիտության խորությունը այս աշխարհի այժմյան, առավել ևս անցյալում ապրողների փոխհարաբերությունների վերաբերմամբ։
ՀԱՏԿԱՆԻՇՆԵՐԻ ՄԵՐՁԵՑՈՒՄԸ[44]
Միստր Հ. Կ. Ուոտսոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 62] կարծում է, որ ես չափազանցրել եմ հատկանիշների տարամիտման սկզբունքի նշանակությունը (որն այնուամենայնիվ, ըստ երևույթին, նա ընդունում է), և մատնանշում է այն, որ հատկանիշների մերձեցումը կամ մոտեցումը նույնպես որոշ դեր է խաղացել։ Եթե տարբեր, թեպետ և մերձավոր, սեռերի պատկանող երկու տեսակներից ամեն մեկն իր հերթին առաջացրել է բազմաթիվ նոր, միմյանցից հեռացող ձևեր, ապա կարելի է ընդունել, որ այդ ձևերից մի քանիսը կարող էին այնքան մոտենալ մեկը մյուսին, որ հարկավոր կլիներ նրանց մտցնել մի ընդհանուր սեռի մեջ. այնպես որ երկու տարբեր սեռերի հաջորդները կմիաձուլվեին ու կկազմեին մի սեռ։ Սակայն չափազանց անմտածված կլիներ, եթե մենք ուզենայինք միմյանց շատ հեռու կանգնած ձևերի փոփոխված հաջորդների մեջ կառուցվածքի ընդհանուր և մոտ նմանությունը վերագրել մերձեցմանը։ Բյուրեղի ձևը որոշվում է բացառապես մոլեկուլային ուժերի խաղով, և դրա համար զարմանալի չէ, եթե տարբեր նյութեր երբեմն ընդունում են նման ձևեր։ Օրգանական էակների վերաբերմամբ մենք չպետք է աչքաթող անենք, որ յուրաքանչյուրի ձևը կախված է ամենաբարդ հարաբերակցությունների անսահման ամբողջությունից, այն փոփոխությունների առաջացումից, որոնց պատճառներն այնքան բարդ են, որ նրանց հետամուտ լինելը նույնիսկ անհնարին է, այն փոփոխությունների բնույթից, որոնք պահպանվել են կամ ընտրվել են, իսկ դրանք բնորոշվում են շրջապատող ֆիզիկական պայմաններով և առավել ևս այն օրգանիզմներով, որոնց հետ նրանք ստիպված էին մրցման մեջ մտնելու և, վերջապես, նախնիքների անսահման շարքում ձեռք բերված ժառանգականությունից (հենց իր բնույթով անհաստատ էլեմենտից), որոնց ձևերն իրենց հերթին որոշվել են նույնպիսի բարդ պայմաններով։ Բոլորովին անհավատալի է այն ենթադրությունը, որ նախապես խիստ կերպով միմյանցից տարբերվող երկու օրգանիզմների հաջորդները կարող են իրար այն աստիճան մոտենալ, որի հետևանքով նրանց կազմվածքները գրեթե լիակատար նմանության հասնեին։ Եթե որևէ նման բան պատահեր, ապա մենք այդ միևնույն ձևին կհանդիպեինք իրարից շատ հեռու գտնվող երկրաբանական ֆորմացիաներում՝ անկախ գենետիկական կապից։ Բայց երկրաբանությունը այդպիսի ենթադրության դեմ է վկայում։
Միստր Ուոտսոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 62] նաև առարկել է, որ բնական ընտրության տևական ներգործությունը հատկանիշների տարամիտման սկզբունքի հետ միասին պետք է առաջ բերեին տեսակային ձևերի թվի անսահմանափակ ավելացում։ Քանի դեռ խոսքը վերաբերում է միմիայն անօրգանական պայմաններից ունեցած կախմանը, ապա հավանական է թվում, որ տեսակների մի որոշ բավարար քանակություն շուտով հարմարված կլինի բոլոր իրարից խիստ տարբերվող ջերմության, խոնավության և այլ պայմաններին. բայց ես միանգամայն ընդունում եմ, որ օրգանիզմների միջև փոխհարաբերություններն ավելի կարևոր դեր են կատարում, իսկ քանի որ տեսակների թիվը յուրաքանչյուր երկրում ժամանակի ընթացքում ավելանում է, ապա և կյանքի օրգանական պայմանները պետք է ավելի և ավելի բարդանան։ Ուրեմն, առաջին հայացքից ակներև է թվում, թե կառուցվածքի մեջ օգտակար զանազանակերպության ձգտելու համար սահման չկա, հետևաբար և սահման չունի այն տեսակների թիվը, որոնք կարող են առաջանալ։ Մենք չգիտենք նույնիսկ ձևերով ամենից ավելի հարուստ մարզերի մասին, թե արդյոք նրանք լիովին ծայր աստիճան հագեցած են տեսակներով, տեսակների զարմանալի քանակություն ունեցող Բարեհուսո հրվանդանի վրա և Ավստրալիայում եվրոպական շատ բույսեր կարողացել են նատուրալիզացիայի ենթարկվել։ Բայց երկրաբանությունը մեզ ուսուցանում է, որ երրորդային դարաշրջանի սկզբից խեցիների տեսակների թիվը, իսկ նրա կեսից սկսած նաև կաթնասունների թիվը շատ քիչ է ավելացել կամ նույնիսկ բոլորովին չի ավելացել։ Իսկ ի՞նչն է սահման դնում տեսակների թվի անսահման ավելացմանը։ Կյանքերի այն քանակությունը (այդ ասելով ես չեմ հասկանում տեսակային ձևերի թիվը), որ կարող է պահպանվել մի որոշ տարածության վրա, պետք է սահման ունենա, որովհետև նա մեծ չափով կախված է ֆիզիկական պայմաններից, ուրեմն, եթե այդ տարածությունը բնակված է շատ բազմաքանակ տեսակներով, այդ դեպքում նրանցից յուրաքանչյուրը կամ գրեթե յուրաքանչյուրը կարող է ունենալ միայն աննշան թվով անհատներ, իսկ այդպիսի տեսակները կենթարկվեն ոչնչացման կլիմայական պայմանների կամ թշնամիների թվի պատահական տատանումների հետևանքով։ Այդպիսի դեպքերում ոչնչացման պրոցեսը պետք է շատ արագ ընթանա, մինչդեռ նոր տեսակների առաջացումը միշտ պետք է չափազանց դանդաղ կատարվի։ Պատկերացրեք մի ծայրահեղ դեպք, որ Անգլիայում լինեին նույնքան տեսակներ, որքան որ անհատներ կան, և առաջին դաժան ձմեռը կամ չոր ամառը կոչնչացներ հազարավոր ու հազարավոր տեսակներ։ Սակավանդամ տեսակները (իսկ նրանց թվի անսահման աճման դեպքում բոլոր տեսակները դառնում են սակավանդամ) բազմիցս պարզաբանված սկզբունքի հիման վրա մի որոշ ժամանակաշրջանում առաջացնում են քիչ օգտակար փոփոխություններ։ Ուրեմն, նոր տեսակների սկզբնավորման բուն պրոցեսը կարգելակվի։ Եթե որևէ տեսակ դառնում է շատ սակավանդամ, այդ դեպքում ազգակցության մոտ աստիճաններում կատարվող խաչաձևումը կօժանդակի նրա ոչնչացմանը, մի քանի հեղինակներ հայտնել են այն կարծիքը, թե այդ պատճառով է բացատրվում զուբրի այլասերվելը Լիտվիայում, եղջերվինը՝ Շոտլանդիայում, արջինը՝ Նորվեգիայում և այլն։ Վերջապես, և դա, ես կարծում եմ, գլխավոր հանգամանքն է, որ տիրապետող տեսակը, որ իր բնակության մարզում արդեն հաղթել է շատ մրցակիցների, կձգտի ավելի ևս լայն կերպով տարածվելու և դուրս մղելու նաև այլ ձևեր։ Ալֆոնս դը Կանդոլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] ցույց է տվել, որ լայն տարածված տեսակները սովորաբար ցուցաբերում են շատ լայն տարածվելու ձգտում. հետևաբար, նրանք կձգտեն փոխարինել և ոչնչացնել մի քանի տեսակներ մի քանի մարզերում և այսպիսով կարգելակեն տեսակային ձևերի անսահման ավելացումը երկրի վրա։ Հուկերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] վերջերս ցույց է տվել, որ Ավստրալիայի հարավ-արևելյան անկյունում, որտեղ, ըստ երևույթին, երևացել են շատ եկվորներ աշխարհիս զանազան ծայրերից, ավստրալիական էնդեմիկ տեսակները զգալիորեն նվազել են թվով։ Ես հանձն չեմ առնի ասելու, թե այդ զանազան կշռադատումներին ինչ ուժ պետք է վերադրել, բայց այդ հանգամանքները միասին վերցրած պետք է սահմանափակին յուրաքանչյուր երկրում տեսակային ձևերի թիվն անսահման ավելացնելու ձգտումը։
ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ ԱՅՍ ԳԼԽԻՑ
Եթե փոփոխվող կենսապայմաններում օրգանական էակները կարող են հասնել անհատական շեղումների իրենց կազմվածքի գրեթե ամեն մի մասում, իսկ այդ ժխտել անհնարին է. եթե երկրաչափական պրոգրեսիայով բազմանալու հետևանքով սկսվում է կյանքի համար մղվող մի դաժան կռիվ բոլոր հասակներում, տարվա բոլոր եղանակներին, իսկ այդ, իհարկե, անհնարին է վիճարկել. նմանապես եթե հիշենք օրգանիզմների անսահմանորեն բարդ հարաբերությունները ինչպես միմյանց հետ, այնպես էլ կենսապայմանների հետ և այղ հարաբերություններից ծագում առնող կառուցվածքի ներքին կերտվածքի և սովորությունների օգտակար առանձնահատկությունների անսահման բարդությունը,— եթե այդ բոլորը ուշադրության առնենք, ապա չափազանց անհավատալի կլինի, որ երբեք հանդես չեն եկել շեղումներ, որոնք օգտակար են դրանցով օժտված օրգանիզմներին, ճիշտ այնպես, ինչպես առաջացել են մարդու համար օգտակար շեղումները։ Բայց եթե երբևիցե երևան են գալիս որևէ օրգանիզմի համար օգտակար շեղումներ, այդ դեպքում դրանցով օժտված օրգանիզմները, իհարկե, ավելի շատ շանսեր կունենան պահպանվելու գոյության կռվում, իսկ ժառանգականության հզոր սկզբունքի շնորհիվ նրանք կձգտեն այդ շեղումները փոխանցել իրենց հաջորդներին։ Ամենահարմարվածների պահպանվելու կամ գերապրելու այդ սկզբունքը ես անվանել եմ բնական ընտրություն։ Դա տանում է յուրաքանչյուր էակի դեպի կատարելագործություն նրա կյանքի օրգանական և անօրգանական պայմանների վերաբերմամբ և, հետևաբար, մեծ մասամբ նաև դեպի այն, որը կարելի է համարել կազմվածքի ավելի բարձր աստիճանի վրա բարձրանալ։ Բայց և այնպես պարզ կազմվածք ունեցող ստորին ձևերը երկար ժամանակ կպահպանվեն, եթե միայն նրանք լավ հարմարված են իրենց պարզ կենսապայմաններին։
Բնական ընտրությունը հատկությունների ժառանգման սկզբունքի հիման վրա համապատասխան հասակում կարող է փոփոխել ձուն, սերմը կամ ոչ բոլորովին զարգացած օրգանիզմը ճիշտ այնպես, ինչպես և հասուն օրգանիզմը։ Շատ կենդանիների վերաբերմամբ սեռական ընտրությունը հավանորեն օժանդակել է սովորական ընտրությանը, ապահովելով ամենաուժեղ և ամենից լավ հարմարված արուներից բազմաթիվ հաջորդների առաջացումը։ Սեռական ընտրությունը զարգացնում է նաև հատկանիշներ, որոնք օգտակար են բացառապես միայն արուներին՝ մյուս արուների դեմ կռվի կամ մրցության ընթացքում, և այդ հատկանիշները, նայած թե ինչ ուղղությամբ է արտահայտվում ժառանգականությունը, պետք է փոխանցվեն երկու սեռերին կամ միայն մեկին։
Այն մասին, թե արդյոք իրոք բնական ընտրությունը նման ձևով ազդել է կյանքի բազմազան ձևերի վրա, հարմարեցնելով նրանց իրենց բնակավայրի և գոյության բազմապիսի պայմաններին, կարելի է դատել հաջորդ գլուխներում բերվող ապացույցների ընդհանուր բովանդակությունից և գումարից։ Բայց մենք արդեն տեսանք, որ բնական ընտրությունը իր հետ բերում է մահաջնջում, իսկ թե ինչպիսի հսկայական դեր է կատարել մահաջնջումը օրգանական աշխարհի պատմության մեջ, այդ մասին պարզորոշ կերպով վկայում է երկրաբանությունը։ Բնական ընտրությունը առաջ է բերում նաև հատկանիշների տարամիտում, որովհետև օրգանական էակները ինչքան ավելի շատ են տարբերվում կառուցվածքով, ներքին կերտվածքով և սովորություններով, այնքան նրանք ավելի մեծ թվով կարող են գոյություն ունենալ տվյալ տարածության վրա, որի ապացույցը մենք կարող ենք գտնել, ուշադրություն դարձնելով որևէ փոքրիկ հողակտորի օրգանական բնակչության վրա և օտար երկրում նատուրալիզացիայի ենթարկված օրգանիզմների կազմի վրա։ Հետևապես, որևէ մի տեսակի հաջորդների փոփոխության պրոցեսի ընթացքում և իրենց թիվը ավելացնելու ձգտող բոլոր տեսակների միջև մղվող անընդհատ կռվում որքան ավելի բազմազան կլինեն այդ հաջորդները, այնքան նրանք ավելի հաջողության շանսեր կունենան գոյության կռվում։ Այսպիսով, միևնույն տեսակի տարատեսակները զանազանող փոքրիկ տարբերությունները ձգտում են մեծանալ ու հասնել տեսակային և նույնիսկ սեռային տարբերությունների աստիճանի։
Մենք համոզվեցինք, որ տվյալ դասի մեծ սեռերին պատկանող սովորական, լայնորեն տարածված և հեռու վայրերում տարաբնակված տեսակները ավելի շատ են փոփոխվում. նրանք կձգտեն իրենց փոփոխված հետնորդներին հաղորդել այն գերազանցությունը, որը նրանց դարձնում է տիրապետող իրենց երկրում։ Բնական ընտրությունը, ինչպես հենց նոր նկատեցինք, առաջ է բերում տարամիտում և կյանքի ավելի քիչ կատարելագործված ու միջանկյալ ձևերի զգալի ոչնչացում։ Ելնելով այս սկզբունքներից, հեշտությամբ կարելի է բացատրել նաև ամբողջ աշխարհում և յուրաքանչյուր դասի անհամար օրգանական էակների միջև, այսպես կոչված, ազգակցության և սովորաբար լավ նկատելի տարբերությունների բնույթը։ Այն փաստը, որ մենք չենք նկատում, այնքան նա սովորական է, իսկապես զարմանալի է, որ բոլոր կենդանիներն ու բույսերը տարածության և ժամանակի մեջ կապված են միմյանց հետ՝ կազմելով մեկը մյուսին ստորադասված խմբեր, և ընդսմին այնպիսի ձևով, ինչպես մենք այդ նկատում ենք ամեն մի քայլափոխում, այն է, որ միևնույն տեսակի տարատեսակները ամենից ավելի սերտորեն են կապված միմյանց հետ. ավելի քիչ սերտ և ավելի քիչ հավասարաչափ կերպով են կապված միմյանց հետ միևնույն սեռի տեսակները, որոնք կազմում են սեկցիաներ և ենթասեռեր. ավելի ևս պակաս մոտիկությամբ միմյանց հետ կապված են տարբեր սեռերի տեսակները և, վերջապես, սեռերը, որ ներկայացնում են փոխադարձ մոտիկության տարբեր աստիճաններ, որոնք արտահայտվում են ենթաընտանիքներով, ընտանիքներով, կարգերով, ենթադասերով և դասերով։ Զանազան ստորադասված խմբերը մի դասի սահմաններում չեն կարող դասավորվել ուղղագիծ շարքով, այլ խմբվում են որոշ կետերի շուրջը, որոնք իրենց հերթին դասավորվում են այլ կետերի շուրջը, և այսպես համարյա անվերջ շրջաններով։ Եթե տեսակները ստեղծված լինեին մեկը մյուսից անկախ կերպով, ապա անհնարին կլիներ այդպիսի դասակարգության համար բացատրություն գտնել, բայց նա բացատրվում է ժառանգականությամբ և բնական ընտրության բարդ ներգործությամբ, որը առաջ է բերում մահաջնջում և հատկանիշների տարամիտում, ինչպես այդ ցույց է տրված մեր աղյուսակի վրա։
Միևնույն դասին պատկանող բոլոր էակների ազգակցությունը երբեմն պատկերում են մեծ ծառի ձևով։ Ես կարծում եմ, որ այդ համեմատությունը շատ համապատասխանում է ճշմարտությանը։ Կանաչ ճյուղերը բացվող բողբոջներով ներկայացնում են ապրող տեսակներին, իսկ նախորդ տարիների ճյուղերը համապատասխանում են մահաջնջված տեսակների երկար շարքին։ Յուրաքանչյուր տարի աճող ճյուղերը աշխատել են առաջացնել ընձյուղներ բոլոր ուղղություններով և անցնել ու խեղդել հարևան ընձյուղներին և ճյուղերին, ճիշտ այդպես էլ տեսակները և տեսակների խմբերը բոլոր ժամանակներում գոյության մեծ կռվում հաղթահարել են մյուս տեսակներին։ Բնի ճյուղավորությունները, որոնք իրենց ծայրերում բաժանվում են սկզբում փոքրիկ ճյուղերի, իսկ հետո ավելի ու ավելի մանրիկ ճյուղերի, մի ժամանակ իրենք էլ, երբ ծառը դեռ մատաղ էր, եղել են բողբոջներով ծածկված ընձյուղներ. իսկ առաջվա և այժմյան բողբոջների միջև ճյուղավորվող ընձյուղների միջոցով եղած կապը հիանալի կերպով մեզ ներկայացնում է ապրող և մահաջնջված տեսակների դասակարգությունը, որը նրանց միացնում է խմբերում, որոնք ստորադասվում են այլ խմբերի։ Այն շատ ընձյուղներից, որոնք դուրս էին եկել այն ժամանակ, երբ ծառը դեռ բուն չէր տվել, թերևս ընդամենը երկուսը կամ երեքը պահպանվել են և աճել ու դարձել են մեծ ճյուղեր, որոնց վրա նստած են մնացած ճյուղերը, նույնն է տեղի ունեցել վաղուց անցած երկրաբանական դարաշրջաններում ապրած տեսակների հետ, նրանցից միայն քչերն են իրենցից հետո թողել դեռևս ապրող փոփոխված հաջորդներ։ Այդ ծառի կյանքի սկզբից շատ ոստեր ու ճյուղեր են չորացել ու թափվել. այդ թափված զանազան մեծության ճյուղերը ներկայացնում են ամբողջ կարգեր, ընտանիքներ և սեռեր, որոնք այժմ չունեն կենդանի ներկայացուցիչներ և մեզ հայտնի են միայն բրածո մնացորդներով։ Եվ ինչպես այստեղ որոշ տեղերում, հին ճյուղերի միջև երկճյուղավորման տեղում, նշվում է վտիտ մի ընձյուղ, որը պատահականության շնորհիվ ողջ է մնացել և շարունակում է իր ծայրին կանաչ մնալ, այդպես էլ որևէ բադակտուց կամ լեպիդոսիրեն, որը հավանորեն պաշտպանված բնակավայրի շնորհիվ փրկվել է օրհասական կռվից, իր ազգակցությամբ որոշ չափով միացնում է կյանքի երկու մեծ ճյուղեր։ Ինչպես որ բողբոջները աճման շնորհիվ սկիզբ են տալիս նոր բողբոջների, իսկ սրանք, եթե միայն նրանք բավականաչափ ուժեղ են, դառնում են ընձյուղներ, որոնք ճյուղավորվելով ծածկում և խեղդում են հյուծված շատ ճյուղեր, այդպես էլ, կարծում եմ, եղել է վերարտադրության շնորհիվ նաև կյանքի մեծ ծառը, որն իր մեռած թափված ճյուղերով լցրել է երկրի կեղևը և նրա մակերեսը ծածկում է իր հավետ ճյուղավորվող և հավետ հրաշալի ճյուղավորություններով։
ԳԼՈՒԽ V։ ՓՈՓՈԽԱԿԱՆՈՒԹՅԱՆ ՕՐԵՆՔՆԵՐԸ
ՓՈՓՈԽՎԱԾ ՊԱՅՄԱՆՆԵՐԻ ՆԵՐԳՈՐԾՈՒԹՅՈՒՆԸ։— ՕՐԳԱՆՆԵՐԻ ՎԱՐԺԵՑՈՒՄԸ ԵՎ ՈՉ ՎԱՐԺԵՑՈՒՄԸ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ԿԱՊԱԿՑՈՒԹՅԱՄԲ. ԹՌԻՉՔԻ ԵՎ ՏԵՍՈՂՈՒԹՅԱՆ ՕՐԳԱՆՆԵՐԸ։— ԱԿԿԼԻՄԱՏԻԶԱՑԻԱ։— ՀԱՐԱԲԵՐԱԿՑԱԿԱՆ ՓՈՓՈԽԱԿԱՆՈՒԹՅՈՒՆ։— ՓՈԽՀԱՏՈՒՑՈՒՄ (ԿՈՄՊԵՆՍԱՑԻԱ) ԵՎ ԱՃԻ ՏՆՏԵՍՈՒՄ։— ԿԵՂԾ ՀԱՐԱԲԵՐԱԿՑՈՒԹՅՈՒՆՆԵՐ։— ԲԱԶՄԻՑՍ ԿՐԿՆՎՈՂ ԹԵՐԱՃ ԵՎ ՑԱԾՐ ԿԱԶՄՎԱԾՔՈՎ ՄԱՐՄՆԱՄԱՍԵՐՆ ԱՄԵՆԻՑ ԱՎԵԼԻ ՓՈՓՈԽԱԿԱՆ ԵՆ։— ԱՐՏԱՍՈՎՈՐ ԿԵՐՊՈՎ ԶԱՐԳԱՑԱԾ ՄԱՐՄՆԱՄԱՍԵՐԸ ՎԵՐԻՆ ԱՍՏԻՃԱՆԻ ՓՈՓՈԽԱԿԱՆ ԵՆ։— ՏԵՍԱԿԱՅԻՆ ՀԱՏԿԱՆԻՇՆԵՐՆ ԱՎԵԼԻ ՓՈՓՈԽԱԿԱՆ ԵՆ, ՔԱՆ ՍԵՌԱՅԻՆՆԵՐԸ։— ԵՐԿՐՈՐԴԱԿԱՆ ՍԵՌԱԿԱՆ ՀԱՏԿԱՆԻՇՆԵՐԸ ՓՈՓՈԽԱԿԱՆ ԵՆ։— ՄԻԵՎՆՈՒՅՆ ՍԵՌԻ ՏԵՍԱԿՆԵՐԸ ՓՈՓՈԽՎՈՒՄ ԵՆ ՄԻԱՆՄԱՆ ՁԵՎՈՎ։— ՎԱՂՈՒՑ ԿՈՐՑՐԱԾ ՀԱՏԿԱՆԻՇՆԵՐԻ ՎԵՐԱԴԱՐՁԸ։— ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ։
Ես մինչև այժմ երբեմն այնպես եմ խոսել, կարծես թե այն փոփոխությունները, որ այնքան սովորական ու բազմազան են ընտանի էակների մեջ և ավելի հազվադեպ են բնական վիճակում, պատահականության արդյունք են։ Այդ արտահայտությունը, իհարկե, միանգամայն սխալ է, բայց դա պարզ կերպով երևան է հանում մեր անգիտակությունը փոփոխությունների պատճառի մասին յուրաքանչյուր մասնավոր դեպքում։ Մի քանի հեղինակներ կարծում են, թե վերարտադրողական սիստեմի գործունեության մեջ մտնում են հավասարապես թե զավակների ու ծնողների ճիշտ նմանության պահպանումը և թե կառուցվածքում անհատական տարբերությունների կամ թույլ շեղումների իրականացումը։ Բայց այն փաստը, որ շեղվող ձևերը և այլանդակությունները ավելի հաճախ հանդիպում են ընտանեցման, քան բնական դրության մեջ, ինչպես նաև լայն տարածված տեսակների փոփոխականությունը ավելի մեծ է, քան սահմանափակ տարածվածություն ունեցող տեսակներինը, բերում է հետևյալ եզրակացության, որ փոփոխականությունը սովորաբար շաղկապված է կենսապայմանների հետ, որոնց ենթարկվել է տեսակը մի քանի հաջորդական սերունդների ընթացքում։ Առաջին գլխում ես փորձել եմ ցույց տալ, որ փոփոխված պայմանները ներգործում են երկու ձևով՝ անմիջականորեն ամբողջ կազմվածքի վրա կամ նրա մի որոշ մասի վրա և անուղղակիորեն վերարտադրողական սիստեմի միջոցով։ Յուրաքանչյուր դեպքում մասնակցում են երկու գործոններ՝ օրգանիզմի բնույթը, որ երկուսից ամենակարևորն է, և ներգործող պայմանների բնույթը։ Փոփոխված պայմանների անմիջական ներգործությունը առաջ է բերում որոշակի և անորոշ հետևանքներ։ Երկրորդ դեպքում ամբողջ կազմվածքը կարծես դառնում է պլաստիկ, և մենք ստանում ենք մեծ, բայց տատանվող փոփոխականություն։ Առաջինում օրգանիզմի բնույթն այնպես է, որ նա հեշտությամբ է ենթարկվում որոշ պայմանների ներգործության և բոլոր կամ գրեթե բոլոր անհատները փոփոխվում են միանման ձևով։
Չափազանց դժվար է որոշել, թե որքան հեռուն է տարածվում կլիմայի, սննդի և այլ փոփոխված պայմանների ներգործությունը այդ ուղղությամբ։ Հիմք կա կարծելու, որ առաջներում նրանց ներգործությունը ավելի մեծ է եղել քան այն, ինչ կարելի է լիակատար ստուգությամբ ապացուցել։ Սակայն մենք կարող ենք վստահ լինել, որ անթիվ բարդ փոխադարձ հարմարումները, որոնք մենք ամենուրեք նկատում ենք բնության մեջ բազմազան օրգանիզմների մոտ, չի կարելի վերագրել միայն այդ ներգործությանը։ Հետևյալ դեպքերում կենսապայմաններն ըստ երևույթին ունեցել են ինչ որ, թեպետև թույլ, որոշակի ներգործություն։ Է. Ֆորբսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] պնդում է, որ կակղամորթների խեցիները նրանց տարածման հարավային սահմանում և միաժամանակ ծանծաղ ջրերում ունենում են ավելի վառ գույն, քան ավելի մեծ խորություն ունեցող ջրերում կամ ավելի հյուսիսային տեղերում ապրող միևնույն տեսակի խեցիները, սակայն այդ, իհարկե, ընդհանուր օրինաչափություն չէ։ Գուլդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 89] կարծում է, որ միևնույն տեսակի թռչունները պարզ մթնոլորտ ունեցող կլիմաներում ունենում են ավելի վառ գույն, քան ծովափնյա երկրներում կամ կղզիների վրա. իսկ Ուոլլաստոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 65] համոզված է այն բանում, որ ծովի հետ հարևանությամբ ապրելը ազդում է միջատների գույնի վրա։ Մոկեն-Տանդոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 90] բերում է այնպիսի բույսերի մի ցուցակ, որոնք ապրելով ծովափին, ձեռք են բերում որոշ չափով մսոտ տերևներ, թեև այլ վայրերում նրանք մսոտ չեն։ Այս թույլ փոփոխվող օրգանիզմները հետաքրքրական են այն տեսակետից, որ նրանց փոփոխությունների բնույթը համապատասխանում է այն տեսակների հատկություններին, որոնք մշտապես սահմանափակված են նման պայմաններով։
Եթե մի փոփոխություն, որ թեկուզ թույլ չափով օգտակար է դրանով օժտված էակին, ապա մենք չենք կարող ասել, թե ինչ չափով մենք պետք է այն վերագրենք բնական ընտրությանը, և ինչ չափով կենսապայմանների որոշակի ներգործությանը։ Այսպես, մորթեվաճառներին լավ հայտնի է այն փաստը, որ միևնույն տեսակի կենդանին որքան ավելի հյուսիսում է ապրում, այնքան ավելի խիտ և ավելի լավ կլինի նրա մորթին, բայց ով կկարողանա ասել, թե այդ փաստը ինչ չափով կախված է այն բանից, որ այդ տաք հագնված անհատները, որպես ավելի բարենպաստվածներ, մյուսների հանդեպ գերադասորեն պահպանվել են բազմաթիվ սերունդների ընթացքում, և ինչ չափով կախված է դաժան կլիմայի անմիջական ներգործությունից, որովհետև, ըստ երևույթին, կլիման իրոք անմիջական ներգործություն է ունենում մեր ընտանի չորքոտանիների մորթու վրա։
Կարելի է բերել օրինակներ, որ միևնույն տեսակի տարատեսակներ միանգամայն նման են միմյանց և առաջացել են այնքան տարբեր պայմաններում, որքան կարելի է պատկերացնել, իսկ մյուս կողմից բերել օրինակներ, որ միմյանց չնմանվող տարատեսակներ առաջացել են, ըստ երևույթին, միանգամայն միանման պայմաններում։ Նմանապես ամեն մի բնախույզի հայտնի են լիակատար անփոփոխության անհամար օրինակներ, չնայած նրան, որ նրանք ապրում են ամենահակառակ կլիմայական պայմաններում։ Նման նկատառումներն էլ ինձ դրդում են ավելի քիչ կշիռ տալու շրջապատող պայմանների անմիջական ներգործությանը, քան փոփոխականության ինչ որ ձգտման, որը կախված է մեզ բոլորովին անհայտ պատճառներից։
Միայն մի իմաստով կենսապայմանների ներգործությունը, կարելի է ասել, ոչ միայն փոփոխականություն է պատճառում անմիջականորեն կամ անուղղակի կերպով, այլև իր մեջ պարփակում է բնական ընտրությունը, այսինքն այն իմաստով, որ այդ պայմաններն են որոշում, թե արդյոք կապրի այս տարատեսակը, թե մյուսը։ Միայն երբ մարդն է հանդիսանում ընտրության ազդակ տվողը, մենք պարզ նկատում ենք փոփոխության այդ երկու տարբերի անկախ ներգործությունը, որովհետև այսպես թե այնպես փոփոխականությունը արտահայտվում է, բայց միայն մարդու կամքն է կուտակում այդ փոփոխությունները որոշ ուղղություններով, և ամենահարմարվածի գերապրումը բնության մեջ համապատասխանում է հենց այդ վերջին գործունեությունը։
ՕՐԳԱՆՆԵՐԻ ՈՒԺԵՂԱՑՈՂ ՎԱՐԺԵՑՄԱՆ ԵՎ ՈՉ ՎԱՐԺԵՑՄԱՆ ՆԵՐԳՈՐԾՈՒԹՅՈՒՆԸ ԿԱՊՎԱԾ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՀԵՏ
Առաջին գլխում բերված փաստերի հիման վրա, ինձ թվում է, չի կարելի կասկածել այն բանում, որ մեր ընտանի կենդանիների մի քանի օրգանների վարժեցումը ամրապնդել է այդ օրգանները և մեծացրել նրանց չափերը, իսկ ոչ վարժեցումը, ընդհակառակը, փոքրացրել է նրանց, ինչպես նաև այն բանում, որ նման փոփոխությունները ժառանգաբար փոխանցվում են։ Ազատ բնության մեջ համեմատության համար մենք չունենք նմուշներ, որոնց հիման վրա կարողանայինք դատել օրգանների տևական վարժեցման կամ ոչ վարժեցման ազդեցության մասին, որովհետև մեզ անհայտ են նախասկզբնական ձևերը։ Բայց շատ կենդանիներ ունեն այնպիսի օրգաններ, որոնց կառուցվածքը լավագույն կերպով կարելի է վերագրել նրանց ոչ վարժեցմանը։ Ինչպես նկատում է պրոֆեսոր Օուենը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] ամբողջ բնության մեջ չի կարելի գտնել ավելի մեծ անոմալիա, քան թռչել չկարողացող թռչունը. բայց և այնպես կան այդպիսի մի քանի թռչուններ։ Հարավ-ամերիկական կարճաթև բադը (logger headed duck) կարողանում է միայն ջրի մակերեսին խփել թևերով, որոնք նրա մոտ նույն վիճակում են, ինչ որ ընտանի էլսբերիական բադինը (Aylesbury duck)։ Ուշագրավ է այն, որ Կյոնինգհեմի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 91] դիտումների համաձայն, երիտասարդ թռչունները կարող են թռչել, մինչդեռ մեծերը կորցրել են այդ ընդունակությունը։ Քանի որ արածող խոշոր թռչունները հազվադեպ են թռչում, բացի այն դեպքերից, երբ փրկվում են վտանգից, ուստի հավանական է թվում, որ գիշատիչ գազաններից զուրկ օվկիանոսային կղզիների վրա ապրող կամ ոչ վաղուց ապրած թռչունների թևազուրկ վիճակը առաջացել է նրանց թռչելու օրգանների ոչ վարժեցման հետևանքով։ Ջայլամը, ճիշտ է, ապրում է մայր ցամաքների վրա և ենթարկվում է վտանգների, որոնցից չի կարող փրկվել թռիչքի միջոցով, բայց դրա փոխարեն նա պաշտպանվում է, աքացի տալով ոչ ավելի վատ, քան որևէ չորքոտանի։ Մենք կարող ենք ենթադրել, որ ջայլամի նախահայրը կենսակերպով նմանվել է արոսին և նրա մարմնի մեծության ու կշռի ավելացման հետ զուգընթաց մեկը մյուսին հաջորդող մի շարք սերունդների ընթացքում նրա ոտները ավելի ու ավելի են վարժվել, իսկ թևերը ավելի քիչ, մինչև որ, վերջապես, վերջնականապես անպետքացել են թռիչքի համար։
Կերբին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 92] նկատել է (ես էլ կարող եմ հաստատել նրա դիտողությունը), որ գոմաղբով սնվող բզեզների արուներից շատերի առջևի վերջավորությունների թաթերը հաճախ կոտրտված են։ Նա զննել է իր հավաքածուի մեջ եղած տասնյոթ օրինակ և ոչ մեկի վրա ոտների այդ մասի հետքն էլ չի գտել։ Onites appelles բզեզի ոտնաթաթերն այնպես միշտ բացակայում են, որ բզեզի նկարագրություններում նա նշված է որպես այդ մասերից զուրկ։ Մի քանի այլ սեռերի մեջ նրանք գտնվում են սաղմնային վիճակում։ Եգիպտացիների սրբազան բզեզը (Ateuchus) բոլորովին զուրկ է դրանցից։ Պատահական վնասվածքների ժառանգական փոխանցումը հաստատող վճռական վկայություն դեռևս չկա, բայց այն զարմանալի դեպքերը, որ դիտել է Բրոուն-Սեկարը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 93] երբ ծովախոզուկների վրա կատարված վիրահատության հետևանքները ժառանգական են եղել, պետք է մեզ դրդեն զգուշավոր լինել նաև այդ փաստի հնարավորությունը ժխտելու մեջ։ Համենայն դեպս Ateuchus-ի թաթերի լրիվ բացակայությունը և այլ սեռերի մեջ նրանց թերզարգացումը ավելի շրջահայաց է դիտել ոչ թե որպես ժառանգական վնասվածքների դեպքեր, այլ որպես վարժեցման տևական բացակայության հետևանքներ։ Քանի որ աղբաբզեզներից շատ շատերին մենք գտնում ենք կոտրատված թաթերով, ուստի այդ կորուստը պետք է տեղի ունենա նրանց կյանքի վաղ շրջանում, իսկ դա ցույց է տալիս, որ տվյալ օրգանները մեծ նշանակություն չեն կարող ունենալ և հետևապես զգալի վարժեցման չեն կարող ենթարկվել։
Այլ դեպքերում մենք կարող ենք վարժեցման և ոչ վարժեցման վերագրել կառուցվածքի այնպիսի ձևափոխությունները, որոնք բացառապես կամ գլխավորապես առաջացել են ընտրության միջոցով։ Միստր Ուոլլաստոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 65] հայտնագործել է մի ուշագրավ փաստ, որ Մադեյրայում ապրող բզեզների 550 տեսակից 200 տեսակը (այժմ հայտնի են ավելի շատ տեսակներ) այնքան թևազուրկ են, որ բոլորովին չեն կարողանում թռչել. տեղական քսանևինը սեռերից ոչ պակաս քան 23-ի բոլոր տեսակները աչքի են ընկնում այդ հատկությամբ։ Մի շարք փաստեր, այն է՝ որ աշխարհիս շատ երկրներում բզեզները քամուց քշվում են ծովը և ոչնչանում են, որ Ուոլլաստոնի դիտումների համաձայն, բզեզները Մադեյրայում թաքնվում են, մինչև որ քամին հանդարտում է և արևն սկսում է փայլել, որ անթև միջատների հարաբերական թիվն ավելի ևս մեծ է Դեզերտաս կղզու վրա, որը քամիների առջև ավելի բաց է, քան Մադեյրան, և առանձնապես այն արտասովոր փաստը, որի վրա հատկապես խիստ կերպով պնդում է Ուոլլաստոնը, որ բզեզների որոշ խմբեր, որոնք անպայման իրենց թևերի կարիքն զգում են և այլ երկրներում բազմաթիվ են, Մադեյրայում գրեթե բացակայում են,— այս բոլոր կշռադատումներն ինձ ստիպում են ենթադրելու, որ մադեյրական այդքան բազմաթիվ բզեզների թևազուրկ վիճակը կախված է գլխավորապես բնական ընտրությունից, կարող է պատահել, և ոչ վարժեցման հետ զուգորդված։ Որովհետև այդ բզեզների յուրաքանչյուր հաջորդ սերնդում ամեն մի անհատ, որ ավելի քիչ թռիչք է գործել թեկուզ թևերի աննշան թերզարգացման պատճառով կամ ավելի ծույլ սովորությունների պատճառով, ավելի շատ շանսեր է ունեցել գերապրելու, քանի որ ավելի քիչ է ենթարկվել ծովը քշվելու վտանգին, իսկ մյուս կողմից, այն բզեզները, որոնք ավելի հաճույքով են նետվել թռիչքներ գործելու, ավելի հաճախ են տարվել ծովը և ոչնչացել։
Բայց Մադեյրայի վրա այն միջատները, որոնք կերակրվում են ոչ թե գետնի վրա սողալով, այլ ծաղիկներով սնվող բզեզների ու թիթեռների նման ստիպված են սնունդ հայթայթելու համար բանեցնել իրենց թևերը, ինչպես կարծում է Ուոլլաստոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 65] ունեն ոչ թե փոքրացած, այլ նույնիսկ մեծացած թևեր։ Դա միանգամայն համատեղելի է բնական ընտրության ներգործության հետ։ Որովհետև, կղզու վրա որևէ միջատի առաջին անգամ երևալու դեպքում նրա թևերը փոքրացնելու կամ մեծացնելու համար բնական ընտրության ցուցաբերած ձգտումը պետք է կախված լիներ այն հանգամանքներից, թե արդյոք անհատների մեծ մասը փրկվում էր քամու դեմ հաջողությամբ պայքարելով, թե՞ այդ պայքարից խուսափելով և քիչ կամ բոլորովին չթռչելով։ Այդպես էլ կլիներ ափի մոտ խորտակված նավի նավաստիների հետ. լավ լողորդներին ձեռնտու կլիներ որքան կարելի է ավելի երկար լողալ և ցամաքին հասնել, իսկ վատ լողորդները ավելի լավ է, բոլորովին չփորձեն լողալ, այլ կառչեն նավի բեկորներին։
Խլուրդների և գետնի տակ ապրող մի քանի կրծողների աչքերը իրենց մեծությամբ սաղմնային են և որոշ դեպքերում բոլորովին ծածկված են մաշկով և մազով։ Նրանց աչքերի այդ վիճակը ամենայն հավանականությամբ կախված է այլասերումից, որն առաջացել է վարժեցման բացակայության հետևանքով, գուցե և բնական ընտրության օժանդակությամբ։ Հարավային Ամերիկայում գետնի տակ ապրող մի կրծող թուքու-թուքոն (Ctenomis) ավելի ևս ստորերկրյա կենսակերպ ունի, քան մեր խլուրդը, և մի իսպանացի, որը հաճախ էր որսացել այդպիսի կրծողներ, ինձ պատմում էր, որ նրանք հաճախ կույր են լինում։ Դրանցից մեկը, որին ես միառժամանակ ողջ պահում էի, իսկապես կույր էր, և ինչպես ցույց տվեց դիահերձումը, դրա պատճառը թարթաթաղանթի բորբոքումն է եղել։ Քանի որ աչքերի հաճախակի բորբոքումը պետք է վնասակար լինի ամեն մի կենդանու համար և քանի որ ստորերկրյա կենսակերպ ունեցող կենդանուն աչքերը հարկավոր չեն, ուստի նրանց փոքրացումը, որին ուղեկցում է կոպերի մոտենալը և նրանց մազակալելը, կարող է միայն օգտակար լինել, իսկ այդ դեպքում բնական ընտրությունը, ի հարկե, կօժանդակե ոչ վարժեցման հետևանքներին։
Հայտնի է, որ շատ կենդանիներ, որոնք պատկանում են ամենատարբեր դասերի և ապրում են Կարնիոլիայի և Կենտուկկիի[45] ստորերկրյա այրերում, բոլորովին կույր են։ Մի քանի խեցեմորթների մոտ աչքի ցողունիկը պահպանվել է, բայց աչքն ինքը անհետացել է, այսինքն հեռադիտակի հաստատոցը պահպանվել է, բայց հեռադիտակն իր ապակիների հետ միասին արդեն գոյություն չունի։ Քանի որ դժվար է ենթադրել, որ աչքերը, թեկուզև օգուտ չտվող, կարող էին այս կամ այն ձևով վնասել մթության մեջ ապրող օրգանիզմներին, ուստի նրանց կորուստը պետք է վերագրել վարժեցման բացակայությանը։ Այդ կույր կենդանիներից մեկը, այն է՝ քարանձավային մեծամկները — (Neotoma), որոնցից մի զույգ բռնել է պրոֆեսոր Սիլլիմանը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 94] քարանձավի մուտքից կես մղոն հեռավորության վրա և, հետևաբար, քարանձավի ոչ ամենախոր տեղում, ունեցել են փայլուն և զգալի մեծությամբ աչքեր։ Այդ կենդանիները, ինչպես ինձ հաղորդում է պրոֆ. Սիլլիմանը, երբ նրանց ենթարկել են մոտավորապես մի ամսվա ընթացքում շարունակ ուժեղացող լույսի ներգործությանը, սկսել են աղոտ կերպով զանազանել առարկաները։
Դժվար է պատկերացնել ավելի նման կենսապայմաններ, քան այն պայմանները, որ ներկայացնում են գրեթե միանման կլիմա ունեցող կրաքարային խոր քարանձավները, այնպես որ, հին տեսակետի համաձայն, ըստ որի կույր կենդանիները ամերիկական և եվրոպական քարանձավների համար առանձին առանձին են ստեղծվել, պետք էր սպասել, որ նրանց կազմվածքը և սիստեմատիկական ազգակցությունը մոտ նմանություն կունենան միմյանց հետ։ Ակներևորեն դա չի արդարանում, երբ երկու կենդանական աշխարհն ամբողջությամբ համեմատում ենք միմյանց հետ։ Շիեդտեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 95] մի քանի միջատների վերաբերմամբ նկատում է. «Հետևաբար, մենք այդ երևույթն ամբողջությամբ վերցրած պետք է համարենք մի ինչ որ բացառապես տեղական բան, իսկ այն նմանության մեջ, որ նկատելի է Մամոնտի այրի (Կենտուկկիի) և Կարնիոլիայի քարանձավների սակավաթիվ ձևերի միջև, կարելի է տեսնել այն համանմանության միայն մի պարզ արտահայտությունը, որ գոյություն ունի առհասարակ Հյուսիսային Ամերիկայի և Եվրոպայի կենդանական աշխարհների միջև»։ Իմ տեսակետից՝ ամերիկական կենդանիները, որ օժտված են եղել տեսողության սովորական սրությամբ, դանդաղորեն ամբողջ մի շարք սերունդների ընթացքում արտաքին աշխարհից շարունակ ավելի ու ավելի թափանցել են Կենտուկկիի քարանձավների խորքը. նույնն է տեղի ունեցել նաև եվրոպական քարանձավներում եղած կենդանիների հետ։ Մենք որոշ վկայություններ ունենք կենսակերպում տեղի ունեցած այդ աստիճանական փոփոխության օգտին, որովհետև Շիեդտեն նկատում է. «Հետևաբար մենք դիտում ենք ստորերկրյա կենդանական աշխարհները որպես հարակից վայրերի աշխարհագրական տեսակետից մեկուսացած կենդանական աշխարհների փոքրիկ ճյուղեր, որոնք թափանցել են երկրի խորքը և որքան նրանք տարածվել են խավարի այդ շրջանում, այնքան ավելի ու ավելի հարմարվել են շրջապատող պայմաններին։ Սովորական ձևերից քիչ տարբերվող կենդանիները նախապատրաստում են անցումը լույսից դեպի մթություն։ Այնուհետև գալիս են այն ձևերը, որոնք հարմարված են մշտական աղջամուղջին և, վերջապես, այն ձևերը, որոնք նախանշված են լիակատար խավարի համար, սրանք արդեն ունեն բոլորովին առանձնահատուկ կառուցվածք»։ Շիեդտեի դիտողությունները, չպետք է մոռանալ, վերաբերում են ոչ թե մի, այլ տարբեր տեսակների։ Այն ժամանակամիջոցում, երբ կենդանին անթիվ սերունդների շարքից հետո հասել է ամենախոր խորխորատներին, մեր արտահայտած տեսակետի հիման վրա վարժեցման բացակայությունը առաջ է բերել աչքերի ավելի կամ պակաս չափով կորուստ, իսկ բնական ընտրությունն արդեն կարողացել է առաջացնել այլ փոփոխություններ, օրինակ, բողկուկների կամ շոշափուկների մեծացում՝ որպես տեսողության կորստին փոխհատուցում։ Չնայած այդ փոփոխություններին, մենք, այնուամենայնիվ, իրավունք ունենք սպասելու, որ Ամերիկայի քարանձավային միջատները ազգակցության գծեր կունենան մայր ցամաքի այլ բնակիչների հետ, ճիշտ այդպես էլ այն կենդանիները, որոնք ապրում են Եվրոպայի քարանձավներում, ազգակցությամբ, կապված կլինեն այդ մայր ցամաքի բնակիչների հետ։ Եվ դա արդարանում է ամերիկական քարանձավային միջատների վերաբերմամբ, ինչպես ինձ հաղորդել է պրոֆ. Դանան.[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 96] նմանապես էլ մի քանի եվրոպական քարանձավային միջատներ մոտ ազգակցություն ունեն շրջապատող երկրի միջատների հետ։ Չափազանց դժվար կլիներ բանական բացատրություն տալ քարանձավային կույր կենդանիների և երկու մայր ցամաքների այլ բնակիչների միջև եղած այդ ազգակցությանը, կողմնակից լինելով այն սովորական պատկերացման, որ նրանք ստեղծված են իրարից անկախ կերպով։ Այն փաստը, որ ձին և նոր աշխարհի քարանձավային կենդանիներից մի քանիսը ներկայացնում են մոտ ազգակցություն, կարելի էր նախատեսել այդ երկու աշխարհների կենդանիների մեծ մասի լավ հանրածանոթ ազգակցության հիման վրա։ Քանի որ Battyscia-ի մի կույր տեսակը առատորեն հանդիպում է քարանձավներից հեռու ստվերոտ ժայռերի վրա, ուստի այդ միակ սեռի քարանձավային տեսակների տեսողության կորուստը հավանորեն ոչ մի առնչություն չունի նրա մութ բնակավայրի հետ։ Մի այլ կույր սեռ (Anophthalmus) ունի այն զարմանալի առանձնահատկությունը, որ ըստ Մուրրեյի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 43] խոսքերի, նրա տեսակները դեռ ոչ մի տեղ չեն գտնված, բացի քարանձավներից. և այնուամենայնիվ Եվրոպայի և Ամերիկայի զանազան քարանձավներում հանդիպող տեսակները տարբերվում են իրարից։ Սակայն հնարավոր է, որ այդ զանազան տեսակների նախնիները անցյալում, երբ նրանք դեռ օժտված էին աչքերով, ցրված են եղել երկու մայր ցամաքների վրա և հետագայում մահաջնջվել են ամենուրեք, բացի իրենց այժմյան մեկուսացած ապաստարաններից։ Ես հեռու եմ զարմանք արտահայտելուց այն անոմալիաների առթիվ, որ ներկայացնում են քարանձավային կենդանիները, ինչպես այն արտահայտել է Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] կույր ձկան (Amblyopsis) վերաբերմամբ, բայց ավելի շուտ այդ կարելի էր անել կույր պրոտեի առթիվ, եթե նրան համեմատում ենք Եվրոպայի այլ երկկենցաղների հետ։ Ես զարմանում եմ նրա վրա, որ այդ մռայլ ապաստարաններում նախկին կյանքի ավելի շատ մնացորդներ չեն պահպանվել շնորհիվ ավելի քիչ դաժան մրցության, որին ենթարկվել է նրանց սակավաթիվ բնակչությունը։
ԱԿԼԻՄԱՏԻԶԱՑԻԱ
Բույսերն ունեն ժառանգական սովորություններ, որոնք դրսևորվում են ծաղկելու և քնելու ժամանակի, սեղմերի ծլման համար անձրևաջրի անհրաժեշտ քանակության մեջ և այլն.— այս հանգամանքն ինձ դրդում է մի քանի խոսք ասել ակլիմատիզացիայի մասին։ Քանի որ շատ հաճախ միևնույն սեռի սահմաններում հանդիպում են տեսակներ, որոնք ապրում են տաք և ցուրտ երկրներում, ապա, ընդունելով այն դրույթի ճշտությունը, որ միևնույն սեռի բոլոր տեսակներն առաջացել են ընդհանուր ծնողներից, մենք պետք է ակնկալենք, որ նրանք սերունդների երկար շարանների ընթացքում կարող են հաջողությամբ կլիմայավարժվել։ Ամենքին հայտնի է, որ յուրաքանչյուր տեսակ հարմարված է իր բնակլիմայական պայմաններին. արկտիկական կամ նույնիսկ բարեխառն գոտու տեսակները չեն դիմանում արևադարձային կլիմային և ընդհակառակը։ Այդպես էլ մսալի բույսերից շատերը չեն դիմանում խոնավ կլիմայի։ Բայց իրենց սովորական կլիմային տեսակների հարմարվածության աստիճանը հաճախ չափազանցնում են։ Մենք այդ իրավունք ունենք եզրակացնելու այն փաստից, որ հաճախ անհնարին է նախօրոք ասել, թե այդպիսի բույսը կդիմանա մեր կլիմային կամ ոչ, ինչպես նաև այն դիտողությունից, որ ամենատարբեր երկրների բույսերն ու կենդանիները մեզ մոտ միանգամայն առողջ են լինում։ Մենք լիովին հիմք ունենք ենթադրելու, որ տեսակներն իրենց բնական դրության մեջ մնում են իրենց տարաբնակման սահմաններում որքան որոշ կլիմայական պայմանների հարմարվելու շնորհիվ, նույնքան էլ, եթե ոչ ավելի, այլ օրգանիզմների դեմ մրցելու շնորհիվ։ Արդյոք միշտ այդ հարմարումը սազացնում է բույսերին կլիմայական պայմաններին թե ոչ, համենայն դեպս մի քանի բույսերի վերաբերմամբ մենք ունենք պարզորոշ ապացույց, որ նրանք կարող են բնականորեն վարժվել զանազան ջերմաստիճանների, այսինքն՝ կլիմայավարժվել։ Այսպես, օրինակ, սոճիները և ռոդոդենդրոնները, որոնք ստացված են Հիմալայների զանազան բարձրություններից Հուկերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] հավաքած սերմերից, Անգլիայում ցուցաբերել են տարբեր չափի ցրտադիմացկունություն։ Միստր Թվեյտսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 98] հաղորդել է ինձ, որ նա նման երևույթներ նկատել է Ցեյլոնում. նույնպիսի դիտումներ կատարել է Ուոտսոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 62] Ազորյան կղզիներից Անգլիա բերված եվրոպական տեսակների վրա. իսկ ես կարող եմ բերել նաև այլ օրինակներ։ Ինչ վերաբերում է կենդանիներին, ապա կարելի է բերել տեսակի լայն տարածման մի քանի արժանահավատ դեպքեր, որ տեղի են ունեցել պատմական ժամանակաշրջանում տաք երկրներից դեպի ավելի ցուրտ երկրները և ընդհակառակը։ Սակայն մենք ստույգ չգիտենք, թե արդյոք այդ կենդանիները խիստ կերպով հարմարված են իրենց բնական կլիմային, թեև մենք այդ համարում ենք որպես ապացուցված, մենք չգիտենք նույնպես և այն, թե արդյոք նրանք իրենց նոր հայրենիքում կլիմայավարժվե՞լ են ժամանակի ընթացքում, արդյոք հետագայում ավելի լա՞վ են հարմարվել նրա պայմաններին, քան եղել են սկզբում։
Քանի որ մենք իրավունք ունենք ենթադրելու, որ մեր ընտանի կենդանիներին սկզբնապես ընտրել է կիսավայրենի մարդը նրանց օգտակար հատկությունների համար և այն պատճառով, որ հեշտությամբ կարող էին բազմանալ գերության մեջ, այլ ոչ թե այն պատճառով, որ հետագայում ընդունակ եղան լայնորեն տարաբնակվելու, ուստի մեր ընտանի կենդանիների սովորական և զարմանալի այն առանձնահատկությունը, որ նրանք ոչ միայն դիմանում են զանազան կլիմաների, այլ նաև միաժամանակ պահպանում են իրենց պտղաբերությունը (որն անհամեմատ ավելի խիստ փորձանշան է) կարող է ներկայացվել որպես փաստարկ այն դրույթի օգտին, որ բնական վիճակում ապրող կենդանիների մեծ մասը կարող է գոյություն ունենալ խիստ տարբեր կլիմաներում։ Բայց մենք չպետք է չափազանց լայն կերպով կիրառենք այս փաստարկը, հիշելով որ մեր ընտանի ցեղերը հավանորեն ծագել են մի քանի տեսակներից. այսպես, օրինակ, մեր շների մեջ կարող է խառնված լինել արևադարձային կամ արկտիկական որևէ գայլի արյունը։ Թեև մուկը և մեծամուկը չեն կարող ընտանեցված կենդանիներ անվանել, այնուամենայնիվ մարդու միջոցով նրանք տարածվել են աշխարհի զանազան կողմերը և այժմ ավելի լայն տարածում ունեն, քան որևէ այլ կրծող, նրանք ապրում են հյուսիսում՝ Ֆարերյան կղզիների ցուրտ կլիմայում և հարավում՝ Ֆալկլանդյան կղզիների, ինչպես նաև տաք գոտու շատ կղզիների վրա։ Հետևաբար, որևէ հատուկ կլիմայի հարմարվելու ընդունակությունը կարելի է դիտել որպես մի առանձնահատկություն, որը շատ հեշտությամբ պատվաստվում է կենդանիների մեծամասնությանը հատուկ կառուցվածքային բնածին մեծ ճկունության հողի վրա։ Այս տեսակետից մարդու և նրա ընտանի կենդանիների կողմից ամենաբազմազան կլիմաների հարմարվելու ընդունակությունը, ինչպես նաև այն փաստը, որ մահաջնջված փղերը և ռնգեղջյուրները առաջներում դիմացել են սառցադաշտային շրջանին, մինչդեռ նրանց այժմ ապրող տեսակները իրենց սովորություններով պատկանում են բացառապես արևադարձային և մերձարևադարձային կլիմային,— այս փաստերը չպետք է դիտվեն որպես անոմալիա, այլ միայն որպես օրգանիզմների բնույթի շատ սովորական ճկունության օրինակներ, մի ճկունություն, որ երևան է գալիս առանձին հանգամանքներում։
Թե որևէ յուրահատուկ կլիմայի վերաբերմամբ տվյալ տեսակի հարմարեցման երևույթը ինչ չափով է կախված միայն նրա հատկություններից, ինչ չափով զանազան բնածին առանձնահատկություններ ունեցող տարատեսակների բնական ընտրությունից և, վերջապես, ինչ չափով ամբողջությամբ միասին վերցրած երկու պատճառներից էլ, այդ բոլորը դեռևս մութ հարցեր են։ Որ սովորությունը իր ներգործությունն է ունենում, դրանում ինձ համոզում են ոչ միայն անալոգիաները, այլև գյուղատնտեսական աշխատությունների մեջ, սկսած հնագույն չինական հանրագիտարաններից, հաճախակի հանդիպող մշտական խորհուրդները, որ կենդանիներին տեղից տեղ փոխադրելիս պետք է ծայրահեղ զգուշություն պահպանել։ Որովհետև անհավանական է, որ մարդը կարողանար տվյալ վայրերին հատկապես հարմարված կառուցվածքով այդքան մեծ թվով ցեղեր և ենթացեղեր ընտրել, ապա այդ արդյունքը, ես կարծում եմ, պետք է վերագրել սովորությանը։ Մյուս կողմից, բնական ընտրությունը անխուսափելիորեն պետք է հակվեր այն անհատների պահպանման կողմը, որոնք ծնվել են իրենց բնակված երկրներին ամենից ավելի հարմարված կառուցվածքով։ Մշակովի այս կամ այն բույսին նվիրված մասնագիտական աշխատություններում հիշատակվում է, որ մի քանի տարատեսակներ ավելի հեշտությամբ են դիմանում մի որոշ կլիմայի, քան մյուսները. դա առանձնապես զարմանալիորեն դրսևորվում է պտղաբուծության վերաբերյալ աշխատություններում, որ հրատարակվում են Միացյալ Նահանգներում։ Այդ աշխատությունների մեջ սովորաբար որոշ տարատեսակներ հանձնարարվում են հյուսիսային, իսկ մյուսները հարավային նահանգների համար և քանի որ այդ տարատեսակների մեծ մասը նորագույն ծագում ունի, ապա ակներև է, որ նրանք իրենց կազմությամբ պարտական չեն սովորությանը։ Գետնախնձորը, որ Անգլիայում երբեք սերմերով չի բազմացվում, և հետևապես, նոր տարատեսակներ չի տվել, բերվում է իբրև ապացույց կլիմայավարժեցման փաստի անհնարինությունը ցույց տալու համար, որովհետև նա այժմ էլ նույնքան զգայուն է, ինչքան միշտ էլ եղել է։ Նույն նպատակով էլ և մեծ հիմնավորմամբ հաճախ բերվել է լոբու օրինակը։ Բայց քանի դեռ որևէ մեկը չի փորձի ցանել իր լոբին առնվազն քսան սերունդների ընթացքում այնքան վաղ, որ մեծ մասը սառնամանիքից ոչնչանա, չի հավաքի ողջ մնացածներից սերմեր, միաժամանակ մանրազնին կերպով խուսափելով պատահական խաչաձևումներից և կրկին չի ստանա սերմեր այդ սերմնաբույսերից, պահպանելով նույն նախազգուշությունները,— մինչև այդ կարելի է համարել, որ այդ իմաստով դեռևս ոչ մի փորձ չի ձեռնարկվել։ Եվ չի կարելի կարծել, թե լոբու ծիլերի կազմության մեջ ոչ մի տարբերություն երևան չի բերվել. ցուցումներ այն մասին, թե ինչքան տարբեր է լինում այդ բույսի զանազան ծիլերի դիմացկունությունը, հանդիպում են մամուլում, և ես ինքս դիտել եմ այդ փաստի ապշեցուցիչ օրինակներ։
Ընդհանուր առմամբ մենք կարող ենք գալ այն եզրակացության, որ սովորությունը, վարժեցումը կամ ոչ վարժեցումը որոշ դեպքերում մեծ դեր են խաղացել կազմության և կառուցվածքի փոփոխության մեջ, բայց այդ պատճառների հետևանքները զգալի չափով ամրապնդվել կամ գերազանցվել են բնածին փոփոխությունների բնական ընտրությամբ։
ԿՈՌԵԼՅԱՏԻՎ (ՀԱՐԱԲԵՐԱԿՑԱԿԱՆ) ՓՈՓՈԽԱԿԱՆՈՒԹՅՈՒՆ
Այդ արտահայտությամբ ես հասկանում եմ այն փաստը, որ էակի ամբողջ կազմվածքը աճման ու զարգացման ժամանակ գտնվում է այնպիսի սերտ փոխադարձ կապի մեջ, որ երբ նրա որևէ մասում երևան են գալիս թույլ փոփոխություններ և կուտակվում են բնական ընտրության միջոցով, մյուս մասերը նույնպես ենթարկվում են փոփոխության։ Սա մի չափազանց կարևոր հարց է, որը դեռ պարզ կերպով չի հասկացվում և, անկասկած, այստեղ միանգամայն տարբեր փաստերի ամբողջ խմբեր կարող են խառնվել։ Մենք ամենից առաջ կհամոզվենք, որ ժառանգականության պարզ փաստը կարող է ընդունվել որպես նման հարաբերակցություն։ Ամենից ավելի ակներև դեպքերից մեկն է հանդիսանում թրթուրների կամ ձագերի կառուցվածքում փոփոխությունների և չափահաս կենդանու կառուցվածքի միջև եղած կախումը։ Մարմնի զանազան մասերը, որ հոմոլոգ են միմյանց հետ, վաղ, սաղմնային վիճակում ունեն նման կառուցվածք և անհրաժեշտորեն ենթակա են նման պայմանների, ըստ երևույթին առանձնապես հակում ունեն միանման ձևով փոփոխվելու։ Դրա օրինակները մենք տեսնում ենք մարմնի բոլորովին միանման ձևով փոփոխվող աջ և ձախ մասերի մեջ, առջևի և հետևի վերջավորությունների և նույնիսկ վերջավորությունների ու ծնոտների համատեղ փոփոխությունների մեջ, որովհետև ստորին ծնոտները, մի քանի կազմախոսների կարծիքով, վերջավորությունների հոմոլոգներ են։ Այդ ձգտումները, անկասկած, կարող են լիովին կամ մասամբ հաղթահարվել բնական ընտրության ներգործությամբ, այսպես, օրինակ, հայտնի է մի դեպք, երբ եղջերուների մի ամբողջ ընտանիք եղջյուրներ ունեցել է գլխի միայն մի կողմում, եթե այդ առանձնահատկությունը կարողանար որևէ օգուտ բերել, ապա նա հավանորեն կարող էր կայունացվել բնական ընտրությամբ։
Հոմոլոգ մասերը, ինչպես այդ նկատել են մի քանի հեղինակներ, հակում ունեն սերտաճելու, դա հաճախ նկատվում է այլանդակ բույսերի վրա, և ոչ մի բան այնքան սովորական չէ, որքան հոմոլոգ գոյացումների սերտաճումը նորմալ կառուցվածքում, ինչպես, օրինակ, երբ պսակաթերթիկները սերտաճում և կազմում են խողովակ։ Կոշտ մասերը ներգործում են իրենց հարակից փափուկ մասերի վրա. մի քանի հեղինակներ կարծում են, որ թռչունների կոնքի ձևի բազմազանությունը ազդում է նրանց երիկամների ձևի զարմանալի բազմազանության վրա։ Ուրիշները կարծում են, որ մարդու մոտ մոր կոնքի ձևը իր գործած ճնշման միջոցով ազդում է երեխայի գլխի ձևի վրա։ Շլեգելի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 99] կարծիքով օձերի մարմնի ձևը և սնունդը կուլ տալու եղանակը որոշում են մի քանի ներքին ամենակարևոր օրգանների դիրքն ու ձևը։
Այդ հարաբերակցության պատճառը հաճախ բոլորովին մութ է. Իսիդոր Ժոֆրուա Սենտ-Իլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 16] պնդել է, որ մի քանի այլանդակություններ շատ հաճախ, իսկ մյուսները, ընդհակառակը, շատ հազվագյուտ դեպքերում են ուղեկցում միմյանց, թեպետև մենք ի վիճակի չենք այդ փաստին որևէ բացատրություն տալու։ ի՞նչը կարող է ավելի տարօրինակ լինել, քան կատուների մեջ հանդիպող այն կախումը, որ գոյություն ունի մի կողմից բոլորովին սպիտակ մազերի ու կապույտ աչքերի և մյուս կողմից խլության միջև, կամ կրիային նմանվող խայտաբղետ գույնի և իգական սեռի միջև։ Իսկ աղավնիների մեջ՝ ի՞նչը կարող է օտարոտի լինել, քան այն կապը, որ մենք տեսնում ենք փետրավորման և արտաքին մատները միացնող թաղանթի միջև, կամ ձվից նոր դուրս եկած ձագուկի աղվամազով ծածկվելու աստիճանի և նրա փետուրի ապագա գույնի միջև։ Եվ, վերջապես, մերկ տաճկական շան մազի և ատամների միջև եղած կապը, թեև այստեղ, անկասկած, դեր է խաղում նաև հոմոլոգիան։ Ինչ վերաբերում է այս վերջին հարաբերակցությանը, ապա ես կարծում եմ, որ հազիվ թե կարելի է պատահական համարել այն փաստը, որ կաթնասունների երկու կարգեր, որոնք իրենց մաշկային ծածկույթի տեսակետից ամենից ավելի անկանոն են, այսինքն կետանմանները (Cetacea) և թերատամավորները (Edentata, ինչպես, օրինակ, զրահակիրները, թեփամորթները և այլն), միաժամանակ ամենից ավելի անկանոն են նաև իրենց ատամներով։ Այնուամենայնիվ՝ միստր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] այդ կանոնից այնքան բացառություններ է նշել, որ նա քիչ նշանակություն ունի։
Ինձ հայտնի չէ մի օրինակ, որ ավելի հարմար լիներ հարաբերակցության և փոփոխականության օրենքների կարևորությունը պարզաբանելու համար, անկախ օգտակարությունից և, հետևաբար, անկախ բնական ընտրության մասնակցությունից, քան այն, որ ընձեռում է մեզ բարդածաղիկների և հովանոցավորների ծաղկաբույլերի արտաքին ու ներքին ծաղիկների միջև եղած տարբերությունը։ Ո՞ւմ հայտնի չէ, օրինակ, մարգարտածաղկի եզրային և միջին ծաղիկների միջև եղած տարբերությունը, և այդ տարբերությունը հաճախ ուղեկցվում է վերարտադրողական օրգանների մասնակի կամ լիակատար թերզարգացմամբ։ Այդ բույսերից մի քանիսների նաև սերմերը ձևով և մակերեսի կառուցվածքով ներկայացնում են տարբերություններ։ Այդ տարբերությունները երբեմն վերագրվել են ծածկույթի թերթիկների ճնշմանը ծաղիկների վրա կամ ծաղիկների փոխադարձ ճնշմանը, և բարդածաղիկների եզրային ծաղիկներում սերմերի ձևը որոշ չափով հաստատում է այդ բացատրությունը։ Սակայն, ինչպես ինձ հաղորդում է Հուկերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] հովանոցավորների ամենևին ոչ այն տեսակները, որոնք ունեն ամենից ավելի խիտ ծաղկաբույլեր, աչքի են ընկնում արտաքին ու ներքին ծաղիկների միջև ամենամեծ տարբերությամբ։ Կարելի էր կարծել, որ եզրային պսակաթերթիկների զարգացումը խլելով վերարտադրողական օրգաններից սննդանյութեր, նրանց թերզարգացման պատճառ է դառնում, բայց դա հազիվ թե միակ պատճառն է, որովհետև բարդածաղիկներից շատերի արտաքին ու ներքին ծաղիկների սերմերը տարբերվում են միմյանցից, թեև նրանց պսակների միջև տարբերություն չկա։ Գուցե այդ տարբերությունները կապ ունեն դեպի եզրային և միջին ծաղիկները գնացող սննդանյութերի հոսանքի տարբերության հետ, համենայն դեպս մենք գիտենք, որ անկանոն ծաղիկներ ունեցող բույսերի այն ծաղիկները, որոնք գտնվում են ծաղկի առանցքի գագաթին, ամենից ավելի հաճախ են ներկայացնում պելորիայի երևույթ, այսինքն աննորմալ վերադարձ դեպի կանոնավոր ձևը։ Կարող եմ իբրև օրինակ, որպես հարաբերակցության մի զարմանալի դեպք բերել այն, որ շատ պելարգոնիաների ծաղկաբույլի մեջ միջին ծաղկի վերին երկու պսակաթերթիկները հաճախ կորցնում են մգագույն բծերի և այդ դեպքում նրանց հարակից նեկտարանոցը լինում է բոլորովին անզարգացած, այսպիսով, միջին ծաղիկը պելորիկ է, կամ կանոնավոր, իսկ երբ մգագույն բիծը բացակայում է միայն մեկ պսակաթերթիկի վրա, նեկտարանոցը բոլորովին չի ոչնչանում, այլ միայն զգալիորեն կարճանում է։
Պսակի զարգացման վերաբերմամբ շատ հավանական է Շպրենգելի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 82] ենթադրությունը, որ եզրային ծաղիկները ծառայում են միջատներին գրավելու համար, որոնց այցելությունը չափազանց ձեռնտու է կամ նույնիսկ անհրաժեշտ է այդ բույսերի բեղմնավորման համար, իսկ եթե այդպես է, ապա բնական ընտրությունը կարող է այստեղ ցուցաբերել իր ներգործությունը։ Սակայն սերմերի վերաբերմամբ անհնարին է թվում, որ նրանց ձևի մեջ եղած տարբերությունները, որոնք ոչ միշտ հարաբերակցության մեջ են գտնվում պսակաթերթիկներում եղած որևէ տարբերությունների հետ, կարող են որևէ ձևով օգտակար լինել։ Բայց և այնպես հովանոցավորների այդ տարբերությունները այնքան ակներևորեն կարևոր են, եզրային ծաղիկները երբեմն տալիս են ուղիղ էնդոսպերմով սերմեր (Orthospermeae), իսկ միջին ծաղիկները՝ կոր էնդոսպերմով (Ceolospermeae),— որ դը-Կանդոլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] հովանոցավորների ընտանիքի գլխավոր ստորաբաժանումների հիմքում դրել է այդ հատկանիշները։ Այստեղից հետևում է, որ այն տարբերությունները, որոնք կարգաբանների կողմից ընդունվում են որպես շատ կարևոր, կարող են կախված լինել բացառապես փոփոխականության և հարաբերակցության (կոռելյացիայի) օրենքներից, չներկայացնելով, որքան մենք կարող ենք այդ մասին դատել, նվազագույն օգուտ անգամ։
Հաճախ մենք կարող ենք սխալմամբ հարաբերակցական փոփոխականության վերագրել կառուցվածքի այն առանձնահատկությունները, որոնք ընդհանուր են տեսակների ամբողջ խմբերի համար, իրականում իրենց ծագումով պարզապես պարտական են ժառանգականությանը։ Մի որևէ հեռավոր նախահայր կարող էր բնական ընտրության շնորհիվ ձեռք բերել այս կամ այն փոփոխությունն իր կառուցվածքում, և ապա հազարավոր սերունդներ անցնելուց հետո՝ մի ուրիշը, առաջինից բոլորովին անկախ կերպով. այդ փոփոխությունները փոխանցվելով տարբեր կենսակերպ ունեցող հաջորդների մի ամբողջ խմբի, մեզ կարող է թվալ, կարծես թե գտնվում են անհրաժեշտ հարաբերակցության մեջ։ Մի քանի այլ հարաբերակցություններ ըստ երևույթին կարող են առաջանալ միայն այն ներգործություններից, որոնք բացառապես հատուկ են բնական ընտրությանը։ Այսպես, օրինակ, Ալֆոնս դը-Կանդոլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] ցույց է տվել, որ թռիչքաթևեր ունեցող սերմերը երբեք չեն հանդիպում չպայթող պտուղներում. ես այդ կանոնը կբացատրեի ելնելով այն բանից, որ բնական ընտրությունը չէր կարողանա առաջացնել թևավոր սերմեր, եթե տուփիկները չբացվեին. միայն այդ դեպքում կարող էին թևավոր սերմերը, որոնք ավելի լավ են տարվում քամուց, ստանալ առավելություններ լայն տարածման համար ավելի քիչ հարմարվածների հանդեպ։
ՓՈԽՀԱՏՈՒՑՈՒՄ (ԿՈՄՊԵՆՍԱՑԻԱ) ԵՎ ԱՃԻ ՏՆՏԵՍՈՒՄ
Ժոֆֆրուա ավագը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 3] և Գյոթեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 101] գրեթե միաժամանակ հայտարարել են աճման փոխհատուցման կամ հավասարակշռության իրենց օրենքը, որը Գյոթեն արտահայտել է այսպես. «Բնությունը, որպեսզի առատաձեռն լինի մի ուղղությամբ, ստիպված է ժլատանալու մի այլ ուղղությամբ»։ Ես կարծում եմ, որ այդ որոշ իմաստով վերաբերում է մեր ընտանի ցեղերին։ Եթե սննդարար հյութերը առատությամբ առհոսում են դեպի մի օրգանը, ապա նրանք հազվադեպ են հոսում, համենայն դեպս առատորեն չեն հոսում դեպի մի այլ օրգան. դժվար է հասնել այն բանին, որ կովը շատ կաթ տա և միևնույն ժամանակ հեշտությամբ գիրանա։ Կաղամբի միևնույն տարատեսակները չեն տալիս միաժամանակ առատ և սննդարար տերևներ և յուղատու սերմերի առատ բերք։ Երբ մեր պտուղներում սերմերը ատրոֆիայի են ենթարկվում, պտուղները դրանից շահում են մեծությամբ և որակով։ Մեր հավացեղերի մեջ գլխի վրա փետուրների մեծ փուփուլը միշտ ուղեկցվում է կատարի փոքրացմամբ, իսկ կզակի տակի փետրափունջը՝ ականջօղերի փոքրացմամբ։ Բնական վիճակում գտնվող տեսակների վերաբերմամբ հազիվ թե կարելի լինի պաշտպանել այս օրենքի ընդհանրականությունը, բայց մանրազնին դիտողներից շատերը, առավելապես բուսաբանները, համարում են այն որպես ճշմարտություն։ Այնուամենայնիվ ես այստեղ չեմ բերի օրինակներ որովհետև գրեթե հնարավոր չեմ համարում տարբերել այդ երկու երևույթները, այն է՝ մի կողմից օրգանի զգալի զարգացումը բնական ընտրության շնորհիվ և մի այլ, հարևան օրգանի թերզարգացումը միևնույն պատճառով կամ ոչ վարժեցման հետևանքով, մյուս կողմից նույնպիսի հետևանքները, որ առաջանում են մի օրգանից սննդանյութերը դեպի մյուս, ուժեղ աճող, հարևան օրգանին հատկացնելուց։
Ես կասկածում եմ, որ փոխհատուցման առաջադրված դեպքերից մի քանիսը, ինչպես նաև մի քանի այլ փաստեր կարող են մի ավելի լայն սկզբունքի հիմք կառուցել, այն է՝ բնական ընտրությունը միշտ ձգտում է խնայողություն պահպանել կազմվածքի բոլոր մասերի վերաբերմամբ։ Եթե փոփոխված կենսապայմաններում առաջուց օգտակար եղած օրգանը դառնում է ավելի քիչ օգտակար, այդ դեպքում բնական ընտրությունը կնպաստի նրա կրճատմանը, որովհետև անհատների համար ձեռնտու է սննդանյութ չվատնել անօգուտ մասի կառուցման վրա։ Միայն այս տեսակետով ես կարող եմ ինձ բացատրել այն փաստը, որը ինձ չափազանց զարմացրել էր փնջոտնյա խեցգետինների (Cirripedia) ուսումնասիրման ժամանակ, և դրան համանման շատերը, այն է՝ երբ մի փնջոտնյա խեցգետին մակաբույծ կյանք է վարում մի ուրիշի մարմնի մեջ և այդպիսով պաշտպանություն է գտնում, ապա նա կատարելապես կամ մասամբ կորցնում է իր կեղևը կամ խեցին։ Այդ մենք նկատում ենք Ibla-ի արուի մոտ և զարմանալի ձևով Proteolepas-ի մոտ։ Մնացած բոլոր փնջոտնյաների խեցին բաղկացած է գլխի առջևի չափազանց կարևոր հատվածներից, որոնք հսկայական չափով են զարգացած և օժտված են ուժեղ մկաններով և նյարդերով։ Բայց մակաբուծական կյանք վարող և այդ պատճառով էլ պաշտպանված Proteolepas-ի գլխի ամբողջ առջևի մասը այլասերվել ու դարձել է մի աննշան թերաճիկ, որն ամրակցված է բռնիչ շոշափուկների հիմքին։ Հասկանալի է, որ այդպիսի մի մեծ ու բարդ, արդեն ավելորդ դարձած օրգանից զրկվելը այդ տեսակի ամեն մի անհատի համար բացահայտ օգուտ կներկայացներ, որովհետև գոյության կռվում, որին ենթակա է ամեն մի կենդանի, նրանցից յուրաքանչյուրը իր գոյությունը պահպանելու համար կունենար այնքան ավելի շատ շանսեր, որքան նրան ավելի քիչ հատուկ լիներ սննդանյութերի ապարդյուն ծախսումը։
Այսպիսով, ես կարծում եմ, բնական ընտրությունը ժամանակի ընթացքում կձգտի կրճատել կազմվածքի ամեն մի մաս, հենց որ նա փոփոխված կենսակերպի պատճառով ավելորդ կդառնա, առանց որևէ այլ մասի համապատասխան ավելացրած զարգացում ստանալու։ Եվ ընդհակառակը, բնական ընտրությունը կարող է առաջ բերել որևէ օրգանի զարգացում, կարիք չունենալով պարտադիր փոխհատուցման համար կրճատել այդ օրգանի հարևան որևէ մի մաս։
ԲԱԶՄԻՑՍ ԿՐԿՆՎՈՂ, ԹԵՐԱՃ ԵՎ ՑԱԾՐ ԿԱԶՄՎԱԾՔՈՎ ՄԱՐՄՆԱՄԱՍԵՐԸ ՓՈՓՈԽԱԿԱՆ ԵՆ
Ըստ երևույթին, պետք է որպես կանոն ընդունել այն, ինչպես այդ նկատել է Իսիդոր Ժոֆֆրուա Սենտ-Իլերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 16] որ ինչպես տեսակների, այնպես էլ տարատեսակների մեջ փոփոխական է լինում այն օրգանների թիվը, որոնք միևնույն անհատի մոտ բազմիցս կրկնվող են (օրինակ, օձերի ողները, առէջքները բազմայր ծաղիկներում). ընդհակառակը, եթե նույն օրգանը կամ նույն մարմնամասը թվով ավելի քիչ է, ապա այդ թիվը հաստատուն է լինում։ Նույն գիտնականը, իսկ նրանից հետո նաև մի քանի բուսաբաններ նկատել են, որ բազմիցս կրկնվող մարմնամասերը փոփոխական են նաև իրենց կառուցվածքով։ Քանի որ «վեգետատիվ կրկնությունը», եթե այստեղ օգտագործենք պրոֆ. Օուենի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] այդ արտահայտությունը, հանդիսանում է ավելի ցածր կազմվածքի հատկանիշ, ապա հենց նոր արտահայտված դրույթները համընկնում են բնախույզների մեջ ավելի լայն տարածված այն կարծիքի հետ, որ կազմվածքի ավելի ցածր աստիճանների վրա կանգնած էակներն ավելի փոփոխական են, քան ավելի բարձր կազմվածքով օրգանիզմները։ Ես կարծում եմ, որ կազմվածքի ցածր աստիճան ասելով այստեղ հասկանում ենք առանձին օրգանների գործունեության մասնագիտացման ավելի ցածր աստիճան, իսկ քանի դեռ մի մարմնամասը կատարում է մի քանի տարբեր աշխատանքներ, մենք կարող ենք բացատրել, թե ինչու նա պահպանում է իր փոփոխականությունը, այսինքն, թե ինչու բնական ընտրությունը չի վերացնում ամեն մի փոքրիկ շեղում, ինչպես այդ արվում է այն դեպքում, երբ որոշ մաս կատարում է միայն մի հատուկ աշխատանք։ Նույն հիման վրա էլ դանակը, որով կտրում են ամեն տեսակի առարկաներ, կարող է համարյա ամեն ձևի լինել, մինչդեռ միայն մի որոշակի աշխատանքի համար նախասահմանված գործիքը պետք է ունենա միանգամայն որոշակի ձև։ Բնական ընտրությունը, այդ երբեք չպետք է մոռանալ, ներգործում է միայն տվյալ էակի օգտին և այդ օգուտի միջոցով։
Թերզարգացած (ռուդիմենտար) մարմնամասերը, ինչպես ընդունում են բոլորը, չափազանց փոփոխական են։ Այդ խնդրին մենք դեռ վերադառնալու ենք. այստեղ ես միայն կնկատեմ, որ նրանց փոփոխականությունը հավանորեն տեղի է ունենում նրանց անօգտակարությունից, որի հետևանքով բնական ընտրությունը չէր կարող սահմանափակել շեղումների առաջացման այդ ձգտումը։
ՄԻ ՄԱՐՄՆԱՄԱՍ, ՈՐԸ ՉԱՓԱԶԱՆՑ ԿԱՄ ԲԱՑԱՌԻԿ ՁԵՎՈՎ ԶԱՐԳԱՑԱԾ Է ՄԻ ՈՐԵՎԷ ՏԵՍԱԿԻ ՄՈՏ՝ ՀԱՄԵՄԱՏԱԾ ԱԶԳԱԿԻՑ ՏԵՍԱԿՆԵՐԻ ՆՈՒՅՆ ՄԱՐՄՆԱՄԱՍԵՐԻ ՀԵՏ, ԵՐԵՎԱՆ է ՀԱՆՈՒՄ ՈՒԺԵՂ ՓՈՓՈԽԱԿԱՆՈՒԹՅԱՆ ՀԱԿՈՒՄ
Մի քանի տարի առաջ ես զարմացել էի միստր Ուոտերհաուզի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 87] այդ իմաստով արած դիտողության վրա։ Ըստ երևույթին պրոֆ. Օուենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] նույնպես եկել է նման եզրակացության։ Բայց ինձ թվում է, թե համարյա անհուսալի է որևէ մեկին համոզելու փորձը այդ դրույթի անկասկածելիության մասին, չբերելով իմ հավաքած փաստերի երկար շարանները, որոնք, իհարկե, այստեղ բերվել չեն կարող։ Ես միայն կարող եմ իմ համոզմունքը արտահայտել այն բանում, որ այդ կանոնը ամենալայն կիրառություն է գտնում։ Ես շատ լավ գիտակցում եմ, որ նման դատողությունների ժամանակ հնարավոր են սխալների մի քանի աղբյուրներ, բայց հույսով եմ, որ դրանք ես նկատի եմ առել։ Ամենից առաջ պետք է առանձին ուշադրություն դարձնել այն հանգամանքի վրա, որ այդ կանոնը ոչ մի դեպքում չի վերաբերում օրգաններին, ինչքան էլ նրանք արտասովոր կերպով զարգացած լինեն, եթե միայն այդ օրգաններով օժտված տեսակը կամ տեսակները դրանով չեն տարբերվում իրենց մոտիկ այլ շատ տեսակներից։ Այսպիսով, չղջիկի թևը գոյացում է, որը չափազանց աննորմալ է կաթնասունների ամբողջ դասի համար, և, այնուամենայնիվ, այդ կանոնը նրա վերաբերմամբ չի կարող կիրառվել, քանի որ չղջիկների ամբողջ խումբն օժտված է թևերով. այդ կանոնը կիրառելի կլիներ միայն այն դեպքում, եթե որևէ տեսակ նույն սեռին պատկանող տեսակների համեմատությամբ ունենար անսովոր կերպով զարգացած թևեր։ Այդ կանոնը առանձնապես խիստ կերպով վերաբերում է երկրորդական սեռական հատկանիշներին, երբ նրանք արտահայտվում են որևէ անսովոր ձևով։ «Երկրորդական սեռական հատկանիշներ» տերմինը Հունտերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 102] (Hunter) կիրառում է այն հատկանիշների վերաբերմամբ, որոնք պատկանում են միայն մի սեռի, թեև անմիջական առնչություն չունեն վերարտադրման բուն գործողության հետ։ Կանոնը վերաբերում է թե՛ արուներին և թե՛ էգերին, բայց վերջիններին ավելի փոքր չափով, որովհետև նրանք ավելի հազվագյուտ դեպքերում են ունենում աչքի ընկնող երկրորդական սեռական հատկանիշներ։ Պատճառը, թե ինչու այդ կանոնը այդքան խստորեն կիրառելի է երկրորդական սեռական հատկանիշներին, թերևս այն է, որ այդ հատկանիշները աչքի են ընկնում մեծ փոփոխականությամբ, անկախ նրանից, թե արդյոք նրանք բացառիկ զարգացում ունեն, թե ոչ։ Այս փաստի վերաբերմամբ, ես կարծում եմ, հազիվ թե կարելի է կասկածել։ Բայց որ մեր կանոնը չի սահմանափակվում միայն երկրորդական սեռական հատկանիշներով, պարզ կերպով երևում է հերմոֆրոդիտ փնջոտնյաների օրինակից. ուսումնասիրելով այդ ընտանիքը, ես հատկապես ի նկատի եմ ունեցել Ուոտերհաուզի դիտողությունը և լիովին համոզված եմ, որ այդ կանոնը համարյա միշտ արդարանում է։ Ես մի այլ աշխատության մեջ կբերեմ ամենից ավելի ուշագրավ դեպքերի ցուցակը, իսկ այստեղ կսահմանափակվեմ միայն մեկ օրինակով, որը կպարզաբանի կանոնն իր ամենալայն կիրառման մեջ։ Նստակյաց կյանք վարող փնշոտնյաների (Cirripedia ծովային ժայռերի) ծածկափեղկերը, որ նրանց կազմվածքի բառիս լիակատար իմաստով չափազանց կարևոր մասեր են, շատ քիչ են տարբերվում նույնիսկ տարբեր սեռերի մեջ. բայց Pyrgoma սեռի մի քանի տեսակների մոտ այդ փեղկերը ունենում են զարմանալի չափերով բազմազանություն, զանազան տեսակների հոմոլոգ փեղկերը իրենց ձևով բոլորովին տարբեր են լինում։ Միևնույն տեսակի պատկանող անհատների մոտ այդ տարբերության աստիճանը այնքան մեծ է, որ առանց չափազանցության մեջ ընկնելու կարելի է ասել, որ միևնույն տեսակին պատկանող տարատեսակները այդ կարևոր օրգանների կառուցվածքում ներկայացնում են ավելի խոր տարբերություններ, քան տարբեր սեռերի պատկանող տեսակների միջև եղած տարբերությունները։
Քանի որ միևնույն երկրում ապրող թռչունների միևնույն տեսակի անհատները միմյանցից շատ քիչ են տարբերվում, ուստի ես հատուկ ուշադրություն եմ դարձրել նրանց վրա, և այս կանոնը, իհարկե, այդ դասի վերաբերմամբ էլ արդարանում է։ Ես չեմ կարողացել համոզվել, թե այս կանոնը կիրառելի՞ է արդյոք նաև բույսերի նկատմամբ և այդ, իհարկե, զգալիորեն կխախտեր իմ հավատը դեպի այդ կանոնի ճշտությունը, եթե միայն բույսերի ընդհանուր փոփոխականության զգալի չափերը առանձնապես չեն դժվարացնի նրանց հարաբերական փոփոխականության աստիճանի գնահատումը։
Երբ մենք տեսնում ենք, որ բույսի որոշ տեսակի որևէ օրգան կամ մաս զարգացած է բացառիկ կերպով կամ անսովոր չափով, բնական է ենթադրել, որ նրանք կարևոր նշանակություն ունեն տեսակի համար։ Այնուամենայնիվ, հենց այդ դեպքերում նրանք աչքի են ընկնում իրենց առանձնապես խիստ արտահայտված փոփոխականության հակումով։ Բայց ինչո՞ւ է այդպես։ Ելնելով այն ենթադրությունից, որ յուրաքանչյուր տեսակ իր բոլոր մասերով ստեղծվել է մյուսներից անկախ կերպով, ապա ես բացատրություն գտնելու հնարավորություն չեմ տեսնում։ Բայց ենթադրելով, որ տեսակների խմբերը առաջացել են այլ տեսակներից և փոփոխվել են բնական ընտրության միջոցով, ինձ թվում է, թե կարելի է որոշ լույս սփռել այդ հարցի վրա։ Ամենից առաջ թույլ տվեք ինձ մի քանի նախնական դիտողություններ անելու։ Եթե մեր ընտանի կենդանիների օրգանիզմի որևէ մասը կամ ամբողջ կենդանին անուշադրության մատնվի և նրա վերաբերմամբ ընտրություն չկիրառվի, ապա այդ մասը (օրինակ, դորկինգյան հավերի կատարը) կամ ամբողջ ցեղը կկորցնի իր միակերպությունը և, կարելի է ասել, կսկսի այլասերվել։ Թերաճ օրգաններում և այն օրգաններում, որոնք բավարար չափով չեն հարմարված որևէ որոշակի նպատակի համար, կարող է պատահել նաև պոլիմորֆ խմբերում, մենք տեսնում ենք դրան զուգահեռ դեպք, որովհետև այստեղ բնական ընտրությունը կամ չի կարողացել երևան գալ, կամ պարզապես չի դրսևորվել իր ամբողջ ուժով, կազմվածքը թողնելով ինչ որ երերվող վիճակում։ Բայց այստեղ ավելի մոտ առնչություն ունի մի այլ փաստ, այսինքն այն, որ այն առանձնահատկությունները, որոնք մեր ընտանի կենդանիների կազմվածքում մշտական ընտրության ազդեցության տակ ամենից ավելի արագ են ենթարկվում փոփոխության, նույնպես երևան են հանում փոփոխվելու ծայրահեղ ենթարկվողություն։ Համեմատեցեք աղավնիների միևնույն ցեղի անհատներին միմյանց հետ, և դուք կզարմանաք փոփոխականության այն հսկայական չափերի վրա, որ նկատվում են թուրմանների կտուցների, փոստատար աղավնու կտուցների և ականջաօղակների, փողապոչ աղավնու ընդհանուր կերպարանքի և պոչի մեջ և այլն, այսինքն այն մասերի, որոնց վրա ուշադրություն են դարձրել անգլիական աղավնասերները։ Նույնիսկ միևնույն ենթացեղի, օրինակ կարճակտուց թուրմանի սահմաններում խիստ դժվար է ստանալ կատարելության մոտ թռչուններ, որովհետև շատերը շեղվում են տիպիկ նմուշից։ Իսկապես կարելի է ասել, որ անընդհատ պայքար է տեղի ունենում մի կողմից օրգանիզմների ավելի նվազ կատարյալ ձևի վերադառնալու հակման, ինչպես նաև նոր շեղումներ առաջացնելու բնածին ձգտման և մյուս կողմից ցեղի մաքրությունը պահպանող անշեղ ընտրության ուժի միջև։ Վերջ ի վերջո հաղթահարում է ընտրությունը, և մենք երբեք չենք վախենա այնպիսի անհաջողություն կրելու, որ կարճակտուց թուրմանից կստանանք այնպիսի կոպիտ թռչուն, ինչպիսին է սովորական թուրմանը։ Բայց քանի դեռ ընտրությունը առաջ է ընթանում, միշտ էլ կարելի է սպասել փոփոխականության զգալի աստիճան այն մասերում, որոնք ենթարկվում են նրա ներգործությանը։
Իսկ այժմ դառնանք դեպի բնությունը։ Երբ մի որևէ տեսակի մեջ կազմվածքի որոշ մասը նույն սեռի այլ տեսակների համեմատությամբ զարգացել է բացառիկ ձևով, մենք կարող ենք եզրակացնել, որ այդ մասը ենթարկվել է արտասովոր փոփոխության արդեն այն ժամանակաշրջանից հետո, երբ այդ բոլոր տեսակները բաժանվել են միմյանցից որպես ամբողջ սեռի մի ընդհանուր նախահորից առաջացած ճյուղեր։ Այդ ժամանակաշրջանը չի կարող լինել շատ հեռավոր, որովհետև տեսակները հազվագյուտ դեպքում են պահպանվում մեկ երկրաբանական ժամանակաշրջանից ավելի երկար։ Փոփոխության անսովոր մեծությունը ենթադրել է տալիս նաև փոփոխականության անսովոր զգալի մեծություն և տևականություն, որի ընթացքում բնական ընտրության միջոցով փոփոխությունները կուտակվել են հօգուտ տեսակի։ Բայց քանի որ արտասովոր կերպով զարգացած մասի կամ օրգանի փոփոխականությունը եղել է այդքան մեծ և այդքան տևական, այն էլ մի ժամանակաշրջանում, որն առանձնապես հեռու չէ, ուստի մենք իրավունք ունենք, որպես ընդհանուր կանոն, սպասելու, որ այդ մասերում էլ կգտնենք պլաստիկության ավելի մեծ աստիճան, քան մնացած մասերում, որոնք ավելի երկար ժամանակաշրջան պահպանվել են համարյա անփոփոխ։ Ես համոզված եմ, որ հենց այդպես էլ լինում է իրոք։ Ես ոչ մի առիթ չեմ տեսնում կասկածելու այն բանում, որ մի կողմից բնական ընտրության և մյուս կողմից վերադարձի հակման ու փոփոխականության միջև եղած պայքարը, վերջապես, կդադարի և ամենաաննորմալ կերպով զարգացած օրգանները կարող են կայունանալ։ Նշանակում է, որևէ օրգան, ինչքան էլ նա նորմայից շեղվի, եթե գրեթե անփոփոխ վիճակում փոխանցվել է բազմաթիվ փոփոխված հաջորդների,— ինչպես այդ եղել է չղջիկի թևերի հետ,— ապա պետք է նա մեր տեսության հիման վրա գոյություն ունենար հսկայական ժամանակաշրջանների ընթացքում գրեթե նույն դրության մեջ, որպեսզի դառնար նույնքան քիչ ենթակա փոփոխականության, ինչպես կազմվածքի ամեն մի այլ մասն է։ Միայն այն դեպքերում, երբ փոփոխությունը վաղուց չի տեղի ունեցել և շատ մեծ է, մենք դեռ պետք է հանդիպենք այդ բնածին փոփոխականությանը (generative variability), ինչպես կարող ենք այն անվանել, որովհետև այդ դեպքերում հազվադեպ է, որ փոփոխականությունը լիովին ամրացած լինի պահանջվող ուղղությամբ ու չափովի փոփոխվող անհատների երկարատև ընտրության միջոցով և նախկին, ավելի քիչ փոփոխված վիճակին վերադառնալու հակում ունեցող անհատների տևական վերացման միջոցով։
ՏԵՍԱԿԱՅԻՆ ՀԱՏԿԱՆԻՇՆԵՐՆ ԱՎԵԼԻ ՓՈՓՈԽԱԿԱՆ ԵՆ, ՔԱՆ ՍԵՌԱՅԻՆՆԵՐԸ
Այն սկզբունքը, որը քննարկման առարկա է եղել նախորդ պարագրաֆում, կարող է կիրառվել նաև ներկա խնդրի նկատմամբ։ Հանրածանոթ է, որ տեսակային հատկանիշներն ավելի փոփոխական են, քան սեռայինները։
Պարզաբանենք այդ օրինակով. եթե մի մեծ սեռի մեջ մի քանի տեսակներ ունենային կապույտ ծաղիկներ, իսկ մյուսները՝ կարմիր, ապա նրանց գույնը կլիներ միայն տեսակային հատկանիշ, և ոչ ոքի չէր զարմացնի, եթե կապույտ ծաղիկներ ունեցող տեսակներից մեկը փոխակերպվեր կարմրի կամ ընդհակառակը. բայց եթե բոլոր ծաղիկները կապույտ լինեին, ապա այդ արդեն կլիներ սեռային հատկանիշ, և նրա փոփոխությունը կներկայացներ արդեն ավելի անսովոր երևույթ։ Ես դիտմամբ ընտրեցի այդպիսի օրինակ, որի վերաբերմամբ կիրառելի չէ այն բացատրությունը, որ սովորաբար այդպիսի դեպքերում առաջարկում են մի քանի բնախույզներ, այն է՝ տեսակային հատկանիշներն ավելի փոփոխական են, որովհետև նրանք վերաբերում են ֆիզիոլոգիական տեսակետից ավելի պակաս էական մասերի, քան նրանք, որոնց հիման վրա որոշում են սեռերը։ Ես կարծում եմ, որ այդ բացատրությունը մասամբ ճիշտ է, թեև ոչ ուղղակի իմաստով, դրան ես կվերադառնամ դասակարգության նվիրված գլխում։ Ավելորդ կլիներ փաստեր բերել հաստատելու, որ սովորական տեսակային հատկանիշներն ավելի փոփոխական են, քան սեռայինները, բայց էական հատկանիշների վերաբերմամբ բնապատմական աշխատություններում ես բազմիցս նկատել եմ հետևյալը՝ եթե հեղինակը զարմանքով նշում է, որ որևէ օրգան կամ մաս, որը տեսակների մի մեծ խմբի մոտ հաստատուն է, մերձավոր որևէ տեսակների մեջ խիստ տարբերվում է, ապա միաժամանակ այդ օրգանը կամ մասը այդ տեսակի տարբեր անհատների մոտ լինում է փոփոխական։ Իսկ այդ փաստը ապացուցում է, որ սովորաբար սեռային նշանակություն ունեցող հատկանիշը այն դեպքերում, երբ նա իջնում է մինչև տեսակային մակարդակի, հաճախ դառնում է փոփոխական, ընդ որում չփոխելով իր ֆիզիոլոգիական նշանակությունը։ Նման մի բան կիրառելի է նաև այլանդակությունների նկատմամբ. գոնե Իսիդոր Ժոֆֆրուա Սենտ-Իլերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 16] ըստ երևույթին, չի կասկածում, որ որքան նորմալ օրգանը միևնույն խմբի տեսակների մոտ ավելի է տարբերվում, այնքան ավելի շատ նա ենթակա է անոմալիաների առանձին անհատների մոտ։
Տեսակները որպես անկախ ստեղծագործական ակտերի արդյունքներդ ընդունող տեսակետի հիման վրա ինչո՞վ կարելի է բացատրել, թե ի՞նչ պատճառով այն օրգանը, որը տարբերվում է նման մասերից միևնույն սեռին պատկանող անկախ կերպով ստեղծված տեսակների մեջ, պետք է ավելի մեծ չափով փոփոխական լինի, քան կազմվածքի այն մասերը, որոնք տարբեր տեսակների մոտ նման են միմյանց։ Ես որևէ բացատրություն տալու հնարավորություն չեմ տեսնում։ Բայց ելնելով այն դրույթից, որ տեսակները միայն ցայտուն կերպով արտահայտված և ավելի կայուն տարատեսակներ են, մենք իրավունք ունենք սպասելու, որ հաճախ նրանց կգտնենք դեռ փոփոխման պրոցեսի մեջ հատկապես նրանց այն մասերի վերաբերմամբ, որոնք արդեն փոփոխվել են համեմատաբար ոչ վաղ ժամանակաշրջանում և այդպիսով սկսել են տարբերվել մերձավոր ձևերի նման մասերից։ Այդ հարցը կարելի է շարադրել նաև այսպես՝ այն հատկանիշները, որոնցով մի սեռի պատկանող տեսակները նմանվում են իրար և որոնցով նրանք տարբերվում են հարևան սեռերից, կոչվում են սեռային. այդ հատկանիշները կարելի է վերագրել մի որևէ ընդհանուր նախահոր, որովհետև ավելի կամ պակաս տարբեր կենցաղաձևի հարմարված զանազան տեսակները միայն շատ հազվագյուտ դեպքերում կարող էին փոփոխվել բնական ընտրության միջոցով միանգամայն միանման ձևով. և քանի որ այդ այսպես կոչված սեռային հատկանիշները ժառանգվել են դեռ այն ժամանակաշրջանից առաջ, երբ ընդհանուր նախահորից առաջին անգամ բաժանվել են այն ճյուղերը, որոնք համապատասխանում են հետագայում փոփոխված և տարբերություններ դրսևորած ժամանակակից տեսակներին, ապա և քիչ հավանական է, որ նրանք սկսել են փոփոխվել ներկա ժամանակներում։ Մյուս կողմից, այն հատկանիշները, որոնցով միևնույն սեռին պատկանող տեսակները տարբերվում են միմյանցից, կոչվում են տեսակային, քանի որ այդ հատկանիշները առաջացել են արդեն այն ժամանակաշրջանից հետո, երբ տեսակները ճյուղավորվել են, հեռանալով իրենց ընդհանուր նախահորից, ապա հավանական է, որ նրանք այժմ էլ դեռ պահպանել են փոփոխականության մի որոշ բաժին, համենայնդեպս ավելի մեծ, քան կազմվածքի այն մասերում, որոնք շատ երկար ժամանակաշրջանի ընթացքում պահպանվել են առանց փոփոխությունների։
ԵՐԿՐՈՐԴԱԿԱՆ ՍԵՌԱԿԱՆ ՀԱՏԿԱՆԻՇՆԵՐԸ ՓՈՓՈԽԱԿԱՆ ԵՆ
Ես կարծում եմ, բնախույզները կհամաձայնվեն ինձ հետ առանց մանրամասնությունների մեջ ընկնելու, որ երկրորդական սեռական հատկանիշները խիստ փոփոխական են։ Նմանապես, ես կարծում եմ, հեշտությամբ կընդունեն, որ միևնույն խմբի տեսակները միմյանցից ավելի շատ են տարբերվում իրենց երկրորդական սեռական հատկանիշներով, քան իրենց կազմվածքի այլ գծերով։ Համեմատեցեք, օրինակ, այն տարբերության աստիճանը, որ ունեն հավերի ընտանիքի թռչունների արուները, որոնց երկրորդական սեռական հատկանիշները այնպես խիստ զարգացած են, էգերի տարբերության աստիճանի հետ։ Այդ հատկանիշների սկզբնական փոփոխականության պատճառը պարզ չէ, բայց մենք կարող ենք բացատրել, թե ինչու նրանք դարձել են այնպես կայուն և միակերպ, ինչպես մյուս հատկանիշները, որովհետև նրանք կուտակվում են սեռական ընտրության ազդեցությամբ, որն իր հետևանքով այնքան խիստ չէ, ինչպես բնական ընտրությունը, վերջինիս հետևանքը մահվան է հանգում, մինչդեռ առաջինը միայն սահմանափակում է ավելի քիչ հարմարված արուի սերունդը։ Ինչ էլ ուզում է լինի երկրորդական սեռական հատկանիշների փոփոխականության պատճառը, համենայն դեպս նրանց չափազանց փոփոխականության շնորհիվ սեռական ընտրության ներգործության համար լայն ասպարեզ է բացվել, և նա կարողացել է այդ խմբի տեսակներին հաղորդել տարբերության ավելի զգալի աստիճան սեռական և այլ հատկանիշների տեսակետներից։
Ուշագրավ է այն փաստը, որ միևնույն տեսակի երկու սեռերը միմյանցից տարբերող երկրորդական սեռական հատկանիշները սովորաբար վերաբերում են կազմվածքի հատկապես այն մասերին, որոնցով միևնույն սեռի տեսակները տարբերվում են միմյանցից։
Այդ փաստը հաստատելու համար ես կբերեմ իմ ցուցակի սկզբին գտնվող առաջին երկու օրինակը, և քանի որ այդ դեպքերում երևան եկող տարբերությունները շատ արտասովոր բնույթ ունեն, ուստի հարընչությունը հազիվ թե կարող է պատահական լինել։ Թաթի մեջ հոդերի միանման թիվ բզեզների ամբողջ մեծ խմբերի համար կայուն հատկանիշ է, բայց Engidae-ի մոտ ինչպես նկատել է Ուեստվուդը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 71] նրանց թիվը զգալիորեն տատանվում է, և միաժամանակ այդ թիվը տատանվում է միևնույն տեսակի թե՛ արուների և թե՛ էգերի մոտ։ Ճիշտ նույնն է նաև այն թաղանթաթևերի մոտ, որոնք իրենց համար բնիկներ են փորում ավազի մեջ, ջղերի դասավորությունը թևերի վրա չափազանց կարևոր հատկանիշ է, որովհետև նա կայուն է մեծ խմբերի համար, բայց մի քանի սեռերում տարբեր տեսակների մոտ ջղավորումը տարբերվում է, ինչպես նաև այն տարբեր է միևնույն տեսակի տարբեր սեռի անհատների մոտ։
Սըր Ջ. Լեբբոկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 60] վերջերս նկատել է, որ մի քան մանր խեցեմորթներ ներկայացնում են այդ օրենքի հիանալի օրինակ. «Pontella-ի, օրինակ, սեռական հատկանիշները վերաբերում են միայն առջևի բողկուկներին և ոտների հինգերորդ զույգին, տեսակային հատկանիշները նույնպես սահմանափակվում են գլխավորապես այդ օրգաններով»։ Այդ հարաբերությունը ինձ համար միանգամայն պարզ է, որովհետև ինձ համար մի սեռի տեսակները, անշուշտ, նույնպես ծագում են մեկ ընդհանուր նախահորից, ինչպես որ միևնույն տեսակի երկու սեռերը։ Ուրեմն՝ եթե ընդհանուր նախահոր կամ նրա ամենամերձավոր հաջորդների կազմվածքի որևէ մասը փոփոխական է դարձել, ապա շատ հավանական է, որ նրա փոփոխությունները աջակցություն են ստացել բնական ու սեռական ընտրությունների կողմից, որպեսզի հարմարեցնեն զանազան տեսակները համապատասխան տեղերին բնական տնտեսության մեջ, կամ երկու սեռերը մեկը մյուսին, կամ, վերջապես, արուներին այն պայքարին, որ նրանք մղում են միմյանց հետ էգերին տիրանալու համար։
Վերջում ես գալիս եմ հետևյալ ընդհանուր եզրակացության, փոփոխականության ամենամեծ աստիճանը տեսակային հատկանիշների համար (այսինքն այն հատկանիշների, որոնք մեզ հնարավորություն են տալիս տարբերելու տեսակը տեսակից), քան սեռային հատկանիշների (այսինքն այն հատկանիշների, որոնք ընդհանուր են մի սեռի պատկանող տեսակներին). հաճախ արտահայտվող խոր փոփոխականություն որևէ մասի, որը խիստ զարգացած է որոշ տեսակի մոտ մերձավոր տեսակների համեմատությամբ, կամ թույլ փոփոխականություն այն մասի, որը թեև ուժեղ կերպով, բայց միանման չափով զարգացած է մի ամբողջ խմբի տեսակների մոտ. երկրորդական սեռական հատկանիշների մեծ փոփոխականությունը և նրանց զգալի տարբերությունը միմյանց մոտիկ տեսակների մեջ. վերջապես, երկրորդական սեռական հատկանիշների ու սովորաբար կազմվածքի միևնույն մասերին վերաբերող սովորական տեսակային հատկանիշների զուգադիպումը միմյանց, այս բոլորը սկզբունքներ են, որոնք փոխադարձ սերտ կապի մեջ են գտնվում։ Նրանք բոլորը հիմնականում կախված են նրանից, որ մի խմբի տեսակները առաջացել են մեկ ընդհանուր նախահորից, որը նրանց փոխանցել է ընդհանուր շատ առանձնահատկություններ, նրանից, որ ոչ վաղուց և խոր կերպով փոփոխված մասերը հավանորեն կշարունակեն ավելի շատ փոփոխվել, քան վաղուց ժառանգված ու չփոփոխված մասերը, նրանից, որ բնական ընտրությունը անցած ժամանակամիջոցի համեմատ կկարողանա մեծ կամ փոքր չափով հաղթահարել վերադարձի կամ հետագա փոփոխականության ձգտումները, նրանից, որ սեռական ընտրությունը այնքան խիստ չի ներգործում, ինչպես սովորական ընտրությունը, և վերջապես նրանից, որ նույն մասերի փոփոխությունները, որոնք կուտակվել են բնական ու սեռական ընտրության ներգործությամբ, հարմարված են եղել օրգանիզմների թե՛ երկրորդական սեռական և թե՛ սովորական պահանջներին։
ՄԻՄՅԱՆՑԻՑ ՏԱՐԲԵՐ ՏԵՍԱԿՆԵՐԸ ՈՒՆԵՆՈՒՄ ԵՆ ԱՆԱԼՈԳ ՓՈՓՈԽՈՒԹՅՈՒՆՆԵՐ, ԱՅՆՊԵՍ ՈՐ ՈՐՈՇ ՏԵՍԱԿԻ ՄԻ ՏԱՐԱՏԵՍԱԿ ՀԱՃԱԽ ՁԵՌՔ է ԲԵՐՈՒՄ ԱԶԳԱԿԻՑ ՄԻ ՏԵՍԱԿԻՆ ՀԱՏՈՒԿ ԱՌԱՆՁՆԱՀԱՏԿՈՒԹՅՈՒՆՆԵՐ, ԿԱՄ ՎԵՐԱԴԱՌՆՈՒՄ է ԻՐ ՄԻ ԱՎԵԼԻ ՎԱՂ ԺԱՄԱՆԱԿԻ ՆԱԽԱՀՈՐ ՀԱՏԿԱՆԻՇՆԵՐԻՆ
Այս դրույթները մեզ իսկույն հասկանալի կդառնան, եթե մենք դրանք կիրառենք մեր ընտանի ցեղերի նկատմամբ։ Մեկը մյուսից հեռու գտնվող երկրներում աղավնիների ամենատարբեր ցեղերը ներկայացնում են այնպիսի ենթացեղեր, որոնք ունենում են գլխի վրա խճճված փետուրներ, փետրածածկ ոտքեր, այսինքն հատկանիշներ, որ հատուկ չեն իրենց տեղական նախահորը, ժայռաղավնուն, հետևաբար դրանք անալոգ փոփոխություններ են երկու կամ ավելի թվով ցեղերի մեջ։ Փքաղավնու պոչի հաճախ տասնչորս կամ նույնիսկ տասնվեց փետուրներ ունենալը կարելի է դիտել որպես մի փոփոխություն, որը համապատասխանում է մի այլ ցեղի՝ սիրամարգաաղվանու նորմալ ձևին։ Ոչ ոք, ես կարծում եմ, չի կասկածում, որ նման բոլոր անալոգ փոփոխություններն իրենց ծագումով պարտական են այն հակումին, որ աղավնիների զանազան ցեղերը ժառանգել են մի ընդհանուր նախահորից, այն է՝ նման, մեզ համար անհայտ ազդեցությունների ներգործությամբ փոփոխվելու հակումը։ Բույսերի մեջ անալոգ փոփոխության մի դեպք են ներկայացնում, սովորաբար, արմատներ կոչվող հաստացած ցողունները շվեդական գոնգեղի և սովորական կերաշաղգամի մոտ, որոնք մի քանի բուսաբանների կողմից համարվում են մի ընդհանուր նախահորից առաջացած կուլտուրական տարատեսակներ. բայց եթե այդ ենթադրությունը ճիշտ չէ, ապա մենք այստեղ ևս կունենանք երկու, այսպես կոչված, առանձին տեսակների անալոգ փոփոխության դեպք, իսկ դրանց կարելի է միացնել նաև երրորդը, այն է՝ սովորական շաղգամը։ Տեսակների մասին տարածված այն տեսակետի հիման վրա, որ նրանք առանձին ստեղծագործական ակտերի արդյունքներ են, մենք չենք կարող ընդունել, որ այդ երեք բույսերի ցողունների միանման հաստացման vera causa-ն[46] գտնվում է նրանց ընդհանուր ծագման և այստեղից բխող միանման փոփոխության ձգտելու մեջ, այլ պետք է ընդունենք երեք ինքնուրույն, թեև մեկը մյուսին շատ մոտիկ ստեղծագործական ակտեր։ Անալոգ փոփոխությունների նման շատ դեպքեր նկատել է Նոդենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 19] դդմազգիների ընտանիքում և զանազան հեղինակներ՝ մեր հացազգիների մեջ։ Նման օրինակներ հանդիպում են նաև միջատների մեջ բնական վիճակում և վերջերս մեծ հմտությամբ մշակվել են միստր Ուոլշի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 103] կողմից, որը խմբավորել է այդ երևույթները «համազոր փոփոխականության» (equable variability) օրենքի անվան տակ։
Բայց աղավնիների մեջ մենք հանդիպում ենք նաև մի այլ դեպքի, այն է՝ աղավնիների բոլոր ցեղերի մեջ պատահական երևան են գալիս գորշակապույտ (թխագորշ) գույնով, թևերի վրա երկու սև լայնակի շերտեր ունեցող, սպիտակ փորով, պոչածայրին սպիտակ շերտով, հիմքի մոտ սպիտակ եզրապատում ունեցող արտաքին փետուրներով անհատներ։ Քանի որ այդ բոլոր հատկանիշները բնորոշ են նախահայր ժայռաղավնու համար, ուստի ես կարծում եմ, որ ոչ ոք չի կասկածի, որ այստեղ մեր առջև ունենք վերադարձի դեպք, այլ ոչ թե նոր և անալոգ փոփոխություն, որն առաջանում է զանազան ցեղերի մեջ։ Մենք կարող ենք վստահությամբ ընդունել այդ եզրակացությունը, որովհետև այդ գունավոր նշանները առանձնապես հեշտությամբ են դրսևորվում երկու տարբեր և զանազանագույն ցեղերի խաչաձևման ժամանակ, իսկ այդ դեպքերում ոչինչ չի փոխվում արտաքին կենսապայմաններում, բացի խաչաձևման ակտից, որն ազդում է ժառանգականության օրենքի վրա։
Անտարակույս զարմանալի է այն փաստը, որ շատ, ամենայն հավանականությամբ, հարյուրավոր սերունդներ առաջ անհետացած հատկանիշները երևան են գալիս։ Բայց երբ որևէ ցեղ խաչաձևում են մի այլ ցեղի հետ, ապա ստացված խառնացեղերը պահպանում են այդ օտար ցեղի հատկանիշներին վերադառնալու ձգտումը,— ոմանք պնդում են, թե մինչև տասներկուերորդ և նույնիսկ մինչև քսաներորդ սերունդը։ Տասներկու սերունդներից հետո օտար նախորդի արյան բաժինը, եթե թույլատրելի է գործածել այդ սովորական արտահայտությունը, կլինի 1/2048, և այնուամենայնիվ այդ օտար արյան մնացորդը, ըստ տարածված կարծիքի, արտահայտվում է վերադարձի ձգտմամբ։ Այն ցեղը, որ երբեք չի ենթարկվել խաչաձևման, բայց որի մեջ երկու ծնողներն էլ կորցրել են իրենց նախնիքի հատկանիշներից որևէ մեկը, կորցրած հատկանիշը վերարտադրելու այդ ընդունակությունը թույլ կամ ուժեղ չափով կարող է, ինչպես մենք ասել ենք վերևում, պահպանվել անորոշ քանակությամբ սերունդներում, թեև մենք սովորաբար տեսնում ենք հակառակը։ Երբ մի որևէ հատկանիշ, որը որոշ ցեղի մեջ անհետացել էր, սերունդների երկար շարքից հետո նորից է երևում, ապա ամենից ավելի հավանական է ոչ թե այն պատկերացումը, որ իբր թե մի անհատ հանկարծակի կերպով իրենից հարյուրավոր սերունդներ առաջ եղած նախորդին է քաշել, այլ այն ենթադրությունը, որ յուրաքանչյուր հաջորդ սերնդում այդ հատկանիշը պահվում է թաքնված վիճակում և միայն մեզ անհայտ բարենպաստ պայմանների շնորհիվ վերջապես հայտնվել է։ Այսպես, շատ հազվագյուտ դեպքերում կապտագույն անհատներ առաջացնող հնդկական աղավնու (Barb), շատ հնարավոր է, յուրաքանչյուր սերնդում գոյություն ունի կապտագույն փետրավորում առաջացնելու այդ թաքնված ընդունակությունը։ Վերացական տեսակետից դատելով, սերունդների երկար շարքի ընթացքում այդպիսի ձգտման փոխանցման անհավանականությունը ավելի մեծ չէ, քան բոլորովին անօգուտ ու այլասերված թերաճ օրգանների նույնպիսի փոխանցումը։ Իսկ նման թերաճ օրգաններ առաջացնելու ձգտումը հաճախ ժառանգական է լինում։
Քանի որ ենթադրվում է, որ միևնույն սեռի բոլոր տեսակները առաջացել են մի ընդհանուր նախահորից, ապա բնական է սպասել, որ նրանք երբեմն կփոփոխվեն համանման ձևով, այնպես որ երկու կամ մեծ թվով տեսակների տարատեսակները նման կլինեն մեկը մյուսին, կամ մի տեսակի տարատեսակ իր որոշ հատկանիշներով կնմանվի մի այլ տեսակի, որն ինքը, մեր տեսակետի հիման վրա, միայն ավելի խիստ արտահայտված ու հաստատուն տարատեսակ է։ Սակայն բացառապես համանման փոփոխությունների շնորհիվ առաջացած հատկանիշները, ամենայն հավանականությամբ էական չեն լինի, որովհետև օրգանիզմի աշխատանքների համար կարևոր բոլոր հատկանիշները կպահպանվեն բնական ընտրությամբ տեսակների տարբեր կենցաղին համապատասխան կերպով։ Վերջապես, այնուհետև կարելի է սպասել, որ միևնույն սեռի տեսակները երբեմն կդրսևորեն վերադարձ դեպի վաղուց կորցրած հատկանիշները։ Բայց քանի որ մեզ անհայտ են մեր բնական խմբերի ընդհանուր նախնիները, ուստի մենք ի վիճակի չենք տարբերելու անալոգ հատկանիշները վերադարձի հատկանիշներից։ Եթե մենք, օրինակ, չգիտենայինք, որ մեր աղավնացեղերի նախահայր ժայռաղավնին չի ունեցել ոչ փետրածածկ ոտներ, ոչ էլ փուփուլավոր գլուխ, մենք չէինք կարողանա ասել, թե մեր ընտանի ցեղերի այդ հատկանիշներն արդյոք անալոգ են, թե վարդարձային։ Մենք գուցե կընդունենք թխակապույտ գույնը որպես վերադարձի դեպք, որովհետև այդ գույնի հետ կապված նշանների մեծ թիվը անհավանական է դարձնում նրանց բոլորի միասին երևան գալը պարզ փոփոխակման շնորհիվ։ Առավել ևս մեզ կբերեր այդ եզրակացության այն փաստը, որ թխակապույտ գույնը և նշանները շատ հաճախ են երևում, երբ խաչաձևվում են ամենաբազմազան գույն ունեցող ցեղերը։ Հետևաբար, թեև բնական վիճակում սովորաբար կասկածի տակ կմնա, թե արդյոք մենք մեր առջև ունե՞նք նախկինում գոյություն ունեցող հատկանիշներին վերադառնալու դեպք, թե նոր անալոգ փոփոխության դեպք, այնուամենայնիվ, մեր տեսության հիման վրա մենք պետք է սպասենք, որ ժամանակ առ ժամանակ երևան կգան փոփոխություններ, որոնք կմոտեցնեն որևէ տեսակի հատկանիշները նույն խմբի այլ տեսակների հատկանիշներին։ Եվ այդ անկասկած այդպես է։
Փոփոխական տեսակները միմյանցից տարբերելը դժվար է լինում մեծ չափով այն պատճառով, որ տարատեսակները, այսպես ասած, ընդօրինակում են միևնույն սեռի այլ տեսակներին։ Կարելի է բերել նաև այն ձևերի երկար ցուցակը, որոնք միջին տեղ են բռնում երկու ձևերի միջև, որոնց կարելի էր միայն կասկածանքով բարձրացնել տեսակների աստիճանի, իսկ այդ ապացուցում է,— եթե միայն այդ միմյանց մերձավոր ձևերը չընդունենք որպես առանձին ստեղծագործական ակտերի արդյունք,— նրանք ձեռք են բերել մյուս ձևերի հատկանիշներին նման հատկանիշներ։ Անալոգ փոփոխությունների լավագույն ապացույց են ծառայում այն մասերը կամ օրգանները, որոնք սովորաբար աչքի են ընկնում իրենց հատկանիշների կայունությամբ, բայց երբեմն այնպես են փոփոխվում, որ սկսում են որոշ չափով նմանվել հարևան տեսակի համապատասխան մասին կամ օրգանին։ Ես այդպիսի դեպքերի մի երկար ցուցակ եմ հավաքել. բայց այստեղ, ինչպես որ առաջ, ես գտնվելով աննպաստ պայմաններում, ի վիճակի չեմ այն բերելու։ Կարող եմ միայն կրկնել, որ այդպիսի դեպքեր իրոք գոյություն ունեն և ինձ չափազանց զարմանայի են թվում։
Սակայն ես կբերեմ մի հետաքրքիր և բարդ դեպք, ոչ այն պատճառով, որ նա վերաբերում է մի կարևոր հատկանիշի, այլ այն պատճառով, որ նա ընդգրկում է մի քանի տեսակներ, որոնք մասամբ գտնվում են բնական վիճակում, մասամբ էլ ընտանեցված են։ Դա համարյա հաստատապես վերադարձի դեպք է։ Էշի ոտքերի վրա երբեմն լինում են լայնական պարզորոշ զոլեր, ինչպես զեբրի ոտքերի վրա. պնդում են, որ նրանք ավելի պարզ կերպով երևում են քուռակի վրա, և իմ հավաքած տեղեկությունների հիման վրա, ես կարծում եմ, որ այդ ճիշտ է։ Ուսի վրա զոլը երբեմն կրկնակի է և խիստ տատանվում է ինչպես երկարությամբ, այնպես էլ ձևով։ Հայտնի է մի դեպք, որ սպիտակ, բայց ոչ ալբինիոս էշը ոչ ուսի, ոչ էլ մեջքի զոլեր չի ունեցել, իսկ մուգ թույր ունեցող էշերի վրա այդ զոլերը երբեմն աննշմարելի են կամ բոլորովին բացակայում են։ Պալլասի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 104] կուլանի վրա պատահել է կրկնակի ուսազոլ։ Միստեր Բլիտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 39] տեսել է մի ջիգետայ (hemionus), որն ունեցել է ուսի վրա պարզորոշ զոլ, թեև տիպականորեն ջիգետայը չի ունենում այն. իսկ գնդապետ Պուլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 105] հաղորդել է ինձ, որ այդ տեսակի քուռակները սովորաբար ունենում են զոլավոր ոտներ և ոչ պարզորոշ ուսազոլ։ Կվագգան, որի մարմինը զեբրին բոլորովին նման ծածկված է զոլերով, իր ոտների վրա զոլեր չունի, բայց դոկտոր Գրեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 106] նկարագրել է մի օրինակ, որ ունեցել է զեբրին հիշեցնող պարզ զոլեր։
Ինչ վերաբերում է ձիուն, ապա ես Անգլիայում հավաքել եմ ամենաբազմազան ցեղերի և բոլոր թույրերի ձիերի մեջքի զոլին վերաբերող փաստեր, ոտների վրա լայնակի զոլերը հազվադեպ չեն բաց աշխետ, մկնագույն աշխետ ձիերի վրա և մի դեպքում նկատվել են մուգ շագանակագույն ձիու վրա։ Թույլ ուսազոլը երբեմն կարելի է տեսնել բաց աշխետների վրա, և ես մի անգամ նրա հետքերը տեսել եմ դեղնակարմիր ձիու վրա։ Իմ որդին մանրազնին կերպով զննել և նկարել է ինձ համար մի բելգիական բաց աշխետ ծանրաքաշ ձի, որն ուներ կրկնակի ուսազոլ և ոտների վրա զոլեր։ Ես ինքս տեսել եմ դեվոնշիրյան բաց աշխետ մի պոնի, իսկ մի ուրիշ փոքրիկ էլի բաց աշխետ վալիսյան պոնի գիտեմ ըստ մանրամասն նկարագրությունների, երկուսն էլ ունեին երեքական զուգահեռ զոլեր յուրաքանչյուր ուսի վրա։
Հյուսիս-արևմայան Հնդկաստանում կատտիվարյան ցեղի ձիերը այնքան միշտ լինում են զոլավոր, որ ինչպես ինձ հաղորդել է գնդապետ Պուլը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 105] որն ուսումնասիրել է այդ ցեղը հնդկական կառավարության հանձնարարությամբ, այդ զոլերը չունեցող ձին զտարյուն չի համարվում։ Մեջքի վրա միշտ գտնվում է մի զոլ, ոտները սովորաբար զոլավոր են, իսկ ուսի զոլերը, թվով երկու կամ երեք հատ, նույնպես հազվադեպ չեն, բացի դրանից, զոլեր կան նաև այտերի վրա։ Հաճախ զոլերը ավելի նկատելի են մտրուկի վրա, իսկ ծեր ձիերի վրա երբեմն բոլորովին անհետանում են։ Գնդապետ Պուլը նկատել է, որ կատտիվարյան թե՛ գորշ և թե՛ դեղնակարմիր զամբիկները առաջին անգամ ծնելիս բերում էին զոլավոր մտրուկներ։ Միստեր Էդվարդսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 86] ինձ հաղորդած տեղեկությունների հիման վրա ես առիթ ունեմ ենթադրելու, որ անգլիական արշավաձիու մեջքի զոլը ավելի սովորական է մտրուկի վրա, քան չափահաս ձիու վրա։ Ես ինքս նույնպես վերջերս ստացել եմ մի մտրուկ դեղնակարմիր զամբիկից (որն առաջացել է թուրքմենական հովատակից և ֆլամանդական զամբիկից) և դեղնակարմիր անգլիական արշավաձիու հովատակից, երբ այդ մտրուկը մոտավորապես մեկ շաբաթական էր, նա ուներ իրանի հետևի մասի և ճակատի վրա բազմաթիվ նեղ մգագույն ղողեր, ինչպես զեբրինն է, և երևան էր բերում ոտների թույլ զոլավորություն. բայց շուտով այդ բոլոր զոլերն անհետացան։ Չմտնելով հետագա մանրամասնությունների մեջ, միայն կասեմ, որ ես ուսերի և ոտների վրա եղած զոլերի մասին փաստեր հավաքել եմ ամենաբազմազան ցեղերի վերաբերյալ և զանազան երկրներում՝ սկսած Բրիտանիայից մինչև Արևելյան Չինաստան և հյուսիսում գտնվող Նորվեգիայից մինչև Մալայան արշիպելագը հարավում։ Բոլոր երկրներում այդ զոլերը ամենից ավելի հաճախ հանդիպում են բաց աշխետ, մկնագույն աշխետ ձիերի վրա. այդ աշխետ տերմինի տակ հասկացվում է թույրերի բավական լայնածավալություն սկսած սևագորշից մինչև սերուցքի գույնը։
Ես շատ լավ գիտեմ, որ գնդապետ Համիլտոն Սմիտը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 107] որը գրել է այդ հարցի մասին, կարծում է, թե ձիերի զանազան ցեղերը առաջացել են մի քանի նախնական տեսակներից, որոնցից մեկը՝ բաց աշխետը եղել է զոլավոր և որ նկարագրված առանձնահատկություններն առաջանում են այդ բաց աշխետ ցեղի հետ կատարված հին խաչաձևումներից։ Բայց այդ տեսակետը կարելի է համարձակորեն ժխտել, որովհետև վերին աստիճանի անհավանական է, որ բելգիական ծանրաքաշը (բեռնակիրը), վալլիսյան պոնին, նորվեգիական ձին, նիհարակազմ կատիվարյան ցեղը, որոնք ապրում են աշխարհիս ամենահեռավոր ծայրերում, բոլորը խաչաձևված լինեն մի ենթադրական նախնական ցեղի հետ։
Այժմ դառնանք ձիու սեռի տարբեր տեսակների խաչաձևման արդյունքներին։ Ռոլլենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 108] պնդում է, որ էշից ու ձիուց ստացված խառնացեղ սովորական ջորին առանձնապես հաճախ երևան է բերում ոտների վրա զոլեր. ըստ միստեր Գոսսեյի՝[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 109] Միացյալ Նահանգների որոշ մասերում տասը ջորուց ինը ունեն զոլավոր ոտներ։ Ես մի անգամ տեսել եմ մի ջորի, որի ոտներն այնպես զոլավոր էին, որ նրան կարելի էր ընդունել որպես զեբրի խառնացեղ, իսկ Վ. Մարտինը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 110] ձիու մասին գրած իր հիանալի աշխատության մեջբերում է նման մի ջորու պատկերը։ Էշից և զեբրից ստացված հիբրիդների գունազարդ չորս նկարների վրա, որ ես տեսել եմ, ոտները այնքան էլ խիստ զոլավոր չէին, ինչպես մարմնի մնացած մասերը, իսկ նրանցից մեկի վրա կար կրկնակի ուսազոլ։ Լորդ Մորտոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 111] նշանավոր հիբրիդը, որն ստացվել էր մուգ շագանակագույն զամբիկից և կվագայի հովատակից, ինչպես նաև դրանից հետո նույն զամբիկից և արաբական սև հովատակից ստացված զտարյուն քուռակը, ունեցել են ոտների վրա ավելի ևս ցայտուն զոլեր, քան նույնիսկ զտարյուն կվագան։ Վերջապես, այս էլ չափազանց ուշագրավ դեպք է, դոկտոր Գրեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 106] տալիս է էշից ու ջիգետայից ստացված հիբրիդի պատկերը (իսկ ինձ նա հաղորդում է, որ իրեն հայտնի է նաև մի երկրորդ այդպիսի դեպք)։ Չնայած այն բանին, որ էշը ոտների վրա երբեմն է ունենում զոլեր, իսկ ջիգետայը երբեք չի ունենում, և նույնիսկ ոչ մի հատ ուսազոլ չի ունենում, այդ հիբրիդի չորս ոտներն էլ եղել են զոլավոր, իսկ ուսերի վրա երեք կարճ զոլեր, ինչպես դեվոնշիրյան և վալլիսյան բաց աշխետ պոնիների վրա, և նույնիսկ ուներ մի քանի զոլեր այտերի վրա, ինչպես զեբրինն է։ Այս վերջին փաստի առթիվ ես համոզված էի այն բանում, որ ոչ մի նույնիսկ գույնի զոլ չի կարող, այսպես ասած, պատահականության արդյունք լինել, որ էշի և ջիգետայի այդ հիբրիդի այտերի զոլերի հիման վրա ես հարցումով դիմել եմ գնդապետ Պուլին,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 105] ցանկանալով իմանալ, թե արդյոք նրանք լինո՞ւմ են նաև ձիերի կատտիվարյան ցեղի վրա, որն այնքան աչքի է ընկնում էր զոլավորությամբ, և ստացել եմ, ինչպես արդեն տեսանք, դրական պատասխան։
Ի՞նչ կասենք մենք այդ զանազան փաստերի մասին։ Մենք տեսնում ենք ձիու սեռի մի քանի ինքնուրույն տեսակներ, որոնց ոտները պարզ փոփոխականության միջոցով դառնում են զոլավոր, ինչպես որ զեբրինն է, կամ զոլերը երևան են գալիս ուսերի վրա, ինչպես որ էշինն է։ Ձիու մոտ այդ հակումն առանձնապես ուժեղ կերպով արտահայտված ենք տեսնում, երբ նրա թույրը մոտենում է բաց աշխետին՝ այսինքն նրան, որն ավելի մոտ նմանություն ունի այդ սեռի մյուս տեսակների թույրին։ Զոլերի երևան գալը չի ուղեկցվում ձևի կամ որևէ այլ հատկանիշների ոչ մի փոփոխություններով։ Զոլավորություն արտահայտելու այդ հակումը առանձնապես ուժեղ կերպով է դրսևորվում, երբ միմյանցից ամենաշատ տարբերվող տեսակներ են խաչաձևվում։ Հիշենք աղավնիների զանազան ցեղերի օրինակը։ Նրանք առաջացել են մեկ աղավնուց (ներառնելով այստեղ երկու կամ երեք ենթատեսակ կամ աշխարհագրական ռասաներ), որն ունի թխակապույտ գույն՝ որոշ զոլերով ու նշաններով և ամեն անգամ, երբ որևէ ցեղ փոփոխականության հետևանքով ձեռք է բերում թխակապույտ գույն, նրա հետ միասին միշտ երևան են գալիս նաև այդ զոլերն ու նշանները, բայց առանց միաժամանակ ձևի կամ այլ հատկանիշների որևէ փոփոխության։ Երբ զանազան գույն ունեցող ամենահին և կայուն ցեղերը խաչաձևում են միմյանց հետ, այդ դեպքում հիբրիդները ցուցաբերում են ուժեղ հակում դեպի թխակապույտ գույնը, զոլերը և նշանները։ Ես կարծիքս արդեն հայտնել եմ, որ ամենից հավանական է թվում այն հիպոթեզը, որը բացատրում է հին հատկանիշների երևալը նրանով, որ յուրաքանչյուր սերունդ իր մատաղ հասակում հակում ունի վերարտադրելու այդ վաղուց կորսված հատկանիշները՝ մի հակում, որը անհայտ պատճառներով երբեմն հաղթում է։ Իսկ այժմ մենք տեսնում ենք, որ ձիու սեռի մի քանի տեսակների մեջ զոլերը և ավելի պարզորոշ և ավելի հաճախ են երևում մատղաշ կենդանիների վրա, քան ծերերի վրա։ Անվանեցեք աղավնիների ցեղերը, որոնք որոշ դեպքերում իրենց կայունությունը պահպանել են դարերի ընթացքում, անվանեցեք նրանց տեսակներ, և այդ դեպքը արդյոք չի՞ ներկայացնի մեզ զուգահեռություն այն երևույթի հետ, ինչ որ մենք տեսնում ենք ձիու սեռի տեսակների մեջ։ Ինչ վերաբերում է ինձ, ապա ես համոզվածությամբ եմ նայում հեռու անցյալին, որից մեզ բաժանում են հազարավոր ու հազարավոր սերունդներ, և տեսնում եմ այնտեղ զեբրի նման զոլավոր կենդանուն, բայց որը այլ տեսակետներից թերևս բոլորովին այլ կերպ էր կառուցված և եղել է մեր ընտանի ձիու (միևնույնն է, թե արդյոք նա առաջացել է մեկ կամ մի քանի վայրենի ձևերից), ջիգետայի, կվագայի և զեբրի ընդհանուր նախահայրը։
Բայց նա, ով հավատում է, թե ձիու յուրաքանչյուր տեսակ ստեղծվել է մնացածներից անկախ կերպով, հավանորեն կպնդի, որ յուրաքանչյուր տեսակ ստեղծվել է բնական կամ ընտանեցված վիճակում մի որոշակի ուղղությամբ այնպես փոփոխվելու հակումով, որպեսզի այդ սեռի մյուս տեսակների նման դառնա զոլավոր և այնուհետև կպնդի, որ յուրաքանչյուր տեսակ ստեղծվել է ուժեղ ձգտումով՝ աշխարհի այլ ծայրում ապրող տեսակների հետ խաչաձևվելու ժամանակ տալ հիբրիդներ, որոնք իրենց զորավորությամբ նմանվում են ոչ թե իրենց ծնողներին, այլ նույն սեռի այլ տեսակներին։ Ընդունել այդ տեսակետը՝ կնշանակեր հրաժարվել իսկական պատճառից հանուն երևակայական, կամ առնվազն անհայտ պատճառի։ Դա կնշանակեր աստծո արարածներին համարել ծաղր ու խաբեություն, ես նույնպիսի հեշտությամբ կհամաձայնեի հավատալ դրան, ինչպես հին տիեզերածնությունների տգետ ստեղծողներին, որոնք պնդում էին, թե բրածո խեցիները երբեք կենդանի չեն եղել, այլ միայն այդպես ստեղծվել են քարից, որպեսզի ընդօրինակեն ծովափին ապրող խեցիներին։
ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ
Մեր անգիտակությունը փոփոխականության օրենքների մասին խորն են։ Հարյուր դեպքից ոչ մեկում մենք ի վիճակի չենք ցույց տալու այն պատճառը, թե ինչու կազմվածքի այս կամ այն մասը փոփոխվել է։ Բայց բոլոր դեպքերում, որտեղ հնարավոր է լինում համեմատել, մենք համոզվում ենք, որ միևնույն օրենքներն են ղեկավարում տարատեսակների միջև եղած ավելի քիչ խոր տարբերությունները և տեսակների միջև եղած ավելի խոր տարբերությունները։ Պայմանների փոփոխությունը սովորաբար հարուցում է տատանվող փոփոխականություն, սակայն երբեմն ուղեկցվում է նաև ուղղակի, որոշակի հետևանքներով, և այդ հետևանքները ժամանակի ընթացքում կարող են ավելի ուժեղ կերպով նկատվել, թեև այդ ենթադրության օգտին բավարար վկայություն դեռ չկա։ Սովորությունը, որն առաջացնում է ընդհանուր կառուցվածքի որոշ առանձնահատկություններ, ինչպես նաև օրգաններն ուժեղացնող վարժեցումը, թուլացնող ու փոքրացնող ոչ վարժեցումը, ըստ երևույթին շատ դեպքերում ցուցաբերել են իրենց ներգործությունը։ Հոմոլոգ մասերը դրսևորում են միանման փոփոխվելու, ինչպես նաև միաձուլվելու ձգտում։ Պինդ և արտաքին մասերի փոփոխությունները իրենց ազդեցությունն են գործում փափուկ և ներքին մասերի վրա։ Երբ մի որևէ մաս անչափ է զարգանում, այդ դեպքում նա կարող է իրեն հարակից մասերից քաշել սննդանյութեր, և ամեն մի մաս, որը կարող է կյանքի համար առանց վնասի հեռացվել, կվերացվի։ Կազմվածքի մեջ վաղ հասակում երևան եկող փոփոխությունները կարող են անդրադառնալ այն մասերի վրա, որոնք զարգանում են ավելի ուշ, ինչպես նաև հանդիպում են հարաբերակցական փոփոխականության դեպքեր, որոնց պատճառը մեզ համար պարզ չէ։ Միևնույն օրգանիզմի մեջ բազմակի կրկնվող մասերը փոփոխական են ինչպես իրենց թվով, այնպես էլ կառուցվածքով, և դա գուցե բխում է նրանից, որ նրանք, ճշգրտորեն մասնագիտացած չլինելով իրենց աշխատանքներում, բնական ընտրության խիստ վերահսկողության չեն ենթարկվել։ Նույն պատճառի հետևանքն է թերևս նաև այն փաստը, որ էակների շարքում ավելի ցածր աստիճանի վրա կանգնածներն ավելի փոփոխական են, քան ավելի բարձր կանգնած էակները, որոնց կազմվածքն ավելի մասնագիտացված է։ Թերաճ օրգանները, անօգուտ լինելով, բնական ընտրության ներգործության չեն ենթարկվում և այդ պատճառով փոփոխական են։ Տեսակային հատկանիշները, այսինքն տարբերության այնպիսի գծերը, որոնցով տեսակներն սկսել են տարբերվել միմյանցից այն ժամանակից սկսած, երբ նրանք ճյուղավորվել են իրենց ընդհանուր նախահորից, ավելի փոփոխական են, քան սեռային կամ այնպիսի հատկանիշները, որոնք վաղուց ի վեր ժառանգված են և չեն փոփոխվել այդ ժամանակաշրջանի ընթացքում։ Այդ դիտողություններում մենք շոշափեցինք միայն օրգաններն ու առանձին մասերը, մատնանշելով, որ նրանք պահպանել են փոփոխականության ձգտումն այն պատճառով, որ դեռ նոր են փոփոխվել ու զանազանակերպվել, բայց մենք երկրորդ գլխում տեսանք, որ նույն կշռադատումները ճիշտ են նաև ամբողջությամբ վերցրած անհատի վերաբերմամբ։ Մենք համոզվեցինք, որ տվյալ սեռին պատկանող շատ տեսակներ պարփակող մարզում, այսինքն այնտեղ, որտեղ վերջերս են տեղի ունեցել զգալի փոփոխություն և դիֆերենցում, կամ ինչպես մենք արտահայտվել ենք, որտեղ տեսակների ֆաբրիկացիան գործուն է եղել, այնտեղ ևս այդ տեսակների մեջ մենք հանդիպում ենք, միջին հաշվով, տարատեսակների ամենամեծ առատության։ Երկրորդական սեռական հատկանիշները շատ փոփոխական են. նրանք ամենից ավելի փոփոխական են միևնույն սեռի տեսակների մեջ։ Կազմվածքի միևնույն մասերում փոփոխականությունը նյութ է ծառայել ինչպես երկրորդական սեռական հատկանիշների համար, այնպես էլ միևնույն սեռի տեսակների միջև տեսակային տարբերությունների համար։ Ամեն մի մաս կամ օրգան, որը զգալի մեծության է հասել կամ զարգացել է արտասովոր ձևով՝ համեմատած ազգակից տեսակների միևնույն օրգանների կամ մասերի հետ, պետք է ուժեղ կերպով փոփոխվեր այդ սեռի ծագման ժամանակից սկսած. այստեղից մեզ հասկանալի է, թե ինչու նրանք դեռ պահպանել են փոփոխվելու ընդունակությունն անհամեմատ ավելի մեծ չափով, քան մյուս մասերը, որովհետև փոփոխականությունը ներկայացնում է դանդաղ ու երկարատև պրոցես և բնական ընտրությունը դեռ չի կարողացել հաղթահարել հետագա փոփոխականության և ավելի քիչ փոփոխված վիճակի վերադառնալու ձգտումները։ Բայց այն դեպքում, երբ արտասովոր զարգացած օրգան ունեցող տեսակը կարողացել է սկիզբ տալ բազմաթիվ փոփոխված հետնորդների, որը, մեր կարծիքով, պետք է կատարվեր ծայրահեղ դանդաղությամբ, երկար ժամանակամիջոցների ընթացքում, այդ դեպքում բնական ընտրությունն արդեն կարողացել է այդ օրգանին կամ հատկանիշին տալ կայունության դրոշմ՝ չնայած նրա արտասովոր զարգացմանը։ Այն տեսակները, որոնք իրենց ընդհանուր նախնիներից ժառանգել են գրեթե միանման ընդհանուր կերտվածք և ենթարկված են միանման պայմանների, բնականաբար ձգտում են համանման փոփոխությունների կամ երբեմն վերադառնում են իրենց հեռավոր նախնիների հատկանիշներին։ Այսպիսով, թեև վերադարձի, կամ անալոգ փոփոխականության շնորհիվ նոր և կարևոր փոփոխություններ չեն առաջանա, այնուամենայնիվ նրանք կնպաստեն բնության հիանալի ու ներդաշնակ բազմազանությանը։
Ինչ էլ որ լինի ծնողների և նրանց հաջորդների միջև եղած յուրաքանչյուր չնչին տարբերության պատճառը,— իսկ այդպիսի պատճառ պետք, է միշտ գոյություն ունենա,— մենք լիակատար հիմք ունենք կարծելու, որ հենց բարենպաստ տարբերությունների անշեղ կուտակումն է ծնել կազմվածքի այն բոլոր էական փոփոխությունները, որոնք կապված են յուրաքանչյուր տեսակի կենսակերպի հետ։
ԳԼՈՒԽ VI։ ԴԺՎԱՐՈՒԹՅՈՒՆՆԵՐ, ՈՐՈՆՑ ՀԱՆԴԻՊՈՒՄ Է ՏԵՍՈՒԹՅՈՒՆԸ
ԴԺՎԱՐՈՒԹՅՈՒՆՆԵՐ, ՈՐՈՆՑ ՀԱՆԴԻՊՈՒՄ է ՓՈՓՈԽՈՒԹՅԱՆ ՄԻՋՈՑՈՎ. ՏԵՍԱԿՆԵՐԻ ԾԱԳՄԱՆ ՏԵՍՈՒԹՅՈՒՆԸ։— ԱՆՑՈՒՄՆԱՅԻՆ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԲԱՑԱԿԱՅՈՒԹՅՈՒՆԸ ԿԱՄ ՀԱԶՎԱԴԵՊ ԼԻՆԵԼԸ։— ԱՆՑՈՒՄՆԵՐ ԿԵՆՍԱԿԵՐՊԻ ՄԵՋ։— ՍՈՎՈՐՈՒԹՅՈՒՆՆԵՐԻ ԲԱԶՄԱԶԱՆՈՒԹՅՈՒՆԸ ՄԻԵՎՆՈՒՅՆ ՏԵՍԱԿԻ ՍԱՀՄԱՆՆԵՐՈՒՄ։— ՏԵՍԱԿՆԵՐ, ՈՐՈՆՑ ՍՈՎՈՐՈՒԹՅՈՒՆՆԵՐԸ ԽԻՍՏ ԿԵՐՊՈՎ ՏԱՐԲԵՐՎՈՒՄ ԵՆ ԱՄԵՆԱՄԵՐՁԱՎՈՐ ՏԵՍԱԿԻ ՍՈՎՈՐՈՒԹՅՈՒՆՆԵՐԻՑ։— ՎԵՐԻՆ ԱՍՏԻՃԱՆԻ ԿԱՏԱՐԵԼԱԳՈՐԾՎԱԾ ՕՐԳԱՆՆԵՐ։— ԱՆՑՄԱՆ ԵՂԱՆԱԿՆԵՐԸ։— ԴԺՎԱՐԻՆ ԴԵՊՔԵՐ։— NATURA NON FACIT SALTUM։— ՔԻՉ ԿԱՐԵՎՈՐ ՕՐԳԱՆՆԵՐ։ ՕՐԳԱՆՆԵՐԸ ՈՉ ԲՈԼՈՐ ԴԵՊՔԵՐՈՒՄ ԲԱՑԱՐՁԱԿԱՊԵՍ ԿԱՏԱՐՅԱԼ ԵՆ։ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՏԵՍՈՒԹՅՈՒՆՆ ԸՆԴԳՐԿՈՒՄ Է ՏԻՊԻ ՄԻԱՍՆԱԿԱՆՈՒԹՅԱՆ ԵՎ ԳՈՅՈՒԹՅԱՆ ՊԱՅՄԱՆՆԵՐԻ ՕՐԵՆՔՆԵՐԸ։
Դեռ ամենևին չհասած գրքիս այս կետին, ընթերցողը երևի արդեն նկատել է, որ իր առջև առաջանում են անթիվ դժվարություններ։ Նրանցից մի քանիսն այնքան էական են, որ ես մինչև այժմ չեմ կարողացել նրանց մասին մտածել առանց որոշ հուզմունքի, բայց որքան ես կարող եմ դատել, նրանց մեծամասնությունը միայն թվացող է, իսկ նրանք, որոնք իրական են, ես կարծում եմ չեն կարող համարվել տեսության համար ճակատագրական։ Այդ դժվարությունները և առարկությունները կարելի է բաժանել չորս խորագրի տակ։
Առաջին, եթե տեսակները առաջացել են մյուս տեսակներից անզգալի աստիճաններով, ապա մենք ամենուրեք ինչո՞ւ չենք հանդիպում անթիվ անցողիկ ձևերի։ Ինչո՞ւ ամբողջ բնությունը չի ներկայացնում մեզ համատարած, անլուծելի խառնաշփոթություն, այլ ընդհակառակը, տեսակները լինում են միանգամայն սահմանազատված խմբեր։
Երկրորդ, հնարավո՞ր է արդյոք, որ չղջիկի սովորություններն ու կառուցվածքը ունեցող մի կենդանի կարող էր առաջանալ բոլորովին այլ սովորություններ ու կառուցվածք ունեցող մի ուրիշ կենդանու փոփոխության հետևանքով։ Կարելի՞ է արդյոք հավատալ, որ բնական ընտրությունը կարող էր առաջ բերել մի կողմից այնպիսի չնչին նշանակություն ունեցող օրգան, ինչպիսին է ընձուղտի պոչը, որը օգտագործվում է միայն ճանճեր քշելու համար, և մյուս կողմից, այնպիսի զարմանալի օրգան, ինչպիսին է աչքը։
Երրորդ, կարո՞ղ են արդյոք բնազդները ձեռք բերվել և փոփոխվել բնական ընտրությամբ։ Ի՞նչ կասենք մենք մեղվի բնազդի մասին, որի շնորհիվ նա կառուցում է մեղրահացեր և գործնականում գերազանցել է խորամիտ մաթեմատիկոսների հայտնագործություններին։
Չորրորդ, ինչպե՞ս մենք կարող ենք մեզ բացատրել այն փաստը, որ տեսակները իրենց խաչաձևման ժամանակ լինում են անպտուղ կամ արտադրում են անպտուղ սերունդ, մինչդեռ տարատեսակների խաչաձևման ժամանակ նրանց պտղաբերությունը մնում է անփոփոխ։
Դժվարությունների առաջին երկու կատեգորիաները կպարզաբանվեն այս գլխում, մի քանի բազմազան առարկություններ կքննարկվեն հաջորդ գլխում, իսկ «բնազդը» և «հիբրիդացումը» կկազմեն առանձին գլուխների խնդիրը։
ԱՆՑՈՒՄՆԱՅԻՆ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԲԱՑԱԿԱՅՈՒԹՅԱՆ ԿԱՄ ՀԱԶՎԱԴԵՊ ԼԻՆԵԼՈՒ ՄԱՍԻՆ
Քանի որ բնական ընտրությունը ներգործում է միայն ձեռնտու փոփոխությունների պահպանման շնորհիվ, ուստի օրգանական բնակչությամբ արդեն գերլցված երկրում ամեն մի նոր առաջացած ձև կձգտի զբաղեցնել իր ավելի քիչ կատարելագործված նախնիքների կամ այլ, ավելի քիչ առավելություն ունեցող ձևերի տեղը, որոնց հետ մտնում է մրցության մեջ, և վերջապես կոչնչացնի նրանց։ Այսպիսով, մահաջնջումը և բնական ընտրությունն ընթանում են ձեռք ձեռքի տված։ Հետևաբար, եթե մենք տեսակներն ընդունենք որպես մեզ անհայտ ձևերի հաջորդներ, ապա այդ սկզբնական ձևերը և բոլոր միջակա տարատեսակները պետք է բնաջնջված լինեն նոր ձևի առաջացման և կատարելագործման պրոցեսի միջոցով։
Բայց քանի որ այդ տեսության հիման վրա, անթիվ անցումային ձևերը պետք է, այնուամենայնիվ, գոյություն ունենային, ապա ինչո՞ւ մենք չենք հանդիպում նրանց անհամար թվով երկրի կեղևում պարփակված։ Ավելի հարմար կլինի քննարկել այդ հարցը երկրաբանության տարեգրության անլրիվությանը նվիրված գլխում, իսկ այստեղ ես միայն կասեմ, որ իմ կարծիքով, դրա պատասխանը գտնվում է այն փաստի մեջ, որ այդ տարեգրությունը բոլորովին այնպես լրիվ չէ, ինչպես սովորաբար ընդունված է կարծել։ Երկրի կեղևը ընդարձակ թանգարան է, բայց նրա կոլեկցիաները հավաքել են շատ անկատար ձևով և երկար ընդմիջումներից հետո։
Բայց կարող է պատահել, որ այն դեպքում, երբ մի քանի միմյանց մոտ տեսակներ բնակվում են միևնույն մարզում, մենք, իհարկե, պետք է ներկայումս էլ գտնենք նրանց մեջ շատ անցումնային ձևեր։ Կանգ առնենք մի պարզ օրինակի վրա. որևէ մայր ցամաքով ճանապարհորդելիս հյուսիսից դեպի հարավ, մենք սովորաբար հանդիպում ենք հաջորդականորեն, մեկը մյուսին հաջորդող միմյանց ազգակից կամ այսպես կոչված փոխարինող տեսակների, որոնք հավանորեն գրեթե միևնույն տեղերն են գրավում երկրի բնական տնտեսության մեջ։ Այդ տեսակները երբեմն հանդիպում են և միահյուսվում են միմյանց հետ, բայց քանի որ մեկը նոսրանում է, մյուսը դառնում է ավելի ու ավելի սովորական, մինչև որ չի փոխարինում իրենով առաջինին։ Բայց եթե մենք համեմատենք նմուշահատերը այն գոտուց, որտեղ հանդիպում են երկու ձևերը, ապա նրանք սովորաբար նույնպես խիստ կերպով տարբերվում են միմյանցից կառուցվածքի բոլոր մանրամասնություններով, ինչպես որ այն նմուշահատերը, որոնք վերցվել են այն մարզի կենտրոնից, որտեղ բնակվում է նրանցից յուրաքանչյուրը։ Իմ տեսության հիման վրա այդ ազգակից տեսակները ծագում են մի ընդհանուր նախահորից և իրենց փոփոխման պրոցեսի ժամանակ նրանցից յուրաքանչյուրը հարմարվել է իր մարզի կենսապայմաններին, ոչնչացնելով և փոխարինելով իր նախասկզբնական ձևը և բոլոր միջակա տարատեսակները, որոնք կապում են նրա նախկին վիճակը արդիականի հետ։ Այդ պատճառով էլ մենք առիթ չունենք սպասելու, որ ներկայումս հանդիպեն բազմաթիվ անցումային տարատեսակներ մարզերից յուրաքանչյուրում, թեև այդ տարատեսակները պետք է գոյություն ունենային և գուցե պահպանվել են բրածո վիճակում։ Բայց ինչո՞ւ միջակա գոտում, որը ներկայացնում է նաև միջակա կենսապայմաններ, ապա այնտեղ մենք չենք հանդիպում այնպիսի տարատեսակների, որոնք կազմում են սերտ կերպով մոտեցած տեսակները միմյանց հետ շաղկապող օղակներ։ Այս դժվարությունը երկար ժամանակ տարակուսանք էր պատճառում ինձ։ Սակայն ես կարծում եմ, որ այն կարող է զգալի չափով բացատրված լինել։
Նախ և առաջ, այն փաստից, որ մի որոշ մարզ այժմ մեզ ներկայանում է անընդհատ, կարելի է միայն մեծ զգուշությամբ եզրակացնել, որ այն երկարատև ժամանակաշրջանի ընթացքում էլ այդպես անընդհատ է եղել։ Երկրաբանությունը համոզում է մեզ այն բանում, որ մայր ցամաքների մեծ մասը բաժանված է եղել կղզիների, մինչև անգամ երրորդային շրջանի վերջում, իսկ այդպիսի կղզիներում ինքնուրույն տեսակներ կարող էին առաջանալ անկախ կերպով, առանց նրանց միջև միջակա տարատեսակների առաջացման որևէ հնարավորության, միջակա գոտիներում։ Ցամաքի գծագրությունների և կլիմայի փոփոխությունների հետևանքով, ծովային մարզերը, որոնք այժմ անընդհատ են, դեռ ոչ այնքան վաղ ժամանակներում կարող էին լինել ավելի անջատված և ավելի քիչ միանման։ Կանգ չեմ առնի դժվարությունից խուսափելու այդ ձևի վրա, որովհետև ես կարծում եմ, որ բազմաթիվ պարզորոշ սահմանազատված տեսակները առաջացել են անընդհատ տարածություններում. բայց միևնույն ժամանակ ես բնավ չեմ կասկածում, որ տարածությունների նախկին բաժանվածությունը, որոնք ներկայումս անընդհատ են, կարևոր դեր է խաղացել տեսակների առաջացման պրոցեսում, առանձնապես միմյանց հետ ազատ կերպով խաչաձևվող, թափառական կյանք վարող կենդանիների վերաբերմամբ։
Ուշադրության առնելով տեսակների ժամանակակից բաշխումը լայնարձակ տարածության վրա, մենք սովորաբար նկատում ենք, որ նրանք բավականաչափ բազմաթիվ են նրա տարածության մեծ մասում, այնուհետև նրա սահմանների մոտ բավական անսպասելիորեն սկսում են նոսրանալ և վերջապես անհետանում են։ Այսպիսով, երկու տեսակների ներկայացուցիչների միջև գտնվող չեզոք գոտին սովորաբար նեղ է նրանցից յուրաքանչյուրին պատկանող մարզերի համեմատությամբ։ Նույնը տեսնում ենք լեռները բարձրանալիս և երբեմն ուղղակի զարմանալի է, ըստ Ալֆոնս-դը-Կանդոլի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] դիտողության, թե ինչպես հանկարծ անհետանում են սովորական ալպիական տեսակները։ Նույն փաստը նկատել է Է. Ֆորբսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] խորջրյա ծովային ուսումնասիրությունների ժամանակ, որոնք կատարվել են հողհանի միջոցով։ Նրանց, ովքեր տեսնում են կլիմայի և ֆիզիկական պայմանների մեջ օրգանիզմների տարաբնակեցումը որոշող բացառիկ տարրեր, այդ փաստերը պետք է ապշեցնեն, որովհետև կլիման և բարձրությունը և կամ խորությունը փոփոխվում են անզգալի աստիճանականությամբ։ Բայց, երբ մենք աչքաթող չենք անում այն նկատառումը, որ գրեթե յուրաքանչյուր տեսակ, մինչև անգամ իր մայր երկրում կարող էր բազմանալ հսկայական չափերով, եթե չհանդիպեր այլ տեսակների մրցությանը, որ գրեթե բոլոր տեսակները ապրում են այլ տեսակների հաշվին, կամ իրենք են ծառայում նրանց որպես որս, մի խոսքով, որ ամեն մի օրգանական էակ, ուղղակի կամ անուղղակի ձևով էականորեն կապված է այլ էակների հետ,— այդ ժամանակ մենք հեշտությամբ համոզվում ենք, որ որևէ երկրի բնակիչների տարածումը ոչ մի կերպ կախված չէ բացառապես անզգալիորեն փոփոխվող ֆիզիկական պայմաններից, այլ մեծ չափով կախված է այլ տեսակների ներկայությունից, որոնց հետ նրանք մտնում են մրցման մեջ։ Բայց քանի որ այդ տեսակներն իրենցից ներկայացնում են խիստ սահմանազատված, այլ ոչ թե անզգալի անցումներով միաձուլվող մի ինչ որ բան, ուստի և յուրաքանչյուր տեսակի տարաբնակման սահմանը, կախված լինելով այլ տեսակների տարաբնակման սահմանից, կլինի խիստ նկատելի։ Բացի դրանից, յուրաքանչյուր տեսակ իր տարաբնակման սահմանների մոտ, որտեղ նա գոյություն ունի անհատների փոքր թվով, նրա թշնամիների կամ որսի քանակության տատանումների դեպքում, կամ կլիմայական անբարենպաստ պայմանների դեպքում շատ հեշտությամբ կենթարկվի լիակատար ոչնչացման և դրա հետևանքով նրա աշխարհագրական սահմանները ավելի խիստ նկատելի կլինեն։
Որովհետև ազգակից կամ փոխարինող տեսակները, որոնք ապրում են անընդհատ տարածության վրա, սովորաբար բաշխվում են այնպես, որ յուրաքանչյուրը տարաբնակված է լայն սահմաններում՝ ունենալով իրենց միջև համեմատաբար նեղ չեզոք գոտի, որտեղ նրանք գրեթե հանկարծ նոսրանում են, ապա և տարատեսակները, որոնք էականորեն չեն տարբերվում տեսակներից, պետք է ենթարկվեն նույն կանոնին, այսինքն, եթե մենք կանգ առնենք խիստ փոփոխվող և մեծ տարածություն գրավող տեսակի վրա, ապա մենք պետք է ենթադրենք, որ երկու տարատեսակները հարմարված են երկու մեծ տարածություններին, իսկ երրորդը՝ նեղ, միջին գոտուն, միջակա տարատեսակը, որպես փոքր և նեղ տարածության վրա գոյություն ունեցող, ներկայացված կլինի ավելի սակավաթիվ անհատներով և, որքան որ ես կարող եմ նկատել, այդ կանոնը իրոք արդարանում է բնական վիճակում գտնվող տարատեսակների վերաբերմամբ կիրառելու դեպքում։ Ես հանդիպել եմ այդ կանոնի ապշեցուցիչ օրինակներին այն տարատեսակների վերաբերմամբ, որոնք միջակա տեղ են գրավում Balanus սեռի խիստ արտահայտված տարատեսակների միջև։ Եվ, ըստ երևույթին, միստր Ուոտսոնի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 62] Ազա-Գրեյի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 68] և Ուոլլաստոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 65] կողմից ինձ հայտնած տեղեկությունների հիման վրա, այն տարատեսակները, որոնք միջակա են երկու ձևերի միջև, սովորաբար ներկայացված են լինում զգալիորեն ավելի քիչ թվով, քան այն ձևերը, որոնց նրանք շաղկապում են։ Բայց եթե միայն մենք վստահենք այդ վկայություններին ու փաստերին և գանք այն եզրակացության, որ երկու այլ տարատեսակներ շաղկապող տարատեսակները սովորաբար գոյություն ունեն անհատների ավելի քիչ թվով, քան նրանցով շաղկապված ձևերը, այն ժամանակ մենք կըմբռնենք, թե ինչու այդ միջակա տարատեսակները չեն կարող իրենց գոյությունը պահպանել առանձնապես երկար ժամանակ, թե ինչու, որպես ընդհանուր կանոն, նրանք պետք է ենթարկվեն ոչնչացման և վերանան ավելի շուտ, քան այն ձևերը, որոնց սկզբում նրանք շաղկապել են։
Ամեն մի ձև, որն ունի ավելի քիչ թվով անհատներ, ինչպես առաջ արդեն նկատել ենք, ոչնչացված լինելու ավելի շատ հավանականությունների է ենթարկվում, քան բազմաթիվ անհատներ ունեցողը, իսկ ներկա հատուկ դեպքում միջակա ձևը առանձնապես ենթակա կլինի իր երկու կողմում ապրող մերձակա ձևերի ներխուժմանը։ Բայց ավելի կարևոր է այն նկատառումը, որ հետագա փոփոխման պրոցեսի ժամանակ, որի հետևանքով երկու ենթադրյալ տարատեսակները փոխարկվում են ինքնուրույն տեսակների, այդ երկու տարատեսակները, որոնք ներկայացված են ավելի մեծ թվով և ապրում են ավելի մեծ տարածությունների վրա, կունենան զգալի առավելություններ միջակա տարատեսակի հանդեպ, որը սակավաթիվ է և բնակվում է միջակա նեղ գոտում, որովհետև անհատներով հարուստ ձևերը կունենան ամեն մի տվյալ ժամանակաշրջանում բարենպաստ փոփոխությունների առաջացման ավելի շատ շանսեր, որոնց կարող է կառչել բնական ընտրությունը։ Այսպիսով, կենսական մրցության մեջ սովորական ձևերը կհաղթեն և դուրս կմղեն ավելի քիչ տարածված ձևերին, որովհետև այս վերջիններն ավելի դանդաղ են փոփոխվում ու կատարելագործվում։ Ելնելով այդ սկզբունքից, կարծում եմ, մենք նաև կարող ենք բացատրել երկրորդ գլխում շարադրված այն փաստը, որ սովորական ձևերը ներկայացնում են միջին հաշվով ավելի մեծ քանակությամբ ցայտուն տարատեսակներ, քան հազվագյուտ տարատեսակները։ Կպարզաբանեմ այդ օրինակով։ Ենթադրենք, թե որևէ մի տեղ պահում են ոչխարի երեք տարատեսակ։ Մեկը հարմարված է ընդարձակ լեռնային երկրի, մյուսը՝ համեմատաբար նեղ բլրաշատ գոտու, երրորդը՝ այդ լեռների ստորոտում փռված լայն հարթավայրին։ Այնուհետև ենթադրենք, թե տեղական բնակիչները միատեսակ ուշադրությամբ ու տոկունությամբ հոգում են այդ ցեղերի կատարելագործման մասին ընտրության միջոցով։ Հաջողության ավելի շատ շանսեր կունենան լեռնային և հարթավայրային ավելի մեծ սեփականատերերը, քան միջակա բլրաշատ գոտու, փոքր հողամասերի սեփականատերերը և հետևաբար լեռնային և տափաստանային կատարելագործված ցեղերը շուտով դուրս կմղեն բլրաշատ գոտու ավելի քիչ կատարելագործված ցեղին։ Հետևանքը այն կլինի, որ երկու ավելի բազմաթիվ ցեղերն անմիջական շփման մեջ կմտնեն միմյանց հետ և այլևս բաժանված չեն լինի միմյանցից միջակա ցեղով։
Ամփոփելով ասվածը, ես կարծում եմ, որ տեսակները դառնում են ինչ որ կատարելապես որոշակի սահմաններ ունեցող բան և ընդհանրապես իրենցից չեն ներկայացնում անզգալիորեն փոփոխվող և միահյուսվող օղակների անլուծելի քաոս հետևյալ պատճառներով։
Առաջին, այն պատճառով, որ նոր տարատեսակները առաջանում են խիստ դանդաղ, որովհետև փոփոխման պրոցեսն ինքը կատարվում է ծայրահեղ դանդաղությամբ. իսկ բնական ընտրությունը ոչինչ չի կարող անել, մինչև որ չարտահայտվեն բարենպաստ անհատական փոփոխությունները կամ տարբերությունները կամ մինչև որ տվյալ երկրի բնական կարգում չբացվի մի տեղ, որը կարող են հաջողությամբ զբաղեցնել նրա մեկ կամ մի քանի փոփոխված բնակիչները։ Իսկ այդ նոր տեղերը կախման մեջ կլինեն կլիմայական դանդաղ փոփոխություններից, նոր ձևերի պատահական ներգաղթից և, ամենայն հավանականությամբ, էլ ավելի զգալի չափով երկրի հին բնակիչների դանդաղ փոփոխությունից ու այդ նոր առաջացած և հին ձևերի միջև տեղի ունեցող փոխազդեցությունից։ Այնպես որ, որևէ տվյալ երկրում և տվյալ ժամանակում, մենք կարող ենք սպասել միայն փոքր թվով տեսակներ, որոնք ունեն կառուցվածքի մեջ մի քիչ կայուն հատկությամբ աննշան փոփոխություններ, և այդ իրականում նկատվում է։
Երկրորդ, այժմյան անընդհատ տարածությունները, հաճախ դեռ ոչ այնքան հեռավոր անցյալում ներկայացրել են բաժանված մասեր, որտեղ բազմաթիվ ձևերը,— մանավանդ յուրաքանչյուր ծննդի համար զուգավորվող և շարժուն կենսաձև վարող դասերին պատկանողները, կարող էին առանձնանալ և հասնել տարբերության այնպիսի աստիճանի, որը բավարար է նրանցից, այսպես կոչվող փոխարինող տեսակների առաջացման համար։ Այդ դեպքում այդ փոխարինող տեսակների և նրանց ընդհանուր նախահոր միջև անցումային ձևերը պետք է գոյություն ունենային յուրաքանչյուր մեկուսացած մարզում, բայց ընտրության պրոցեսի միջոցով այդ միացնող օղակները փոխարինվել են այլ ձևերով և ոչնչացել, այնպես որ չեն պահպանվել կենդանի վիճակում։
Երրորդ, երբ երկու տարատեսակ, կամ ավելի մեծ թվով տարատեսակներ առաջացել են անընդհատ տարածության տարբեր մասերում, ապա միջակա գոտում կարող էին, շատ հավանական է, առաջանալ նաև միջակա տարատեսակներ, բայց նրանց գոյությունը թերևս եղել է վաղանցիկ, որովհետև այդ տարատեսակները, առաջներում արդեն նշված (և մերձավոր փոխարինող տեսակների, ինչպես նաև ընդունված տարատեսակների բաշխման մասին մեր տեղեկություններից վերցված) նկատառումների համաձայն գոյություն ունեն միջակա գոտում ավելի քիչ թվով, քան այն տարատեսակներր, որոնց նրանք շաղկապում են իրար հետ։ Հենց միայն այդ պատճառով, միջակա տարատեսակները ավելի ենթակա կլինեն պատահական ոչնչացման, իսկ հետագա փոփոխման պրոցեսում, բնական ընտրության միջոցով կհաղթվեն, կփոխարինվեն ձևերով, որոնց նրանք շաղկապում են, որովհետև այդ վերջինները, շնորհիվ իրենց մեծաքանակության, ընդհանուր առմամբ կներկայացնեն ավելի շատ փոփոխություններ և հետևապես ավելի շատ կկատարելագործվեն ընտրությամբ և ձեռք կբերեն էլի նոր առավելություններ։
Եվ, վերջապես, նկատի ունենալով օրգանիզմների փոփոխումները ոչ թե այս կամ այն ժամանակում, այլ ընդհանրապես ժամանակի մեջ,— եթե միայն իմ տեսությունը ճիշտ է,— մենք պետք է ընդունենք, որ մի խմբի բոլոր տեսակներին մերձավոր օղակներով շաղկապող անթիվ միջակա տարատեսակները մի ժամանակ անկասկած գոյություն են ունեցել, բայց որ բնական ընտրության պրոցեսն ինքը մշտապես թեքվում է դեպի այն, որպեսզի, ինչպես արդեն բազմիցս նշվել է, ոչնչացնի նախասկզբնական ձևերը և նրանց շաղկապող օղակները։ Հետևաբար, նրանց նախկին գոյության մասին վկայություն կարելի է գտնել միայն բրածո մնացորդների մեջ, որոնք, ինչպես մենք կաշխատենք ցույց տալ հետագա գլուխներից մեկում, ներկայացնում են միայն շատ թերի, կցկտուր տարեգրություն։
ՅՈՒՐԱՀԱՏՈՒԿ ՍՈՎՈՐՈՒԹՅՈՒՆՆԵՐ ԵՎ ԿԱՌՈՒՑՎԱԾՔ ՈՒՆԵՑՈՂ ՕՐԳԱՆԱԿԱՆ ԷԱԿՆԵՐԻ ԾԱԳՄԱՆ ՄԱՍԻՆ ԵՎ ՆՐԱՆՑ ՄԻՋԵՎ ՏԵՂԻ ՈՒՆԵՑՈՂ ԱՆՑՈՒՄՆԵՐԻ ՄԱՍԻՆ
Իմ պաշտպանած հայացքների հակառակորդները շատ անգամ առաջ են քաշել մի հարց, թե ինչպե՞ս, օրինակ, կարող էր ցամաքային գիշակեր կենդանին փոխվել ջրայինի, որովհետև ի՞նչպես կարող էր գոյություն ունենալ այդ կենդանին իր անցումային վիճակում։ Դժվար չէր լինի ցույց տալ, որ ներկայումս էլ գոյություն ունեն գիշակեր կենդանիներ, որոնք ներկայացնում են բոլոր հաջորդական և մերձավոր անցումային ձևերը խիստ ցամաքայինների և ջրայինների միջև։ Բայց քանի որ նրանցից յուրաքանչյուրը գոյություն ունի, դիմանալով գոյության կռվին, ապա ակներև է, որ նրանք լավ են հարմարված իրենց տեղին բնության մեջ։ Նայեցեք Հյուսիս-ամերիկյան Mustela vison-ին, որը մատների միջև թաղանթներ ունի և իր մորթիով, կարճ ոտքերով և պոչի ձևով հիշեցնում է ջրասամույրին։ Ամռանը այդ կենդանին կերակրվում է ձկնով, որը որսալու համար սուզվում է ջրի մեջ, իսկ երկարատև ձմռան ընթացքում թողնում է սառած ջրերը և այլ ժանտաքիսների նման որսում է մկներ և այլ ցամաքային կենդանիներ։ Եթե այդ հարցը տվողները գլխի ընկնեին և հարցնեին, թե ի՞նչպես միջատակեր չորսոտանին կարող էր փոխվել չղջիկի, ապա այս անգամ պատասխանելը ավելի դժվար կլիներ։ Բայց և այնպես ես կարծում եմ, որ այդ դժվարությունները մեծ կշիռ չունեն։
Այս անգամ էլ, ինչպես և այլ դեպքերում, ես կրկին գտնվում եմ աննպաստ վիճակում, որովհետև իմ կողմից հավաքած բազմաթիվ զարմանալի օրինակներից այստեղ կարող եմ բերել մեկը կամ երկուսը, որոնք պարզաբանում են միմյանց մոտ տեսակների կենսաձևի ու կազմվածքի անցումները և միևնույն տեսակի ներկայացուցիչների սովորությունների պատահական կամ հաստատուն բազմազանությունը։ Բայց ինձ թվում է, որ օրինակների երկար մի շարք կարող է լիովին հարթել դժվարությունները այնպիսի մասնավոր դեպքերում, որ նման են, օրինակ, չղջիկին։
Նայեցեք սկյուռների ընտանիքին։ Դուք այստեղ կհանդիպեք ամենանուրբ անցումների՝ սկսած թեթևակի տափակած պոչերով կենդանիներից կամ մյուսներից, որոնց մոտ, ըստ սըր Ջ. Ռիչարդսոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 112] դիտողության, մարմնի հետին կեսը թեթևակի լայնացած է ու կողերի մաշկը մի քիչ պարկանման է մինչև իսկական, այսպես կոչված, թռչող սկյուռները։ Իսկ թռչող սկյուռների վերջավորությունները իրար հետ և մինչև անգամ պոչի հիմքի հետ կապված են լայն թաղանթով, որը ծառայում է որպես պարաշյուտ և թույլ է տալիս նրանց սահել օդի մեջ ծառից ծառ զարմանալի տարածությունների վրա։ Հազիվ թե կարելի է կասկածել այն բանում, որ կառուցվածքի յուրաքանչյուր առանձնահատկությունը օգուտ է բերում յուրաքանչյուր սկյուռին իր բնական, շրջադրության մեջ, օգնելով նրան ազատվելու գիշատիչ գազաններից ու թռչուններից և ավելի արագ հավաքելու իրեն համար սնունդ, կամ թե, որն ավելի հավանական է, պաշտպանելով նրան հանկարծակի հարձակման վտանգից։ Բայց ղրանից չի հետևում, որ յուրաքանչյուր սկյուռի կառուցվածքը լավագույնն է, որը կարելի է երևակայել ամենատարբեր պայմաններում։ Թող փոխվեն կլիման և բուսականությունը, թող նրանց հետ մրցող այլ կրծողներ կամ նոր գիշատիչներ ներխուժեն երկիրը և թող հները փոփոխվեն և ըստ անալոգիայի կգանք այն եզրակացության, որ սկյուռներից գոնե մի քանիսը կսակավանան և վերջապես կանհետանան, եթե միայն չփոփոխվեն և չկատարելագործվեն համապատասխան ձևով։ Այդ պատճառով էլ ես խոչընդոտ չեմ տեսնում ենթադրելու, որ, մանավանդ փոփոխվող կենսապայմաններում, մշտապես պահպանվեն ավելի ու ավելի զարգացած կողքի թաղանթներ ունեցող անհատները, որովհետև ամեն մի փոփոխություն այդ ուղղությամբ օգտակար կլիներ և կփոխանցվեր, մինչև որ կուտակման և բնական ընտրության այդ ճանապարհով չէր ստացվի լիովին կատարելագործված թռչող սկյուռ։
Իսկ այժմ նայեցեք, այսպես կոչված, թռչող լեմուրին (Galeopithecus), որին նախկինում դասում էին չղջիկների, իսկ այժմ դասում են միջատակերների շարքը (Insectivora)։ Նրա չափազանց լայն կողքի թաղանթը տարածվում է ծնոտների անկյունից մինչև պոչը և ընդգրկում է երկարացված մատներ ունեցող վերջավորությունները։ Այդ թաղանթը օժտված է նրան ձգող մկանով։ Թեև ներկայումս չեն պահպանվել օդում սահելու համար հարմարված այդ օրգանի միջակա օղակները, որոնք կշաղկապեին գալեոպիթեկին այլ միջատակերների հետ, այնուամենայնիվ չկա ոչ մի դժվարություն ենթադրելու համար, որ այդպիսի օղակներ նախկինում եղել են և առաջացել են, ինչպես որ ավելի քիչ հաջողությամբ թռչող սկյուռների մոտ, ընդորում կատարելագործման պրոցեսի ընթացքում, յուրաքանչյուր միջակա քայլ եղել է օգտակար նրա տիրոջ համար։ Ես անհաղթահարելի խոչընդոտ չեմ տեսնում նաև հետագա ենթադրության համար, որ գալեոպիթեկի թաղանթով միացած մատները և նախաբազուկը կարող էին բնական ընտրության ազդեցության ներքո զգալիորեն երկարել, իսկ այդ արդեն նրան կձևափոխեին չղջիկի գոնե թռչելու օրգանների տեսակետից։ Որոշ չղջիկների մոտ, որոնց թռիչքաթաղանթը տարածվում է ուսի գագաթից մինչև պոչը և ներառնում է ետևի վերջավորությունները, մենք ամենայն հավանականությամբ պետք է այդ օրգանում տեսնենք ավելի շուտ սկզբնական հարմարում օդում սահելու, քան թռչելու համար։
Եթե թռչունների մոտ մեկ դյուժին սեռեր վերջնականապես անհետանային, ում մտքով կարող էր անցնել, որ գոյություն են ունեցել թռչուններ, որոնք Էյտոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 113] Micropterus-ի նման կարող էին միայն թափահարել իրենց թևերը, օգտագործել նրանց որպես լողաթևեր կամ որպես առջևի ոտքերի պինգվինի նման, որպես առագաստ ջայլամի նման, կամ, վերջապես պահպանել նրան առանց որևէ գործունեության, ինչպես ապտերիքսի մոտ։ Եվ այնուամենայնիվ, այդ թռչուններից յուրաքանչյուրի կառուցվածքը պիտանի է իրեն համար, որովհետև նրանք բոլորը պետք է ապրեն պայքար մղելով, բայց այդ ոչ մի կերպ չի կարող համարվել ամենալավը ամեն տեսակի պայմաններում։ Ասվածից չպետք է անել այնպիսի եզրակացություն, թե բերված հաջորդական աստիճանները թևի կառուցվածքում ներկայացնում են իրական քայլեր թռիչքի կատարյալ ընդունակության մշակման ճանապարհին, կարող է պատահել, որ այդ միայն վարժության բացակայության հետևանք է, բայց համենայն դեպս այդ օրինակները ցույց են տալիս, թե ինչպիսի բազմազան անցումներ կարող են միանգամայն հնարավոր լինել։
Գիտենալով, որ գլխավորապես ջրում շնչելու համար հարմարված դասերում, ինչպես խեցեմորթներն ու կակղամորթներն են, հանդիպում են ցամաքային կյանքին հարմարված ներկայացուցիչներ, գիտենալով, որ գոյություն ունեն թռչող թռչուններ և կաթնասուններ, ինչպես և ամեն տեսակի միջատներ, իսկ մի ժամանակ գոյություն են ունեցել նաև թռչող սողուններ, միանգամայն կարելի է ընդունել, որ թռչող ձկներն էլ, որոնք սահում են օդում, թեթևակի բարձրանալով և շրջվելով ընթացքի ժամանակ իրենց թպրտացող լողաթևերի օգնությամբ, կարող էին ձևափոխվել իսկական թևավոր կենդանիների։ Եթե այդ իսկապես իրականանար, ապա ո՞ւմ մտքով կանցներ, որ այդ թևավոր կենդանիները իրենց վաղ անցումային վիճակում եղել են երբևէ բաց օվկիանոսի բնակիչներ և օգտագործել են իրենց թերաճ թռչելու ապարատները, ըստ երևույթին, բացառապես նրա համար, որպեսզի փրկվեն այլ ձկների հետապնդումներից։
Երբ մենք հանդիպում ենք մի օրգանի, որը որոշ գործունեության համար իր հարմարվածությամբ հասցված է կատարելագործման բարձր աստիճանի, ինչպիսին է թռչունի թևը թռիչքի համար իր հարմարվածությամբ, մենք չպետք է մոռանանք, որ կենդանիները, որոնք դրանով օժտված են նրա զարգացման վաղ անցումային աստիճաններում, միայն հազվագյուտ դեպքերում կարող էին ապրել մինչև ներկա ժամանակները, որովհետև փոխարինվել են բնական ընտրությամբ կատարելագործված հաջորդներով։ Այնուհետև մենք պետք է եզրակացնենք, որ անցումային դրությունները երկու այնպիսի ձևերի միջև, որոնք հարմարված են բոլորովին տարբեր կենսաձևերի, միայն հազվագյուտ դեպքերում կարող էին հայտնվել զգալի թվով և ներկայացնել երկրորդային ձևափոխություններ։ Այսպիսով, եթե մենք դառնանք թռչող ձկան մեր երևակայական օրինակին, ապա քիչ հավանական է թվում, որ իսկական թռչելու ընդունակ ձկները կարող էին առաջանալ բազմաթիվ երկրորդային փոփոխություններով, որոնք հարմարված էին բազմազան որսակենդանիների հետապնդման համար, այն էլ տարբեր եղանակներով ցամաքի վրա և ջրի մեջ, մինչև որ նրանց օրգանները չեն հասել կատարելագործության այնպիսի աստիճանի, որը կապահովեր նրանց համար վճռական գերակշռություն գոյության կռվում այլ կենդանիների հանդեպ։ Այստեղից էլ օրգանների կառուցվածքում անցումային աստիճաններով օժտված տեսակներ բրածո վիճակում գտնելու շանսերը շատ չնչին կլինեն, քան միանգամայն զարգացած օրգաններով օժտված տեսակներ գտնելու շանսերը, որովհետև առաջինները միշտ ավելի սակավաթիվ են եղել։
Այժմ ես կբերեմ միևնույն տեսակին պատկանող անհատների կենսակերպի բազմազանության ու փոփոխության երկու կամ երեք օրինակներ։ Այդ դեպքերից յուրաքանչյուրում բնական ընտրությունը կարող էր կենդանու կառուցվածքը հարմարեցնել կամ ընդհանրապես նրա փոփոխված սովորություններին, կամ նրա մի քանի սովորություններից բացառապես մեկին։ Սակայն դժվար է որոշել, և մեզ համար էլ էական չէ, արդյոք սկզբից փոխվում են սովորությունները և հետո օրգանների կառուցվածքը, թե՞ կառուցվածքի մեջ հենց փոփոխություններն առաջացնում են սովորությունների փոփոխություններ․ ամենայն հավանականությամբ, թե՛ մեկը և թե՛ մյուսը հաճախ տեղի են ունենում գրեթե միաժամանակ։ Որպես փոփոխված սովորությունների օրինակ, արժե մատնանշել բրիտանական միջատներից շատերին, որոնք սնվում են էկզոտիկ բույսերով, կամ բացառապես արհեստական մթերքներով։ Ինձ պատահել է հետևել հարավ-ամերիկական ճանճորսին (Saurophagus sulphuratus), մերթ ճախրելիս մի տեղի վրա և հետո բազեի պես թռչելով անցնելիս մի ուրիշ տեղ, մերթ ափին կանգնած հանկարծ նետվելիս ջրի մեջ, ձկան ետևից, ինչպես ալկիոն։ Անգլիայում մեծ երաշտահավը (Parus major) երբեմն մագլցում է ծառերի վրա, համարյա ինչպես սիտեղ, կամ կարմրաշամփրուկի պես սպանում է փոքրիկ թռչուններին կտուցով հարվածելով գլուխներին, և ես շատ անգամ տեսել և լսել եմ, ինչպես նա ջարդում էր կենու սերմերը, նրանց խփելով ճյուղերին, ինչպես սիտեղը, Հյուսիսային Ամերիկայում սև արջը, Հերնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 114] դիտումների համաձայն, լողում է ժամերով լայն բացած երախով և այսպիսով որսում է ջրային միջատներին, ճիշտ կետի նման։
Քանի որ մենք երբեմն հանդիպում ենք անհատների, որոնք ունեն իրենց տեսակի ներկայացուցիչներին և նույն սեռի այլ տեսակներին հատուկ սովորություններից բոլորովին տարբերվող սովորություններ, ապա կարող ենք ենթադրել, որ այդպիսի անհատները երբեմն կարող են սկիզբ տալ անկանոն նոր տեսակների, որոնք ունեն անկանոն սովորություններ ու կազմվածք, որը թեթևակի, իսկ երբեմն էլ զգալիորեն տարբերվում է տիպի կազմվածքից։ Եվ այդպիսի դեպքեր իրոք հանդիպում են բնության մեջ։ Արդյոք կարելի է բերել հարմարվածության ավելի զարմանալի օրինակ, քան այն, որը ներկայացնում է մեզ ծառերի բների վրայով մագլցող և կեղևի ճեղքվածքներում միջատներ որսացող փայտփորիկը։ Եվ, այնուամենայնիվ, Հյուսիսային Ամերիկայում հանդիպում են պտուղներով սնվող փայտփորիկներ և ուրիշներ, որոնք ունեն երկարած թևեր և միջատներ են որսում թռչելիս։ Լա-Պլատայի հարթավայրում, որտեղ հազիվ թե կարելի է գտնել գեթ մի ծառ, հանդիպում է փայտփոր (Colaptes campestris), որի երկու մատը ուղղված են առաջ և երկուսը ետ, լեզուն երկար ու սրածայր է, պոչի փետուրները սրածայր են և բավականաչափ կոշտ, որպեսզի թռչունին պահեն ուղղաձիգ վիճակում ձողի վրա, բայց ոչ այնքան կոշտ, ինչպես տիպիկ փայտփորինը և ուժեղ ուղիղ կտուց։ Թեև կտուցը այնքան էլ ուղիղ չէ ու այնքան էլ ուժեղ չէ, ինչպես տիպիկ փայտփորիկինը, բայց և այնպես բավականաչափ ուժեղ է ծառի կեղևը ծակելու համար։ Այսպիսով, այդ Colaptes-ը իր բոլոր էական մասերով իսկական փայտփորիկ է. մինչև անգամ այնպիսի մանր առանձնահատկություններում, ինչպիսիք են գույնը, ձայնի սուր տոնը և ալիքաձև թռիչքը, պարզ կերպով դրսևորվում է նրա արենակցությունը մեր փայտփորիկի հետ։ Բայց և այնպես ես կարող եմ հաստատել ոչ միայն իմ սեփական, այլև իր ցուցմունքներում միշտ մանրազնին Ազարայի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 74] դիտումների հիման վրա, որ այդ թռչունը որոշ ընդարձակ մարզերում երբեք չի մագլցում ծառերի վրա և իր բունը շինում է գետերի զառիթափ ափերին։ Ընդհակառակը, այլ մարզերում, միևնույն փայտփորիկը, միստր Հուդզոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 115] վկայության համաձայն, այցելում է ծառերը և իրեն համար բույն է փորում նրանց բների մեջ։ Կարող եմ բերել մեկ օրինակ ևս, որը ապացուցում է այդ սեռի թռչունների սովորությունների բազմազանությունը։ Մի մեքսիկական Colaptes, ըստ դը Սոսյուրի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 116] նկարագրությունների, կարծր ծառի մեջ ծակեր է փորում և լցնում է դրանք խոզակաղնու պաշարներով։
Մրրկահավերը թռչունների մեջ ամենից շատ օդային և օվկիանոսային են, բայց Հրո երկրի հանդարտ նեղուցներում հանդիպող Puffinuria berardi-ն իր կենսաձևի ընդհանուր կացույթով, սուզվելու զարմանալի ընդունակությամբ, լողալու և թռչելու եղանակով, երբ նրան խրտնեցնում են, հեշտությամբ կարելի է ընդունել ծովային սուզաբադի կամ սուզահավի տեղ, և այնուամենայնիվ դա իսկական մրրկահավ է, միայն իր նոր կենցաղին համապատասխան խոր կերպով փոփոխվել է կազմվածքի շատ գծերով, մինչդեռ Լա-Պլատայի փայտփորիկի կազմվածքը փոփոխված է միայն թեթևակի։ Ամենախորաթափանց դիտողը, ուսումնասիրելով ջրային կեռնեխին մեռած վիճակում, չէր կասկածի, որ նա ունի ստորջրյա սովորություններ, մինչդեռ այդ թռչունը, որը ազգակից է կեռնեխայիններին, իր գոյությունը պահպանում է միայն շնորհիվ իր սուզվելու ընդունակության, ջրի տակ օգտվելով իր թևերով և ոտքերով կառչելով քարերից։ Թաղանթաթևավորների ընդարձակ կարգի բոլոր ներկայացուցիչները ցամաքի վրա ապրող միջատներ են. բացառություն է կազմում Proctotrupes-ը, որն ունի, ըստ սըր Ջոն Լեբբոկի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 60] դիտումների, ջրային միջատի սովորություններ. նա հաճախակի ընկղմվում է ջրի մեջ և սուզվում է, ընդ որում օգտագործում է ոչ թե թաթիկները, այլ թևերը, և հաճախ մնում է ջրի տակ չորս ժամ շարունակ և, այնուամենայնիվ, չնայած այդ բոլորովին շեղված սովորություններին, նա ոչ մի համապատասխան փոփոխություն չունի իր կառուցվածքի մեջ։
Նա, ով որ հավատում է, թե բոլոր էակները ստեղծվել են այնպես, ինչպես մենք նրանց այժմ տեսնում ենք, պետք է երբեմն զարմանք զգա, տեսնելով մի կենդանի, որի սովորությունները չեն համապատասխանում նրա կառուցվածքին։ Կարո՞ղ է որևէ մի բան ավելի ակներև լինել նրանից, որ սագերի ու բադերի թաղանթավոր ոտքերը ստեղծված են լողալու համար։ Այնուամենայնիվ կան լեռնային թաղանթավոր ոտքերով սագեր, որոնք միայն հազվադեպ են մոտենում ջրին։ Ոչ ոք, բացի Օդյուբոնից,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 117] չի տեսել, որ ճղլակն իջնի օվկիանոսի մակերեսի վրա, մինչդեռ այդ թռչունի բոլոր չորս մատները միացված են թաղանթով։ Մյուս կողմից, սուզահավերը ու կերկերները անկասկած ունեն ջրային թռչունների սովորություններ, բայց նրանց մատները միայն բոլորազարդված են թաղանթով։ Թվում է, թե ի՞նչը կարող է ավելի ակներև լինել, քան այն, որ բարձրասրունքների (Grallatores) մոտ հանդիպող թաղանթազուրկ, երկար մատները հարմարված են ճահիճներում ու լողացող բույսերի, վրայով քայլելու համար։ Սակայն ջրային հավիկը և մարգալորը պատկանում են այդ կարգին, և առաջինը գրեթե նույնպիսի ջրային թռչուն է, ինչպես կերկերը, իսկ երկրորդը նույնպիսի ցամաքային թռչուն է, ինչպես լորը կամ կաքավը։ Այս դեպքերում, իսկ նույնանման օրինակներ դեռ շատ կարելի էր բերել,— սովորությունը փոխվել է առանց կազմվածքի համապատասխան փոփոխման։ Լեռնային սագի թաղանթավոր ոտքերը, կարելի է ասել, դառել են գրեթե թերաճ, ռուդիմենտար ըստ իրենց գործունեության, բայց ոչ թե ըստ կազմվածքի։ Ճղլակի մատների միջև թաղանթների խոր կտրվածքները ցույց են տալիս, որ կազմվածքն էլ սկսել է փոփոխվել։
Նա, ով հավատում է առանձին-առանձին ու բազմաթիվ ստեղծագործական ակտերի, իհարկե, կարող է ասել, որ արարիչը կամեցել է, որպեսզի որոշ տիպի էակ գրավեր մի այլ տիպի էակի տեղը, բայց ինձ թվում է, որ դա կլիներ փաստի պարզ կրկնողությունը, միայն ավելի վերամբարձ ոճով։ Իսկ նա, ով ընդունում է գոյության կռվի և բնական ընտրության սկզբունքը, պետք է հնարավոր համարի, որ յուրաքանչյուր օրգանական էակ մշտապես ձգտում է ավելացնել իր թիվը, և եթե որևէ մի էակ փոփոխվում է իր կազմվածքով կամ սովորություններով, ձեռք բերելով դրանով նույն երկրի մի այլ բնակչի հանդեպ թեկուզ և չնչին առավելություն, ապա նա տիրանում է այս վերջինի տեղին, ինչքան էլ որ այդ չնմանվի այն տեղին, որ ինքն է զբաղեցրել։ Եվ այդ տեսակետից նա չի զարմանա հանդիպելով թաղանթավոր ոտերով սագերին ու ճղլակին, բայց որոնք բնակվում են ցամաքի վրա ու հազվադեպ են իջնում ջրի վրա, հանդիպելով մարգալորերին, որ ճահիճների փոխարեն ապրում են մարգագետիններում, հանդիպելով փայտփորիկներին այնտեղ, որտեղ ծառեր չկան, հանդիպելով սուզվող կեռնեխներին և թաղանթաթևավորներին ու սուզաբադերի սովորություններ ունեցող մրրկահավերին։
ՎԵՐԻՆ ԱՍՏԻՃԱՆԻ ԿԱՏԱՐԵԼՈՒԹՅԱՆ ԵՎ ԲԱՐԴՈՒԹՅԱՆ ՀԱՍԱԾ ՕՐԳԱՆՆԵՐ
Այն ենթադրությունը, որ աչքը, առարկայի հեռավորության համապատասխան իր կիզակետային տարածություներ փոխելու, թափանցող լույսի քանակը կանոնավորելու, գնդաձև ու խրոմատիկ խոտորումը ուղղելու անընդօրինակելի հարմարեցում է, այն ենթադրությունը, թե այդ օրգանը կարող էր մշակվել բնական ընտրության միջոցով, անկեղծորեն այդ բանում խոստովանվում եմ, կարող է վերին աստիճանի անհեթեթ թվալ։ Բայց չէ որ երբ առաջին անգամ էլ արտահայտվել է այն կարծիքը, որ արևը կանգնած է, իսկ երկիրը պտտվում է, մարդկային առողջ բանականոլթյունը հայտաարարել է այն կեղծ, փիլիսոփայորեն մտածող ամեն մի մարդ շատ լավ գիտի, որ հին ասացվածքը «ժողովրդի ձայնը աստծո ձայնն է» անկիրառելի է գիտական հարցերում։ Բանականությունը թելադրում է ինձ, որ եթե կարելի լիներ ցույց տալ բազմաթիվ անցումային աստիճանները հասարակ և անկատարելագործված աչքից դեպի ավելի բարդ կառուցված ու կատարելագործված աչքը, ընդորում ամեն մի աստիճանը օգտակար է նրա տիրոջը, իսկ դա կասկածի ենթակա չէ. այնուհետև, եթե աչքը ենթակա է փոփոխության և այդ փոփոխությունները ժառանգական են, իսկ այդ նույնպես կասկածից վեր է. եթե վերջապես, այդ փոփոխությունները կարող էին օգտակար լինել կենդանուն նրա փոփոխված կենսապայմաններում, այդ դեպքում առաջացած այն դժվարությունը, երբ մտածում ենք, թե բարդ և կատարյալ աչքը ծագել է բնական ընտրության միջոցով, ինչքան էլ դա անհաղթահարելի է մեր երևակայության համար, չի կարելի ճակատագրական, խորտակիչ համարել ամբողջ տեսության համար։ Թե ինչպես նյարդը դարձավ լույսի նկատմամբ զգայուն, մեզ գրեթե նույնպես քիչ է վերաբերում, ինչպես և այն հարցը, թե ինչպես է առաջացել կյանքը։ Միայն պետք է նշեմ, որ եթե պարզագույն օրգանիզմներից մի քանիսը, որոնց նյարդերը չեն հայտնաբերված, ընդունակ են ընկալելու լույսի ազդեցությունը, ապա հազիվ թե առիթ կա աներևակայելի համարել, որպեսզի սարկոդայի[47] որոշ զգայուն տարրերը կարողանային կազմել ագրեգատ, որը դարձել է այդ հատուկ զգացողությամբ օժտված նյարդեր։
Փնտրելով որևէ տեսակի մոտ միջակա աստիճանները, որոնցով անցել է օրգանը հետզհետե կատարելագործվելով, մենք, իհարկե, պետք է որ կանգ առնենք առանձնապես նրա նախնիների վերընթաց տոհմագրության վրա, բայց այդ հազիվ թե իրագործելի է, և այդ պատճառով էլ ստիպված ենք կանգ առնել միևնույն խմբի նույնանման տեսակների ու սեռերի վրա, այսինքն միևնույն ընդհանուր նախնիներից առաջացած կողմնակի գծերի վրա։ Այսպիսով, մենք հույս ունենք իմանալու, թե ինչպիսի աստիճանական անցումներ են հնարավոր և ինչպիսին է նրանց անփոփոխ կամ քիչ փոփոխված վիճակում փոխանցման հավանականությունը։ Բայց օրգանի վիճակը, մինչև անգամ տարբեր դասերում, երբեմն կարող է լույս սփռել այն միջակա աստիճանների վրա, որոնցով նա անցել է։ Ամենապարզ օրգանը, որը կարելի է աչք անվանել, կազմված է օպտիկական նյարդից, որը շրջապատված է պիգմենտային բջիջներով և ծածկված է թափանցիկ մաշկով, բայց առանց որևէ մի ոսպնյակի կամ լուսաբեկող մարմնի։ Ժուրդենի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 118] կարծիքով, մենք թերևս կարող ենք մի աստիճան ցած իջնել և էլի կգտնենք պիգմենտային բջիջների կուտակումներ, որոնք հավանորեն ծառայում են որպես նյարդերից զուրկ և կենդանի հյուսվածքի վրա գտնվող տեսողության օրգաններ։ Վերոհիշյալ պարզ կազմությամբ աչքերը զրկված են պարզորոշ տեսողության ընդունակությունից և ծառայում են միայն լույսը մթությունից տարբերելու համար։ Որոշ ծովաստղերի մոտ, նույն հեղինակի նկարագրությամբ, նյարդը շրջապատող պիգմենտային շերտում փոսիկները լցված են թափանցիկ դոնդողանման նյութով, որը իրենից ներկայացնում է բարձրակարգ կենդանիների եղջերաթաղանթի նման կորնթարթ մակերես։ Նա այն ենթադրությունն է անում, որ այդ օրգանը ծառայում է ոչ թե պատկեր ստանալու, այլ միայն ճառագայթներ հավաքելու համար, որով ուժեղանում է նրանց ներգործությունը։ Ճառագայթների համակենտրոնացման մեջ մենք տեսնում ենք առաջին և ամենակարևոր քայլի իրագործումը դեպի պատկեր տվող, իսկական աչքի առաջացումը. մնում է միայն օպտիկական նյարդի մերկ վերջավորությունը տեղավորել, որը որոշ պարզագույն կենդանիների մոտ ընկղմված է մարմնի մեջ խոր, իսկ ուրիշների մոտ դուրս է գալիս մակերեսին մոտիկ համակենտրոնացնող ապարատից պատշաճ հեռավորության վրա, և այդ վերջավորության վրա կստացվի պատկերը։
Հոդվածոտանիների ընդարձակ դասում մենք կարող ենք սկսել օպտիկական ներվից, որը ուղղակի ծածկված է պիգմենտով, ընդորում վերջինը կազմում է բբի նման ինչ որ մի բան, բայց առանց ոսպնյակի կամ որևիցե օպտիկական ապարատի հետքերի։ Միջատների վերաբերմամբ այժմ հայտնի է, որ նրանց մեծ, բարդ աչքերի եղջերաթաղանթի բազմաթիվ նիստերը իրենցից ներկայացնում են իսկական ոսպնյակներ, իսկ կոները պարունակում են արտասովոր կերպով փոփոխված նյարդային թելիկներ։ Բայց օրգանները հոդվածոտանիների մոտ այնպես բազմազան են, որ դեռ Մյուլլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] բաժանել է նրանց երեք ընդհանուր դասի յոթ ստորաբաժանումներով, չհաշված դեռ հավաքական պարզ աչքերի չորրորդ դասը։
Եթե կանգ առնենք փաստերի վրա, որոնք բերված են այստեղ չափազանց համառոտ՝ համեմատած աչքի ձևերի ընդարձակ, բազմազան աստիճանական շարքի հետ, որ ներկայացնում են պարզագույն կենդանիները։ Եթե հիշենք, թե որքան չնչին է ապրող ձևերի թիվը՝ համեմատած մահաջնջվածների հետ, ապա դժվարությունները, որոնք խոչընդոտ են հանդիսանում ընդունելու, որ բնական ընտրությունը կարող էր օպտիկական նյարդից կազմված, պիգմենտով և թափանցիկ թաղանթով ծածկված պարզ ապարատը դարձնել մի այնպիսի կատարելագործված օպտիկական գործիք, ինչպիսին է հոդվածոտանիների յուրաքանչյուր ներկայացուցչի աչքը, այդ դժվարությունները մեզ առանձնապես մեծ չեն թվա։
Նա, ով այդքան հեռու կգնա, չպետք է վարանի մի նոր քայլ ևս անելու, եթե կարդալով այս գիրքը, նա համոզվի, որ այլ տեսակետից չբացատրվող փաստերի ընդարձակ շարքերը բացատրվում են բնական ընտրության միջոցով փոփոխության տեսությամբ, ապա նա պետք է ընդունի որ մինչև անգամ այնքան կատարելագործված կառուցվածքը, ինչպիսին է արծվի աչքը, կարող է առաջանալ նույն ճանապարհով, թեև այս անգամ նա չունի միջակա ստադիաներ։ Ներկայացվել է նաև այսպիսի առարկություն, աչքը փոփոխելու և այն որպես կատարյալ կերպով աշխատող գործիք մշտապես պահպանելու համար, պետք է մի քանի զուգահեռ փոփոխություններ կատարվեն միաժամանակ, պնդելով, իբրև թե դա անհասկանալի է բնական ընտրության միջոցով, բայց ինչպես ես փորձել եմ ապացուցել ընտելացված կենդանիների փոփոխությունների մասին իմ գրքում, հարկ չկա ենթադրելու, որ այդ փոփոխությունները կատարվել են միաժամանակ, միայն եթե նրանք կատարվել են հաջորդաբար՝ չափազանց աննշան աստիճաններով։ Եվ բացի դրանից, տարբեր ձևի փոփոխություններ կարող էին ծառայել միևնույն ընդհանուր նպատակին, ինչպես նշել է Ուոլլեսը.[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] «Եթե ոսպնյակն ունի չափազանց մեծ կամ չափազանց փոքր կիզակետային հեռավորություն, այն կարելի է ուղղել փոփոխելով կորությունը, կամ նրա նյութի խտությունը։ Եթե կորությունը անկանոն է և ճառագայթները չեն հավաքվում մի կետում, այդ դեպքում կորության ճշտության յուրաքանչյուր մեծացումը կլինի արդեն բարելավում։ Ոչ բբի կծկումները, ոչ աչքի մկանային շարժումները չի կարելի համարել, որ անպայման անհրաժեշտ են տեսողության համար, այլ ներկայացնում են միայն կատարելագործումներ, որոնք կարող էին ավելանալ ու բարելավվել այդ գործիքի մշակման ամեն մի ստադիայում։ Կենդանական թագավորության բարձրակարգ բաժնում, հատկապես ողնաշարավորների մոտ, մենք տեսնում ենք այնքան պարզ աչք, որ նա նշտարաձկան մոտ կազմված է թափանցիկ մաշկի փոքրիկ պարկից, որն օժտված է նյարդով և պատած է պիգմենտով, բայց զրկված է որևէ այլ ապարատից։ Ձկների և սողունների մոտ, ինչպես նկատել է Օուենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] «Դիօպտրիկական ապարատը ներկայացնում է կատարելագործվածության աստիճանների մի երկար շարք»։ Զարմանալի է, որ մինչև անգամ մարդու մոտ, Վիրխովի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 119] հեղինակավոր վկայության հիման վրա, հիանալի ոսպնյակ է առաջանում սաղմի մոտ մաշկի պարկանման ծալքում տեղավորված վերնամաշկի բջիջների կուտակումից, իսկ ապակենման մարմինը առաջանում է սաղմնային ենթամաշկային հյուսվածքից։ Բայց աչքի կառուցվածքի զարմանալի, թեև ոչ բացարձակ կատարյալ մանրամասնությունների ծագման մասին ճիշտ եզրակացության հանգելու համար, անհրաժեշտ է, որ մեր բանականությունը հաղթահարի երևակայությանը։ Սակայն, ես ինքս չափազանց խիստ զգացել եմ այդ դժվարությունը, ուստի չեմ զարմանա, որ ուրիշները ևս կարող են տատանվել այդ մտքի հանդեպ, թե հնարավոր է կիրառել բնական ընտրության սկզբունքն այդպիսի լայն սահմաններում։
Դժվար է խուսափել աչքը հեռադիտակի հետ համեմատելուց։ Մենք գիտենք, որ այդ գործիքը կատարելագործվել է մարդկային բարձր մտքերի երկարատև ջանքերով, դրանից մենք եզրակացնում ենք, որ աչքն էլ առաջացել է նման ճանապարհով։ Բայց արդյոք այդպիսի եզրակացությունը թույլ չէ՞ հիմնավորված։ Արդյոք մենք իրավունք ունե՞նք վերագրել արարչին մտավոր հատկություններ, որոնք նման են մարդկայինին։ Եթե մենք կամենում ենք աչքը համեմատել օպտիկական գործիքի հետ, մենք պետք է երևակայության ճիգով պատկերացնենք թափանցիկ հյուսվածքի շերտ, որի արանքները լցված են հեղուկով և նրա տակ փռվող նյարդով, որը զգայուն է լույսի նկատմամբ, այնուհետև մենք պետք է պատկերացնենք, որ այդ շերտի յուրաքանչյուր մասը դանդաղորեն տրոհվում է տարբեր խտություն ունեցող երկրորդական շերտերի, որոնք դասավորված են մեկը մյուսից տարբեր հեռավորության վրա և սահմանափակված են իրենց գծագրությունները աստիճանաբար փոխող մակերեսներով։ Այնուհետև մենք պետք է պատկերացնենք, որ գոյություն ունի գործունյա սկզբունք, բնական ընտրության կամ ամենահարմարվածի կենդանի մնալու ձևով, որը ուշադրությամբ հետևում է այդ թափանցիկ շերտերի յուրաքանչյուր ամենամանր փոփոխությանը և խնամքով պահպանում է նրանցից յուրաքանչյուրը, որը անընդհատ փոփոխվող պայմաններում այսպես թե այնպես նպաստում է ավելի պարզ պատկերի ստացմանը։ Մենք պետք է ենթադրենք, որ գործիքի ամեն մի նոր ձևափոխումը կատարվում է միլիոնավոր օրինակներով և պահպանվում է միայն այնքան ժամանակ, մինչև որ կստացվի ամենալավը, որից հետո բոլոր հները ենթարկվում են ոչնչացման։ Կենդանի մարմիններում փոփոխականությունը կառաջացնի թեթևակի փոփոխություններ, օրգանիզմների վերարտադրողական ընդունակությունը կբազմացնի նրանց գրեթե անսահման քանակությամբ, իսկ բնական ընտրությունը անսխալ հմտությամբ կվերցնի յուրաքանչյուր կատարելագործություն։ Ենթադրենք, թե այդ պրոցեսը կատարվում է միլիոնավոր տարիների ընթացքում և մշտապես կատարվում է բազմաթիվ կենդանի էակների միլիոնավոր անհատների վրա, և մի՞թե մենք չենք կարող պատկերացնել, որ ի վերջո կստացվի կենդանի օպտիկական գործիք, որը նույն չափով գերազանցում է ապակյա գործիքին, ինչ չափով, որ արարչի գործերը գերազանցում են մարդու արտադրանքներին։
ԱՆՑՄԱՆ ԵՂԱՆԱԿՆԵՐԸ
Եթե հնարավոր լիներ ապացուցել, որ գոյություն ունի մի բարդ օրգան, որը չէր կարող առաջանալ բազմաթիվ իրար հաջորդող թույլ փոփոխությունների միջոցով, իմ տեսությունը վերջնականապես կխորտակվեր։ Բայց ես չեմ կարողանում գտնել մի այդպիսի դեպք։ Անկասկած, գոյություն ունեն բազմաթիվ օրգաններ, որոնց վերաբերյալ մենք չգիտենք անցումային աստիճաններ, առանձնապես եթե մենք ուշադրությունը սևեռենք չափազանց մեկուսացած տեսակների վրա, որոնց շուրջը, իմ տեսության հիման վրա, տեղի է ունեցել զգալի բնաջնջում։ Կամ եթե մենք կանգ առնենք այն օրգանի վրա, որն ընդհանուր է մի ամբողջ դասի բոլոր ներկայացուցիչների համար, ապա մենք կընկնենք նույնպիսի վիճակի մեջ, որովհետև օրգանը պետք է առաջանար այն հեռավոր ժամանակաշրջանում, որից հետո զարգացել են այդ դասի բոլոր բազմաթիվ անդամները, և վաղ անցումային ձևերը որոնելով, ստիպված կլինենք հասնել արդեն վաղուց անհետացած, չափազանց հին նախածնողական ձևին։
Մենք պետք է ծայրահեղ զգուշությամբ գանք այն եզրակացության, որ այս կամ այն օրգանը չէր կարող առաջանալ մի շարք ու զգալի անցումների միջոցով։ Կարելի է բերել այդպիսի օրգանի բազմաթիվ օրինակներ, որը ստորին կենդանիների մոտ միաժամանակ կատարում է բոլորովին տարբեր ֆունկցիաներ. այսպես, օրինակ, շերեփակոթիկի թրթուրի ու Cobites ձկան մոտ կերակուր ընդունող խողովակը շնչում է, մարսում է կերը և արտաթորում է։ Հիդրային կարելի է շուռ տալ, և կենդանու արտաքին մակերեսը այդ դեպքում կսկսի մարսել կերակուրը, իսկ ստամոքսը կսկսի շնչել։ Այդպիսի դեպքերում բնական ընտրությունը, եթե այդ ձեռնտու լիներ, կարող է մասնագիտացնել մի քանի ֆունկցիա կատարող մասեր կամ օրգաններ, հարմարեցնելով նրանց որևէ մեկ ֆունկցիայի, և այսպիսով անզգալի աստիճաններով խորապես կփոփոխեր նրանց կառուցվածքը։ Հայտնի են շատ բույսեր, որոնք միաժամանակ արտադրում են տարբեր կազմություն ունեցող ծաղիկներ, եթե այդ բույսերը սկսեին արտադրել մեկ ձևի ծաղիկներ, ապա դրա հետևանքը կլիներ այն, որ տեսակի հատկանիշների մեջ կստացվեր խոր փոփոխություն, որը կիրականանար համեմատաբար հանկարծակի կերպով։ Այնուամենայնիվ, հավանական է, որ միևնույն բույսի վրա հանդիպող այդ երկու տեսակի ծաղիկները սկզբում դիֆերենցվել են աստիճանաբար, անզգալի անցումների միջոցով, որոնք մի քանի դեպքերում կարելի է նկատել նաև այժմ։
Այնուհետև, երկու տարթեր օրգան կամ միևնույն օրգանը երկու խիստ տարբեր ձևերով կարող են նույն անհատի մեջ կատարել միևնույն ֆունկցիան և այդ ներկայացնում է անցման չափազանց կարևոր եղանակ։ Կանգ առնենք մի օրինակի վրա. կան ձկներ, որոնք շնչում են խռիկների միջոցով՝ ջրի մեջ լուծված օդով և միևնույն ժամանակ ազատ օդով, որը գտնվում է նրանց լողափամփուշտում, ընդ որում այդ օրգանը օժտված է անոթներով չափազանց հարուստ միջնապատերով և օդ մատակարարող օդային անցքով (ductus pneumaticus)։ Բերենք մի այլ օրինակ բուսական աշխարհից, մագլցող բույսերը բարձրանում են երեք տարբեր եղանակներով պարուրաձև ոլորվելով, կառչելով իրենց զգայուն խիղբերով կամ կպչելով իրենց օդային արմատիկներով. այդ երեք եղանակները սովորաբար հանդիպում են տարբեր խմբերում, բայց որոշ փոքրաթիվ տեսակներին հատուկ են երկու կամ մինչև անգամ երեք եղանակները, որոնք միավորված են միևնույն անհատի մեջ։ Բոլոր այդպիսի դեպքերում, օրգաններից մեկը կարող է փոփոխվել և կատարելագործվել, այնպես որ կհարմարվի միայնակ կատարել ամբողջ աշխատանքը, բայց հենց փոփոխության պրոցեսի ընթացքում կօգտվի մյուսի օժանդակությունից։ Իսկ հետո, այդ երկրորդ օրգանը կարող է հարմարվել բոլորովին մի այլ գործունեության կամ իսպառ անհետանալ։
Ձկների լողափամփուշտի օրինակն առանձնապես լավ է, որովհետև ակնառու կերպով դրսևորում է վերին աստիճանի կարևոր մի փաստ, որ մի գործունեության՝ լողացող մարմինը պահելու համար կառուցված օրգանը կարող է հարմարվել բոլորովին մի այլ գործունեության, այսինքն շնչելու համար։ Որոշ ձկների լողափամփուշտը կարող է նաև մշակվել, դառնալ լսելիքի օրգանի լրացուցիչ ապարատ։ Բոլոր ֆիզիոլոգներն ընդունում են, որ լողափամփուշտը դիրքով և կառուցվածքով հոմոլոգ է կամ «իդեալական կերպով նման է» բարձրակարգ ողնաշարավոր կենդանիների թոքերին,— հետևաբար առիթ չկա կասկածելու, որ լողափամփուշտը իսկապես դարձել է թոք կամ բացառապես շնչառության համար գործածվող օրգան։
Այդ տեսակետին համապատասխան կարելի է եզրակացնել, որ իսկական թոքեր ունեցող բոլոր ողնաշարավոր կենդանիները ծագել են մի հին ու անհայտ նախատիպից, որն օժտված է եղել լողացող դրության մեջ մարմինը պահելու ապարատով կամ լողափամփուշտով։ Այսպիսով, մենք կարող ենք բացատրել, ինչպես որ ես այդ եզրակացնում եմ այդ մասերի վերաբերյալ Օուենի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] տված հետաքրքրական նկարագրությունից, այդ տարօրինակ փաստը, որ մեր ուտելիքի կամ ըմպելիքի յուրաքանչյուր պատառը պետք է անցնի շնչափողի անցքի վերևից, ենթարկվելով թոքերի մեջ ընկնելու վտանգին, չնայած այն հիանալի հարմարումին, որով փակվում է կոկորդի ճեղքը։ Բարձրակարգ ողնաշարավորների մոտ խռիկները վերջնականապես անհետացել են, բայց սաղմերի մոտ պարանոցի կողքերին ճեղքերը և արտերիաների օղականման ծռվածքները դեռևս նշում են նրանց նախկին դիրքը։ Հնարավոր է, որ բոլորովին կորցված խռիկները բնական ընտրությամբ աստիճանաբար վերամշակվել են որևէ մի այլ դեր կատարելու համար, այսպես, օրինակ, Լանդուան[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 120] ցույց է տվել, որ միջատների թևերն առաջացել են տրախեաներից։ Ուրեմն, շատ հավանական է, որ այդ լայնածավալ դասի մեջ օրգանները, որոնք մի ժամանակ ծառայել են շնչելու համար, իսկապես դարձել են թռչելու օրգաններ։
Օրգանների միջև անցումների հարցը քննարկելիս մի գործունեությունը մյուսի փոխարկելու հնարավորությունը աչքաթող չանելն այնքան կարևոր է, որ ես կբերեմ մի օրինակ ևս այդ հնարավորությունը հաստատելու համար։ Ցողունավոր փնջոտնյաներն ունենում են մաշկի երկու փոքրիկ ծալքեր (ես անվանել են «ձվակիր երասանիկ»), որոնք ծառայում են մածուցիկ արտաթորանքի միջոցով ձվերը պարկում պահելու այնքան ժամանակ, մինչև որ մատղաշ կենդանիները դուրս կգան ձվերից։ Այդ փնջոտնյաները խռիկներ բոլորովին չունեն, իսկ նրանց մարմնի մակերեսը պարկի և փոքրիկ երասանիկի հետ միասին ծառայում են շնչառության համար։ Մյուս կողմից, Balanidae-ը կամ նստակյաց փնջոտնյաները չունեն այդ ձվակիր երասանիկը, ձվերն ազատ կերպով պառկած են պարկի հատակին խնամքով փակված խեցու ներսում. բայց այն տեղում, որտեղ տեղավորվում է երասանիկը, նրանց մոտ լինում է մեծ, խիստ ծալքավոր թաղանթ, որն ազատ կերպով հաղորդակից է պարկի և ամբողջ մարմնի շրջանառական խոռոչների հետ և բոլոր բնախույզների կողմից համարվում է խռիկներ։ Ես կարծում եմ, որ ոչ ոք չի վիճի, որ մի ընտանիքում ձվակիր երասանիկը և մյուսում խռիկները միմյանց հետ խիստ հոմոլոգ են. և իրոք այդ օրգաններն անզգալի աստիճաններով փոխարկվում են մեկը մյուսին։ Ուրեմն, առիթ չկա կասկածելու, որ մաշկի այդ երկու փոքրիկ ծալքերը, որոնք սկզբում ծառայել են որպես ձվակիր երասանիկ և միևնույն ժամանակ էլ թեթևակիորեն մասնակցել են շնչառությանը, աստիճանաբար բնական ընտրության ազդեցության տակ մշակվել են, դարձել խռիկներ, ընդ սմին նրանք միայն մեծացել են իրենց չափերով և կորցրել իրենց կպչուն գեղձերը։ Եթե բոլոր ցողունավոր փնջոտնյաները մահաջնջվեին,— և նրանք իրականության մեջ ավելի շատ են ենթարկվել բնաջնջման, քան նստակյացները,— ո՞ւմ մտքով կանցներ, որ այս վերջին ընտանիքում խռիկներն առաջացել են այն օրգաններից, որոնք սկզբում ծառայել են միայն նրա համար, որպեսզի ձվերը պարկից ջրի միջոցով դուրս չքշվեն։
Գոյություն ունի անցման նաև մի այլ հնարավոր եղանակ՝ դա վերարտադրության շրջանի արագացման կամ դանդաղեցման միջոցն է։ Պրոֆեսոր Կոպը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 121] և մի քանի այլ գիտնականներ Միացյալ Նահանգներում վերջերս առանձնապես պնդում են դրա վրա։ Այժմ հայտնի է, որ որոշ կենդանիներ կարող են բազմանալ շատ վաղ հասակում, մինչև իսկ ավելի շուտ, քան նրանք լիովին ձեռք են բերել իրենց հատկանիշները, եթե որևէ տեսակի այդ ընդունակությունը ամուր կերպով հաստատվել է, ապա հավանական է թվում, որ նրա անհատները վաղ թե ուշ կկորցնեն իրենց զարգացման բարձր ստադիաները, և այդ դեպքում, մանավանդ եթե թրթուրները խիստ տարբերվում են հասունացած կենդանուց, տեսակի կազմվածքը ուժեղ կերպով կփոփոխվեր ու կթուլանար։ Այնուհետև, մեծ թվով կենդանիներ, հասունության հասնելուց հետո արդեն, շարունակում են փոփոխվել իրենց հատկանիշներով գրեթե իրենց ամբողջ կյանքի ընթացքում։ Այսպես, օրինակ, կաթնասունների գանգի ձևը հասակի հետ միասին զգալիորեն փոփոխվում է, որի մասին դոկտոր Մյուրին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 122] բերել է զարմանալի օրինակներ փոկերի վերաբերմամբ։ Ամեն ոք գիտե, որ եղջերուի եղջյուրների ճյուղավորումները տարիքի հետ միասին ավելի ու ավելի բարդանում են, ինչպես նաև թռչունների փետրավորումը դառնում է ավելի փարթամ։ Պրոֆ. Կոպը վկայում է, որ մի քանի մողեսների ատամների ձևը տարիքի հետ միասին զգալիորեն փոփոխվում է։ Խեցեմորթների մոտ, ըստ Ֆրից Մյուլլերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] դիտողությունների, ոչ միայն բազմաթիվ ոչ էական, այլև մի քանի կարևոր մասերը բոլորովին նոր տեսք են ընդունում լրիվ հասունության հասնելուց հետո։ Բոլոր այսպիսի դեպքերում, իսկ դրանցից կարելի էր շատ բերել, եթե վերարտադրության հասակը հետաձգվել է, այդ դեպքում նաև տեսակի հատկությունները, համենայն դեպս նրանց չափահաս վիճակում, կփոփոխվեին, նույնպես անհավանական չէ այն, որ զարգացման նախորդ ավելի վաղ ստադիաները անցնեին կրճատված ձևով և, վերջապես, իսպառ անհետանային։ Արդյոք հաճա՞խ է կատարվում փոփոխության այդպիսի պրոցես և ընդհանրապես այն երբևիցե կատարվե՞լ է արդյոք, հանձն չեմ առնում վճռել, բայց, համենայն դեպս, եթե նա երբևիցե կատարվել է, ապա հավանորեն մատաղ հասակի ու հասունացածի միջև և հասունացած հասակի ու ծեր հասակի միջև եղած բուն տարբերությունները սկզբում ձեռք են բերվել միայն աստիճանական քայլերով։
ՄԱՍՆԱՎՈՐ ԴԺՎԱՐՈՒԹՅՈՒՆՆԵՐ, ՈՐՈՆՑ ՀԱՆԴԻՊՈՒՄ է ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՏԵՍՈՒԹՅՈՒՆԸ
Թեպետ մեծ զգուշությամբ միայն պետք է մոտենանք այն եզրակացության, որ մի որևէ օրգան չէր կարող առաջանալ հաջորդական փոքրիկ անցումային աստիճանների միջոցով, այնուամենայնիվ հանդիպում են դեպքեր, որոնք լուրջ դժվարություններ են ներկայացնում։
Ամենադժվարին փաստերից մեկն այն է, որ անսեռ միջատներն իրենց կազմվածքով հաճախ տարբերվում են միևնույն տեսակի թե՛ արուներից և թե՛ պտղաբերելու ընդունակ էգերից. այդ հարցը կքննարկվի հաջորդ գլխում։ Ձկների էլեկտրական օրգանները ներկայացնում են երկրորդ բացառիկ դժվարությունը, որովհետև դժվար է պատկերացնել, թե ինչպիսի միջակա աստիճաններով կարող էին անցնել այդ զարմանալի օրգանները։ Բայց այս դեպքում հազիվ թե առիթ կա զարմանալու, որովհետև մենք նույնիսկ չգիտենք, թե ինչի համար են ծառայում նրանք։ Գիմնոտի և տորպեդոյի մոտ նրանք, անկասկած, հանդիսանում են պաշտպանության և գուցե նաև որսին հետապնդելու հզոր գործիք, բայց կատվաձկան մոտ, ինչպես նկատել է Մատտեուչին,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 123] համանման մի օրգան պոչում քիչ էլեկտրականություն է զարգացնում, նույնիսկ երբ կենդանին խիստ գրգռված է՝ այնքան քիչ, որ նա հազիվ թե կարող է ծառայել մատնանշված նպատակների համար։ Բացի դրանից, կատվաձուկը, պոչում նշված օրգանից զատ, ինչպես ցույց է տվել դ-ր Մակ Դոննելը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 124] ունի գլխի մոտ մի ուրիշը, որի վերաբերմամբ հայտնի չէ, թե պե՞տք է արդյոք նրան էլեկտրական օրգան համարել, բայց որն ըստ երևույթին հոմոլոգ է տորպեդոյի էլեկտրական մարտկոցին։ Բոլորն ընդունում են, որ այդ օրգանները սերտ անալոգիա ունեն սովորական մկանների հետ ինչպես նյարդերի ներքին կառուցվածքով ու բաշխմամբ, այդպես էլ նրանց վրա զանազան ռեակտիվների ներգործությամբ։ Պետք է նկատել, որ մկանի կծկումը ուղեկցվում է էլեկտրական պարպումով և, ըստ դ-ր Ռադկլիֆի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 125] խոսքերի, «տորպեդոյի անշարժ էլեկտրական ապարատում գոյություն ունի էլեկտրական լիցք, որը բոլորովին նման է անշարժ մկանի և նյարդի մեջ եղածին, իսկ պարպումը տորպեդոյի մոտ փոխանակ մի ինչ-որ բացառիկ բան ներկայացնելու՝ կարող է լինել միայն մկանի և շարժողական նյարդի գործողության ժամանակ դրսևորվող պարպումի մի այլ ձև»։ Դրանից ավելի հեռու մեր բացատրությունն առայժմ չի տարածվում, բայց քանի որ մեր տեղեկություններն այդ օրգանների մասին այնքան անբավարար են և մենք բոլորովին ոչինչ չգիտենք գոյություն ունեցող էլեկտրական ձկների նախնիների կառուցվածքի և կենսակերպի մասին, ուստի չափազանց համարձակ կլիներ պնդել, որ չէին կարող գոյություն ունենալ որևէ բանի համար պիտանի ոչ մի միջակա ստադիաներ, որոնցով այդ օրգանները անցնելով՝ կարող էին աստիճանաբար զարգանալ։
Բայց այդ օրգաններն առաջին հայացքից ներկայացնում են նաև մի այլ, անհամեմատ ավելի կարևոր դժվարություն, այն է՝ նրանք հանդիպում են համարյա մի դյուժին զանազան ձկների մոտ, որոնցից մի քանիսը կապված են իրար հետ շատ հեռավոր ազգակցությամբ։ Երբ միևնույն օրգանը հանդիպում է նույն դասի մի քանի ներկայացուցիչների մոտ, մանավանդ այնպիսիների մոտ, որոնք ապրում են շատ տարբեր կենսապայմաններում, մենք սովորաբար կարող ենք նրա ներկայությունը վերագրել մի ընդհանուր նախահորից ժառանգելուն, իսկ մի քանի ներկայացուցիչների մոտ նրա բացակայությունը վարժության պակասությանը կամ բնական ընտրությանը։ Այնպես որ եթե էլեկտրական օրգանը ժառանգված լիներ որևէ հին նախահորից, մենք պետք է սպասենք, որ բոլոր էլեկտրական ձկները միմյանց հետ մոտ ազգակցության մեջ են գտնվում, բայց իրականում այդ չի լինում։ Երկրաբանությունը նույնպես առիթ չի տալիս ենթադրելու, որ ձկների մեծամասնությունը երբևիցե ունեցել է էլեկտրական օրգաններ, որոնց կորցրել են այդ ձկների փոփոխված հաջորդները։ Բայց երբ մենք ավելի մոտիկից զննում ենք գործը, համոզվում ենք նրանում, որ տարբեր ձկների մոտ այդ օրգանները տեղավորված են մարմնի տարբեր մասերում, որ նրանք տարբեր ձևով են կառուցված ինչպես թիթեղների դասավորման տեսակետից, այնպես էլ, ըստ Պաչինիի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 126] վկայության, նրանց մեջ էլեկտրականություն հարուցելու եղանակով և, վերջապես, որ նրանք օժտված են նյարդերով, որոնք ուղղվում են զանազան կենտրոններից, այս վերջին տարբերությունը թերևս ամենաէականն է։ Այսպիսով, էլեկտրական օրգաններով օժտված ձկների այդ օրգանները չեն կարող դիտվել որպես հոմոլոգ մասեր, այլ միայն որպես օրգաններ, որոնք իրենց աշխատանքով անալոգ են։ Այստեղից առիթ չկա ենթադրելու, որ նրանք ժառանգվել են մի ընդհանուր նախահորից, որովհետև նման դեպքում նրանք պետք է բոլոր տեսակետներից մոտիկ նմանություն ունենային։ Ուրեմն, այն դժվարությունը, որ պարփակվում է շատ հեռավոր ազգակցության մեջ գտնվող տեսակների, ըստ երևույթին, միևնույն օրգանի երևան գալու փաստի մեջ, ինքնըստինքյան բացառվում է, թռցնելով միայն թեև ավելի քիչ էական, բայց և այնպես մի կարևոր դժվարություն, այսինքն՝ այն աստիճանների անհայտությունը, որոնցով պետք է անցներ այդ օրգանը ձկների յուրաքանչյուր առանձին խմբում։
Ճառագայթման օրգանները, որոնք հանդիպում են մեկը մյուսից շատ հեռավոր ընտանիքների պատկանող մի քանի միջատների մոտ և զետեղված են մարմնի տարբեր մասերում, մեր արդի անգիտակության պատճառով հանդիսանում են դժվարության օրինակ, որը միանգամայն զուգահեռ է այն դժվարություններին, որ ներկայացնում են էլեկտրական օրգանները։ Կարելի է բերել դարձյալ նման օրինակներ, այսպես, օրինակ, բույսերի մեջ ծաղկափոշու զանգվածի, որն օժտված է, ծայրին գեղձ ունեցող կոթունով, վերին աստիճանի զարմանալի կառուցվածքը համարյա միևնույնն է խոլորձու մոտ և ծիծեռնախոտի (Asclepias) մոտ՝ այսինքն երկու այնպիսի սեռերի մոտ, որոնք այնքան հեռու են մեկը մյուսից, որքան այդ հնարավոր է երկու ծաղկավոր բույսերի միջև, բայց այս անգամ էլ մասերը միմյանց հետ հոմոլոգ չեն։ Բոլոր դեպքերում, երբ կենդանի էակները մեկը մյուսից կարգաբանական տեսակետից հեռացած են, բայց օժտված բացառիկ և նման օրգանով, այդ օրգանը, ինչպիսին էլ ուզում է լինի նրա նմանությունը թե՛ կառուցվածքով և թե՛ աշխատանքով, միշտ ցուցաբերում է արմատական տարբերություն։ Այսպես, օրինակ, գլխոտանիների կամ թանաքաձկան և ողնաշարավորների աչքերն արտաքուստ զարմանալի կերպով նման են իրար, բայց այնքան լայն կերպով անջատված խմբերում նմանության նվազագույն բաժինն անգամ չի կարող վերագրվել մի ընդհանուր նախահորից ժառանգվելուն։ Մ-ր Մայվարտն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] առաջադրել է այդ օրինակը, որն իբրև թե, նրա կարծիքով, ներկայացնում է բացառիկ դժվարություն, բայց ես չեմ նկատում, թե իսկապես ինչումն է նրա փաստարկի ուժը։ Տեսողության ամեն մի օրգան պետք է կազմված լինի թափանցիկ հյուսվածքից և պետք է բովանդակի որոշ ձևի ոսպնյակ, որպեսզի պատկերն անդրադարձնի մութ խցիկի հետևի պատի վրա։ Այս մակերեսային համեմատությունից ավելի՝ այլևս դժվար թե որևէ ընդհանուր բան կարելի է գտնել թանաքաձկան և ողնաշարավորների աչքի միջև, ինչպես այդ ակներևությամբ բխում է գլխոտանիների աչքի մասին Հենզենի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 127] կատարած հիանալի հետազոտությունից։ Այստեղ, իհարկե, անհնարին է մտնել մանրամասնությունների մեջ, բայց և այնպես ես կարող եմ նշել տարբերության մի քանի գծեր։ Բարձրակարգ գլխոտանիների ոսպնյակը կազմված է երկու մասերից, որոնք զետեղված են մեկը մյուսի հետևին, ինչպես երկու ոսպեր, թե՛ մեկը և թե՛ մյուսը ինչպես կառուցվածքով, այնպես էլ դիրքով արմատական կերպով տարբերվում են ողնաշարավորների համապատասխան մասից։ Ցանցենին կառուցված է բոլորովին այլ կերպ, նրա տարրերն ունեն հակառակ դրություն, և աչքի բուն թաղանթների ներսում է գտնվում նյարդային հանգույցը։ Մկանների հարաբերությունն այնքան տարբեր է, որքան միայն կարելի է պատկերացնել. նույնը ճիշտ է նաև մյուս մասերի վերաբերմամբ։ Այստեղից բավական դժվար է նույնիսկ լուծել այն հարցը, թե որչափ ճիշտ է միևնույն տերմինների կիրառումը գլխոտանիների և ողնաշարավորների աչքը նկարագրելու ժամանակ։ Իհարկե, ամեն ոք ազատ է ժխտելու, որ թե՛ մեկ և թե՛ մյուս դեպքում աչքը մշակվել է թույլ հաջորդական փոփոխությունների բնական ընտրության հետևանքով, բայց եթե այդ մեկնաբանությունն ընդունվի մի դեպքում, ապա այն նույնպես հնարավոր է մյուս դեպքում, և երկու խմբերում օրգանի կառուցվածքում եղած արմատական տարբերությունը կարելի էր կանխատեսել նրանց ծագման նկատմամբ արտահայտված տեսակետի հիման վրա։ Նման այն բանին, երբ երկու մարդ երբեմն իրարից անկախ կերպով անում են միևնույն հայտնագործությունը, այդպես էլ, ըստ երևույթին, վերևում բերված դեպքերից մի քանիսում բնական ընտրությունը, գործելով յուրաքանչյուր էակի օգտին և օգտագործելով յուրաքանչյուր փոփոխություն, առաջացրել է օրգաններ, որոնք նման են, որչափ որ այդ վերաբերում է նրանց աշխատանքին տարբեր էակների մոտ, բայց իրենց նման գծերից ոչ մեկով պարտական չեն մի ընղհանուր նախահորից ժառանգելուն։
Ֆրից Մյուլլերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] ցանկանալով ստուգել այն եզրակացությունները, որոնց ես հանգում եմ այս գրքում, աշխատել է հետևել պատճառաբանության շատ նման ուղղության։ Խեցեմորթների որոշ ընտանիքներ իրենց մեջ պարփակում են մի քանի տեսակներ, որոնք օժտված են շնչառական ապարատով և հարմարված են ջրից դուրս ապրելու համար։ Այդ ընտանիքներից երկուսի մեջ, որոնք առանձնապես մանրազնին կերպով ուսումնասիրվել են Մյուլլերի կողմից և մոտ ազգակից են իրար հետ, տեսակները նման են միմյանց իրենց բոլոր էական հատկանիշներով, այն է՝ զգայարաններով, արյան շրջանառության սիստեմով, բարդ ստամոքսի ներսում մազափնջերով և, վերջապես, ջրի մեջ շնչելու խռիկների կառուցվածքի բոլոր մանրամասնություններով՝ մինչև այն միկրոսկոպիկական կարթիկները ներառյալ, որոնք ծառայում են խռիկները մաքրելու համար։ Այստեղից կարելի էր սպասել, որ այդ երկու ընտանիքների ցամաքի վրա ապրող սակավաթիվ տեսակների մեջ նաև օդային շնչառության համար հավասարապես կարևոր ապարատները պետք է նույնպես նման լինեն։ Ինչո՞ւ, թվում է թե, միայն այդ միակ ապարատը, որ ծառայում է միանման նպատակի համար, պետք է տարբերվեր, մինչդեռ մյուս կարևոր օրգանները միմյանց լիովին նման են, կամ ավելի ճիշտ նույնական են։
Ֆրից Մյուլլերն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] ապացուցում է, որ կառուցվածքի այդքան բազմաթիվ գծերով այդ մոտ նմանությունը, համաձայն իմ առաջադրած տեսակետների, պետք է բացատրվեր ժառանգական փոխանցմամբ մի ընդհանուր նախահորից։ Բայց քանի որ տեսակների հսկայական մեծամասնությունը նշված երկու ընտանիքներում, ինչպես նաև բոլոր խեցեմորթների մեծամասնությունը վարում է ջրային կենդանիների կենցաղ, ուստի վերին աստիճանի անհավանական է թվում, որ նրանց ընդհանուր նախահայրը հարմարված լիներ օդային շնչառության։ Այսպիսով, Մյուլլերը հանգել է օդով շնչող տեսակների ապարատի ուսումնասիրությանը և համոզվել է, որ նրանցից յուրաքանչյուրի մոտ նա տարբերվել է մի քանի շատ էական կողմերով, ինչպիսիք են անցքերի դիրքը, նրանց բացման ու փակման եղանակը և մի քանի այլ երկրորդական մանրամասնություններ։ Այդպիսի տարբերությունները միանգամայն հասկանալի են, դրանց մասին կարելի էր նույնիսկ կանխագուշակել, ենթադրելով, որ տարբեր ընտանիքների պատկանող տեսակներն աստիճանաբար ավելի ու ավելի հարմարվել են, որպեսզի ապրեն ջրից դուրս և շնչեն օդով։ Քանի որ այդ տեսակները պատկանել են զանազան ընտանիքների ուստի և նրանք պետք է որոշ չափով տարբերվեին, և այն սկզբունքի շնորհիվ, որ ամեն մի փոփոխության բնույթ կախված է երկու գործոններից, բուն օրգանիզմի բնույթից և շրջապատող պայմանների բնույթից,— նրանց փոփոխությունները չէին կարող միանգամայն նույնական լինել։ Հետևաբար, բնական ընտրությանը տրամադրված է եղել մշակելու և միանման գործող հետևանքի բերելու համար տարբեր նյութ, այսինքն զանազան փոփոխություններ։ Հասկանալի է, որ այդ ձևով ձեռք բերված օրգաններն անխուսափելիորեն պետք է տարբերվեին իրարից։ Առանձին ստեղծագործական ակտեր ընդունող հիպոթեզի հիման վրա բերված ամբողջ օրինակը անհասկանալի կլիներ։ Ըստ երևույթին այդ ձևի փաստարկումը ամենից ավելի ազդել է Ֆրից Մյուլլերի վրա և դրդել է նրան ընդունելու այն տեսակետները, որոնք ես բերում եմ այս գրքում։
Մի այլ ականավոր կենդանաբան, հանգուցյալ պրոֆ. Կլապարեդը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 128] նման ձևով դատելով, եկել է նույն եզրակացությանը։ Նա մատնանշում է այն բանի վրա, որ գոյություն ունեն մակաբույծ տիզեր (Acaridae,), որոնք պատկանում են ինքնուրույն ենթաընտանիքների ու ընտանիքների և օժտված են ճանկերով մազեր բռնելու համար։ Այդ օրգանները պետք է առաջ գային մեկը մյուսից անկախ կերպով, որովհետև չէին կարող ժառանգված լինել մի ընդհանուր նախահորից, և տարբեր խմբերում նրանք առաջ են եկել առջևի ոտների, ետևի ոտների, ծնոտների կամ շրթունքների կամ, վերջապես, իրանի ետևի մասի ստորին մակերեսի հավելուկների ձևափոխությամբ։
Նախորդ օրինակներում մենք տեսանք, թե ինչպես միմյանց հետ միայն քիչ ազգակից կամ նույնիսկ բոլորովին ոչ ազգակից էակների մոտ արտաքին տեսքով շատ նման, բայց զարգացման եղանակով տարբեր օրգանների օգնությամբ ձեռք է բերվել միևնույն հետևանքը և իրագործվել է միևնույն աշխատանքը։ Մյուս կողմից ամբողջ բնության մեջ կարելի է լայն տարածված համարել այն կանոնը, որ նույնիսկ միմյանց մոտիկ ազգակից ձևերի սահմաններում միևնույն նպատակը ձեռք է բերվում ամենաբազմազան միջոցներով։ Ինչքան տարբեր ձևով են կառուցված թռչունի փետրավոր թևը կամ չղջիկի թաղանթավոր թևը, կամ առավել ևս թիթեռի չորս թևերը, ճանճի երկու թևերը կամ բզեզների վերնաթևերով օժտված թևերը։ Երկփեղկանի խեցիները կառուցված են այնպես, որ կարող են բացվել ու փակվել, բայց որքան բազմազան ձևեր է ստանում այդ փականային ապարատը, սկսած պինդ կերպով իրար մեջ մանող ատամիկներից, ինչպես Nucula-ինը, և վերջացրած պարզ կապանով, ինչպես անատամինն է։ Սերմերը տարածվում են կամ պարզապես իրենց փոքրիկ մեծության շնորհիվ, կամ նրանց տուփիկները վերածվում են թեթև, օդապարիկի նման թաղանթի, կամ նրանք սուզվում են ամենաբազմազան ծագում ունեցող վառ գունավորված սննդարար պտղամսի մեջ, որը գրավում է թռչուններին, որոնք և ուտում են նրանց, կամ նրանք օժտված են բազմազան կարթերով և խարիսխներով կամ սղոցաձև քիստերով, որոնցով կպչում են չորքոտանիների մազերին. կամ թե, վերջապես, նրանք օժտված են թռիչքաթևերով կամ փուփուլներով, որոնք նույնքան տարբեր են իրենց ձևով, որքան որ նուրբ են իրենց կառուցվածքով և որոնց շնորհիվ այդ սերմերը քշվում են ամենաթեթև քամու միջոցով։ Բերեմ էլի մի օրինակ, որովհետև միևնույն նպատակին զանազան միջոցներով հասնելու այս հարցը արժանի է ուշադրության։ Մի քանի հեղինակներ պնդում են, թե օրգանական էակներն ստեղծվել են տարբեր ձևով, պարզապես բազմազանության համար՝ ինչպես խաղալիքի խանութում խաղալիքները, բայց բնության վրա այդպիսի հայեցակետը անհավատալի է։ Բաժանասեռ բույսերը և այն բույսերը, որոնց ծաղիկները թեև երկսեռ են, բայց ծաղկափոշին ինքը չի ընկնում սպիի վրա, բեղմնավորման համար կողմնակի օգնություն են պահանջում։ Բույսերից մի քանիսը դրան հասնում են այն բանի շնորհիվ, որ ծաղկափոշին, լինելով սրսուռ ու թեթև, տարվում է քամու միջոցով և պատահականորեն ընկնում է սպիի վրա։ Դա, իհարկե, ամենապարզ եղանակն է, ինչպիսին միայն կարելի է պատկերացնել։ Համարյա նույնպես պարզ, թեև փոքր ինչ այլ եղանակ է իրականանում շատ բույսերի մոտ, այսինքն՝ մի քանի կաթիլ նեկտար արտադրող համաչափ ծաղիկներին այցելում են միջատները, որոնք այդ ժամանակ փոշանոթներից ծաղկափոշին տեղափոխում են սպիի վրա։
Մեկնելով այս պարզագույն ստադիայից, մենք կարող ենք հետամուտ լինել մի շարք գրեթե անսպառ հարմարումների, որոնք ուղղված են միևնույն նպատակին և էապես նման են, բայց կապված են ծաղկի համարյա բոլոր մասերի փոփոխության հետ։ Նեկտարը կարող է կուտակվել ամենաբազմազան ձևերի զետեղարանների մեջ, իսկ առէջքներն ու վարսանդը կարող են փոփոխվել ամենատարբեր եղանակներով, երբեմն առաջացնելով յուրատեսակ թակարդներ, երբեմն դրսևորելով չափազանց ճիշտ հարմարեցված շարժումներ, որոնք առաջ են գալիս գրգռականությամբ կամ առաձգականությամբ։ Մենք կարող ենք այս օրգաններից է՛լ ավելի հեռու գնալ, մինչև որ կհանդիպենք մի այնպիսի բացառիկ հարմարեցման, ինչպիսին է այն, որ նկարագրել է վերջերս դ-ր Կրյուգերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 129] Coryanthes-ի մոտ։ Այդ խոլորձու labellum-ի, կամ շրթունքի մի մասը ծռված է մեծ դույլի ձևով, որի վրա տեղավորված երկու եղջերանման ելուստները նրա մեջ անընդհատ արտադրում են գրեթե մաքուր ջրի կաթիլներ։ Երբ դույլիկը կիսով չափ լցվում է, ջուրը նրա միջից հոսում է կողքի ակոսիկով։ Շրթունքի հիմնական մասը գտնվում է դույլերից ավելի վեր և սնամեջ է՝ երկու կողքերից անցքեր ունեցող խցիկի նման, որի մեջ տեղավորված են հետաքրքրական, մսալի սանրիկներ։ Ամենահնարագետ մարդը, եթե նա չի տեսել, թե ինչ է տեղի ունենում այդ ծաղկի մեջ, չի կարողանա կռահել, թե ինչի համար են ծառայում այդ բոլոր մասերը։ Սակայն դ-ր Կրյուգերը տեսել է բազմաթիվ մեծ իշամեղուներ, որոնք այցելում են այդ խոլորձու հսկա ծաղիկները ոչ նրա համար, որպեսզի ծծեն նրա նեկտարը, այլ նրա համար, որպեսզի կրծոտեն դույլիկի վրայի շրթունքի վերին խցիկում եղած այդ սանրիկները։ Խռնվելով, նրանք հաճախ մեկը մյուսին հրում են ջրի մեջ, և նրանք, որոնց թևերը թրջվել են, արդեն չէին կարողանում թռչել, այլ ստիպված էին դուրս սողալ բացատար ակոսիկի միջով։ Դ-ր Կրյուգերն առիթ է ունեցել ներկա լինելու այն իշամեղուների «անընդհատ երթին», որոնք դուրս էին գալիս ակամայից ընդունած լոգանքից։ Անցքը շատ նեղ է և վերևից, ինչպես կամարով, ծածկված է ծաղկի մասով, որը կրում է փոշանոթներ և սպի, այնպես որ իշամեղուն հազիվհազ անցնում է ծաղկից դուրս, ստիպված է մեջքով քսվել սկզբում մածուցիկ սպիին, իսկ հետո ծաղկափոշու զանգվածների մածուցիկ գեղձիկներին։ Նոր բացված ծաղկի միջից դուրս եկած իշամեղվի մեջքին այս ձևով կպած այդ ծաղկափոշու զանգվածները դուրս են բերվում ծաղկից։ Դ-ր Կրյուգերն ինձ ուղարկել է սպիրտի մեջ պահված ծաղիկը իշամեղվի հետ, որը սպանվել է նախքան ծաղկից դուրս գալը, և նրա մեջքին կպած ծաղկափոշու զանգվածի հետ։ Երբ իշամեղուն, որն օժտված է այդպիսի ծաղկափոշու զանգվածով, թռչում է մի ուրիշ ծաղկի վրա կամ կրկին նույն ծաղկի վրա, և նրա ընկերները նրան նորից կհրեն ու կգցեն դույլիկի մեջ, իսկ նա դուրս կսողա ակոսիկի միջով, այդ դեպքում ծաղկափոշու զանգվածն անհրաժեշտորեն շփման մեջ է մտնում սկզբում մածուցիկ սպիի հետ, կկպչի նրան, և ծաղիկը կբեղմնավորվի։ Այսպիսով մենք, վերջապես, նկատում ենք, թե ինչ նշանակություն ունեն ծաղկի բոլոր մասերը՝ ջուր արտադրող եղջյուրները, դույլիկը, որի մինչև կեսը լցված է ջրով, որը խանգարում է իշամեղվին թռչել և ստիպում է նրան սողալ ակոսիկի միջով ու քսվել պատշաճ ձևով դասավորված մածուցիկ սպիին և մածուցիկ ծաղկափոշու զանգվածներին։
Մի այլ, կարգաբանական տեսակետից մոտիկ խոլորձու՝ Catasetum-ի ծաղկի կառուցվածքը բոլորովին ուրիշ է, թեև ոչ պակաս զարմանալի է և ծառայում է նույն նպատակի համար։ Իշամեղուները այցելում են այգ ծաղիկներին նույնպես, ինչպես որ Coryanthes-ի ծաղիկներին, որպեսզի կրծոտեն նրա շրթունքը, որի ժամանակ նրանք դիպչում են երկար, սրածայր զգայուն ելունին կամ, ինչպես ես այն անվանել եմ, անտեննային։ Այդ անտեննան ամենափոքր շփման դեպքում գրգիռը կամ ցնցումը հաղորդում է փոքրիկ թաղանթին, որը իսկույն պայթում է. այդ պայթելուց արձակվում է զսպանակը, որը նետի նման դուրս է շպրտում ծաղկափոշու զանգվածը հարկ եղած ուղղությամբ, այնպես որ իր մածուցիկ վերջավորությամբ կպչում է իշամեղվի մեջքին։ Փոշանոթներ ունեցող բույսի (այդ խոլորձու ծաղիկները բաժանասեռ են) ծաղկափոշու զանգվածը, այսպիսով, տեղափոխվում է վարսանդավոր ծաղկի վրա, շփման մեջ է մտնում սպիի հետ, որի մակերեսը բավականաչափ մածուցիկ է, որպեսզի կտրատի ծաղկափոշին կապկպող թելիկները ու պահի նրան, որի հետևանքով տեղի է ունենում բեղմնավորումը։
Հարց է ծագում, թե ի՞նչպես մենք կարող ենք բացատրել այս վերջին դեպքում և անթիվ այլ դեպքերում ևս կազմվածքի աստիճանական բարդացումը և միևնույն նպատակի իրագործումը ամենաբազմազան միջոցներով։ Պատասխանը, անկասկած, նույնն է, ինչ որ բերված է ավելի առաջ, այն է՝ երբ երկու ձևեր, որ արդեն փոքր ինչ տարբերվում են իրարից, կփոփոխվեն, այդ փոփոխությունները չեն կարող լինել բոլորովին միանման հատկությամբ և հետևաբար բնական ընտրության միջոցով ձեռք բերված արդյունքներն էլ, թեև ուղղվում են միանման նպատակի, չեն կարող իրենց բնույթով միանման լինել։ Մենք չպետք է աչքաթող անենք նաև այն, որ յուրաքանչյուր բարձր կազմակերպված էակ իր գոյության պայմաններում անցել է բազմաթիվ փոփոխությունների շարքով, որ կառուցվածքի յուրաքանչյուր փոփոխություն ժառանգվում է այնպես, որ ոչ մի փոփոխություն չի կարող հեշտությամբ կորչել, այլ միայն նորից ու նորից կփոփոխվի։ Այսպիսով, ամեն մի տեսակի յուրաքանչյուր մասի կառուցվածքը, ինչի համար էլ նա ծառայելիս լինի, հանդիսանում է այն ժառանգված փոփոխությունների գումարը, որոնց ենթարկվել է տեսակն իր կյանքի փոփոխված պայմաններին ու ձևին հարմարվելու ընթացքում։
Վերջիվերջո, թեպետ շատ դեպքերում նույնիսկ դժվար է պատկերացնել, թե այս կամ այն օրգանը ինչպիսի անցումային ձևերի շարաններով անցնելով, հասել է իր այժմյան վիճակին, այնուամենայնիվ ի նկատի ունենալով, թե որքան չնչին է մեզ հայտնի ձևերի թիվը գոյություն ունեցածների և մեզ անհայտների համեմատությամբ, ես կարող եմ միայն զարմանալ այն օրգանների թվի աննշանության վրա, որոնց վերաբերմամբ մեզ հայտնի չէ ոչ մի անցումային միջակա ստադիա։ Անկասկած ճիշտ է այն դրույթը, որ նոր օրգանները, որոնք կարծես ստեղծված են որևէ որոշակի նպատակի համար, հազվագյուտ դեպքում կամ նույնիսկ երբեք չեն առաջանում մի որևէ էակի մոտ, ինչպես արտահայտված է հին, թեև կարող է պատահել մի քիչ չափազանցված ասացվածում, «Natura non facit saltum»[48]։ Մենք այդ կարծիքին հանդիպում ենք գրեթե բոլոր հմուտ բնախույզների գրվածքներում։ Միլն Էդվարդսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 86] հիանալի կերպով է արտահայտել նույն միտքը հետևյալ խոսքերով՝ բնությունը առատաձեռն է բազմազանության նկատմամբ, բայց չափազանց ժլատ է նորամուծությունների նկատմամբ։ Առանձին ստեղծագործական ակտերի տեսության հիման վրա ի՞նչպես բացատրել, որ բնությունը մեզ ներկայացնում է այդ բազմազանության հարստությունը և այդքան քիչ իսկական նորություն իր արտադրանքներում։ Ինչո՞ւ համար բազմաթիվ, բոլորովին անկախ էակների, որոնք այդ ենթադրության հիման վրա ստեղծված են առանձին-առանձին բնության մեջ որոշակի տեղեր գրավելու համար, տարբեր մասերը սովորաբար պետք է շաղկապված լինեին միմյանց հետ աստիճանական անցումներով։ Ի՞նչ պատճառով բնությունը հանկարծական թռիչքներով չի անցել մի կառուցվածքից մյուսին։ Բնական ընտրության տեսության հիման վրա մենք հստակ կերպով ըմբռնում ենք՝ թե ինչու։ Բնական ընտրությունը ներգործում է միայն օգտագործելով յուրաքանչյուր թույլ հաջորդական շեղում։ Նա երբեք չի անում հանկարծական, մեծ թռիչքներ, այլ միշտ շարժվում է կարճ, բայց հաստատուն, թեպետև դանդաղ քայլերով։
ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՆԵՐԳՈՐԾՈՒԹՅՈՒՆԸ ՔԻՉ ԿԱՐԵՎՈՐ ԹՎԱՑՈՂ ՕՐԳԱՆՆԵՐԻ ՎՐԱ
Քանի որ բնական ընտրությունը ներգործում է կյանքի և մահվան միջոցով, ամենահարմարվածի գերապրելու և միաժամանակ ավելի քիչ հարմարվածներին ոչնչացնելու միջոցով, ուստի ես երբեմն շատ դժվարանում էի, թե ինչպես բացատրեմ օրգանիզմի սակավ կարևոր մասերի ծագումը կամ առաջացումը։ Այդ դժվարությունը թեև բոլորովին այլ կարգի է, բայց համարյա նույնքան մեծ է, ինչքան այն, որ ներկայացնում են բարձր աստիճանի բարդության nւ կատարելության հասած օրգանները։
Նախ, մենք որևէ օրգանական էակի ընդհանուր էկոնոմիկայի վերաբերյալ շատ քիչ բան գիտենք, որպեսզի կարողանանք վստահությամբ խոսել այն մասին, թե որ աննշան ձևափոխությունն է էական, որն է ոչ էական։ Նախորդ գլուխներից մեկում ես բերել եմ աննշան հատկանիշների օրինակներ, ինչպիսիք են պտուղների խավապատումը կամ նրանց պտղամսի գույնը, չորքոտանիների մաշկի կամ բրդի գույնը, որոնք ընդհանուր կարգի այլ առանձնահատկությունների հետ առնչություն ունենալու պատճառով կամ որպես միջատների հարձակումների դեմ պաշտպանություն, իսկապես կարող էին բնական ընտրության նյութ դառնալ։ Ընձուղտի պոչը հիշեցնում է արհեստական ճանճասպանին. առաջին հայացքից գրեթե անհավատալի է թվում, որ այդ օրգանը կարող էր իր ներկայիս դերի համար մշակվել թույլ փոփոխությունների միջոցով, որոնք շարունակ ավելի ու ավելի հարմարվել են այնպիսի մի աննշան նպատակի համար, ինչպիսին է ճանճերին քշելը։ Այնուամենայնիվ, նույնիսկ այդ դեպքում էլ չի խանգարի ձեռնպահ մնալ չափազանց հապճեպ դատողությունից, քանի որ մենք գիտենք, որ եղջերավոր անասունների և այլ կենդանիների տեղաբաշխումն ու բուն գոյությունը Հարավային Ամերիկայում ուղղակի կապված է միջատների հարձակումներից նրանց պաշտպանվելու ընդունակության հետ։ Այսպիսով, այն անհատները, որոնք կարող էին պաշտպանվել այդ չնչին թշնամիներից, կարող էին տեղափոխվել նոր արոտավայրեր և այդ ճանապարհով ձեռք բերել մեծ առավելություն։ Ճանճերը ոչ թե կարող էին ոչնչացնել խոշոր չորքոտանիների (բացառությամբ մի քանի հազվագյուտ դեպքերի), այլ այդ թշնամիները շարունակ անհանգստացնելով կենդանիներին, այնպես ուժասպառ են անում նրանց, որ նրանք ավելի շատ ենթակա են լինում հիվանդությունների և ավելի քիչ ընդունակ են դառնում սովի ժամանակ իրենց համար սնունդ որոնելու կամ գիշատիչներից ազատվելու։
Հիմա չնչին նշանակություն ունեցող օրգանները որոշ դեպքերում կարող էին անցյալում շատ օգտակար եղած լինել առանձին նախահոր համար և երկարատև, դանդաղ կատարելագործումից հետո նույն վիճակում ժառանգաբար փոխանցվել են գոյություն ունեցող տեսակին, որի համար նրանք արդեն անօգուտ են։ Բայց որևէ իսկապես վնասակար շեղման առջև բնական ընտրությունն անհապաղ սահման կդներ։ Գիտենալով, թե ինչպիսի կարևոր նշանակություն ունի պոչը, որպես ջրային կենդանիների մեծամասնության շարժողության օրգան, կարելի է բացատրել նրա առկայությունը և նրա գործածությունը այդքան շատ կարիքների համար այդպիսի մեծ թվով ցամաքային կենդանիների մոտ, որոնց թոքերը, այսինքն փոփոխված լողափամփուշտը, ցուցաբերում են ջրային ծագում։ Քանի որ ջրային կենդանու մոտ լավ զարգացած պոչն արդեն մշակվել էր, նա կարող էր հետագայում իրեն համար կիրառում գտնել նաև բոլորովին այլ ուղղություններով, այն է՝ որպես ճանճասպան, որպես բռնիչ օրգան կամ որպես օգնություն շրջվելու դեպքում, օրինակ, շների մոտ, թեև այս վերջին դեպքում նրա դերը հազիվ թե մեծ է, որովհետև պոչից գրեթե զուրկ նապաստակը համարյա երկու անգամ ավելի արագ է կատարում շրջադարձեր։
Երկրորդ, մենք կարող ենք սխալվել, որոշ հատկանիշների վերագրելով նշանակություն և ենթադրելով, թե նրանք մշակվել են բնական ընտրության օժանդակությամբ։ Մենք ոչ մի դեպքում աչքաթող չպիտի անենք նաև այլ պատճառների ներգործությունները, ինչպիսիք են, փոփոխված կենսապայմանների որոշակի ազդեցությունը՝ այսպես կոչված, ինքնաբերական փոփոխությունները, որոնք շատ փոքր չափով կախված են կենսապայմաններից. վաղուց կորցված հատկանիշներին վերադառնալու ձգտումը, զանազան մարմնամասերի աճման, փոխհարաբերության, փոխադարձ ճնշման բարդ օրենքները և այլն, վերջապես, սեռական ընտրության ազդեցությունը, որի միջոցով մի սեռի համար օգտակար հատկանիշները հաղորդվում են նաև մյուս սեռին, որի համար այդ հատկանիշները կարող են անօգուտ լինել։ Բայց այսպիսի անուղղակի ճանապարհով ձեռք բերված և տեսակի համար սկզբնապես բոլորովին անօգուտ հատկությունները կարող էին հետագայում օգտակար դառնալ փոփոխված հետնորդների համար՝ նրանց կյանքի նոր պայմաններում և նոր ձեռք բերված սովորությունների դեպքում։
Եթե գոյություն ունենային միայն կանաչ փայտփորիկներ և մենք չգիտենայինք սևերի և խայտաբղետների գոյության մասին, մենք ամենայն հավանականությամբ կերևակայեինք, թե կանաչ գույնը հիանալի հարմարանք է ներկայացնում նրա համար, որպեսզի թաքցնի այդ ծառաբնակ թռչունին իր թշնամիներից, և հետևաբար շատ էական հատկանիշ է և ձեռք է բերված բնական ընտրությամբ, իսկ իրականում այդ գույնը, ամենայն հավանականությամբ, իր ծագմամբ գլխավորապես պարտական է սեռական ընտրությանը։ Մալայան արշիպելագում աճող մի մագլցող արմավենի բարձրանում է ամենաբարձր ծառերի վրա իր զարմանալիորեն հաջող կառուցված կեռիկների օգնությամբ, որոնք հավաքված են ճյուղերի ծայրերին, և այդ հարմարումը անկասկած, կարևոր ծառայություն է մատուցում բույսին․ բայց քանի որ համարյա նույնպիսի կեռիկներ հանդիպում են չմագլցող շատ ծառերի վրա, որտեղ, ինչպես կարելի է դատել ըստ Աֆրիկայի և Հարավային Ամերիկայի փշոտ տեսակների բաշխման, նրանք ծառայում են արածող չորքոտանիներից պաշտպանվելու համար, ապա մեր արմավենու կեռիկները կարող էին սկզբնապես ունենալ նույն նշանակությունը և միայն հետագայում բույսը դառնալով մագլցող, օգտվել է նրանցով և կատարելագործել է նրանց։ Ուրուրի գլխի վրա լերկ մաշկը սովորաբար դիտվում է որպես հարմարեցում լեշը կտցահարելու համար, այդ բացատրությունը գուցե և ճիշտ է, կարող է պատահել, որ այդ առանձնահատկությունն առաջացել է նեխող նյութերի անմիջական ներգործությունից, բայց միայն թե այդ փաստից մենք մեծ զգուշությամբ կկարողանայինք որևէ եզրակացություն անել միշտ մաքուր կերպով սնվող հնդուհավի գլխի վրայի մերկ մաշկի վերաբերմամբ։ Մատղաշ կաթնասունների գանգի կարերը դիտվել են որպես հիանալի մի հարմարում, որը հեշտացնում է ծննդաբերության գործողությունը և անկասկած է, որ նրանք կարող են նրան նպաստել և նույնիսկ անհրաժեշտ լինել, բայց որովհետև այդ կարերը գոյություն ունեն նաև թռչունների և սողունների գանգերի վրա, որոնք միայն դուրս են գալիս ձվերից, ուստի մենք իրավունք ունենք եզրակացնելու, որ այդ հարմարումը բխում է հենց աճման օրենքներից, և միայն հետագայում դրանից օգուտ է ստացվել բարձրակարգ կենդանիների ծննդյան գործողության ժամանակ։
Մենք համարյա ոչինչ չգիտենք յուրաքանչյուր թույլ փոփոխության կամ անհատական տարբերության պատճառի մասին․ դրանում դժվար չէ համոզվել, եթե ուշադրություն դարձնենք մեր ընտանեցված կենդանիների ցեղերի միջև եղած տարբերության վրա զանազան երկրներում, առանձնապես քիչ քաղաքակրթված երկրներում, որտեղ գրեթե չի կիրառվել մեթոդական ընտրությունը։ Զանազան երկրներում վայրենիների պահած անասունները հաճախ ստիպված են իրենց գոյության միջոցների համար պայքարի մեջ մտնել և որոշ չափով ենթարկվել բնական ընտրության, ըստ որում թեթևակի փոփոխված կերտվածք ունեցող անհատները կարող են մեծ հաջողություն ունենալ զանազան կլիմայական պայմաններում։ Եղջերավոր անասունների այն ընդունակությունը, որ նրանք դիմանում են միջատների հարձակմանը, ինչպես նաև թունավոր բույսերի ներգործությանը, կապված է նրանց թույրի հետ, այնպես որ կենդանու գույնը նույնիսկ կենթարկվի բնական ընտրության ներգործությանը։ Մի քանի դիտողներ խորապես համոզված են, որ խոնավ կլիման նպաստում է մազերի աճին, և որ մազերի աճման հետ կախման մեջ է գտնվում նաև եղջյուրների աճը։ Լեռնային ցեղերը սովորաբար տարբերվում են հարթավայրերում տարածված ցեղերից․ լեռնոտ վայրը, ամենայն հավանականությամբ, կարող է ետևի վերջավորությունների ավելի զգալի վարժեցման հետևանքով ազդել նրանց վրա, միգուցե նաև կոնքի ձևի վրա․ հոմոլոգ փոփոխականության օրենքի հետևանքով այդ փոփոխությունը թերևս կարող է անդրադառնալ առջևի վերջավորությունների և գլխի վրա։ Կոնքի ձևն իր հերթին ճնշման հետևանքով կարող է ազդել նաև ձագերի ձևի վրա։ Բարձրադիր վայրերում ուժեղ շնչառությունը կարող է առաջ բերել կրծքի չափերի ավելացում, որի հետ միասին կազմվածքի մեջ կառաջանան էլի նոր հարաբերակցություններ։ Առատ սնման դեպքում նվազած վարժեցման ազդեցությունը թերևս ավելի ևս խոր կերպով է ներգործում ամբողջ կազմվածքի վրա. ըստ երևույթին դրանում, ինչպես ցույց է տվել Նատուզիուսն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 130] իր հիանալի աշխատության մեջ, գտնվում է այն խոր փոփոխությունների գլխավոր պատճառը, որոնց ենթարկվել են խոզերի ցեղերը։ Բայց մեր անտեղյակությունն այնպես խորն է, որ մենք չենք կարող գնահատական տալ փոփոխականության մեզ հայտնի ու անհայտ պատճառների նշանակության մասին։ Ես այս դիտողությունները բերեցի միայն այն բանի համար, որպեսզի ցույց տամ, որ մենք ի վիճակի չենք բացատրելու մեր ընտանի ցեղերի բնորոշ տարբերությունների ծագումը, որոնց վերաբերմամբ, այնուամենայնիվ, բոլորն էլ համաձայն են, որ նրանք ծագել են սովորական ճանապարհով մեկ կամ մի քանի նախնական ձևերից։ Այդ պատճառով էլ մենք առիթ չունենք առանձին նշանակություն տալու իսկական տեսակների միջև եղած նման թույլ տարբերությունների ամենամոտ պատճառների չիմանալուն։
ՈՐՔԱՆ ՃԻՇՏ է ՈՒՏԻԼԻՏԱՐ ՍԿԶԲՈՒՆՔԸ. ԻՆՉՊԵՍ Է ՁԵՌՔ ԲԵՐՎՈՒՄ ԳԵՂԵՑԿՈՒԹՅՈՒՆԸ
Նախորդ դիտողություններն ինձ ստիպում են մի քանի խոսք ասել այն բողոքի առթիվ, որ վերջերս հայտնել են մի քանի բնախույզներ ուտիլիտար ուսմունքի դեմ, որը ենթադրում է, թե կազմվածքում եղած ամեն մի մանրամասնություն մշակվել է բացառապես դրանով օժտված էակի օգտի համար։ Նրանք կարծում են, թե կազմվածքի բազմաթիվ առանձնահատկությունները ստեղծված են բացառապես նրանց գեղեցկության համար, մարդու կամ արարչի իր իսկ զվարճության համար (այս վերջին ենթադրությունը գիտական քննարկման սահմաններից դուրս է գալիս), կամ թե պարզապես բազմազանության համար, այս վերջին ենթադրությունը մենք արդեն առիթ ունեցել ենք քննարկելու։ Այդպիսի պատկերացումը, եթե ճիշտ լիներ, անպայման ճակատագրական կլիներ իմ ուսմունքի համար։ Ես միանգամայն ընդունում եմ, որ կազմվածքի շատ առանձնահատկություններ ոչ մի նշանակություն չունեն իրենց տերերի համար, և գուցե նշանակություն չեն ունեցել նաև նրանց նախնիների համար, բայց այդ բնավ չի ապացուցում, որ նրանք ստեղծվել են բացառապես գեղեցկության կամ բազմազանության համար։ Կասկածի ենթակա չէ, որ փոփոխված պայմանների որոշակի ներգործությունը և փոփոխականության հենց նոր թվարկված բազմազան պատճառները բոլորն էլ թողել են իրենց, թերևս շատ խոր, ազդեցությունը, առանց որևէ առնչություն ունենալու այդ եղանակով իրագործվող օգտի հետ։ Բայց ավելի ևս կարևոր է այն նկատառումը, որ ամեն մի կենդանի էակի կազմվածքի բոլոր գլխավոր գծերն իրենց ծագմամբ պարտական են ժառանգականությանը․ սրանից հետևում է, որ թեև յուրաքանչյուր կենդանի էակ անկասկած հիանալի կերպով հարմարված է բնության մեջ իր գրաված տեղին, բայց և այնպես նրա կազմվածքի բազմաթիվ կողմերը ճիշտ ու անմիջական առնչություն չունեն ներկայիս կենսապայմանների հետ։ Ուրեմն, մենք հազիվ թե կարող ենք ենթադրել, որ լեռնային սագի կամ ճղլակ թռչունի թաղանթավոր ոտները հատկապես օգտակար են այդ թռչուններին․ մենք չենք կարող նաև ենթադրել, որ կապկի ձեռքերում, ձիու առջևի ոտներում, չղջիկի թևում և փոկի մաշկաթաթերում միանման ոսկորների ներկայությունը հատկապես օգտակար է եղել այդ կենդանիներին։ Մենք կարող ենք այդ հանգամանքը համոզվածությամբ վերագրել ժառանգականությանը։ Բայց, առանց որևէ կասկածի, լեռնային սագի կամ ճղլակի նախահոր թաղանթավոր թաթերը նրան նույնքան օգտակար են եղել, ինչքան որ այժմ ապրող թռչուններից ամենից ավելի ջրլողներին։ Ճիշտ այդպես էլ մենք կարող ենք համոզված լինել, որ փոկի նախահայրը ունեցել է ոչ թե լողաթաթ, այլ քայլելու կամ բռնելու համար հարմարված հինգ մատներով թաթ․ մենք կարող ենք նույնիսկ ենթադրել և ավելին, որ կապկի, ձիու և չղջիկի վերջավորություններում եղած զանազան ոսկորները սկզբնապես զարգացել են օգտակարության սկզբունքի հիման վրա, ամենայն հավանականությամբ, ամբողջ դասի որևէ հնագույն ձկնակերպ նախահոր լողաթևի բազմաթիվ ոսկորների թվի կրճատման ճանապարհով։ Հազիվ թե հնարավոր է որոշել, թե երևույթի որ մասը պետք է վերագրել փոփոխականության այնպիսի պատճառների բաժնին, ինչպիսիք են արտաքին պայմանների ազդեցությունը, այսպես կոչված, ինքնակամ փոփոխությունները և աճման բարդ օրենքները։ Բայց, անելով այդ կարևոր բացառությունները, մենք կարող ենք ասել, որ յուրաքանչյուր կենդանի էակի կազմվածքի մեջ եղած բոլոր մանրամասները, ուղղակի կամ անուղղակի կերպով, օգտակար են կամ օգտակար են եղել դրանք ունեցողին։
Ինչ վերաբերում է այն համոզմունքին, որ օրգանական էակները հիանալի են ստեղծված նրա համար, որպեսզի մարդուն զվարճություն պատճառեն, մի համոզմունք, որը ոմանց կարծիքով կտրում է իմ տեսության արմատը, ուստի ես ամենից առաջ պետք է նկատեմ, որ գեղեցկության զգացմունքը, հավանորեն, կախված է խելքի բնույթից՝ անկախ զվարճության առարկային հատուկ որևէ իրական որակից, ինչպես նաև այն, որ գեղեցկության գաղափարը չի կարելի համարել բնատուր և անփոփոխ։ Դրան որպես ապացույց ծառայում է այն փաստը, որ զանազան ռասաների մարդիկ հիանում են կանացի գեղեցկության բոլորովին տարբեր տիպերով։ Եթե գեղեցիկ առարկաներն ստեղծվել են բացառապես մարդու բավականության համար, ապա պետք էր ապացուցել, որ մինչև մարդու երևան գալը, երկրի վրա ավելի քիչ գեղեցկություն է եղել, քան նրա ասպարեզ ելնելուց հետո։ Մի՞թե էոցենի դարաշրջանի Conus և Voluta սքանչելի խեցիները կամ երկրորդային ժամանակաշրջանի նրբագեղ կերտված ամմոնիտներն ստեղծվել են նրա համար, որպեսզի մարդը դարեր անցնելուց հետո կարողանար նրանցով զմայլվել բնապատմական կաբինետներում։ Քիչ կգտնվեն ավելի գեղեցիկ առարկաներ, քան դիատոմների կայծաքարային մանրագույն զրահները, արդյոք մենք կենթադրե՞նք, թե սրանք էլ ստեղծված են նրա համար, որպեսզի կարելի լիներ խիստ ուժեղ կերպով մեծացնող մանրադիտակով նրանց զննել ու նրանցով զմայլվել։ Գեղեցկությունը այդ և այլ շատ դեպքերում ըստ երևույթին բացառապես առաջանում է համաչափ աճումից։ Ծաղիկները համարվում են բնության ամենագեղեցիկ արտադրանքները, բայց նրանք խիստ տարբերվում են կանաչ սաղարթից, և դրանով իսկ գեղեցիկ են միայն նրա համար, որպեսզի իրենց վրա գրավեն միջատների ուշադրությունը։ Ես այդ եզրակացությանը եկել եմ այն անփոփոխ կանոնի հիման վրա, որ երբ ծաղիկը բեղմնավորվում է քամու միջոցով, նա երբեք չի ունենում վառ գույնի պսակ։ Որոշ բույսեր միշտ տալիս են երկու տեսակի ծաղիկներ, մի քանիսը բաց են ու գունավորված, միջատներին գրավող, մյուսները փակ են, նեկտարից զուրկ և միջատները նրանց երբեք չեն այցելում։ Սրանից մենք իրավունք ունենք եզրակացնելու, որ եթե երկրի երեսին միջատներ չլինեին, ապա բույսերը զարդարված չէին լինի հիանալի ծաղիկներով, այլ կտային այնպիսի խղճուկ ծաղիկներ, ինչպիսին մենք տեսնում ենք սոճու, կաղնու, պնդուկենու, հացենու, կամ մեր հացաբույսերի, սպանաղի և եղինջի վրա, որոնք բոլորն էլ բեղմնավորվում են քամու միջոցով։ Կշռադատության նույն եղանակը կիրառելի է նաև պտուղների վերաբերմամբ, որ հասուն ելակը կամ բալը միանման հաճելի են թե՛ աչքի համար և թե՛ համով, որ իլենու վառ գունավորված պտուղը կամ չիչխանի կարմիր հատապտուղներն ինքնըստինքյան գեղեցիկ են, դրա հետ յուրաքանչյուր ոք կհամաձայնի, բայց այդ գեղեցկությունը ծառայում է թռչուններին և գազաններին գրավելու համար, որպեսզի նրանք ուտեն պտուղը և տարածեն պարարտացրած սերմերը։ Ես գալիս եմ այդ եզրակացության այն կանոնի հիման վրա, որը ոչ մի բացառություն չունի, որ պտղի մեջ գտնվող սերմերը (այսինքն օժտված են մսալի կամ հյութալի պատյանով) տարածվում են այն դեպքում, երբ նրանք աչքի են ընկնում իրենց վառ գույնով կամ լինում սպիտակ կամ սև։
Մյուս կողմից, ես հոժարությամբ ընդունում եմ, որ զգալի թվով արուները, ինչպես, օրինակ, մեր բոլոր ամենագեղեցիկ թռչունները, որոշ ձկներ, սողուններ ու կաթնասուններ և հիանալի գունավորված թիթեռների բազմությունը գեղեցիկ են միայն գեղեցկության համար. բայց այդ ձեռք է բերվել սեռական ընտրության միջոցով, այսինքն էգերի կողմից գեղեցիկ արուներ նախընտրելու շնորհիվ, այլ ոչ թե մարդու զվարճության համար։ Նույնը ճիշտ է նաև թռչունների երգեցողության վերաբերմամբ։ Սրանից մենք իրավունք ունենք եզրակացնելու, որ գեղեցիկ գույների և երաժշտական ձայների նկատմամբ մոտավորապես միանման ճաշակ ունի կենդանիների թագավորության զգալի մասը։ Երբ էգերը նույնպես գեղեցիկ գունավորված են, ինչպես արուները, որը առանձնապես հազվադեպ չէ թռչունների և թիթեռների մեջ, պատճառը երևի նրանումն է, որ սեռական ընտրության միջոցով ձեռք բերված գույնը, փոխանակ սահմանափակվելու միայն արուներով, փոխանցվել է երկու սեռերին։ Թե ինչպես գեղեցկության զգացմունքն իր ամենապարզ արտահայտությամբ, այսինքն գույնի, ձևերի և ձայների առաջացրած առանձին բավականության զգացումն առաջին անգամ ծագել է մարդու կամ ամենապարզ կենդանիների մտքի մեջ, դա վերին աստիճանի մութ հարց է։ Նույնպիսի դժվարության ենք հանդիպում նաև այն հարցը քննարկելիս, թե ինչու որոշ համի և հոտառական տպավորություններ հաճելի են, իսկ մյուսները տհաճ։ Երևի սովորությունը այդ բոլոր դեպքերում որոշ դեր է խաղացել, բայց պետք է նաև մի ավելի խոր պատճառ լինի, որը գտնվում է յուրաքանչյուր տեսակի նյարդային սիստեմի հենց կազմության մեջ։
Բնական ընտրությունը ոչ մի կերպ չի կարող որևէ տեսակի մեջ առաջացնել փոփոխություն, որը բացառապես օգտակար է մի այլ տեսակի համար, թեև բնության մեջ ամենուրեք մի տեսակ առավելություն և օգուտ է ստանում մյուս տեսակների կազմվածքից։ Բայց բնական ընտրությունը կարող է առաջացնել և հաճախ առաջացնում է օրգաններ, որոնց նպատակն է ուղղակի վնաս պատճառել այլ կենդանիների, մենք տեսնում ենք օձերի թունավոր ատամներից կամ իխնեվմոնի ձվադրից (ovipositor), որի միջոցով վերջինս ձվեր է ածում այլ միջատների կենդանի մարմինների մեջ։ Եթե կարելի լիներ ապացուցել, որ այս կամ այն տեսակի որևէ օրգանը գոյացել է բացառապես ոչ թե իր, այլ մի այլ տեսակի օգտի համար, ապա այդ դեպքը կխորտակեր իմ տեսությունը, որովհետև նման օրգանի գոյացումը չէր կարող ստացվել բնական ընտրության միջոցով։ Թեև բնապատմության վերաբերյալ աշխատություններում կարելի է այդ իմաստով շատ ցուցումների հանդիպել, բայց ես չեմ գտել ոչ մեկը, որին կարելի լիներ որևէ կշիռ տալ. ընդունում են, օրինակ, որ շառաչող օձը թունավոր ատամներ ունի ինքնապաշտպանության և իր զոհերի ոչնչացման համար, բայց միևնույն ժամանակ որոշ հեղինակներ ենթադրում են, թե նրա շառաչող ապարատը իրեն վնասակար է, որովհետև դրանով նա նախազգուշացնում է իր զոհերին։ Ես գրեթե նույնպիսի հեշտությամբ պատրաստ եմ հավատալու, որ կատուն պատրաստվելով ցատկելու, պտտում է իր պոչը, որպեսզի նախազգուշացնի մահվան դատապարտված մկանը։ Շատ ավելի հավանական է այն ենթադրությունը, որ շառաչող օձը բանեցնում է իր շառաչող ապարատը, կոբրան արձակում է իր հավելուկը, իսկ ակնոցավոր օձը ուռչում է բարձրաձայն սուր ֆշշոցով միայն նրա համար, որպեսզի վախեցնի այն թռչուններին և գազաններին, որոնք, ինչպես հայտնի է, հարձակվում են նույնիսկ ամենաթունավոր օձերի վրա։ Օձերն այնտեղ առաջնորդվում են նույն հիմունքով, որն ստիպում է հավին չռել փետուրները և տարածել թևերը, երբ շունը մոտենում է նրա ճուտերին. բայց այստեղ տեղը չէ երկար կանգ առնելու այն բազմազան եղանակների վրա, որոնց միջոցով կենդանիներն աշխատում են վախեցնել իրենց թշնամիներին։
Բնական ընտրությունը երբեք չի կարող որևէ էակի մեջ առաջացնել որևէ օրգան կամ կառուցվածք, որը նրա համար ավելի վնասակար լիներ, քան օգտակար, որովհետև բնական ընտրությունը ներգործում է բացառապես միայն տվյալ էակի բարօրության համար և այդ բարօրության միջոցով։ Երբեք չի կարող գոյանալ մի օրգան, ինչպես նկատել է Պալեյը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 131] որ ցավ կամ որևէ վնաս պատճառեր իր տիրոջը։ Եթե տանք կազմվածքի յուրաքանչյուր մասի պատճառած բարիքի և չարիքի հանրագումարը, ապա այն ամբողջությամբ վերցրած ավելի օգտակար է լինում։ Եթե ժամանակի ընթացքում փոփոխվող կենսապայմաններում որևէ մարմնամաս վնասակար է դարձել, նա կփոփոխվի, իսկ եթե այդ անհնարին է լինում, այդ դեպքում տվյալ տեսակը կանհետանա, ինչպես որ իսկապես անհետացել են բյուրավոր ուրիշները։
Բնական ընտրությունը ձգտում է յուրաքանչյուր օրգանական էակ դարձնել նույնքան կատարյալ կամ քիչ ավելի կատարյալ, քան նույն երկրի մյուս բնակիչները, որոնց հետ նա մտնում է մրցման մեջ։ Եվ մենք տեսնում ենք, որ այդպիսին է բնության մեջ հասանելի կատարելության չափանիշը։ Տեղական օրգանիզմները, օրինակ, Նոր Զելանդիայում կատարյալ են միմյանց հետ համեմատելիս, բայց այժմ նրանք արագ կերպով տեղի են տալիս Եվրոպայից բերված բույսերի և կենդանիների լեգիոնների գրոհի առջև։ Բնական ընտրության հետևանքով չի կարող բացարձակ կատարելություն ստեղծվել, և իսկապես էլ, որքան մենք ի վիճակի ենք դատել, մենք այդ բանին չենք հանդիպում բնության մեջ։ Մյուլլերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] խոսքերով՝ լույսի աբերացիայի շտկումը ոչ լիովին կատարյալ է նույնիսկ օրգաններից ամենից ավելի կատարյալի մեջ, մարդկային աչքի մեջ։ Հելմհոլցը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 132] որի դատողությունները ոչ ոք չի կարող ժխտել, ամենաուժեղ արտահայտություններով նկարագրելով մարդու աչքի զարմանալի հատկությունները, ավելացնում է այս ուշագրավ խոսքերը. «Այն, ինչ որ մենք կարողացել ենք օպտիկական ապարատի մեջ և ցանցաթաղանթի վրա նրա առաջացրած պատկերի մեջ հայտնագործել անճշտության և անկատարյալության իմաստով, ոչինչ բան է այն անհարիրությունների համեմատությամբ, որոնց մենք հանդիպում ենք զգայությունների բնագավառում։ Կարելի է ասել, որ բնությունը կարծես թե զվարճացել է իրար վրա կիտելով հակասությունները նրա համար, որպեսզի խլի ամեն մի հիմք արտաքին ու ներքին աշխարհների միջև նախորդող ներդաշնակության տեսության համար»։ Եթե մեր բանականությունը մեզ զարմանքի զգացմունք է ներշնչում բնության ներկայացրած անզուգական հարմարումների բազմության հանդեպ, ապա նա էլ ուսուցանում է մեզ, թեև սխալները հավասարապես հնարավոր են թե այս և թե այն ուղղությամբ, որ գոյություն ունեն ավելի նվազ կատարյալ ուրիշ հարմարեցումներ։ Կարո՞ղ ենք արդյոք մենք կատարյալ համարել մեղվի խայթոցը, որը նրա բազմազան թշնամիներից որևէ մեկի դեմ գործադրելիս դուրս չի կարող քաշվել, որովհետև նրա ատամիկները դեպի ետ են շրջված, և դրանով էլ անխուսափելի մահ է պատճառում միջատին, պոկելով վերջինիս փորոտիքը։
Եթե մենք ենթադրենք, որ մեղվի խայթոցը գոյություն է ունեցել նրա հեռավոր նախահոր մոտ որպես շաղափող ատամնավոր մի գործիք, ինչպիսիք հանդիպում են այդ մեծածավալ ընտանիքի բազմաթիվ ներկայացուցիչների մոտ, եթե մենք ընդունենք, որ այն ժամանակից սկսած խայթոցը փոփոխվել է, թեև չի կատարելագործվել իր իսկական նպատակի համար, որ ուժեղացել է նաև այն թույնը, որն սկզբում հարմարված է եղել բոլորովին այլ նպատակի համար, օրինակ գխտորներ առաջացնելու համար, եթե մենք այդ բոլորն ընդունենք, այն ժամանակ գուցե հասկանանք, թե ինչու խայթոցի գործածությունը կարող է այդքան հաճախ ուղեկցվել միջատի մահով, ի վերջո, եթե խայթելու ընդունակությունը օգտակար կլիներ սոցիալական համայնքի համար, նա կհամապատասխաներ բնական ընտրության բոլոր պահանջներին, թեկուզ և մահ պատճառեր այդ համայնքի առանձին անդամներին։ Եթե մենք զարմանում ենք հոտառության իսկապես հրաշալի ուժի վրա, որի շնորհիվ միջատների արուները որոնում, գտնում են էգերին, մենք կարո՞ղ ենք արդյոք նույն չափով հիանալ միայն բազմացման նպատակի համար հազարավոր բոռերի արտադրությամբ, որոնք այլ տեսակետներից միանգամայն անօգուտ են համայնքի համար և որոնց աշխատասեր, բայց ամուլ քույրերը նրանց վերջիվերջո սպանում են։ Թեև, կարող է պատահել, այդ դժվար է, բայց մենք պետք է հիանանք մայր մեղվի բնազդային վայրենի ատելությամբ, որը դրդում է նրան ոչնչացնելու երիտասարդ մայրերին, իր դուստրներին հենց նրանց ծննդյան ժամանակ, կամ թե իրեն ոչնչանալու այդ կռվում, որովհետև դա անկասկածորեն ուղղված է դեպի ամբողջ համայնքի բարօրությանը։ Մայրական սերը կամ մայրական ատելությունը, թեև վերջինս բարեբախտաբար անհամեմատ ավելի հազվադեպ է արտահայտվում, հավասար են բնական ընտրության անողոք սկզբունքի առջև։ Եթե մենք հիանում ենք բազմազան ճարտար հարմարեցումների վրա, որոնց շնորհիվ խոլորձիները և շատ այլ բույսեր բեղմնավորվում են միջատների աջակցությամբ, ապա արդյոք կարո՞ղ ենք մենք միանման կատարյալ համարել մեր սոճիների ծաղկափոշու ամբողջ ամպերի արտադրումը միայն նրա համար, որպեսզի մի քանի փոշեհատիկներ, պատահմամբ քամու աջակցությամբ, հասնեն սերմնաբողբոջներից մեկին։
ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ. ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՏԵՍՈՒԹՅՈՒՆՆ ԸՆԴԳՐԿՈՒՄ է ՏԻՊԻ Եվ ԳՈՅՈՒԹՅԱՆ ՊԱՅՄԱՆՆԵՐԻ ՄԻԱՍՆՈՒԹՅԱՆ ՕՐԵՆՔՆԵՐԸ
Մենք այս գլխում քննարկեցինք մի քանի դժվարություններ ու առարկություններ, որոնք կարող են ներկայացվել մեր տեսության դեմ։ Նրանցից շատերը լուրջ են, բայց ես կարծում եմ, որ քննարկության ժամանակ մեզ հաջողվեց լույս սփռել մի քանի փաստերի վրա, որոնք, անկախ արարչագործական ակտերին հավատալու տեսակետից, միանգամայն անըմբռնելի կլինեին։ Մենք տեսանք, որ տեսակները յուրաքանչյուր տվյալ ժամանակաշրջանում անսահմանորեն փոփոխական չեն լինում և միջակա օղակների երկար շղթայով միմյանց հետ կապված չեն լինում մասամբ այն պատճառով, որ բնական ընտրության պրոցեսը կատարվում է չափազանց դանդաղ կերպով և յուրաքանչյուր տվյալ մոմենտում ներգործում է միայն փոքրաթիվ ձևերի վրա, մասամբ էլ այն պատճառով, որ հենց բնական ընտրության պրոցեսը ենթադրում է նախորդողների և միջակա աստիճանների մշտական փոխարինում և ոչնչացում։ Ներկայումս անընդհատ տարածություն գրավող միմյանց մոտ ազգակից տեսակները հաճախ պետք է առաջանային դեռ այն ժամանակ, երբ այդ տարածությունը անընդհատ չէր և երբ կենսապայմանները չէին փոփոխվել այնպիսի անզգալի աստիճաններով տարածության մի մասից մյուսին անցնելիս։ Երբ երկու տարատեսակներ առաջանում են միևնույն անընդհատ տարածության երկու մարզերում, հաճախ կարող է առաջանալ միջակա մի տարատեսակ, որը հարմարված է միջակա գոտուն, վերևում շարադրված պատճառներով այդ միջակա տարատեսակը սովորաբար գոյություն կունենա ավելի սակավաթիվ անհատներով, քան այն երկու ձևերը, որոնց նա միացնում է. դրա հետևանքը կլինի այն փաստը, որ փոփոխությունների հետագա ընթացքում այս վերջիններն իրենց թվի ավելի շատության շնորհիվ կգտնվեն ավելի բարենպաստ պայմաններում այդ սակավաթիվ միջակա տարատեսակի համեմատությամբ, սովորաբար հաջողությամբ դուրս կմղեն ու, վերջապես, կոչնչացնեն նրան։
Այս գլխում մենք համոզվեցինք, որ պետք է չափազանց շրջահայաց լինել, ենթադրություն անելով, որ ամենատարբեր կենցաղաձևերի միջև անցումներ չեն կարող լինել, մենք համոզվեցինք նրանում, որ չղջիկը կարող էր բնական ընտրության միջոցով առաջանալ սկզբում օդում միայն սահող կենդանուց։ Մենք համոզվեցինք, որ տեսակները նոր կենսապայմաններում կարող են փոփոխել իրենց սովորությունները կամ կարող են ունենալ բազմազան սովորություններ, որոնք երբեմն նման չեն իրենց հետ մերձավոր ազգակից ձևերի սովորություններին։ Յուրաքանչյուր օրգանական էակ ձգտում է ապրել այնտեղ, որտեղ նա կարող էր ապրել, այստեղից էլ մեզ համար հասկանալի դարձավ, թե ինչպես կարող են առաջանալ այնպիսի ձևերը, ինչպես լեռնային սագը թաղանթավոր ոտներով, գետնի վրա ապրող փայտփորիկը, սուզվող կեռնեխը և սուզաբադերի կենցաղաձևն ունեցող մրրկահավերը։
Այն համոզմունքը, որ կատարելության այդպիսի բարձր աստիճանի հասած օրգանը, ինչպիսին աչքն է, կարող էր առաջանալ բնական ընտրության միջոցով, կարող է ում ասես տարակուսանքի մեջ գցել։ Այնուամենայնիվ, որևէ մի օրգանի վերաբերմամբ, եթե մենք գիտենք աստիճանական բարդացումների երկար շարքը, ըստ որում նրանցից յուրաքանչյուրն ինքնըստինքյան օգտակար է, չի կարելի տրամաբանորեն անհնարին համարել, որ շարունակ փոփոխվող կենսապայմաններում տվյալ օրգանը բնական ընտրության միջոցով ձեռք չի բերել կատարելության ամեն մի հնարավոր աստիճան։ Այն դեպքերում, երբ մեզ անհայտ են միջակա կամ անցումային աստիճանները, մենք պիտի շատ շրջահայաց լինենք այն եզրակացության մեջ, թե նրանք երբեք էլ գոյություն չեն ունեցել, որովհետև օրգանների կերպարանափոխության բազմաթիվ դեպքերում մենք տեսնում ենք, թե ինչպիսի զարմանալի ձևափոխություններ հնարավոր են լինում օրգանների գործունեության մեջ։ Արժե հիշել, որ լողափամփուշտը դարձել է օդ շնչող թոքեր։ Միևնույն օրգանը, որը միաժամանակ կատարել է զանազան ֆունկցիաներ, իսկ հետո մասամբ կամ բոլորովին հարմարվել է նրանցից մեկի համար, կամ երկու օրգաններ, որոնք միաժամանակ կատարել են նույն ֆունկցիան, ըստ որում մեկը կատարելագործվել մյուսի աջակցության շնորհիվ, շատ դեպքերում զգալիորեն հեշտացրել են այդպիսի անցումը։
Մենք տեսանք, որ էակների սանդուխքի վրա մեկը մյուսից հեռու կանգնած երկու օրգանիզմներ կարող են ունենալ օրգաններ, որոնք ծառայում են միևնույն նպատակի համար և իրար հետ մոտ նմանություն ունեն, բայց առաջացել են առանձին-առանձին ու միանգամայն անկախ կերպով։ Սակայն այդ օրգանների ավելի ուշադիր հետազոտման ժամանակ համարյա միշտ կարելի է էական տարբերություններ գտնել նրանց կառուցվածքում, և այդ, հասկանալի է, բխում է բնական ընտրության սկզբունքից։ Մյուս կողմից, բնության մեջ արտահայտվող ամենածավալված կանոնն է, որ միևնույն նպատակին կարելի է հասնել բազմազան միջոցներով, այս էլ դարձյալ բխում է նույն մեծ սկզբունքից։
Շատ դեպքերում մեր տեղեկությունները այնքան անբավարար են, որ մենք իրավունք չունենք պնդելու, թե որոշ մարմնամասեր կամ օրգաններ այնքան անկարևոր են տեսակի բարեկեցության համար, որ նրանց կառուցվածքում փոփոխությունները չկարողանային դանդաղ կերպով կուտակվել բնական ընտրության միջոցով։ Բազմաթիվ այլ դեպքերում փոփոխություններն ամենայն հավանականությամբ հանդիսացել են փոփոխականության ու աճման օրենքի ուղղակի արդյունք, անկախ այն բանից, թե նրանք իրենց հետ որևէ օգուտ բերել են, թե ոչ։ Բայց մենք կարող ենք համոզված լինել, որ այդպիսի կառուցվածքները ևս հետագայում, նոր կենսապայմաններում, կարող էին օգտակար դառնալ տվյալ տեսակի համար և ենթարկվել հետագա փոփոխությունների։ Մենք կարող ենք համոզված լինել նաև, որ սկզբում շատ կարևոր մի մարմնամաս հաճախ պահպանվել է (ինչպես, օրինակ, ջրային կենդանու պոչը նրա ցամաքային հետնորդների մոտ), թեև այնպիսի չնչին նշանակություն է ստացել, որ իր այժմյան վիճակում չէր էլ կարող ձեռք բերվել բնական ընտրության միջոցով։
Բնական ընտրությունը չի կարող մի տեսակի մեջ առաջացնել որևէ այնպիսի մի բան, որը բացառապես ծառայի հօգուտ կամ ի վնաս մի այլ տեսակի, թեև նա հեշտությամբ կարող է առաջացնել մասեր, օրգաններ կամ արտաթորանքներ, որոնք շատ օգտակար են, նույնիսկ անհրաժեշտ կամ, ընդհակառակը, չափազանց վնասակար մի այլ տեսակի համար, բայց բոլոր այդպիսի դեպքերում նրանք օգտակար կլինեն իրենց տիրոջ համար։ Յուրաքանչյուր խիտ բնակված երկրում բնական ընտրությունը ներգործում է նրա բնակիչների միջև տեղի ունեցող մրցության շնորհիվ և այդ պատճառով էլ գոյության կռվում հաջողություն է ապահովում կատարելության այն չափանիշին համապատասխան, որը հատուկ է տվյալ երկրին։ Այս պատճառով սովորաբար մի փոքր երկրի բնակիչները մեծ մասամբ ենթարկվում են մի այլ սովորաբար մեծ երկրի բնակիչներին։ Որովհետև մեծ երկրում գոյություն են ունեցել մեծ թվով անհատներ և ավելի շատ բազմազան ձևեր, որոնց միջև մրցությունը եղել է ավելի խիստ, ապա հետևաբար և կատարելության չափանիշն ավելի բարձր աստիճանի է հասել։ Բնական ընտրությունն անպայմանորեն չի հանգում բացարձակ կատարելության և, որքան մենք կարող ենք դատել մեր սահմանափակ ընդունակությունների օգնությամբ, չի կարելի պնդել, որ նա ամենուրեք իրականում կենսագործվել է։
Բնական տեսության հիման վրա մենք պարզ կերպով հասկանում ենք բնապատմության վաղեմի կանոնի՝ «Natura non facit saltum»[49] լրիվ իմաստը։ Այս կանոնը, եթե մենք սահմանափակվենք միայն երկրի արդի բնակիչներով, ոչ լրիվ ճիշտ է, բայց եթե այն տարածենք անցյալ ժամանակներում ապրած թե՛ հայտնի և թե՛ անհայտ էակների վրա, ապա այս տեսության հիման վրա նա պետք է ճիշտ լինի իր ամբողջ խստությամբ։
Հանրաճանաչ է համարվում, որ օրգանական էակներն առաջացել են երկու մեծ օրենքների համաձայն՝ տիպի միասնականության և գոյության պայմանների։ Տիպի միասնականություն ասելով հասկացվում է կազմության մեջ եղած այն հիմնական նմանությունը, որ մենք նկատում ենք մի դասի պատկանող էակների մեջ և որը միանգամայն անկախ է նրանց կենսակերպից. իմ տեսությամբ տիպի միասնականությունը բացատրվում է ծագման միասնականությամբ։ «Գոյության պայմաններ» արտահայտությունը, որի վրա այնքան հաճախ պնդել է հռչակավոր Կյուվիեն,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 133] լիովին ընդգրկվում է բնական ընտրության սկզբունքի կողմից։ Բնական ընտրությունը ներգործում է շնորհիվ այն բանի, որ հարմարեցնում է էակների փոփոխվող մասերը նրանց կյանքի օրգանական և անօրգանական պայմաններին, կամ հարմարեցրել է նրանց անցած ժամանակներում, ըստ որում այդ հարմարումները շատ դեպքերում պահպանվել են ուժեղացրած վարժեցման միջոցով կամ, ընդհակառակը, վարժեցման բացակայությամբ, գտնվել են արտաքին պայմանների ուղղակի ներգործության ազդեցության տակ և, համենայն դեպս, ենթարկվել են աճման ու փոփոխականության հայտնի օրենքներին։ Ուրեմն, գոյության պայմանների օրենքը իրականում հանդիսանում է բարձրագույն օրենք, որովհետև նա նախկին փոփոխությունների և հարմարեցումների ժառանգման միջոցով իր մեջ է առնում նաև տիպի միասնականության օրենքը։
ԳԼՈՒԽ VII։ ԶԱՆԱԶԱՆ ԱՌԱՐԿՈՒԹՅՈՒՆՆԵՐ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՏԵՍՈՒԹՅԱՆ ԴԵՄ
ԵՐԿԱՐԱԿԵՑՈՒԹՅՈՒՆ։— ՁԵՎԱՓՈԽՈՒԹՅՈԻՆՆԵՐԸ ՄԻՇՏ ՉԷ, ՈՐ ՄԻԱԺԱՄԱՆԱԿ ԵՆ ԱՌԱՋԱՆՈՒՄ։ ՁԵՎԱՓՈԽՈՒԹՅՈՒՆՆԵՐ, ՈՐՈՆՔ, ԸՍՏ ԵՐԵՎՈՒՅԹԻՆ, ՈՒՂՂԱԿԻ ՕԳՈՒՏ ՉԵՆ ԲԵՐՈՒՄ։— ԱՌԱՋԱԴԻՄԱԿԱՆ ԶԱՐԳԱՑՈՒՄ։— ՖՈՒՆԿՑԻՈՆԱԼ ՏԵՍԱԿԵՏԻՑ ՔԻՉ ԿԱՐԵՎՈՐ ՀԱՏԿԱՆԻՇՆԵՐՆ ԱՄԵՆԻՑ ԱՎԵԼԻ ԿԱՅՈՒՆ ԵՆ։— ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ԵՆԹԱԴՐՅԱԼ ԹԵՐՈՒԹՅՈՒՆԸ ԿԱՌՈՒՑՎԱԾՔԻ ՕԳՏԱԿԱՐ ԱՌԱՆՁՆԱՀԱՏԿՈՒԹՅՈՒՆՆԵՐԻ ՍԿԶԲՆԱԿԱՆ ՍՏԱԴԻԱՆԵՐԻ ԲԱՑԱՏՐՈՒԹՅԱՆ ՀԱՄԱՐ։— ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՄԻՋՈՑՈՎ ՕԳՏԱԿԱՐ ԱՌԱՆՁՆԱՀԱՏԿՈՒԹՅՈՒՆՆԵՐ ՁԵՌՔ ԲԵՐԵԼՈՒՆ ԽՈՉԸՆԴՈՏ ՀԱՆԴԻՍԱՑՈՂ ՊԱՏՃԱՌՆԵՐԸ։— ԿԱՌՈՒՑՎԱԾՔԻ ԱՍՏԻՃԱՆԱԿԱՆ ՓՈՓՈԽՈՒԹՅՈՒՆԸ, ՈՐԻՆ ՈՒՂԵԿՑՈՒՄ է ՖՈՒՆԿՑԻԱՆԵՐԻ ՓՈՓՈԽՈՒԹՅՈՒՆԸ։— ՄԻԵՎՆՈՒՅՆ ԴԱՍԻ ՆԵՐԿԱՅԱՑՈՒՑԻՉՆԵՐԻ ԽԻՍՏ ՏԱՐԲԵՐՎՈՂ ՕՐԳԱՆՆԵՐԸ, ՈՐՈՆՔ ԾԱԳՈՒՄ ԵՆ ՄԻ ԸՆԴՀԱՆՈՒՐ ԱՂԲՅՈՒՐԻՑ։— ԱՅՆ ՀԻՄՔԵՐԸ, ՈՐՈՆՔ ԻՆՁ ՀԱՐԿԱԴՐՈՒՄ ԵՆ ԺԽՏԵԼՈՒ ԽԻՍՏ ԵՎ ՀԱՆԿԱՐԾԱԿԱՆ ՓՈՓՈԽՈՒԹՅՈՒՆՆԵՐԻ ԳՈՅՈՒԹՅՈՒՆԸ։
Այս գլուխը կնվիրեմ իմ տեսակետների դեմ առաջ քաշված խառը բնույթի զանազան առարկությունների քննարկմանը, որովհետև այդ ճանապարհով կարող են ավելի լավ պարզաբանվել որոշ բաներ մինչև այժմ քննարկվածից։ Բայց անօգուտ կլիներ քննության առնել այդ բոլոր առարկությունները, որովհետև նրանցից շատերը արել են այնպիսի գրողներ, որոնք նեղություն չեն կրել ըմբռնելու բուն խնդիրը։ Այսպես, օրինակ, գերմանական մի հայտնի բնախույզ պնդում է, որ իմ տեսության ամենաթույլ կողմն այն է, որ ես բոլոր օրգանական էակները դիտում եմ որպես չափազանց անկատար. մինչդեռ իրականում ես ասել եմ, որ նրանք բոլորը այնքան էլ կատարյալ չեն, ինչպիսին կարող էին լինել իրենց շրջապատող պայմանների նկատմամբ, և այդ ապացուցվում է նրանով, որ երկրագնդի շատ կետերում տեղական բազմաթիվ ձևերն իրենց տեղերը զիջել են ներխուժած օտարերկրացիներին։ Բացի դրանից, եթե օրգանական էակները լինեին էլ մի որոշ ժամանակ լիովին հարմարված շրջապատող պայմաններին, նրանք չեն կարող այդպես մնալ այդ պայմանների փոփոխվելուց հետո, եթե իրենք ևս չեն փոփոխվի համապատասխան ձևով, և ոչ ոք, իհարկե, վիճելի չի կարող համարել այն, որ յուրաքանչյուր երկրի ֆիզիկական պայմանները, ինչպես նաև նրա բնակիչների և՛ քանակությունը, և՛ բնույթը շատ փոփոխություններ են կրել։
Վերջերս մի քննադատ, պճնվելով մաթեմատիկական ճշգրտությամբ, պնդում էր, թե երկարակեցությունը շատ օգտավետ է բոլոր տեսակներին. այնպես որ, ով հավատում է բնական ընտրությանը, «պետք է իր ծննդաբանական ծառը կառուցի» այնպիսի ձևով, որ բոլոր հետնորդները ավելի երկարակյաց լինեն իրենց նախնիներից։ Մի՞թե մեր քննադատը չի կարողանում ըմբռնել այն, ար երկամյա բույսը կամ մեր կենդանիներից մեկը կարող է ընկնել ցուրտ կլիմայի մեջ և այնտեղ ոչնչանալ յուրաքանչյուր ձմեռ, և սակայն, բնական ընտրության միջոցով ձեռք բերված օգտակար հատկությունների շնորհիվ, տարեցտարի ապրել իր սերմերի կամ ձվերի օգնությամբ։ Ռեյ Լանկեստերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 134] վերջերս քննարկել է այդ հարցը և, որքան խնդրի բարդությունը հնարավորություն է տալիս որոշակի դատողություն կազմելու, նա գալիս է այն եզրակացության, որ երկարակեցությունը սովորաբար հարաբերակցության մեջ է գտնվում օրգանական սանդուխքի վրա յուրաքանչյուր տեսակի ունեցած դրության հետ, ինչպես նաև վերարտադրության պրոցեսի ու ընդհանուր գործունեության վրա օրգանիզմի ծախսի աստիճանի հետ։ Իսկ այդ հատկությունները հավանորեն շատ մեծ չափով կախված են բնական ընտրությունից։
Բերվել է նաև այն փաստարկը, թե ինչպես որ եգիպտական այն կենդանիներից ու բույսերից ոչ մեկը, որոնց մասին մենք որևէ բան գիտենք, չի փոփոխվել վերջին երեք կամ չորս հազարամյակների ընթացքում, ճիշտ այդպես էլ հավանորեն չի փոփոխվել նրանցից ոչ մեկը և ոչ մի տեղ աշխարհում։ Բայց, ըստ Ջ. Հ. Լյուիսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 135] դիտողության, փաստարկման այդ եղանակը ոչինչ չի ապացուցում, որովհետև հնագույն ընտանի ցեղերը, որ պատկերված են եգիպտական հուշարձաններում կամ զմռսված են, շատ մոտ են կամ նույնիսկ նման են նրանց, որոնք ապրում են ներկայումս, իսկ մինչդեռ բոլոր բնախույզներն ընդունում են, որ այդ ցեղերն առաջացել են իրենց նախասկզբնական տիպերի ձևափոխման միջոցով։ Շատ կենդանիներ, որոնք առանց փոփոխվելու մնացել են սառցադաշտային շրջանից սկսած, կարող էին անհամեմատ ավելի զարմանալի օրինակ ծառայել, որովհետև նրանք ենթարկվել են կլիմայի մեծ փոփոխությունների ու տեղափոխվել են մեծ հեռավորությունների վրա, մինչդեռ Եգիպտոսում վերջին մի քանի հազարամյակների ընթացքում կենսապայմանները, որքան մենք գիտենք, մնացել են բոլորովին միանման։ Սառցադաշտային շրջանի ժամանակից սկսած փոփոխությունների աննշանության կամ նույնիսկ լիակատար բացակայության փաստը, որոշ հաջողությամբ կարելի է շրջել նրանց դեմ, ովքեր հավատում են զարգացման բնածին ու անհրաժեշտ օրենքին, և այդ փաստը բոլորովին անզոր է բնական ընտրությանը, կամ ամենահարմարվածի գերապրելուն վերաբերող ուսմունքի հանդեպ, մի ուսմունքի, որը ենթադրում է, որ երբ պատահաբար առաջանում են օգտակար հատկություն ունեցող ձևափոխություններ կամ անհատական տարբերություններ, ապա նրանք պահպանվում են, բայց այդ իրագործվում է միայն որոշ բարենպաստ հանգամանքներում։
Նշանավոր հնէաբան Բրոննը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 136] գրքիս իր գերմաներեն թարգմանության վերջում հարցնում է, թե ինչպե՞ս բացատրել բնական ընտրության տեսակետով այն, որ տարատեսակը և նրան ծնունդ տվող տեսակը կարող են ապրել կողք կողքի։ Եթե նրանք երկուսն էլ հարմարվել են թեթևակի տարբերվող կենսական շրջադրության կամ պայմաններին, այդ դեպքում նրանք կարող են միասին ապրել և եթե մի կողմ թողնենք պոլիմորֆ ձևերը, որոնց փոփոխականությունն ըստ երևույթին առանձին բնույթ է կրում, և բոլոր ժամանակավոր ձևափոխությունները, ինչպիսիք են մեծությունը, ալբինիզմը և այլն, ապա ավելի կայուն տարատեսակները, որքան ես կարող եմ դատել, սովորաբար լինում են զանազան ապրելավայրերում, օրինակ, բարձունքներում կամ հարթավայրերում, չոր կամ խոնավ տեղերում բնակվողներ։ Այնուհետև շատ թափառող և հեշտությամբ միմյանց հետ խաչաձևվող կենդանիների տարատեսակները, ըստ երևույթին, սովորաբար կապված են տարբեր վայրերի հետ։
Բրոննը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 136] հաստատամտորեն պնդում է նաև, որ առանձին տեսակները մեկը մյուսից երբեք չեն տարբերվում առանձին հատկանիշներով, այլ միշտ շատ հատկանիշներով. նա հարցնում է, թե ինչպե՞ս է միշտ լինում, որ օրգանիզմի մի քանի մասերը, վարիացիաների և բնական ընտրության միջոցով միաժամանակ ձևափոխվում են։ Բայց այս դեպքում ոչ մի անհրաժեշտություն չկա ենթադրելու, որ որևէ օրգանիզմի բոլոր մասերը փոփոխվել են միաժամանակ։ Ամենազարմանալի ձևափոխությունները, որոնք հիանալի կերպով հարմարված են որևէ նպատակի համար, ինչպես առաջ արդեն նկատված է, կարող են ձեռք բերվել, թեկուզև թույլ, հաջորդական շեղումների միջոցով՝ սկզբում մի մասում, հետո մյուսում, բայց քանի որ նրանք փոխանցվում են բոլորը միասին, ուստի և մեզ վրա այնպիսի տպավորություն են թողնում, որ կարծես թե նրանք միասին են առաջացել։ Վերոհիշյալ առարկությանը ամենալավ պատասխան կարող են ծառայել, ասենք, այն ընտանի կենդանիները, որոնք ձևափոխվել են մարդու կամքով, որը նրանց ընտրել է որևէ հատուկ նպատակով։ Նայենք արշավաձիուն և ծանրաքաշ ձիուն, կամ բարակ շանը և դոգ գամփռին (mastiff)։ Նրանց բոլոր մարմնակազմությունը և նույնիսկ նրանց մտավոր կերտվածքը ձևափոխվել են, մինչդեռ, եթե մենք հնարավորություն ունենայինք հետամտելու նրանց կերպափոխումների պատմության ընթացքում եղած յուրաքանչյուր ստադիային,— իսկ վերջին ստադիաներին հետամտել հնարավոր է,— մենք չէինք տեսնի խոշոր և միաժամանակ տեղի ունեցած փոփոխություններ, այլ միայն թեթև ձևափոխություններ ու կատարելագործություններ սկզբում մի մասում, ապա և մյուս մասում։ Նույնիսկ այն դեպքում, երբ մարդը կիրառել է միայն որևէ մի հատկանիշի ընտրություն,— որի ամենալավ օրինակ կարող են ծառայել մեր մշակովի բույսերը,— մենք անփոփոխ կերպով նկատում ենք, որ եթե մի մասը, լինի դա ծաղիկ, պտուղ կամ տերևներ, շատ ուժեղ կերպով փոփոխվել է, ապա նաև գրեթե բոլոր մյուս մասերը թեթև ձևափոխություններ են կրել։ Դա մասամբ կարելի է բացատրել հարաբերակցական աճման սկզբունքով, մասամբ էլ այսպես կոչված ինքնակամ վարիացիայով։
Անհամեմատ ավելի լուրջ առարկություն է առաջադրել Բրոննը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 136] իսկ վերջերս նաև Բրոկան,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 137] այն է՝ որ շատ հատկանիշներ, ըստ երևույթին, ոչ մի օգուտ չեն բերում իրենց տերերին և այդ պատճառով չէին կարող իրենց վրա կրել բնական ընտրության ազդեցությունը։ Բրոննը մատնանշում է, օրինակ, նապաստակների և մկների տարբեր տեսակների ականջների ու պոչերի երկարությունը, շատ կենդանիների ատամների արծնապակու բարդ ծալքերը և բազմաթիվ համանման օրինակներ։ Բույսերի վերաբերմամբ այդ հարցը պարզաբանել է Նեգելին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 138] իր հիանալի հետազոտության մեջ։ Նա ընդունում է, որ բնական ընտրությունը շատ բան է արել, բայց պնդում է, թե բույսերի ընտանիքները մեկը մյուսից տարբերվում են գլխավորապես մորֆոլոգիական հատկանիշներով, որոնք, ըստ երևույթին, բոլորովին կարևոր չեն տեսակների բարեկեցության համար։ Դրա համար էլ նա հավատում է, որ տեսակներն ունեն առաջադիմելու և կատարելագործվելու բնածին հակում։ Նա մատնանշում է բջիջների դասավորությունը հյուսվածքներում և տերևներինը՝ առանցքային մասերի վրա որպես օրինակներ, երբ բնական ընտրությունը չի կարողացել ներգործել։ Դրան կարելի է ավելացնել նաև ծաղկի մասերի թիվը, սերմնաբողբոջների դիրքը, սերմերի ձևը, երբ վերջինս ոչ մի օգուտ չի բերում սերմնավորման ժամանակ և այլն։
Վերոհիշյալ առարկությունը շատ լուրջ է։ Այնուամենայնիվ, մենք պետք է նախ և առաջ, չափազանց զգույշ լինենք, երբ ձգտում ենք որոշել, թե այս կամ այն տեսակի համար կառուցվածքի ինչպիսի գծերը օգտակար են այժմ, կամ օգտակար են եղել առաջ։ Երկրորդ, միշտ պետք է հիշել, որ երբ ձևափոխվում է մի մաս, նույնն է պատահում նաև մյուսների հետ՝ լիովին չպարզաբանված պատճառների հետևանքով, ինչպիսիք են, օրինակ, սննդի ուժեղացած կամ նվազած առհոսումը դեպի մի մասը, մասերի փոխադարձ ճնշումը, ընդ որում ավելի առաջ զարգացած մասը ճնշող ազդեցություն է գործում իրենից հետո զարգացողի վրա և այլն, ինչպես նաև այլ պատճառների հետևանքով, որոնք առաջ են բերում հարաբերակցության բազմաթիվ և խորհրդավոր ու մեզ համար բոլորովին անհասկանալի փաստեր։ Այդ բոլոր գործոնները, համառոտության համար, կարելի է միասին միացնել «աճման օրենքներ» ընդհանուր անվան տակ։ Երրորդ, մենք պետք է ընդունենք կենսապայմանների փոփոխության ուղղակի և որոշակի ներգործությունը և այսպես կոչված ինքնակամ վարիացիաները, որոնց մեջ պայմանների բնույթը միանգամայն ստորադաս դեր է կատարում։ Բողբոջների վարիացիան, ինչպես, օրինակ, սովորական վարդենու վրա թավոտ վարդի կամ դեղձենու վրա նեկտարինների (հարթ դեղձերի) երևալը, ներկայացնում են ինքնակամ վարիացիաների լավ օրինակներ, բայց նույնիսկ այդ դեպքերում,— եթե մենք մտաբերենք, թե ինչպիսի ուժեղ ազդեցություն է գործում թույնի մի կաթիլը բարդ գղտորների առաջացման վրա,— մենք չենք կարող լիովին վստահ լինել, որ վերոհիշյալ վարիացիաները հյութի բնույթի մեջ մի որևէ տեղական փոփոխության արդյունք չեն, որը կախված է պայմանների փոփոխություններից։ Յուրաքանչյուր անհատական թեթև տարբերության համար պետք է որևէ իրական պատճառ լինի, նույնպես էլ պատահաբար առաջացող ավելի ուժեղ արտահայտված վարիացիների համար[50] և եթե անհայտ պատճառը կներգործի համառ կերպով, ապա համարյա անկասկած է, որ տվյալ տեսակի բոլոր անհատները կփոփոխվեն նման ձևով։
Այս գրքի նախորդ հրատարակություններում, ինչպես թվում է ինձ այժմ, ես հարկ եղած չափով չեմ գնահատել, թե որքան հաճախակի և կարևոր են ինքնակամ փոփոխականությունից կախված ձևափոխությունները։ Բայց անհնարին է այդ պատճառով բացատրել կառուցվածքի այն առանձնահատկությունների անթիվ բազմությունը, որոնք այնքան լավ հարմարված են յուրաքանչյուր տեսակի կենսակերպին։ Այդ ինձ նույնքան քիչ հավանական է թվում, որքան և այն, որ դրա միջոցով կարելի է բացատրել արշավաձիու կամ բարակ շան լավ հարմարված ձևի առաջանալը, որոնք այնպիսի զարմանք են հարուցել, նախկին բնախույզների մտքերում, քանի դեռ լավ չէր ըմբռնվել մարդու ձեռքով կատարվող ընտրության սկզբունքը։
Ավելորդ չի լինի նախորդ դիտողություններից մի քանիսը պարզաբանել օրինակներով։ Զանազան մասերի և օրգանների ենթադրյալ անօգտակարության մասին հազիվ թե հարկ կա նշելու, որ նույնիսկ բարձրակարգ և ամենից ավելի ծանոթ կենդանիներն ունեն կառուցվածքի առանձնահատկություններ, որոնք այնքան բարձր զարգացման են հասել, որ ոչ ոք չի կասկածում նրանց կարևորության մասին, սակայն նրանց նշանակությունը մինչև այժմ պարզված չէ, կամ պարզվել է միայն վերջերս։ Բրոննը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 136] մատնանշում է մկների զանազան տեսակների ականջների ու պոչի երկարությունը որպես կառուցվածքի թեպետև չնչին, բայց այնպիսի տարբերությունների օրինակներ, որոնք ոչ մի հատուկ, նշանակություն չունեն․ այդ առթիվ ես կարող եմ հիշատակել, որ ըստ դոկտոր Շեբլի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 139] հետազոտությունների, սովորական մկան արտաքին ականջը չափազանց հարուստ է նյարդերով, այնպես որ նա անկասկած ծառայում է որպես շոշափելիքի օրգան. հետևաբար, ականջների երկարությունը հազիվ թե բոլորովին անկարևոր հատկանիշ լինի։ Մենք հիմա կտեսնեք նաև, որ պոչը մի քանի տեսակների համար ծառայում է որպես շատ օգտակար բռնիչ օրգան, և այդ դեպքում նրա դերը մեծ կախման մեջ է գտնվում երկարությունից։
Ինչ վերաբերում է բույսերին, որոնցով ես բացառապես կզբաղվեմ հետագայում՝ Նեգելիի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 138] հոդվածի առթիվ, ապա բոլորը, իհարկե, կհամաձայնվեն, որ խոլորձիների ծաղիկները ներկայացնում են կառուցվածքի բազմաթիվ հետաքրքրական մանրամասնություններ, որոնց վրա մի քանի տարի առաջ կնայեին որպես պարզ մորֆոլոգիական առանձնահատկությունների վրա, որոնք ոչ մի հատուկ ֆունկցիա չունեն, մինչդեռ ներկայումս հայտնի է, որ նրանք խոշորագույն նշանակություն ունեն միջատների օգնությամբ տեսակների բեղմնավորման համար և հավանորեն ձեռք են բերվել բնական ընտրության միջոցով։ Մինչև վերջին ժամանակները ոչ ոքի մտքով չէր անցնի, որ դիմորֆ և տրիմորֆ բույսերի առէջքների և վարսանդների անհավասար երկարությունը և նրանց դասավորությունը կարող են որևէ նշանակություն ունենալ, իսկ այժմ մենք գիտենք, որ այդ հենց այդպես էլ է։
Բույսերի որոշ խմբերում բոլոր տեսակների սերմնաբողբոջները ուղիղ են կանգնած, մյուսներինը՝ կախված են, իսկ մի քանի սակավաթիվ բույսերի միևնույն սերմնարանում մի սերմնաբողբոջը գրավում է կանգուն դիրք, իսկ մյուսը՝ կախված։ Սերմնաբողբոջների այդ դիրքերը առաջին հայացքից զուտ մորֆոլոգիական հատկանիշներ են թվում, այսինքն չունեն ոչ մի ֆիզիոլոգիական նշանակություն, բայց դոկտոր Հուկերն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] ինձ հաղորդում է, որ միևնույն սերմնարանի մեջ երբեմն բեղմնավորվում են միայն վերին սերմնաբողբոջները, իսկ այլ դեպքերում միայն ստորինները, և նա ենթադրություն է անում, որ դա հավանորեն կախված է այն ուղղությունից, որով ծաղկափոշու խողովակները մտնում են սերմնարանի մեջ։ Եթե այդ այդպես է, ուրեմն սերմնաբողբոջների դիրքը,— եթե անգամ միևնույն սերմնարանում նրանցից մեկը ուղիղ կանգնած է, իսկ մյուսը՝ կախված,— կարող է լինել դիրքի մեջ տեղի ունեցած այնպիսի թեթև որոշ շեղումների ընտրության հետևանք, որոնք նպաստել են բեղմնավորմանը և սերմերի ստացմանը։
Որոշ բույսեր, որոնք պատկանում են զանազան ընտանիքների, սովորաբար առաջացնում են երկու ձևի ծաղիկներ՝ մի քանիսը բաց, սովորական կառուցվածքով, մյուսները՝ փակ և թերզարգացած։ Ծաղիկների այդ երկու ձևերը երբեմն զարմանալի կերպով տարբերվում են միմյանցից իրենց կառուցվածքով, այնուամենայնիվ կարելի է տեսնել, թե ինչպես նրանք միևնույն բույսի վրա աստիճանաբար փոխակերպվում են մեկը մյուսին։ Սովորական բաց ծաղիկները կարող են միմյանց հետ խաչաձևվել և այդպիսով ապահովում են այն օգտակարությունը, որն անշուշտ ստացվում է խաչաձևման ժամանակ։ Սակայն փակ և թերզարգացած ծաղիկները[51] ակներևորեն շատ մեծ նշանակություն ունեն, որովհետև նրանք ապահովաբար տալիս են որոշ քանակությամբ սերմեր՝ ծախսելով զարմանալի քիչ քանակությամբ ծաղկափոշի։ Այդ երկու ձևի ծաղիկները, ինչպես հենց նոր մատնանշվեց, հաճախ խիստ տարբերվում են միմյանցից իրենց կառուցվածքով, թերի ծաղիկների պսակաթերթերը համարյա միշտ կազմված են միայն թերաճիկներից, և փոշեհատիկների տրամագիծը փոքրացած է։ Ononis columnae-ի միմյանց հաջորդող տասը առէջքներից հինգը սաղմնային վիճակում են, իսկ Viola-ի առէջքներից երեքը գտնվում են այդպիսի վիճակում, այնպես որ միայն երկուսն են պահպանում իրենց նորմալ ֆունկցիան և այն էլ շատ փոքր չափով։ Հնդկական մի մանուշակի (անունն ինձ անհայտ է, որովհետև այդ բույսերն ինձ մոտ լիովին զարգացած ծաղիկներ երբեք չեն տվել) երեսուն փակ ծաղիկներից վեցի մեջ փոխանակ հինգ բաժակաթերթի, որ նորմալ է, զարգացել էին երեքը։ Malpighiaceae-ի մի ենթաընտանիքում, ըստ Ա. դը Ժյուսսյեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 140] հետազոտությունների, փակ ծաղիկները ավելի ևս ձևափոխված են, որովհետև բաժակաթերթերի դիմացի հինգ առէջքները բոլորն էլ ապաճած են և զարգանում է միմիայն վեցերորդը, որը գտնվում է պսակաթերթի դիմաց (այս վերջին առէջքը այդ տեսակի սովորական ծաղիկներում չի լինում). սռնակը զարգացած չէ, և սերմնարանների թիվը երեքի փոխարեն վերածվել է երկուսի։ Եվ ահա թեև բնական ընտրությունը, անտարակույս, կարող էր խոչընդոտ լինել այդ ծաղիկներից մի քանիսի բացվելուն և պակասեցնել ծաղկափոշու ծախսումը, որովհետև նրա նախկին քանակը ավելորդ կլիներ փակ ծաղիկներում, սակայն հազիվ թե վերոհիշյալ հատուկ ձևափոխություններից որևէ մեկը ծագել է հենց այդ ձևով, այլ հավանորեն աճման օրենքների հետևանք է եղել (այդ թվում հաշվելով մասերի ֆունկցիոնալ անգործունեությունը) ծաղկափոշու քանակի աստիճանական կրճատման և ծաղիկների փակման դեպքում։
Աճման օրենքների ներգործության կարևորության պատշաճ գնահատումն այնքան անհրաժեշտ է, որ ես կբերեմ այլ տեսակի մի քանի լրացուցիչ օրինակներ ևս, այսինքն միևնույն մասի կամ օրգանի մեջ եղած այնպիսի տարբերությունների օրինակներ, որոնք կախված են տվյալ բույսի վրա նրանց հարաբերական դիրքի տարբերություններից։ Իսպանական շագանակենու և մի քանի փշատերև ծառերի տերևների բաժանման անկյունները, Շախտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 141] հետազոտությունների համաձայն, միանման չեն համարյա հորիզոնական և ուղիղ կանգնած ճյուղերի վրա։ Սովորական փեգենայի և մի քանի այլ բույսերի վրա մի ծաղիկը, սովորաբար կենտրոնականը կամ գագաթնայինը, առաջինն է բացվում և ունենում է հնգական բաժակաթերթ ու պսակաթերթ և հնգաբաժան սերմնարան, մինչդեռ նույն բույսի վրա մնացած ծաղիկները քառանդամ են։ Բրիտանական Adoxa-ի գագաթնային ծաղկի բաժակը սովորաբար երկբլթականի է և մնացած օրգանները քառանդամ են, մինչդեռ շրջապատող ծաղիկների բաժակները սովորաբար եռաբլթակ են, իսկ մնացած մասերը հնգանդամ են։ Բարդածաղիկներից ու հովանոցավորներից (Compositae, Umbelliferae) շատերի, ինչպես նաև մի քանի այլ բույսերի եզրային ծաղիկների պսակներն անհամեմատ ավելի զարգացած են, քան ներքին ծաղիկներինը և այդ, ըստ երևույթին, հաճախ կապ ունի բազմացման գործարանների ատրոֆիայի հետ։ Առավել ևս հետաքրքրական է մի փաստ, որը մենք շոշափել ենք առաջ, այն է, որ եզրային սերմիկները կամ սերմերը երբեմն խիստ տարբերվում են ներքիններից իրենց ձևով, գույնով և այլ հատկանիշներով։ Carthamus-ի [սաֆլորի] և մի քանի այլ բարդածաղիկների միայն կենտրոնական սերմիկներն են ունենում փուփուլիկներ, իսկ Hyoseris-ի մոտ միևնույն գլխիկը տալիս է երեք տարբեր ձևի սերմիկներ։ Մի քանի հովանոցավորների (Umbelliferae,) արտաքին սերմերը, ըստ Տաուշի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 142] ունենում են ուղիղ էնդոսպերմ, իսկ կենտրոնականները՝ ծռված էնդոսպերմ, մինչդեռ դը-Կանդոլն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] այլ տեսակների մոտ այդ հատկանիշը դիտել է որպես սիստեմատիկայի մեջ շատ մեծ կարևորություն ունեցող։ Պրոֆեսոր Բրաունը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 143] հիշատակում է Fumariaceae-ի մի սեռի մասին, որի մեջ ողկույզի ստորին մասում ծաղիկները տալիս են ձվաձև կողավոր, միասերմ կաղիններ, իսկ վերին մասում՝ նիզակաձև, երկփեղկանի և երկսերմ պատիճներ։ Այդ բոլոր դեպքերում, եթե չհաշվենք լավ զարգացած եզրային ծաղիկները, որոնք օգտակար են նրանով, որ գրավում են միջատների ուշադրությունը, բնական ընտրությունը չէր կարող, որքան ես ըմբռնում եմ, ոչ մի դեր խաղալ, կամ միայն ստորադաս դեր է խաղացել։ Այդ բոլոր ծխափոխությունները մասերի հարաբերական դիրքի և փոխադարձ ազդեցության արդյունք են, և հազիվ թե կարելի է կասկածել այն բանի վրա, որ եթե տվյալ բույսի վրա բոլոր ծաղիկները և տերևները գտնվեին միևնույն արտաքին ու ներքին պայմանների ներգործության տակ, ինչպես այդ լինում է որոշ դիրք գրավող ծաղիկների ու տերևների նկատմամբ, ապա նրանք բոլորն էլ կձևափոխվեին միանման կերպով։
Այլ բազմաթիվ դեպքերում մենք նկատում ենք, որ կառուցվածքում եղած ձևափոխությունները, որոնց բուսաբանները սովորաբար մեծ նշանակություն են վերադրում, վերաբերում են միայն տվյալ բույսի վրա գտնվող մի քանի ծաղիկների կամ պատահում են միևնույն պայմաններում կողք կողքի աճող զանազան բույսերի վրա։ Քանի որ այդ ձևափոխությունները, ըստ երևույթին, առանձին ոչ մի օգուտ չեն բերում բույսերին, ուստի բնական ընտրությունը չէր կարող նրանց վրա ազդեցություն գործել։ Դրանց պատճառը մեզ բոլորովին անհայտ է, մենք չենք կարող դրանք նույնիսկ վերագրել, ինչպես օրինակների նախորդ խմբում, փոխադարձ դիրքի նման մի որևէ մերձավոր գործոնի ազդեցությանը։ Կբերեմ միայն մի քանի օրինակներ։ Միևնույն բույսի վրա թե՛ տետրամեր և թե՛ պենտամեր ծաղիկների հանդիպելը այնքան սովորական դեպք է, որ ես օրինակներ ցույց տալու կարիք չունեմ, բայց քանի որ թվական վարիացիաները համեմատաբար հազվադեպ են, երբ մասերը քիչ են, ուստի ես կարող եմ հիշատակել, որ Paraver bracteatum-ի ծաղիկները, ինչպես հաղորդում է դը-Կանդոլը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] ունենում են կամ երկու բաժակաթերթիկ և չորս պսակաթերթիկ (կակաչների սովորական տիպը), կամ թե երեք բաժակաթերթիկ և վեց պսակաթերթիկ։ Ծաղկաբողբոջում պսակաթերթիկների դասավորման եղանակը խմբերի մեծամասնության համար շատ կայուն մորֆոլոգիական հատկանիշ է հանդիսանում, բայց պրոֆեսոր Ազա-Գրեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 68] հաղորդում է, որ Mimulus սեռի մի քանի տեսակների մոտ բողբոջակազմությունը նույնքան հաճախ կառուցված է ըստ Rhinanthideae տիպի, որքան ըստ Antirrhinideae տիպի, որպիսի խմբի էլ պատկանում է հիշված սեռը։ Օգյուստ Սենտ Իլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 144] բերում է հետևյալ օրինակները. Zanthoxylon-ը պատկանում է Rutaceae ընտանիքի մեջ այն խմբին, որն աչքի է ընկնում իր անբաժան սերմնարանով, բայց միևնույն բույսի վրա կամ նույնիսկ միևնույն հուրանի մեջ կարելի է գտնել ինչպես մեկ, այնպես էլ երկբաժան սերմնարան ունեցող ծաղիկներ. Helianthemum սեռի մեջ սերմնատուփիկը նկարագրվել է թե որպես միաբուն և թե որպես եռաբուն, իսկ H. mutabile-ի մոտ «une lame, plus ou moin large, s’etend entre le pericarpe et le placente»[52]։ Դոկտոր Մաստերսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 145] Saponaria officinalis-ի ծաղիկներում տեսել է ինչպես եզրային, այնպես էլ կենտրոնական սերմնակիրի օրինակներ։ Վերջերս Սենտ-Իլերը Gomphia oleaeformis-ի տարածման հարավային սահմանի մոտ գտել է երկու ձևեր, որոնց նա սկզբում համարել է անկասկածելի երկու տեսակներ, բայց հետո տեսել է, որ նրանք աճել են մեկ արմատից, և այն ժամանակ նա ավելացնում է. «voila donc dans un même individu des boges et un style qui se rattachent tantôt à un axe vertical, et tantôt à un ginobase»[53]։
Այսպիսով, մենք տեսնում ենք, որ բույսերի մորֆոլոգիական շատ փոփոխություններ կարող են կախման մեջ դրվել աճման օրենքներից, և մասերի փոխազդեցությունից՝ անկախ բնական ընտրությունից։ Բայց կանգնելով Նեգելիի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 138] ուսմունքի այն տեսակետի վրա, որ վերաբերում է կատարելագործման կամ առաջադիմական զարգացման բնածին հակումին և գործ ունենալով փոփոխությունների այդ խիստ արտահայտված օրինակների հետ, կարելի՞ է արդյոք ասել, որ բույսերին մենք գտնում ենք դեպի զարգացման բարձր աստիճանը առաջ ընթանալու մոմենտում։ Ընդհակառակը, արդեն այն փաստից, որ մեր կողմից քննարկվող մասերը միևնույն բույսի վրա խիստ կերպով տարբերվում են կամ ձևափոխվում, ես պետք է եզրակացնեմ, որ նման ձևափոխությունները, որքան էլ նրանք կարևոր լինեն մեր դասակարգումների համար, շատ չնչին կարևորություն ունեն հենց բույսերի համար։ Հազիվ թե կարելի է ասել, որ օրգանիզմը, ձեռք բերելով որևէ անօգուտ մաս, բարձրանում է բնության սանդուխքով, և իրոք, եթե մենք օգնության էլ կանչենք մի որևէ սկզբունք՝ բացատրելու համար թերզարգացած փակ ծաղիկների վերևում նկարագրված օրինակը, ապա այդ կլինի հետագնաց շարժման, այլ ոչ թե առաջընթաց շարժման սկզբունք, այդպես է նաև շատ մակաբույծ և թերզարգացած կենդանիների նկատմամբ։ Մենք չգիտենք, թե մատնանշված յուրահատուկ ձևափոխության պատճառն ինչ է, բայց եթե այդ պատճառը ներգործեր գրեթե միանման ձևով երկար ժամանակի ընթացքում, ապա հետևանքն էլ, կարելի է կարծել, կլիներ գրեթե միանման, իսկ այդ դեպքում տվյալ տեսակի բոլոր անհատները կձևափոխվեին միանգամայն միանման ձևով։
Նկատի առնելով այն, որ վերոհիշյալ հատկանիշները կարևոր չեն տեսակի բարեկեցության համար, նրանց մեջ դրսևորվող բոլոր թեթև վարիացիաները բնական ընտրության ազդեցության տակ չեն կուտակվի ու մեծանա։ Երբ կառուցվածքի որոշ առանձնահատկություններ, որոնք զարգացել են երկարատև ընտրության միջոցով, դադարում են օգուտ բերել տեսակին, այդ դեպքում նրանք սովորաբար դառնում են փոփոխական, ինչպես մենք այդ տեսնում ենք թերաճ օրգանների օրինակի վրա. դա բացատրվում է նրանով, որ նրանք արդեն այլևս չեն կրում ընտրության հենց նույն ուժի կանոնավորող ազդեցությունը։ Բայց երբ օրգանիզմի բնույթի և շրջապատող պայմանների ազդեցության տակ առաջացել են այդպիսի ձևափոխություններ, որոնք կարևոր չեն տեսակի բարեկեցության համար, նրանք կարող են փոխանցվել գրեթե նույն ձևով, որը և հավանորեն հաճախ է տեղի ունեցել, այլ տեսակետներից ձևափոխված բազմաթիվ հաջորդներին։ Կաթնասունների, թռչունների կամ սողունների մեծամասնության համար չէր կարող առանձնապես կարևոր լինել, թե արդյոք ծածկվա՞ծ են մազերով, փետուրներով կամ թեփուկներով, բայց և այնպես մազերը ժառանգաբար փոխանցվել են համարյա բոլոր կաթնասուններին, փետուրները՝ բոլոր թռչուններին և թեփուկները՝ բոլոր իսկական սողուններին։ Երբ կառուցվածքի առանձնահատկությունները, ինչպիսիք էլ նրանք լինեն, ընդհանուր են ազգակից շատ ձևերի համար, մենք նրանց տալիս ենք բարձր սիստեմատիկական նշանակություն և այդ պատճառով հաճախ ենթադրում ենք, թե նրանք չափազանց կարևոր կենսական նշանակություն ունեն տեսակի համար։ Ուստի ես հակված եմ կարծելու, որ մորֆոլոգիական տարբերությունները, որոնց մենք կարևորություն ենք տալիս, ինչպիսիք են, օրինակ, տերևների դասավորությունը, ծաղկի կամ սերմնարանի մասերի թիվը, սերմնաբողբոջների դիրքը և այլն, շատ դեպքերում երևացել են սկզբում որպես անկայուն վարիացիաներ, որոնք վաղ թե ուշ դարձել են հաստատուն՝ օրգանիզմ ի բնույթի և շրջապատող պայմանների շնորհիվ, ինչպես նաև առանձին անհատների փոխադարձ խաչաձևման հետևանքով, բայց ոչ թե բնական ընտրության հետևանքով։ Իրոք՝ քանի որ այդ մորֆոլոգիական հատկանիշները չեն ազդում տեսակի բարեկեցության վրա, ապա նրանց մեջ տեղի ունեցած ամեն մի թեթև շեղում չէր կարող որոշվել կամ կուտակվել այդ վերջին գործոնի օգնությամբ։ Այդպիսով, մենք գալիս ենք մի տարօրինակ եզրակացության, որ տեսակների համար կենսական ոչ մեծ նշանակություն ունեցող հատկանիշները միևնույն ժամանակ շատ մեծ կարևորություն ունեն սիստեմատիկայի համար, բայց ինչպես մենք հետո կտեսնենք, երբ կքննարկենք դասակարգման գենետիկական սկզբունքները, այդ այնքան էլ տարօրինակ չէ, ինչպես կարող է թվալ առաջին հայացքից։
Թեև մենք ոչ մի վստահելի ապացույց չունենք, որ օրգանական աշխարհում գոյություն ունի առաջադիմական զարգացման բնածին հակում, սակայն նա, ինչպես ես աշխատել եմ ցույց տալ չորրորդ գլխում, բնական ընտրության անընդմիջվող ներգործության անհրաժեշտ հետևանքն է։ Եվ իրոք՝ կազմվածքի բարձրության ամենալավ որոշումը, որ մինչև հիմա տրվել է, մասերի մասնագիտացման կամ դիֆերենցման աստիճանն է, իսկ բնական ընտրությունը հենց այդ նպատակին է հանգեցնում այն պատճառով, որ այդ դեպքում առանձին մասերը հնարավորություն են ստանում ավելի մեծ հաջողությամբ կատարելու իրենց ֆունկցիաները։
Հայտնի կենդանաբան միստեր Ստ. Գեորգ Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] վերջերս ամփոփել է այն բոլոր առարկությունները, որ երբևիցե ես և ուրիշները արել ենք այն ձևի բնական ընտրության դեմ, ինչ ձևով որ այն արտահայտել ենք միստեր Ուոլլեսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] և ես, և հիանալի հմտությամբ ու համոզարարությամբ պաշտպանել է իր շարադրանքը օրինակներով։ Այդ ձևով կառուցված լինելով, նրանք ուժեղ տպավորություն են թողնում, իսկ որովհետև միստեր Մայվարտը բոլորովին մտադրություն չունի իր եզրակացություններին հակառակ փաստեր ու կշռադատումներ բերելու, ուստի ընթերցողը, որը կցանկանար այդ հարցին երկու կողմից նայել, նվազագույն հնարավորություն էլ չունի այդ անելու։ Քննարկելով առանձին դեպքեր, միստեր Մայվարտը կանգ չի առնում օրգանների աստիճանաբար ավելացող գործածության և անգործածության հետևանքների վրա, որը ես համարել եմ վերին աստիճանի կարևոր հանգամանք և որը ես պարզաբանել եմ «Կենդանիների և բույսերի փոփոխությունն ընտանեցման ազդեցության տակ» աշխատությանս մեջ ավելի լրիվ կերպով, քան այդ արել է, որքան ես գիտեմ, որևէ մեկ ուրիշը։ Նմանապես էլ նա հաճախ հավատացնում է, թե ես փոփոխականությանը նշանակություն չեմ տալիս բնական ընտրությունից անկախ, մինչդեռ հենց նոր հիշատակածս աշխատության մեջ ես այդ ուղղությամբ հավաքել եմ ավելի մեծ քանակությամբ հաստատուն կերպով ապացուցված տվյալներ, քան դրանք կարելի է գտնել որևէ այլ ինձ հայտնի աշխատության մեջ։ Իմ կարծիքը կարող է վստահության արժանի չլինել, բայց ուշադիր կերպով ընթերցելով միստեր Մայվարտի գիրքը և համեմատելով նրա յուրաքանչյուր բաժինը այդ նույն խնդրի մասին իմ ասածի հետ, ես երբեք ինձ այնպես խորապես համոզված չեմ զգացել այն եզրակացությունների ընդհանուր ճշմարտության մեջ, որոնց ես հանգել եմ այստեղ, թեև բնականաբար, այդպիսի խրթին հարցում ես կարող էի առանձին դեպքերում ընկնել նույնիսկ ոչ աննշան սխալի մեջ։
Միստեր Մայվարտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] բոլոր առարկությունները կամ քննարկված են, կամ քննարկվելու են այս հատորում։ Միակ նոր կետը, որը կարծես շփոթեցրել է մի քանի ընթերցողների, այն է, «որ բնական ընտրությունը չի կարող բացատրել կառուցվածքի օգտակար առանձնահատկությունների սկզբնական ստադիաները»։
Այդ հարցը սերտորեն կապված է առանձնահատկությունների աստիճանական ուժեղացման հարցի հետ, որին հաճախ ուղեկցում է օրգանի աշխատանքի փոփոխությունը, օրինակ՝ լողափամփուշտի փոխարկվելը թեքերի, որն արդեն քննարկվել է նախորդ գլխի երկու բաժիններում։ Այնուամենայնիվ ես այստեղ քիչ թե շատ մանրամասնորեն կքննարկեմ Մայվարտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] բերած օրինակներից մի քանիսը, ընտրելով նրանցից ամենից ավելի բնորոշները, որովհետև տեղի սղությունը ինձ թույլ չի տալիս բոլորը քննարկելու։
Ընձուղտն իր բարձր հասակով, շատ երկար վզով, առջևի ոտներով, գլխով և լեզվով հիանալի կերպով հարմարված է ծառերի վերին ճյուղերից տերևներ պոկելու համար։ Դրա շնորհիվ նա կարողանում է իրեն կեր հայթայթել միևնույն երկրում ապրող այլ սմբակավոր կենդանիների, այսպես ասած, բնագավառի սահմաններից դուրս, և այդ կարող է նրան մեծ առավելություն տալ երաշտների ժամանակաշրջաններում։ Հարավ-ամերիկական եղջերավոր անասուն նյատայի վրա իր հերթին կարելի է տեսնել, որ կառուցվածքի նույնիսկ փոքրիկ առանձնահատկությունները այդպիսի չոր ժամանակաշրջաններում կենդանիների պահպանման տեսակետից մեծ տարբերության պատճառ են հանդիսանում։ Այդ անասունը ուրիշների նման կարող է կերակրվել խոտով, բայց ստորին ծնոտի դուրս ցցվածքի պատճառով չի կարողանում հաճախակի կրկնվող անանձրև ժամանակաշրջաններում կերակրվել ծառերի ճյուղերով, եղեգով և այլն, որոնցով կերակրվելուն են անցնում սովորական եղջերավոր անասունները և ձիերը, դրա հետևանքով այդպիսի ժամանակաշրջաններում նյատաները ոչնչանում են, եթե տերերը չեն կերակրում նրանց։ Նախքան միստեր Մայվարտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] առարկություններին անցնելը՝ օգտակար է մի անգամ ևս բացատրել, թե բնական ընտրությունը ինչ ձևով է իր ազդեցությունը թողնում սովորական դեպքերում։ Մարդը փոփոխել է մի քանի կենդանիների, ձեռք չտալով նրանց կառուցվածքի այս կամ այն առանձնահատկություններին, այլ պարզապես պահպանելով ու բազմացնելով, կամ ամենաարագավազ անհատների, ինչպես, օրինակ, արշավաձին ու բարակ շունը ստեղծելիս, կամ բազմացնելով հաղթող թռչուններին՝ կռվան աքլորներ ստեղծելիս։ Ճիշտ այդպես էլ բնական պայմաններում նոր ծնունդ առնող ընձուղտի դեպքում ամենաբարձրահասակ անհատները, որոնք մեկ կամ երկու դյույմով ավելի բարձր էին մյուսներից, հաճախ կկարողանային պահպանվել երաշտների ժամանակաշրջանում, կեր որոնելու համար թափառելով ամբողջ երկրում։ Որ մի տեսակի անհատները հաճախ քիչ տարբերվում են մեկը մյուսից մարմնամասերի հարաբերական երկարությամբ, կարելի է գտնել բնապատմական շատ աշխատություններում, որտեղ բերված են մանրազնին կերպով կատարված չափումներ։ Չափերի այդ աննշան տարբերությունը, որը պայմանավորված է աճման և փոփոխականության օրենքներով, տեսակների մեծամասնության համար նշանակություն չունի։ Բայց այլ բան էր նոր ծնունդ առնող ընձուղտի նկատմամբ, եթե ուշադրության առնենք նրա հավանական կենսակերպը, որովհետև այն անհատները, որոնց որևէ կամ մի քանի զանազան մարմնամասերը ավելի երկար էին, քան սովորականները, ընդհանրապես պետք է գերապրեին։ Խաչաձևվելիս նրանք պետք է թողնեին կամ կառուցվածքի նույն առանձնահատկություններով օժտված կամ միևնույն ուղղությամբ փոփոխվելու հակում ունեցող կենդանիներ, մինչդեռ այդ տեսակետից ավելի քիչ նպաստավոր կերպով կազմակերպված անհատները պետք է ամենից ավելի ենթարկվեին ոչնչացման։
Այստեղից մենք տեսնում ենք, որ ոչ մի անհրաժեշտություն չկա ընտրելու առանձին զույգեր, ինչպես այդ անում է մարդը սիստեմատիկ ձևով ցեղեր ստեղծելու ժամանակ, բնական ընտրությունը և՛ պահպանում է, և՛ դրանով իսկ ջոկում է բոլոր ավելի բարձր անհատներին, տալով նրանց լիակատար հնարավորություն խաչաձևվելու, և նպաստում է բոլոր ավելի ցածր անհատների ոչնչացմանը։ Եթե այդ պրոցեսը, որը բոլորովին համապատասխանում է նրան, որ ես անվանել եմ մարդու կողմից կատարվող անգիտակցական ընտրություն, երկար է տևում, որին անկասկած շատ էական կերպով ուղեկցում է որոշ օրգանների գործածության աստիճանական ավելացման ժառանգական փոխանցումը, ապա ինձ վերին աստիճանի հավանական է թվում, որ սովորական սմբակավոր կենդանին կարող է այդպիսով դառնալ ընձուղտ։
Դրա դեմ մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] բերում է երկու առարկություն։ Մեկն այն է, որ կենդանու չափերի աճող մեծությունը հավանականորեն պետք է առաջ բերի նրա սպառած սննդի քանակության ավելացում, և նա համարում է «չափազանց կասկածելի, թե արդյոք սննդի պակասության ժամանակաշրջաններում դրանից բխող անբարենպաստ պայմաններն ավելի կարևոր են, քան նրանց հակառակ բարենպաստ պայմանների ազդեցությունը»։ Բայց քանի որ ընձուղտները իրականում մեծ թվով գոյություն ունեն հարավային Աֆրիկայում և քանի որ հենց այստեղ բազմաթիվ են աշխարհիս ամենախոշոր, ցուլից ավելի մեծ անտիլոպներից մի քանիսը, հապա էլ ինչո՞ւ մենք պետք է կասկածենք, որ այստեղ առաջներում, որքանով այդ վերաբերում է մեծությանը, գոյություն են ունեցել անցողիկ ձևեր, որոնք նույնպես, ինչպես որ այժմ, ենթարկվել են դաժան երաշտներին։ Իհարկե, նոր ծնունդ առնող կենդանու ընձուղտի համար կարող էր որոշ առավելություն ներկայացնել այն ընդունակությունը, որի շնորհիվ մեծացող հասակի յուրաքանչյուր ստադիայում կենդանին հայթայթում էր այն կերը, որը երկրի մյուս չորքոտանի սմբակավոր կենդանիների համար անմատչելի էր մնում։ Մենք չպետք է մոռանանք նաև այն, որ հասակի ավելացումը պաշտպանություն էր համարյա բոլոր գիշատիչ գազաններից, բացի առյուծից, իսկ նրա նկատմամբ երկար վիզը,— իսկ որքան ավելի երկար է, այնքան ավելի լավ,— կարող էր ծառայել, ըստ միստեր Չոնսեյ Ռայտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 146] դիտողության, ինչպես հսկողական աշտարակ։ Այդ պատճառով, ինչպես ասում է սըր Ս. Բեկերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 147] ընձուղտին ավելի դժվար է գաղտագողի մոտենալ, քան մի որևէ այլ կենդանու։ Նա իր երկար վիզը գործածում է նաև հարձակման ու պաշտպանության համար, ուժեղ կերպով շարժելով գլուխը, որը զինված է կոճղանման եղջյուրներով։ Յուրաքանչյուր տեսակի պահպանումը հազվագյուտ դեպքում է որոշվում միայն մի որևէ առավելությամբ, իսկ սովորաբար բոլոր՝ թե՛ խոշոր և թե՛ մանր առավելությունների ամբողջությամբ։
Մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] հարցնում է (և դրանումն է նրա երկրորդ առարկությունը), արդյոք իսկապե՞ս այդքան մեծ է բնական ընտրության ազդեցությունը, և եթե բարձր նստած տերևները պոկելու ընդունակությանը այդքան մեծ առավելություններ է տալիս, ապա ինչո՞ւ մյուս չորքոտանիները, բացի ընձուղտից և քիչ չափով էլ ուղտից, գուանակոյից և լամայից, չեն ձեռք բերել երկար վիզ ու բարձր հասակ։ Կամ թե չէ՝ ինչո՞ւ այդ խմբի կենդանիներից ոչ մեկը չի ձեռք բերել երկար կնճիթ։ Հարավային Աֆրիկայի վերաբերմամբ, որտեղ առաջ ընձուղտի բազմաթիվ հոտեր էին ապրում, պատասխանը դժվար չէ և ամենից ավելի լավ կարող է տրվել համապատասխան օրինակով։ Անգլիայի ամեն մի արոտավայրում, որտեղ միայն աճում են ծառեր, նրանց ստորին ճյուղերը կարծես քշտված կամ խուզված են մինչև մի որոշ մակարդակի վրա, որովհետև նրանց կրծում են ձիերը և եղջերավոր անասունները։ Այստեղ ի՞նչ առավելություն կարող էր ունենալ, օրինակ, քիչ ավելի երկար վիզ ունեցող ոչխարը։ Յուրաքանչյուր շրջանում կենդանիների որևէ ցեղ գրեթե ապահովաբար կարող է մյուսներից ավելի բարձր սաղարթը պոկել, և գրեթե նույն չափով էլ հավանական է, որ միայն այդ ցեղն էլ կարող էր ունենալ ավելի երկար վիզ՝ շնորհիվ բնական ընտրության ազդեցության և օրգանի շարունակ ավելացող գործածության։ Հարավային Աֆրիկայում ակացիայի և այլ ծառերի բարձր ճյուղերից տերևներ պոկելու գործում մրցում կարող է տեղի ունենալ միայն ընձուղտների միջև, այլ ոչ թե ընձուղտների և այլ սմբակավորների միջև։
Թե ինչու աշխարհի այլ երկրներում ապրող զանազան կենդանիները ձեռք չեն բերել ոչ երկար վիզ, ոչ էլ կնճիթ, չի կարելի վստահությամբ ասել, բայց այդ հարցին պատասխան սպասելը, նույնքան անհիմն է, որքան որ այն հարցին պատասխան սպասելը, թե ինչու մարդկության պատմության մեջ որևէ իրադարձություն տեղի է ունեցել մի երկրում և չի պատահել մի այլ երկրում։ Մենք չգիտենք այն պայմանները, որոնք որոշում են յուրաքանչյուր տեսակի թիվը և տեղը, և չենք կարող նույնիսկ ենթադրել, թե կառուցվածքում ինչպիսի փոփոխություններ կարող են բարենպաստ լինել նոր երկրում՝ տեսակի բազմացման համար։ Սակայն ընդհանրապես կարելի է կարծել, որ երկար վզի ու կնճիթի զարգացման հետ առնչություն ունեն զանազան պատճառներ։ Ծառի բարձր գագաթից տերևներ վերցնելու ընդունակությունը (առանց մագլցելու, որին բոլորովին հարմարված չեն սմբակավոր կենդանիները) ենթադրում է մարմնի զգալի չափով ավելացած մեծություն, իսկ մենք գիտենք, որ որոշ երկրներ զարմանալիորեն քիչ ունեն խոշոր կաթնասուններ, օրինակ Հարավային Ամերիկան, չնայած իր բնության շքեղությանը, ընդհակառակը, հարավային Աֆրիկան դրանցով չափից դուրս առատ է։ Թե ինչու է այդպես, մենք չգիտենք, ինչպես նաև չգիտենք այն, թե ինչու ուշ երրորդային ժամանակաշրջանը անհամեմատ ավելի բարենպաստ է եղել նրանց գոյության համար, քան այժմյան ժամանակը։ Ինչպիսին էլ ուզում է լինեն այդ պատճառները, բայց մենք տեսնում ենք, որոշ մարզեր և որոշ ժամանակաշրջաններ մյուսներից անհամեմատ ավելի բարենպաստ են այնպիսի խոշոր չորքոտանիների զարգացման համար, ինչպիսին է ընձուղտը։
Այն հանգամանքը, որ կենդանու կառուցվածքի որևէ առանձնահատկություն կարող է մի ուղղությամբ շատ մեծ զարգացման հասնել, համարյա անխզելիորեն կապված է այն բանի հետ, որ զանազան մյուս մասերը նույնպես պիտի փոփոխվեն և դրվեն առաջինին համապատասխան դրության մեջ։ Թեև մարմնի յուրաքանչյուր մասը փոփոխվում է աստիճանաբար, սակայն սրանից չի հետևում, որ անհրաժեշտ մասերը միշտ կփոփոխվեն հարկավոր ուղղությամբ և մինչև հարկ եղած աստիճանի։ Մեր ընտանի կենդանիների զանազան տեսակների վերաբերմամբ մենք գիտենք, որ նրանց մարմնի մասերը փոփոխվում են զանազան ուղղությամբ և զանազան չափերով և որ որոշ տեսակներ անհամեմատ ավելի շատ հակում ունեն փոփոխվելու, քան մյուսները։ Նույնիսկ այն դեպքում, եթե առաջանում են օգտակար շեղումներ, ապա դրանից չի հետևում, որ բնական ընտրությունը միշտ կարող է ազդեցություն գործել նրանց վրա և առաջացնել կառուցվածքի այնպիսի առանձնահատկություն, որը բարենպաստ կլիներ տեսակի համար։ Այսպես, օրինակ, եթե որևէ երկրում ապրող անհատների թիվը մեծ չափով կախված է գիշատիչ կենդանիներից, արտաքին կամ ներքին մակաբույծներից և այլն, որոնք ոչնչացնում են նրանց, որ ըստ երևույթին, իրոք հաճախ է լինում, այդպիսի դեպքում բնական ընտրությունը կարող է կամ միայն թույլ ազդեցություն գործել, կամ թե նրա ազդեցությունը կեր հայթայթման համար ծառայող որևէ մասի փոփոխության վրա խիստ կասեցվում է։ Վերջապես, բնական ընտրության պրոցեսը շատ դանդաղ է, և միևնույն բարենպաստ պայմանները պետք է շատ երկար շարունակվեն, որպեսզի նրա ազդեցությունը դրսևորվի նկատելի կերպով։ Բացառելով այդպիսի ամենաընդհանուր ձևի բացատրությունները, մենք չենք կարող գտնել մեկ ուրիշը, որը կպատասխաներ այն հարցին, թե ինչու մի քանի երկրներում սմբակավոր չորքոտանիները ձեռք չեն բերել շատ երկար վիզ կամ այլ առանձնահատկություններ, որոնք օգնում են տերևներ պոկելու ծառերի բարձր ճյուղերից։
Նախորդին նման առարկություններ արել են շատ հեղինակներ։ Բացի հենց նոր նշված ընդհանուր պատճառներից, յուրաքանչյուր դեպքում ամենայն հավանականությամբ նաև զանազան այլ պատճառներ համատեղ մասնակցել են բնական ընտրության միջոցով կառուցվածքի առանձնահատկություններ ձեռք բերելուն, որոնք, կարելի է կարծել, բարենպաստ են եղել որոշ տեսակների համար։ Մի հեղինակ հարցնում է, թե ինչո՞ւ ջայլամը ձեռք չի բերել թռչելու ընդունակություն։ Բայց րոպեական խորհրդածումը բավական է, որպեսզի հասկանանք, թե սննդի ինչպիսի հսկայական քանակություն անհրաժեշտ կլիներ, որ անապատի այդ թռչունին ուժ տար իր ծանր մարմինը օդում տեղափոխելու համար։ Օվկիանոսային կղզիների վրա ապրում են չղջիկներ և փոկեր, բայց ոչ ցամաքային կաթնասուններ, իսկ քանի որ այդ չղջիկներից մի քանիսը ներկայացնում են առանձին տեսակներ, ապա նրանք պետք է երկար ժամանակի ընթացքում գրաված լինեին իրենց այժմյան մարզերը։ Այդ պատճառով սըր Չ. Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] հարց է տալիս և սպասում է ստանալ նրա որոշ չափով հիմնավոր պատասխանը, թե ինչո՞ւ փոկերը և չղջիկները այդ կղզիների վրա չեն տվել ցամաքային կյանքի համար պիտանի հաջորդներ։ Բայց փոկերն անխուսափելիորեն պետք է սկզբում դառնային զգալի մեծության ցամաքային գիշատիչ կենդանիներ, իսկ չղջիկները՝ վերերկրյա միջատակեր կենդանիներ. առաջինների համար որս չպիտի գտնվեր, իսկ չղջիկներին կերակուր պիտի դառնային վերերկրյա միջատները, բայց վերջիններս արդեն հանդիսանում են գերադասելի որս սողունների և թռչունների համար, որոնք առաջիններն են բնակվում և առատությամբ լինում օվկիանոսային կղզիների մեծամասնության վրա։ Կառուցվածքի մեջ տեղի ունեցող հաջորդական փոփոխությունները յուրաքանչյուր ստադիայում փոփոխվող տեսակի համար օգտակար կարող են լինել միայն որոշ պայմաններում։ Բուն վերերկրյա կենդանին պատահականորեն կեր ճարելով ծանծաղ ջրում, հետո գետերում ու լճերում վերջ ի վերջո կարող էր այնքան փոխարկվել ջրային կենդանու, որ կապրեր բաց օվկիանոսում։ Սակայն փոկերը օվկիանոսային կղզիների վրա չէին կարող գտնել նպաստավոր պայմաններ աստիճանաբար վերերկրյա ձևի փոխարկվելու համար։ Իսկ չղջիկները, ինչպես արդեն ասվել է, թևեր ձեռք են բերել հավանորեն սկզբում ճյուղից ճյուղ օդում ճախրելով, այսպես կոչված թռչող սկյուռների նման, թե՛ թշնամիներից ազատվելու և թե՛ վայր չընկնելու համար. բայց երբ արդեն ձեռք էր բերվել թռչելու ընդունակությունը, նա այլևս չէր փոխարկվի ճյուղից ճյուղ Ճախրելու ընդունակության, որը, համենայն դեպս, վերոհիշյալ նպատակների համար ավելի քիչ օգտակար է։ Իհարկե, չղջիկների թևերը թռչունների թևերի նման կարող էին կամ զգալիորեն կրճատվել, կամ անգործածությունից բոլորովին կորցվել, բայց այդ դեպքում անհրաժեշտ կլիներ, որ այդ կենդանիները նախ և առաջ ձեռք բերեին գետնի վրա արագ վազելու ընդունակություն բացառապես ետևի ոտների օգնությամբ, որպեսզի կարողանային մրցել վերերկրյա թռչունների և այլ կենդանիների հետ. իսկ այդպիսի փոփոխության համար չղջիկը զարմանալիորեն քիչ պիտանի է։ Այդ ենթադրություններն արված են բացառապես նրա համար, որպեսզի ցույց տրվի, որ կառուցվածքի անցումային առանձնահատկությունները, որ բարենպաստ են յուրաքանչյուր ստադիայում, շատ բարդ բնույթ ունեն, և որ զարմանալի ոչինչ չկա, եթե որևէ անցողիկ ձև չի հանդիպում այս կամ այն դեպքում։
Վերջապես, շատ հեղինակներ դրել են այս հարցը, թե ինչո՞ւ որոշ կենդանիների մտավոր ընդունակություններն ավելի են զարգացած, քան մյուսներինը, մինչդեռ այդպիսի զարգացումը պետք է օգտակար լիներ բոլորի համար։ Ինչո՞ւ կապիկները ձեռք չեն բերել մարդու մտածողության ուժը։ Այդ կարելի է վերագրել զանազան պատճառների, բայց որովհետև նրանք բոլորը վերածվում են կռահումների և նրանց հարաբերական հավանականությունը չի կարելի գնահատել, ուստի անօգուտ է դրա վրա կանգ առնել։ Վերջին հարցին խիստ որոշակի պատասխան սպասել չի կարելի, որովհետև ոչ ոք չի կարող լուծել նույնիսկ մի ավելի պարզ հարց, թե ինչո՞ւ վայրենիների երկու ցեղերից մեկը հասել է քաղաքակրթության ավելի բարձր աստիճանի, քան մյուսը, իսկ դա հավանորեն ենթադրում է ուղեղային գործունեության մեծացում։
Այժմ դառնանք միստեր Մայվարտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] մյուս առարկություններին։ Միջատները հաճախ, որպես պաշտպանողական առանձնահատկություններ, ձեռք են բերում նմանություն զանազան առարկաների հետ, օրինակ, կանաչ կամ թափված տերևների, չոր ճյուղերի, քարաքոսերի կտորների, ծաղիկների, փշերի, թռչունների աղբի և այլ խմբերի պատկանող կենդանի միջատների հետ. այս վերջին նմանությանը ես հետագայում դեռ կվերադառնամ։ Այդ նմանությունը հաճախ զարմանալիորեն մոտ է լինում և, չսահմանափակվելով գույնով, տարածվում ձևի և նույնիսկ շարժուձևի վրա։ Թրթուրները, որոնք չոր ճյուղի նման անշարժ կերպով կախվում են թփուտից, որի վրա կերակրվում են, այդպիսի նմանության հիանալի օրինակ են ծառայում։ Այնպիսի առարկաների, ինչպես թռչունների աղբն է, նմանվելու դեպքերը հազվագյուտ են և բացառիկ։ Այդ առթիվ մ-ր Մայվարտը նկատում է հետևյալը. «Քանի որ Դարվինի տեսության համաձայն գոյություն ունի անսահման փոփոխելիության մշտական ձգտում և քանի որ նոր սկզբնավորվող փոքրագույն փոփոխությունները տեղի են ունենում բոլոր ուղղություններով, ուստի նրանք պետք է ձգտեն չեզոքացնել մեկը մյուսին և հենց սկզբից առաջացնել այնպիսի անկայուն փոփոխություններ, որ դժվար է ու նույնիսկ անհնարին է ըմբռնել, թե ինչպես անսահմանորեն փոքրիկ սկզբնավորությունների անսահման տատանումները երբևիցե կհանգեն տերևի, եղեգի կամ այլ առարկայի հետ նմանության և այնքան բավարար չափով արտահայտված, որ բնական ընտրությունը կարողանա դրանք իրեն ենթարկել ու ավելի ևս զարգացնել»։
Բայց բոլոր նախորդ դեպքերում միջատներն արդեն իրենց նախասկզբնական վիճակում, անկասկած, ներկայացրել են ընդհանուր պատահական նմանություն իրենց շրջապատող իրադրության մեջ գտնվող որևէ սովորական առարկայի հետ։ Դա բոլորովին չի կարելի անհավանական համարել, ուշադրության առնելով շրջապատող առարկաների գրեթե անսահման քանակությունը և գոյություն ունեցող միջատների ձևի ու գույնի տարբերությունը։ Քանի որ ընդհանուր նմանությունը անհրաժեշտ է սկզբնավորման համար, ապա մեզ հասկանալի է դառնում, թե ինչու խոշոր և բարձրակարգ կազմվածքով կենդանիները (որքան ես գիտեմ, բացառությամբ մեկ ձկան) պաշտպանողական նպատակով չեն նմանվում որոշակի առարկաների, այլ միայն նմանվում են իրենց շրջապատող սովորական մակերևույթին, և այն էլ միայն գույնով։ Եթե ընդունենք, որ միջատներից մեկին պատահաբար վիճակվել է որոշ նմանություն ունենալ չոր ճյուղի կամ թափած տերևի հետ, և որ այդ միջատը փոփոխվում է զանազան ուղղություններով, այդ դեպքում անշուշտ բոլոր այն փոփոխությունները, որոնք նրան ամենից ավելի են նմանեցնում այդ առարկային և նպաստում են նրա թաքնվելուն, կպահպանվեն, մինչդեռ մյուս փոփոխությունները կթուլանան և, վերջապես իսպառ կանհետանան։ Իսկ եթե այդ փոփոխությունները միջատին դարձնում են նմանվելիք առարկային ընդհանրապես ավելի քիչ նմանվող, այդ դեպքում նրանք ևս կանհետանան։ Սակայն մ-ր Մայվարտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] առարկությունը ուժ կունենար, եթե մեզ հաջողվեր վերոհիշյալ նմանությունը բացատրել առանց բնական ընտրության օգնության, պարզապես անորոշ փոփոխականության միջոցով, իսկ իրերի ներկա դրությամբ այն նշանակություն չունի։
Ես չեմ կարող նշանակություն տալ այն դժվարությանը, որին հանդիպում է մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] «միմիկրիայի կատարելության վերջին աստիճաններում», ինչպես, օրինակ, մ-ր Ուոլլեսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] բերած դեպքը ձողիկանման միջատ Ceroxylus laceraetus-ի հետ, որը նման է «սողացող մամուռով կամ jungermannia-ով պատված ձողիկի»։ Այդ նմանությունը այնքան մեծ է, որ մի բնիկ Դայակ[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 148] պնդում էր, որ տերևանման մակաճումները իրոք ներկայացնում են մամուռներ։ Միջատները որպես կեր են ծառայում թռչունների և այլ թշնամիների համար, որոնց տեսողությունն անշուշտ ավելի սուր է մերինից, և նմանության ամեն մի աստիճան, որ օգնում է միջատին աննկատելի մնալու կամ չհայտնաբերվելու, նպաստում է նրա պահպանմանը, իսկ որքան նմանությունը ավելի լրիվ է, այնքան ավելի լավ է միջատի համար։ Ուսումնասիրելով տեսակային տարբերությունների բնույթը այդ խմբում, որին պատկանում է վերոհիշյալ Ceroxylus-ը մենք անհավանական ոչինչ չենք գտնում այն բանում, որ այդ միջատը փոփոխվել է իր մակերեսի անկանոնությունների կողմից, որոնք ձեռք են բերել շատ թե քիչ կանաչ գույն, որովհետև ամեն մի խմբում այն հատկանիշները, որոնք տարբեր են զանազան տեսակների մեջ, ամենից ավելի հակված են փոփոխվելու, մինչդեռ սեռայինները, այսինքն՝ բոլոր տեսակների համար ընդհանուր հատկանիշները, ամենից ավելի կայուն են։
Գրենլանդական կետը աշխարհի ամենազարմանալի կենդանիներից մեկն է և այսպես կոչված կետոսկրը ներկայացնում է նրա ամենից ավելի աչքի ընկնող առանձնահատկություններից մեկը։ Կետոսկրը կազմված է մի շարք եղջերաթիթեղներից, որոնք, գտնվելով վերին ծնոտի յուրաքանչյուր կողմում 300-ական հատ, դասավորված են բերանի խոռոչի երկայնական առանցքին հակառակ ուղղությամբ։ Գլխավոր շարքի շուրջը գտնվում են մի քանի լրացուցիչ շարքեր։ Եղջերաթիթեղների ծայրերը և ներսի եզրերը ճղված ու վերածված են ամուր խոզանների, որոնք պատում են ամբողջ ահռելի քիմքը և ծառայում են ջուր դուրս ծորելու կամ քամելու համար և պահելու համար մանր որսը, որով սնվում է հսկա կենդանին։ Գրենլանդական կետի միջին և ամենաերկար եղջերաթիթեղը հասնում է 10, 12 և նույնիսկ 15 ոտնաչափ երկարության, բայց կետերի տարբեր տեսակների մոտ այդ երկարությունը ներկայացնում է հաջորդական փոփոխություններ։ Ըստ Սկորեսբիի՝[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 149] մի տեսակի միջին եղջերաթիթեղը հասնում է չորս ոտնաչափի, մյուսինը՝ երեքի, երրորդինը՝ տասնևութ դյույմի, Balaenoptera rostrata մոտ՝ միայն մոտավորապես ինը դյույմի։ Կետոսկրի որակը նույնպես փոփոխվում է զանազան տեսակների մոտ։
Կետոսկրի մասին մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] նկատում է, որ «մի անգամ հասնելով այնպիսի մեծության և այնպիսի զարգացման, որով կետոսկրը դարձել է ընդհանրապես օգտակար, նա կարող է արդեն և՛ պահպանվել, և՛ զարգանալ հարկավոր չափերով միայն բնական ընտրության ազդեցության տակ։ Բայց ինչպե՞ս ստանալ այդպիսի օգտակար գոյացման սկզբնավորումը։ Ի պատասխան դրան կարելի է հարցնել, ինչո՞ւ կետերի հեռավոր նախնիները կետոսկրի հետ միասին չպետք է ունենային այնպիսի բերան, որն իր կառուցվածքով մի քիչ նման է բադի ատամնավոր կտուցին։ Բադերը կետերի նման կերակրվում են ցեխը և ջուրը քամելով, և այդ ընտանիքը երբեմն անվանել են Criblatores կամ քամողներ։ Հույսով եմ, որ ինձ սխալ կերպով չեն մեկնաբանի, իբրև թե ես ասում եմ, որ կետերի նախնիները ունեցել են եղջերաթիթեղներով օժտված բերան, որը նմանվել է բադի կտուցին։ Ես ուզում եմ միայն ասել, որ այդ անհավանական չպետք է համարել, և որ գրենլանդական կետի հսկայական եղջերաթիթեղները կարող էին աստիճանաբար զարգանալ այնպիսի թիթեղներից, որոնք զարգացման յուրաքանչյուր ստադիայում օգտակար են եղել իրենց տիրոջ համար։
Լայնակտուց բադի (Spatula clypeata) կտուցն իր կառուցվածքով թե՛ ավելի զարմանալի և թե՛ ավելի բարդ է, քան կետի բերանը։ Վերին ծնոտը յուրաքանչյուր կողմից օժտված է (իմ հետազոտած նմուշահատի վրա) մի շարք 188 բարակ, էլաստիկ թիթեղիկներով, որոնք շեղակի կտրված են այնպես, որ սրածայր են և զետեղված են բերանի երկայնական առանցքին լայնակի ուղղությամբ։ Նրանք դուրս են գալիս քիմքից և ճկուն թաղանթով ամրակցված են ծնոտի կողքերին։ Մեջտեղում գտնվող ամենաերկարները հասնում են մոտավորապես մի երրորդական դյույմ երկարության՝ եզրից ցածր կարկառելով 0,14 դյույմ[54]։
Նրանց հիմքի մոտ գտնվում է շեղալայնական թիթեղներից կազմված լրացուցիչ կարճ շարք։ Այսպիսով, նրանք զանազան տեսակետներից հիշեցնում են կետի բերանում եղած եղջերաթիթեղները, բայց կտուցի ծայրում շատ խիստ տարբերվում են, որովհետև ուղղված են դեպի ներս, այլ ոչ թե կախված են դեպի ցած։ Լայնակտուցի ամբողջ գլուխը, թեև անհամեմատ ավելի քիչ խոշոր է, կազմում է միջին մեծության գաճաճ շերտավորի[55] Balaenoptera rostrata-ի գլխի երկարության մոտավորապես մեկ տասնևութերորդականը, որի կետոսկրն ունի միայն ինը դյույմ երկարություն․ հետևաբար, եթե մենք լայնակտուցի գլուխը պատկերացնենք կետի գլխի երկարության հավասար, այդ դեպքում նրա թիթեղիկները կունենային վեց դյույմ երկարություն, այսինքն կկազմեին կետի տվյալ տեսակի ունեցած կետոսկրի երկու երրորդականը։ Ստորին ծնոտը օժտված է նույն երկարության թիթեղիկներով, ինչ որ վերինը, բայց ավելի բարակ են և այդ տեսակետից խիստ տարբերվում են կետի ստորին ծնոտից, որը զուրկ է եղջերաթիթեղներից։ Մյուս կողմից, այդ ստորին թիթեղիկների եզրերը ճեղքված են բարակ խոզանակաձև ելուստների, որով նրանք զարմանալիորեն նմանվում են կետոսկրին։ Մրրկահավերի մի առանձին ընտանիքի պատկանող Prion սեռի միայն վերին ծնոտն ունի թիթեղիկներ, որոնք լավ զարգացած են և իջնում են եզրից ցած, որի պատճառով այդ թռչունի կտուցը մատնանշված տեսակետից նմանվում է կետի բերանին։
Լայնակտուցի բարձր զարգացած կառուցվածք ունեցող կտուցից մենք կարող ենք (ինչպես ես այդ գիտեմ այն հաղորդումից ու նմուշներից, որ ինձ ուղարկել է մ-ր Սալվինը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 150]), առանց մեծ ընդհատման, որքանով այդ վերաբերում է քամելու համար կտուցի պիտանիությանը, անցնել Merganetta armata-ի կտուցով և որոշ տեսակետներից Aix sponsa-ի կտուցով դեպի սովորական բադի կտուցը։ Վերջին տեսակի մոտ թիթեղիկներն անհամեմատ ավելի կարճ են, քան լայնակտուցինը, և ամուր կերպով առաճել են ծնոտի կողքերին, յուրաքանչյուր կողմից նրանք ընդամենը մոտավորապես հիսունական են և ծնոտի եզրից ցած չեն իջնում։ Իսկ այդ թիթեղիկների քառակուսի գագաթները եզրին վերածվում են թափանցիկ ամուր հյուսվածքի, որը պիտանի է սնունդը մանրացնելու համար։ Ստորին ծնոտի եզրերը ծածկված են բազմաթիվ բարակ լայնակի կողերով, որոնք շատ քիչ են դուրս ցցվում։ Թեև այդպիսի կառուցվածքով կտուցը որպես մաղ անհամեմատ ավելի ցածր է կանգնած, քան լայնակտուցի կտուցը, սակայն այդ թռչունները, ինչպես ամենքին հայտնի է, իրենց կտուցը միշտ օգտագործում են այդ նպատակի համար։ Մյուս տեսակների թիթեղիկները, ինչպես ինձ հաղորդել է մ-ր Սալվինը, նույնիսկ զգալի չափով ավելի քիչ են զարգացած, քան սովորական բադինը, բայց ես չգիտեմ, արդյոք նրանք օգտագործո՞ւմ են իրենց կտուցը ջուր քամելու համար։
Դառնանք նույն ընտանիքի մի այլ խմբի, Եգիպտական սագի (Chenalopex) կտուցը շատ նման է սովորական բադի կտուցին, բայց նրա թիթեղիկներն այնպես բազմաթիվ չեն, այնպես խիստ հեռացած չեն մեկը մյուսից և այնպես չեն կարկառվում դեպի ներս, ինչպես մեր բադինը, սակայն այդ սագը, ինչպես ինձ հաղորդել է մ-ր Բարտլետտը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 151] «իր կտուցը գործ է ածում, ինչպես բադը, ջուրը թիթեղների միջև եղած արանքներով քամելու համար», թեև նրա գլխավոր կերը կազմում է խոտը, որ պոկոտում է սովորական սագի նման։ Վերջին թռչունի վերին ծնոտի թիթեղիկներն անհամեմատ ավելի կարճ են, քան ընտանի բադինը. նրանք համարյա միաձուլվում են, յուրաքանչյուր կողմում գտնվում են մոտավորապես 27-ական հատ և վերջանում են ատամնանման թմբիկներով։ Քիմքը նույնպես ծածկված է ամուր բութ ելուստներով։ Ստորին ծնոտի եզրերը ատամնավոր են, անհամեմատ ավելի զարգացած քառթերի ներկայության շնորհիվ, որոնք թե՛ ավելի խոշոր են և թե՛ ավելի սուր են, քան բադինը։ Սովորական սագը ջուր չի քամում, այլ իր կտուցը օգտագործում է բացառապես նրա համար, որպեսզի խոտ պոկռտի, որին այդ կտուցը այնքան լավ է հարմարված, որ սագը կարող է նրանով խոտը ավելի տակից պոկռտել, քան մյուս կենդանիները։ Սագերի այլ տեսակների մոտ, ինչպես ինձ հաղորդել է մ-ր Բարտլետտը, թիթեղիկներն ավելի քիչ են զարգացած, քան սովորական սագի մոտ։
Այսպիսով, մենք տեսնում ենք, որ բադազգիների ընտանիքի մի քանի անդամներ, որոնց կտուցը կառուցված է սովորական կտուցի նման և հարմարված է բացառապես նրա համար, որ խոտ պռճոկի, կամ նույնիսկ ավելի քիչ զարգացած թիթեղներով կտուց ունեցողները կարող են փոքրիկ փոփոխությունների միջոցով ձևափոխվել այնպիսի տեսակի, ինչպիսին է եգիպտական սագը, վերջինս էլ կարող է ձևափոխվել սովորական բադի նման մի տեսակի, իսկ սա, վերջապես, մի տեսակի, որի կտուցը նման է լայնակտուցի կտուցին և պիտանի է գրեթե բացառապես ջուր քամելու համար, որովհետև հազիվ թե այդ թռչունը կարողանա իր կտուցի որևէ մասը, բացի նրա ուռած գագաթից, գործածել սնունդը բռնելու և մանրացնելու համար։ Դրան ես կարող եմ ավելացնել այն, որ փոքրիկ փոփոխության միջոցով սագի կտուցը կարող է փոխարկվել դուրս ցցված, ետ ծռված ատամիկներ ունեցող կտուցի, որը նման է սղոցակտուց բադի (նույն ընտանիքի ներկայացուցիչ) կտուցին և ծառայում է բոլորովին հատուկ նպատակի՝ կենդանի ձուկ բռնելու համար։
Դառնանք կետերին։ Hyperoodon bidens-ը հասուն վիճակում զուրկ է իսկական ատամներից, բայց դրա քիմքը ծածկված է, ըստ Լասեպեդի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 152] փոքրիկ անհավասար ու ամուր եղջերային կետերով։ Այդ պատճառով ոչ մի անհավանական բան չկա այն ենթադրության մեջ, որ կետերի վաղ ձևերից որևէ մեկը ունենար այնպիսի քիմք, որը ծածկված էր մի քիչ ավելի կանոնավոր դասավորված նույնպիսի եղջերացած տեղամասերով, որոնք սագի կտուցում եղած թմբիկների նման օգնել են կերը բռնելուն և մանրացնելուն։ Բայց եթե այդպես է, ապա հազիվ թե կարելի է ժխտել, որ մատնանշված տեղամասերը փոփոխելիության ու բնական ընտրության հետևանքով կարող էին փոխարկվել թիթեղիկների, որոնք նույնքան զարգացած էին, որքան եգիպտական սագի մոտ, և միաժամանակ պիտանի էին թե՛ ջուր քամելու և թե՛ որս բռնելու համար. այնուհետ և հետևում են ընտանի բադի թիթեղիկներին նմանվող թիթեղիկները, և այսպես էլ հետագայում, մինչև որ նրանք դարձել են նույնքան կատարյալ, ինչքան լայնակտուցինն է, փոփոխվելով ու վերածվելով բացառապես քամելու համար պիտանի ապարատի։ Սկսած այդ ստադիայից, երբ թիթեղիկներն ունենում են Balaenoptera rostrata-ի կետոսկրի թիթեղի երկարության երկու երրորդականը, աստիճանաբար նրանց զարգացումը, որը նկատվում է դեռ ապրող կետերի մոտ, հասնում է գրենլանդական կետի կետոսկրի հսկայական թիթեղներին։ Մենք ամենափոքրիկ հիմք էլ չունենք կասկածելու, որ այդ շարքում յուրաքանչյուր աստիճան կարող էր նույնքան օգտակար լինել նախկին դարաշրջանների կետերից մի քանիսին, երբ ուսումնասիրվող գոյացումները հաջորդական զարգացման ընթացքում կրել են ֆունկցիաների աստիճանական փոփոխություն, որքան որ օգտակար են կտուցի հաջորդական փոփոխությունները բադազգիների ընտանիքի գոյություն ունեցող զանազան անդամների համար։ Մենք պետք է հիշենք, որ բադերի յուրաքանչյուր տեսակ ենթակա է գոյության դաժան կռվին և որ թռչունի մարմնի յուրաքանչյուր օրգանը պետք է լավ հարմարված լինի նրա կենսապայմաններին։
Pleuronectidae կամ տափակաձկները զարմանալի են իրենց մարմնի անհամաչափությամբ։ Նրանք հատակին պառկում են մի կողքի վրա, տեսակների մեծամասնությունը՝ ձախ կողքի վրա, մի քանիսը՝ աջ, պատահմամբ հանդիպում են հակառակ զարգացում ունեցող հասուն անհատներ։ Ներքևի կողմը, որի վրա պառկում է ձուկը, առաջին հայացքից նման է սովորական ձկան որովայնի կողմին. նա սպիտակ գույն ունի, շատ տեսակետներից ավելի քիչ զարգացած է, քան վերևինը, ունի փոքրիկ լողաթևեր։ Բայց ամենագլխավոր առանձնահատկությունը կազմում են աչքերը, որոնք երկուսն էլ գտնվում են գլխի վերին կողմում։ Սակայն վաղ հասակում նրանք գտնվում են մեկը մյուսի հակադիր կողմում, և այդ ժամանակ ամբողջ մարմինը համաչափ է, երկու կողմերն էլ միանման գույն ունեն։ Շուտով ներքևի աչքն սկսում է աստիճանաբար տեղափոխվել դեպի գլխի վերին կողմը, բայց չի անցնում ուղղակի գանգի միջով, ինչպես կարծում էին առաջ։ Ակներև է, որ քանի դեռ ներքևի աչքն անցնում է դեպի վերին կողմը, ձուկը, պառկած լինելով իր սովորական դրության մեջ մի կողմի վրա, չի կարող այն օգտագործել։ Այդ աչքը, բացի դրանից, գտնվելով ներքևի կողմում, պետք է շփվեր ավազոտ գետնի հետ։ Որ Pleuronectidae-ները իրենց տափակ ձևով և անհամաչափ կառուցվածքով հիանալի կերպով հարմարված են իրենց կենսակերպին, ապացուցվում է նրանով, թե ինչքան սովորական են նրանց շատ տեսակները, օրինակ պալտուսը և այլն։ Դրանից բխող ամենակարևոր առավելությունները, ըստ երևույթին, թշնամիներից պաշտպանվելն է և հատակում հեշտությամբ կեր ճարելն է։ Սակայն ըստ Շիեդտեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 95] դիտողության, այդ ընտանիքի զանազան անդամները կազմում են «ձևերի երկար շարք, որոնք ներկայացնում են աստիճանական անցում սկսած Hippoglossus piuguis պալտուսից, որը ձվից դուրս գալիս ստացած տեսքը որևէ նկատելի չափով չի կորցնում, մինչև տափակաձկները, որոնք բոլորովին շրջված են մի կողմի վրա»։
Մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] կանգ է առնում այդ դեպքի վրա և նկատում է, որ աչքերի դիրքի անսպասելի ինքնակամ փոփոխությունը հազիվ հասկանալի է, որի մասին ես նրա հետ համաձայն եմ։ Նա դրան ավելացնում է. «Եթե այդպիսի տեղափոխությունը եղել է աստիճանաբար, ապա բնավ պարզ չէ, թե անհատի օգուտն ինչումն է արտահայտվել աչքերի այդպիսի շատ փոքր տարածության վրա՝ գլխի մի կողմից մյուսը տեղափոխվելուց։ Նույնիսկ թվում է, թե օրգանի դիրքի այդպիսի հազիվ նշմարելի փոփոխությունը կարող էր որոշ չափով վնասակար լինել»։ Բայց նա իր առարկությունների պատասխանը կարող էր գտնել Մալմի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 153] հիանալի դիտողություններում, որ հրատարակված են 1867 թվականին։ Տափակաձկները շատ վաղ հասակում լինելով համաչափ, ունենալով գլխի հակադիր կողմերում գտնվող աչքեր, չեն կարող երկար ժամանակ մնալ ուղղաձիգ գրության մեջ իրենց մարմնի բարձրության, կողքի լողաթևերի շատ փոքրության և լողափամփուշտի թերզարգացման հետևանքով։ Աճման հետ զուգընթաց նրանք ուղղաձիգ դրության մեջ արագորեն հոգնում են և ընկնում հատակին մի կողքի վրա։ Գտնվելով այդպիսի վիճակում, նրանք հաճախ, ինչպես դիտել է Մալմը, ներքևի աչքը շրջում են դեպի վեր, որպեսզի տեսնեն իրենցից վեր, և այդ անում են այնպիսի եռանդով, որ աչքը պինդ սեղմում են ակնախոռոչի վերին եզրին։ Հեշտությամբ կարելի է տեսնել, որ այդ ժամանակ երկու աչքերի միջև գտնվող գլխամասը ժամանակավորապես կրճատվում է լայնությամբ։ Մի դեպքում Մալմը տեսել է, թե ինչպես մատղաշ ձուկը իջեցրել ու բարձրացրել է աչքը, ըստ որում անկյունային տեղաշարժը հասել է 70°-ի։
Մենք պետք է հիշենք, որ գանգը զարգացման վաղ ստադիայում կրճկային է և ճկուն, ուստի և հեշտությամբ ենթարկվում է մկանների ներգործությանը։ Բարձրակարգ կենդանիների մասին հայտնի է նաև, որ նրանց վաղ երիտասարդական շրջանից հետո անգամ գանգը փոփոխություններ է կրում, եթե մաշկը կամ մկանները շարունակ կծկված են հիվանդության կամ որևէ այլ հանգամանքի ազդեցության տակ։ Եթե երկարականջ ճագարների մի ականջը կախված է դեպի առաջ ու ցած, ապա նրա ծանրությունը ձգում է դեպի առաջ գանգի միևնույն կողմի բոլոր ոսկորները, որը ես ցույց եմ տվել իմ «Կենդանիների և բույսերի փոփոխությունն ընտանեցման ազդեցության տակ» աշխատության առաջին հատորում զետեղված մի նկարի վրա։ Մալմը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 153] պնդում է, որ պերկեսների, սալմոնների և զանազան այլ համաչափ ձկների նորածին ձկնիկները սովորություն ունեն երբեմն հանգստանալու, պառկելով հատակին մի կողքի վրա, և նա դիտել է, որ նրանք հաճախ այդ ժամանակ իրենց ներքևի աչքը շրջում են դեպի վեր. դրա հետ միասին նրանց գանգը փոքր ինչ ծռվում է։ Սակայն այդ ձկները շուտով ընդունակ են դառնում շարունակ մնալու ուղղաձիգ գրության մեջ և նրանց ժամանակավոր սովորությունը ոչ մի հետևանք չի առաջացնում։ Մյուս կողմից, Pleuronectidae-ը իրենց մարմնի տափակացումն ավելանալու հետ միասին որքան մեծանում են, այնքան ավելի երկար են մնում մի կողքի վրա, և որի մշտական հետևանքն այն է լինում, որ փոփոխվում է գլխի ձևը և աչքերի դիրքը։ Դատելով անալոգիայի հիման վրա, կասկած չկա, որ ծռվելու հակումը ժառանգության օրենքով ուժեղանում է։ Հակառակ մի քանի այլ բնախույզների, Շիեդտեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 95] կարծում է, որ Pleuronectidae-ը բոլորովին համաչափ չեն լինում նույնիսկ սաղմնային վիճակում, և եթե այդ ճիշտ է, ապա մենք կարող ենք հասկանալ, թե ինչու մի քանի տեսակների ձկնիկները վաղ հասակում սովորաբար ընկնում են հատակին ու մնում ձախ կողմի վրա պառկած, մյուսները՝ աջ։ Մալմը վերոհիշյալ կարծիքը հաստատելու համար ավելացնում է, որ չափահաս Trachiptefus arcticus-ը, որը Pleuronectidae-ի ընտանիքին չի պատկանում, հատակին պառկում է ձախ կողքի վրա, իսկ լողում է շեղակի վիճակում. այդ ձկան գլխի աջ և ձախ կողմերը, ինչպես ասում են, ամենևին համաչափ չեն։ Ձկների վերաբերյալ մեր ականավոր հեղինակություն դոկտոր Գյունտերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 154] Մալմի հոդվածից արած քաղվածքը վերջացնում է դիտողությամբ, որ «հեղինակը Pleuronectidae-ների աննորմալ վիճակին շատ պարզ բացատրություն է տալիս»։
Սրանից կարելի է տեսնել, որ գլխի մի կողմից մյուսը աչքի տեղափոխության ստադիաները, մի հանգամանք, որ մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] համարում է վնասակար, կարող են թե՛ անհատի և թե՛ տեսակի համար անկասկած բարենպաստ այն սովորությանը վերադրվել, որ ձուկը երկու աչքն էլ դեպի վեր է դարձնում, որպեսզի կարողանա նրանցով տեսնել, երբ պառկած է հատակին մի կողքի վրա։ Ճիշտ այդպես էլ մենք կարող ենք գործածության ժառանգված հետևանքին վերագրել այն, որ որոշ տափակամարմին ձկների բերանը ուղղված է դեպի ցած և որ գլխի այդ, աչքերից զուրկ կողմում ծնոտներն ավելի ամուր են և ուժեղ, քան մյուս կողմում, որպեսզի ավելի հեշտությամբ ապահովվի, ինչպես ենթադրում է դոկտոր Տրակվերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 155] կերի հայթայթումը հատակից։ Մյուս կողմից, անգործածությամբ բացատրվում է մարմնի ամբողջ ստորին կեսի, ներառյալ նաև կողքի լողաթևերի թերզարգացումը, թեև Յարրելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 156] կարծում է, որ այդ կողքի լողաթևերի փոքրացումը օգտակար է ձկան համար, որովհետև «ներքևի կողմում նրանց շարժելու համար անհամեմատ ավելի քիչ տեղ կա, քան վերևի կողմում։ Գուցե պալտուսի ծնոտների ստորին կեսերի ունեցած 25—30 ատամների համեմատությամբ վերին կեսերի ատամների թվի նվազումը մինչև 4—7-ի, կարելի է բացատրել նրանց անգործածությամբ։ Ելնելով ձկների մեծամասնության և շատ այլ կենդանիների որովայնի կողմում գունավորման բացակայությունից, մենք լիովին հիմք ունենք ենթադրելու, որ տափակաձկան մի կողմում, այդ միևնույնն է, թե աջն է ներքևում, կամ ձախը, գույնի բացակայությունը լույսի բացակայությունից է։ Բայց չի կարելի կարծել, թե տափակաձկան վերին կողմի յուրահատուկ բծավոր գույնը, որ այնքան նմանվում է ծովի ավազոտ հատակին, ինչպես նաև մի քանի տեսակների ընդունակությունը փոփոխելու իրենց գույնը շրջապատող մակերևույթի համեմատ, որ վերջերս ապացուցել է Պուշեն,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 157] կամ տափակաձկան վերին կողմում ոսկրային թմբիկների ներկայությունը պարտական են լույսի ազդեցությանը։ Այդ դեպքերում հավանորեն արտահայտվել է բնական ընտրության գործունեությունը, որը մի կողմից փոփոխել է այդ ձկների մարմնի ընդհանուր ձևը, իսկ մյուս կողմից՝ նաև նրանց կազմվածքի մասնավոր շատ առանձնահատկություններ, համապատասխանեցնելով նրանց կենսակերպին։ Մենք պետք է հիշենք, որի վրա ես առաջները պնդել եմ, որ օրգանների գործածության, կամ կարող է պատահել, նաև նրանց անգործության հետ կապ ունեցող ժառանգված առանձնահատկությունները ուժեղանում են բնական ընտրության ազդեցության տակ։ Այսպիսով պահպանվում են և՛ բոլոր պատահական օգտակար առանձնահատկությունները, և՛ այն անհատները, որոնք ամենամեծ չափով են ժառանգել որևէ օրգանի ուժեղացած օգտակար գործածության հետևանքները։ Յուրաքանչյուր առանձին դեպքում թե ինչը պետք է վերագրել գործածության հետևանքներին և ինչը բնական ընտրության ազդեցությանը, հավանորեն անհնարին է որոշել։
Ես կարող եմ բերել կառուցվածքի այդպիսի փոփոխության մի այլ օրինակ ևս, որը, ըստ երևույթին, սկիզբ է առել բացառապես օրգանի գործածությունից կամ որոշ սովորությունից։ Ամերիկական կապիկներից մի քանիսի պոչի ծայրը փոփոխվել ու դարձել է բռնելու զարմանալի կատարյալ օրգան և ծառայում է որպես հինգերորդ ձեռք։ Մի բնախույզ, որ իր տեսակետներով մինչև մանրունքները համաձայն է Մայվարտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] հետ, պոչի կառուցվածքի մասին նկատում է հետևյալը. «Անհնարին է կարծել, որ որևէ դրության մեջ առաջին անգամ երևան եկած բռնելու հազիվ նշմարելի հակումը կարող էր ծառայել դրանով օժտված անհատների պաշտպանության համար կամ նպաստեր նրանց ունենալու և դաստիարակելու իրենց սերունդներին»։ Բայց այդպիսի ենթադրության ոչ մի անհրաժեշտություն չկա։ Ամենայն հավանականությամբ այս դեպքում բավական է սովորությունը, որն ինքնին արդեն հարկադրում է ենթադրելու, որ դրա հետ կապված է ավելի կամ պակաս չափով օգուտ։ Բրեմը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 158] տեսել է, որ աֆրիկական փոքրիկ կապիկները (Cercopithecus) ձեռքերով կպչելով մոր մարմնին ցածի կողմից, միաժամանակ իրենց կարճ պոչը փաթաթում են մոր պոչին։ Պրոֆ. Հենսլոուն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 159] անազատ վիճակում պահել է մի քանի դաշտային մկներ (Mus messorius), որոնք չունեն իսկական կառչող պոչ. բայց հաճախ է տեսել, որ նրանք իրենց պոչերով փաթաթվում էին իրենց վանդակում դրված թփի ճյուղին և այդպիսով օգնում էին իրենց մագլցելու ժամանակ։ Համանման դիտողության մասին ինձ հաղորդել է դ-ր Գյունտերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 154] որը տեսել է, թե ինչպես մուկն այդ ձևով կախվել է։ Եթե դաշտային մուկը հարմարված լիներ ծառերի վրա ապրելու համար, ապա թերևս նրա պոչը կառուցվածքով ավելի կառչուն կդառնար, ինչպես այդ տեսնում ենք հենց նույն կարգի մի քանի անդամների մոտ։ Թե ինչու ցերկոպիթեկը իր սովորություններով հանդերձ վաղ հասակում ձեռք չի բերել այդ դժվար է ասել։ Սակայն հնարավոր է, որ այդ կապիկի երկար պոչը նրան ավելի մեծ ծառայություն է մատուցում նրա զարմանալի թռիչքների ժամանակ որպես հավասարակշռության պահպանելու օրգան, քան թե որպես բռնելու օրգան։
Կաթնագեղձերը հանդիպում են կաթնասունների ամբողջ դասի մեջ և անհրաժեշտ են նրանց գոյության համար, ըստ երևույթին, նրանք զարգացել են շատ հեռավոր ժամանակաշրջանում, և մենք նրանց զարգացման եղանակի մասին հաստատական ոչինչ չգիտենք։ Մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] հարց է տալիս. «Կարելի՞ է արդյոք ընդունել, որ մի որևէ կենդանու ձագ երբևիցե մահից ազատվել է նրանով, որ պատահաբար ծծել է նվազ սննդարար հեղուկի մի կաթիլ իր մոր մաշկի հիպերտրոֆիայի ենթարկված գեղձերից։ Եվ եթե այդպես եղել է մի անգամ, ապա ի՞նչ հավանականություններ կան նման փոփոխության կրկնության համար»։ Բայց այստեղ բանն այդպես չէ։ Էվոլյուցիոնիստների մեծամասնությունն ընդունում է, որ կաթնասուններն առաջացել են երկարգանդային ձևից, իսկ եթե այդպես է, այդ դեպքում կաթնագեղձերն սկզբում զարգացել են պարկի մեջ, որը բնորոշում է երկարգանդավորներին։ Մի ձկան (ծովաձի, Hippocampus) ձվերը պահպանվում են և նորածին ձկնիկները որոշ ժամանակ սնվում են նման պարկի մեջ, և ամերիկական բնախույզ մ-ր Լոկվուդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 160] կարծում է, դատելով նրանից՝ ինչ ինքը նկատել է ձկնիկների զարգացման վերաբերմամբ, որ ձկնիկները սնվում են պարկի մաշկագեղձերի արտաթորանքով։ Այժմ, ինչ վերաբերում է կաթնասունների հեռավոր նախնիներին, մոտավորապես այն ժամանակից առաջ, երբ նրանք դեռ հազիվ արժանանում էին այդ անունը կրելու, ապա չի կարելի արդյոք գոնե հնարավոր համարել, որ նրանց զավակները կերակրվել են հենց նման ձևով։ Եվ այս դեպքում այն անհատները, որ արտաթորել են այս կամ այն կերպ ավելի սննդարար և կաթի հատկություն ունեցող հեղուկ, կարող էին երկար ժամանակամիջոցի ընթացքում սնուցել ավելի մեծ թվով լավ կերակրված հաջորդներ, քան այն անհատները, որոնք արտաթորել են ավելի պակաս սննդարար հեղուկ։ Ընդսմին մաշկային գեղձերը, որ հոմոլոգ են կաթնագեղձերին, կարող էին կատարելագործվել և դառնալ ավելի գործունյա։ Այն հանգամանքը, որ պարկի որոշ տարածության մեջ պարփակվող գեղձերը կարող էին մյուսներից ավելի զարգանալ, լիովին համաձայնվում է մասնագիտացման վերաբերյալ լայնորեն տարածված սկզբունքի հետ, նրանք կարող էին գոյացնել նաև «կուրծ» սկզբում առանց պտուկի, ինչպես բադակտուցինն է, որը կանգնած է կաթնասունների շարքի սկզբում։ Ես ինձ վրա չեմ վերցնում որոնելու այն, թե ինչից կախված մի տեղում գեղձերն ավելի մասնագիտացել են, քան մի այլ տեղում, գուցե կախված է եղել մասամբ աճումից, մասամբ որպես գործածության հետևանք կամ բնական ընտրության ազդեցության տակ։
Կաթնագեղձերի զարգացումը նշանակություն չէր ունենա և բնական ընտրության ազդեցության տակ չէր ընկնի, եթե նորածինները չկարողանային միևնույն ժամանակ օգտվել նրանց արտաթորությունից։ Հասկանալ այն, թե ինչպես նորածին կաթնասունը սովորել է բնազդաբար ծծել կուրծը, ավելի դժվար չէ, քան հասկանալ այն, թե ինչպես ձվից դուրս չեկած ճուտերը սովորել են կոտրել ձվի կեղևը, խփելով նրան հատկապես դրա համար հարմարված կտուցով, կամ ինչպես նրանք սովորել են ձվից դուրս գալուց մի քանի ժամ հետո կեր կտցել։ Այդպիսի դեպքերում հարցի ամենահավանական լուծումը, թվում է թե, այն է, որ այս կամ այն սովորությունը սկզբում ձեռք է բերվել ավելի ուշ հասակում, իսկ հետո հաջորդին փոխանցվել է ավելի վաղ հասակում։ Բայց նորածին կենգուրուն, ինչպես ասում են, չի ծծում, այլ միայն կպչում է մոր պտկին, որն ընդունակություն ունի կաթ սրսկելու իր անօգնական և կիսակազմավորված զավակի բերանը։ Դրա վերաբերմամբ մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] նկատում է. «Եթե գոյություն չունենար հատուկ նախապաշտպանում, նորածին ձագը հավանորեն կոչնչանար կաթը շնչափողի մեջ թափանցելուց։ Բայց այդպիսի նախապաշտպանում գոյություն ունի։ Կոկորդի եզրերը հեռացած են այնքան, որ հասնում են քթանցքի վերջի եզրին, որով հնարավոր է լինում օդի թափանցումը թոքերի մեջ, մինչդեռ կաթն անարգել կերպով հոսում է կոկորդի կողքերով և հաջող կերպով անցնում կերակրափողի մեջ»։ Այնուհետև մ-ր Մայվարտը հարցնում է, թե ի՞նչ կերպով բնական ընտրությունը ոչնչացրել է չափահաս կենգուրուի (և մյուս կաթնասունների մեծամասնության, ընդունելով նրանց ծագումը երկար գանգային ձևից) «կառուցվածքի այդ, առնվազն, բոլորովին անմեղ ու անվնաս առանձնահատկությունը»։ Ի պատասխան դրան, կարելի է ենթադրել, որ ձայնը, որը անշուշտ մեծ նշանակություն ունի շատ կենդանիների համար, հազիվ թե հասներ լիակատար ուժգնության, քանի դեռ կոկորդը մտնում էր քթանցքի մեջ, իսկ պրոֆ. Ֆլոուերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 161] ինձ հայտնել է մի կշռադատություն, որ այդպիսի կառուցվածքը մեծ արգելք կլիներ պինդ սնունդ կուլ տվող կենդանու համար։
Այժմ մենք կարճ ժամանակով դառնանք կենդանիների ստորակարգ բաժիններին։ Փշամորթները (ծովաստղերը, ծովաոզնիները և այլն) օժտված են զարմանալի օրգաններով, այսպես կոչված պեդիցելլյարիաներով, որոնք միանգամայն զարգացած տեսակի մեջ ներկայացնում են եռաբաժան ունելի, այսինքն երեք ատամնավոր ելուստներ՝ ամուր կերպով միացած միմյանց հետ և ամրացած ճկուն կոթունի գագաթին, որը շարժման մեջ է դրվում մկանների միջոցով։ Այդ ունելիները կարող են ամուր կերպով բռնել զանազան առարկաներ և Ալեքսանդր Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] տեսել է, թե ինչպես ծովաոզնին իր կղկղանքի մասնիկները արագորեն տալիս էր մի ունելիից մյուսին իր մարմնի վրայով որոշ ուղղությամբ, որպեսզի չաղտոտվի իր կեղևը։ Բայց կասկածից դուրս է, որ բացի զանազան կեղտեր հեռացնելուց, այդ գոյացումները կատարում են նաև այլ աշխատանքներ, և դրանց մեկը պաշտպանությունն է։
Այդ օրգանների վերաբերմամբ մ-ր Մայվարտը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] ինչպես և նախորդ շատ դեպքերում, հարցնում է. «Ի՞նչ ձևի օգուտ կարող էր լինել այդ օրգանների առաջին սկզբնավորումից և ինչպե՞ս այդ չնչին սկզբնավորումները կարող էին պահպանել առանձին ծովաոզնիների կյանքը»։ Եվ նա ավելացնում է. «Նույնիսկ բռնելու գործողության հանկարծական զարգացումը օգուտ չէր բերի առանց շարժական կոթունի, ճիշտ այդպես էլ վերջինս նշանակություն չէր ունենա առանց բռնող ունելիների. սակայն կառուցվածքի մեջ այդ բարդ համապատասխանության զարգացման համար բավական չէին միաժամանակ անսահմանորեն մանր, պարզապես անորոշ փոփոխություններր,— ժխտել այդ՝ նշանակում է, առնվազն պաշտպանել մի կատարյալ պարադոքս»։ Սակայն Մայվարտին այդ ինչքան էլ պարադոքսային թվա, երեք սայրով ունելիները, որ իրենց հիմքով անշարժ ամրակցված են, բայց ընդունակ են բռնելու գործողության համար, անկասկածորեն գոյություն ունեն մի քանի փշամորթների մոտ. և այդ միանգամայն հասկանալի է, եթե նրանք թեկուզև մի քիչ ծառայում են պաշտպանության համար։ Մ-ր Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97], որի սիրալիրությանը ես պարտական եմ այդ խնդրի վերաբերյալ տեղեկությունների մեծ մասի համար, հաղորդել է ինձ, որ մի քանի փշամորթների եռաժանու երեք ճյուղերից մեկը մյուս երկուսին օժանդակելու համար է, այնինչ կան նաև այնպիսի սեռեր, որոնք երրորդ ճյուղը բոլորովին չունեն։ Պերյեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 162] նկարագրությամբ Echinoneus-ի պեդիցելլյարիաները երկու ձև ունեն, մի քանիսը նման են Echinus-ի պեդիցելլյարիաներին, մյուսները՝ սպատանգի պեդիցելլյարիաներին. այդպիսի դեպքերը միշտ հետաքրքրական են, որովհետև ծառայում են բացատրելու համար, ըստ երևույթին, անսպասելի փոփոխությունները օրգանի երկու վիճակներից մեկը կորցնելու միջոցով։
Ինչ վերաբերում է այն գոյացումներին, որոնցից կարող էին զարգանալ այս հետաքրքրական օրգանները, ապա մ-ր Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] իր սեփական հետազոտությունների և Մյուլլերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] հետազոտությունների հիման վրա եկել է այն եզրակացության, որ ինչպես ծովաստղերի, այնպես էլ ծովաոզնիների պեդիցելլյարիաները անկասկածորեն կարելի է դիտել որպես ձևափոխված փշեր։ Դրանում կարելի է համոզվել ինչպես առանձին անհատների մոտ նրանց զարգացումից, այնպես էլ այն անցումների լրիվ շարքից, որոնք զանազան տեսակների և սեռերի մեջ պարզ թմբիկներից տանում են դեպի փշերը և փշերից դեպի լիովին զարգացած եռաճյուղ պեդիցելլյարիաները։ Անցման աստիճանականությունը տարածվում է նույնիսկ սովորական փշի ու պեդիցելլյարիայի և նրանց նեցուկ հանդիսացող կրային թիթեղիկների հետ հոդավորման եղանակի վրա։ Ծովաստղերի մի քանի սեռերի մոտ կարելի է գտնել «ցուցումներ այն մասին, որ պեդիցելլյարիաները ներկայացնում են պարզապես փոփոխված ճյուղավոր փշեր»։ Նրանք ունեն անշարժ փշեր սղոցաձև երեք շարժական ճյուղերով, որոնք բաժանված են հավասար հեռավորություններով և հոդավորված են փշի հիմքում, իսկ ավելի բարձր, միևնույն փշի վրա, նստած են երեք ուրիշ շարժական ճյուղիկներ։
Եթե վերջինները դուրս են գալիս փշի գագաթից, այդ դեպքում մենք մեր առջև ունենք ըստ էության կոպիտ եռաճյուղ պեդիցելլյարիա, որը կարելի է տեսնել փշի վրա երեք հիմնական ճյուղերով միասին։ Այդ դեպքում պեդիցելլյարիայի Ճյուղերի և փշի շարժական ճյուղերի միջև նմանությունը կասկածի ենթակա չէ։ Ընդհանրապես ընդունված է կարծել, որ պարզ փշերը ծառայում են պաշտպանության համար, իսկ եթե այդպես է, ապա հիմք չկա կասկածելու այն բանում, որ այն փշերը ևս, որոնք օժտված են սղոցաձև և շարժուն ճյուղերով, ծառայում են նույն նպատակի համար, բայց նրանց նշանակությունն այդ տեսակետից ավելի ևս կմեծանա, եթե նրանք միևնույն ժամանակ կդառնան նաև բռնող ապարատ։ Այսպիսով, բոլոր անցումները, սկսած պարզ նստած փշից մինչև պեդիցելլյարիան, ունեն օգտակար նշանակություն։
Ծովաստղերի մի քանի սեռերի այդ օրգանները փոխանակ մի անշարժ հիմքի ամրանալու, գտնվում են թեև կարճ, բայց ճկվելու ընդունակ մկանոտ կոթունի գագաթին. այդ դեպքում հավանորեն նրանք, բացի պաշտպանությունից, կատարում են նաև որևէ լրացուցիչ ֆունկցիա։ Ծովաոզնիների մոտ կարելի է հետամուտ լինել, թե ինչպես անշարժ նստած փուշը աստիճանաբար ձեռք է բերում հոդեր և այդպիսով դառնում է շարժական։ Ես կցանկանայի այստեղ տեղ ունենալ, որպեսզի ավելի լրիվ քաղվածքներ բերեի պեդիցելլյարիաների զարգացման մասին մ-ր Ագասսիսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] հետաքրքրական դիտողություններից։ Նա նույնպես ասում է, որ բոլոր հնարավոր անցումները հավասարապես կարելի է գտնել ծովաստղերի պեդիցելլյարիաների և փշամորթների մի խմբի՝ օֆիուրների կարթերի միջև, ինչպես նաև ծովաոզնիների պեդիցելլյարիաների և հոլոթուրների խարիսխների միջև, որոնք պատկանում են միևնույն մեծ դասին։
Գաղութներով ապրող կենդանիներից կամ զոոֆիտներից, ինչպես անվանում են նրանց, մի քանիսը, այն է՝ մամռակենդանիները (Polyzoa), օժտված են ուշագրավ օրգաններով, այսպես կոչված ավիկուլարիաներով, որոնք զանազան տեսակների մոտ ունենում են շատ տարբեր կառուցվածք։ Ամենակատարյալ ձևով նրանք զարմանալի կերպով նմանվում են գառնանգղի փոքրածավալ կտուցին և գլխին, որը նստած է վզի վրա և ընդունակ է շարժվելու միանգամայն այնպես, ինչպես այդ հատուկ է ստորին ծնոտին։ Մի տեսակի մոտ, որը ես դիտել եմ, մեկ ճյուղի վրա նստած ավիկուլարիաները հաճախ հինգ վայրկյանի ընթացքում միաժամանակ շարժվում էին մերթ դեպի առաջ, մերթ դեպի ետ, բաց անելով իրենց ստորին ծնոտը մոտավորապես 90° անկյունով, և այդ շարժումից ցնցվում էր ամբողջ գաղութը։ Ծնոտները փշին շփվելիս նրանք այնքան պինդ էին բռնում նրան, որ կարելի էր ամբողջ ճյուղը ցնցել։
Մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] բերում է այդ օրինակը գլխավորապես այն կարծեցյալ դժվարության պատճառով, որ առաջ է գալիս բնական ընտրության ազդեցությամբ այդ, նրա կարծիքով, «էապես նման» օրգանների զարգացումը կենդանական թագավորության միմյանցից հեռու կանգնած երկու խմբերի մեջ բացատրելու տեսակետից։ Բայց որքան դա վերաբերում է կառուցվածքին, ես նմանություն չեմ տեսնում եռաճյուղ պեդիցելլյարիայի և ավիկուլարիայի միջև։ Վերջինս մի քիչ ավելի նման է խեցեմորթնեըի chelae-ին կամ բռնաչանչին, և մ-ր Մայվարտը նույն իրավունքով կարող էր բերել այդ նմանությունը որպես բացատրության համար դժվարին մի դեպք, ինչպես նաև նրանց նմանությունը թռչունի գլխի և կտուցի հետ։ Կենդանիների այդ խումբը մանրազնին կերպով ուսումնասիրած բնախույզների՝ մ-ր Բյոսկի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 163] դ-ր Սմիտտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 164] և դ-ր Նիտշեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 165] կարծիքով ավիկուլարիաները հոմոլոգ են զոոիդներին ու բջիջներին, որոնցից կառուցված են զոոֆիտները. բջջի շարժական կափարիչը համապատասխանում է ավիկուլարիայի շարժական ստորին ծնոտին։ Սակայն մ-ր Բյոսկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 163] չգիտե անցումներ, որոնք ներկայումս կարող էին գոյություն ունենալ զոոիդի և ավիկուլարիայի միջև, ուստի և չի կարելի դատել, թե ինչպիսի օգտակար անցումային աստիճաններով մի գոյացում կարող էր փոխարկվել մի այլ գոյացման. բայց դրանից երբեք չի հետևում, որ այդպիսի անցումներ գոյություն չեն ունեցել։
Քանի որ խեցեմորթների բռնաչանչերը որոշ չափով նման են ավիկուլարիաներին և երկու օրգանն էլ ծառայում են բռնելու համար, ուստի արժե ցույց տալ, որ առաջին օրգանի համար դեռ գոյություն ունի օգտակար անցումնային աստիճանների երկար շարք։ Առաջին և ամենապարզ ստադիայում վերջավորության վերջին հատվածը կամ ծռվում է դեպի նախավերջին լայն հատվածի լայն ուղիղ գագաթը, կամ դեպի նրա ամբողջ կողքի վրա, որով հնարավորություն է ստացվում առարկան ամուր կերպով բռնելու, բայց միևնույն ժամանակ ոտքը դեռ ծառայում է որպես տեղաշարժման օրգան։ Մոտակա ստադիայում նախավերջին լայն հատվածի վրա մենք գտնում ենք թեթևակի դուրս ցցված ելուստ, որը երբեմն օժտված է անկանոն ատամիկներով, և վերջին հատվածը ծռվում է դեպի այդ ելուստը։ Երբ այդ ելուստի չափերը մեծանում են, ընդ որում թե՛ նրա ձևը և թե՛ վերջին հատվածի ձևը թեթևակի փոփոխվում են և բարելավվում, բռնաչանչերը ավելի ու ավելի կատարելագործվում են, մինչև որ դառնում են այնքան կատարյալ գործիք, ինչպիսին են ծովային տասնոտանի խեցգետնու բռնաչանչերը. այս բոլոր անցումները կարելի է գտնել իրականության մեջ։
Բացի ավիկուլարիաներից, մամռակենդանիներն ունեն էլի մի հետաքրքրական օրգան, այսպես կոչված վիբրակուլները, որոնք սովորաբար լինում են երկար խոզանների տեսքով, ընդունակ են շարժվելու և հեշտությամբ գրգռվելու։ Իմ ուսումնասիրած մի տեսակի վիբրակուլները թեթևակի ծռված են և իրենց արտաքին եզրում ատամնաձև են. մեկ գաղութի բոլոր վիբրակուլները հաճախ շարժվում են միաժամանակ, գործելով երկար թիակների նման, նրանք արագ կերպով տեղաշարժում էին մի ճյուղ իմ մանրադիտակի տեսողության դաշտում։ Եթե ճյուղը զետեղվում էր նրա առջևի կողմում, վիբրակուլները խճճվում էին և մեծ ջանք էին գործադրում, որպեսզի ազատվեն։ Ենթադրում են, որ այգ օրգանները ծառայում են պաշտպանության համար և, ըստ մ-ր Բյոսկի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 163] դիտողության, «կարելի է տեսնել, թե ինչպես նրանք դանդաղորեն և զգուշությամբ սահում են մամռակենդանիների գաղութի մակերեսով, հեռացնելով այն, ինչ որ կարող է անհանգստացնել բջիջների քնքույշ բնակիչներին, երբ շոշափուկները դուրս են հանված»։ Ավիկուլարիաները վիբրակուլների նման հավանորեն նույնպես ծառայում են պաշտպանության համար, բայց նրանք կարող են նաև բռնել ու սպանել մանր ողջ կենդանիների, որոնք դրանից հետո, ինչպես կարծում են, ջրի շարժման միջոցով տարվում են ու հասնում զոոիդի շոշափուկներին։ Մի քանի տեսակներ օժտված են ավիկուլարիաներով և վիբրակուլներով, մի քանիսը՝ միայն ավիկուլարիաներով, քչերը՝ միայն վիբրակուլներով։
Հեշտ չէ պատկերացնել երկու այդքան տարբեր առարկաներ, ինչպիսիք են խոզանաձև վիբրակուլը և թռչունի գլխի նմանվող ավիկուլարիան. սակայն նրանք գրեթե հաստատապես հոմոլոգ են և զարգացել են միևնույն սաղմից, այն է՝ բջիջ ունեցող զոոիդից։ Սրանից մենք կարող ենք հասկանալ, ինչպես ինձ հաղորդում է մ-ր Բյոսկը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 163] թե ինչ կերպով այդ օրգանները որոշ դեպքերում, փոխարկվում են մեկը մյուսին։ Այսպես, Lepralia-ի մի քանի տեսակների ավիկուլարիաների շարժական ծնոտը այնպես զարգացած է և այնքան նման է խոզանի, որ կտուցի վերին կամ անշարժ մասի ներկայությունն է ծառայում որոշելու համար, թե այդ օրգանը ավիկուլարիա է։ Վիբրակուլները կարող էին զարգանալ նաև ուղղակի բջջի կափարիչից, նախապես չանցնելով ավիկուլարիայի ստադիան, բայց ավելի հավանական է, որ նրանք անցել են այդ ստադիայով, որովհետև ձևափոխման վաղ ստադիաների ընթացքում հազիվ թե զոոիդ պարունակող բջջի մյուս մասերը կարող էին միանգամից անհետանալ։ Շատ դեպքերում վիբրակուլները հենվում են առանձին հիմքի վրա, որն, ըստ երևույթին, ներկայացնում է կտուցի անշարժ մասը, թեև մի քանի տեսակներ այդ հիմքը բոլորովին չունեն։ Վիբրակուլների զարգացման այդ ընթացքը, եթե դա իրոք ճիշտ է, հետաքրքրական է, որովհետև եթե ենթադրենք, որ ավիկուլարիաներ ունեցող բոլոր տեսակները մահաջնջվել են, ապա ոչ ոք, նույնիսկ շատ վառ երևակայություն ունենալով, երբեք չէր մտածի, որ վիբրակուլները սկզբում գոյություն են ունեցել որպես թռչունի գլխի, անկանոն արկղի կամ գլխանոցի նմանվող օրգանի մասեր։ Հետաքրքիր է տեսնել, որ երկու այդքան տարբեր օրգաններ զարգացել են միևնույն սկզբնավորությունից, իսկ քանի որ բջջի շարժական կափարիչը ծառայում է զոոիդի պաշտպանության համար, ուստի դժվար չէ պատկերացնել, որ բոլոր աստիճանական անցումները, որոնց միջոցով շարժական կափարիչը սկզբում փոխարկվել է ավիկուլարիայի ստորին ծնոտի, իսկ հետո՝ երկարավուն խոզանի, միանման ձևով ծառայել են այս կամ այն եղանակով և զանազան պայմաններում պաշտպանության համար։
Բուսական թագավորության մեջ մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] կանգ է առնում միայն երկու դեպքի վրա, այն է՝ խոլորձիների ծաղիկների կառուցվածքի և փաթաթվող բույսերի շարժումների վրա։ Առաջինների վերաբերմամբ, ասում է նա «նրանց ծագման մեկնաբանությունը պետք է միանգամայն անբավարար, միանգամայն թերի համարել բացատրելու համար կառուցվածքի անսահմանորեն փոքր փոփոխությունները, որոնք օգտակար են դառնում միայն այն ժամանակ, երբ նրանք հասնում են զգալի զարգացման»։ Քանի որ այս խնդիրը ես քննարկում եմ ամբողջ լրիվությամբ մի այլ աշխատության մեջ, ուստի ես այստեղ կբերեմ միայն մի քանի դիտողություններ խոլորձիների ծաղիկների մեջ եղած ամենից ավելի զարմանալի առանձնահատկություններից մեկի, այն է՝ նրանց ծաղկափոշու զանգվածների (pollinia) կառուցվածքի վրա։ Ծաղկափոշու զանգվածն իր ամենաբարձր զարգացման ժամանակ ներկայացնում է ծաղկափոշու հատիկների մի գնդիկ, որտեղ փոշեհատիկները միացած են ճկուն կոթունի կամ caudicula-ի հետ, որն իր ծայրին կրում է խիստ կպչուն նյութի գնդիկ։ Այսպիսով, ծաղկափոշու այդ զանգվածները միջատների միջոցով տեղափոխվում են մի ծաղկից մյուսի սպիի վրա։ Մի քանի խոլորձիների ծաղկափոշու զանգվածները կոթուն չունեն և փոշեհատիկները միմյանց հետ միացած են բարակ թելիկներով. քանի որ այս վերջինները խոլորձիների բացառիկ առանձնահատկություններ չեն, ուստի կարելի էր այստեղ կանգ չառնել նրանց ծագման վրա. այնուամենայնիվ ես կնշեմ, որ Cypripedium-ի վրա, որ խոլորձիների սիստեմում գրավում է ամենացածր տեղը, մենք կարող ենք նկատել, թե այդ թելիկներն ինչպես են առաջացել սկզբում։ Մյուս խոլորձիների մոտ այդ թելիկները փոշեզանգվածի մի ծայրում մոտենում են միմյանց, որը և ներկայացնում է կոթունի առաջին կամ սկզբնական ստադիան։ Որ այդպիսին է կոթունի ծագումը, նույնիսկ երբ նա հասնում է զգալի երկարության ու զարգացման, մենք իրավունք ունենք եզրակացնելու ծաղկափոշու թերազարգացած հատիկների հիման վրա, որոնք երբեմն գտնվում են նրա կենտրոնական և ամուր մասերի ներսում։
Ինչ վերաբերում է երկրորդ գլխավոր առանձնահատկությանը, այսինքն, կոթունների ծայրին կպչող նյութի փոքրիկ գնդիկների ներկայությանը, ապա այստեղ կարելի է ցույց տալ զարգացման աստիճանների մի երկար շարան, որոնցից յուրաքանչյուրը օգտակար է բույսի համար։ Այլ ընտանիքների պատկանող ծաղիկների մեծամասնության սպին արտադրում է մի քիչ կպչուն նյութ։ Խոլորձիներից մի քանիսը նման մածական նյութ են արտադրում, բայց անհամեմատ ավելի մեծ քանակությամբ երեք սպիներից մեկի վրա, որը հավանորեն այդ առատ արտաթորման պատճառով, անընկալունակ է դառնում։ Երբ միջատը այցելում է այդպիսի ծաղկին, նա սրբում է այդ կպչուն նյութի մի մասը և միևնույն ժամանակ վերցնում է որոշ քանակությամբ ծաղկափոշու հատիկներ։ Սկսած այդ պարզ դրությունից, որը միայն քիչ տարբերվում է նրանից, ինչ որ հանդիպում է սովորական ծաղիկներից շատերի մեջ, մենք գտնում ենք անսահման անցումներ դեպի այնպիսի տեսակները, որոնց ծաղկափոշու զանգվածները վերջանում են շատ կարճ ազատ կոթունով, և դեպի այնպիսիները, որոնց կոթունը ամուր կապված է լինում կպչուն զանգվածի հետ, ըստ որում անընկալունակ սպին խորապես փոփոխված է։ Այս վերջին դեպքում մենք ունենք ծաղկափոշու զանգվածն իր ամենազարգացած և կատարյալ վիճակում։ Ով մանրազնին կերպով կուսումնասիրի խոլորձիների ծաղիկների կառուցվածքը, նա չի կարող ժխտել անցումների վերոհիշյալ շարքի գոյությունը՝ սկսած ծաղկափոշու հատիկների զանգվածից, որոնք միմյանց հետ միացած են միայն թելիկներով, և սպիից, որը հազիվ տարբերվում է սովորական ծաղկի սպիից, և վերջացրած վերին աստիճանի բարդ կառուցված ծաղկափոշու զանգվածով, որը զարմանալիորեն հարմարված է միջատների միջոցով տեղափոխության համար։ Նա չի կարող ժխտել նաև այն, որ բոլոր այդ աստիճանները համապատասխան տեսակների մոտ հիանալի կերպով հարմարված են ծաղկի ընդհանուր կառուցվածքին և նրա բեղմնավորմանը զանազան միջատների օգնությամբ։ Այս, ինչպես և համարյա ամեն մի այլ դեպքում, հետազոտությունը կարելի է տարածել ավելի ևս հեռու. կարող են հարցնել, հապա ի՞նչ կերպով սովորական ծաղկի սպին դարձել է մածուցիկ. բայց քանի որ էակների ոչ մի խմբի լրիվ պատմությունը մեզ հայտնի չէ, ապա նույնպես էլ անօգուտ է հարցնելը, ինչպես և անհուսալի է փորձել այդպիսի հարցերին պատասխանելը։
Այժմ դառնանք մագլցող բույսերին. նրանք կարող են դասվել մի երկար շարքում՝ սկսած նրանցից, որոնք պարզապես փաթաթվում են նեցուկի շուրջը, անցնելով նրանց, որոնք կառչում են տերևներով և վերջացրած նրանցով, որոնք օժտված են խիղբերով։ Այս վերջին երկու դասերի ցողունները սովորաբար, թեև ոչ միշտ, կորցնում են փաթաթվելու ընդունակությունը, բայց պահպանում են իրենց գագաթները պտտելու ընդունակությունը, որով օժտված են նաև խիղբերը։ Անցումները կառչող տերևներից դեպի խիղբեր ունեցողները զարմանալիորեն աստիճանական են, և շատ բույսեր կարող էին առանց խտրության դասվել այդ երկու դասերից ամեն մեկում։ Բայց սովորական փաթաթվող բույսերից դեպի տերևներով կառչող բույսերին անցնելիս առաջանում է բոլորովին մի նոր հատկություն, այն է՝ զգայունություն շփման նկատմամբ, որի հետևանքով տերևակոթերը և ծաղկակոթերը կամ խիղբերի ձևափոխված այդ օրգանները գրգռվում են, ծռվում և փաթաթվում են իրենց շփվող առարկային։ Նա, ով կկարդա իմ հետազոտությունը այդ բույսերի մասին, ես կարծում եմ, կընդունի, որ պարզ ու փաթաթվող բույսերի և խիղբերով օժտված բույսերի միջև եղած կառուցվածքի կամ գործունեության բոլոր բազմաթիվ անցումները յուրաքանչյուր մասնավոր դեպքում վերին աստիճանի օգտակար են նրանցով օժտված տեսակների համար։ Այսպես, օրինակ, ակներև է, որ փաթաթվող բույսի համար շատ օգտակար է իր տերևներով կառչելու ընդունակություն ձեռք բերելը և շատ հավանական է, որ երկար տերևակոթեր ունեցող փաթաթվող բույսը կձևափոխվեր կառչող տերևներ ունեցող բույսի, եթե միայն այդ տերևակոթերն ունենան շփման նկատմամբ հարկավոր աստիճանի զգայունություն։
Քանի որ գալարվելը նեցուկի վրայով սողալու ամենապարզ եղանակն է և փաթաթվող ցողունները ներկայացնում են ամենապարզ ձևը, որը կազմում է ամբողջ շարքի հիմքը, ապա բնականաբար հարց է ծագում՝ իսկ ինչպե՞ս են ձեռք բերել բույսերն այդ ընդունակությունը նրա սկզբնական վիճակում, որը հետագայում միայն զարգացել ու կատարելագործվել է բնական ընտրությամբ։ Փաթաթվելու ընդունակությունը կախված է, նախ, այն բանից, որ շատ մատաղ ցողուններ չափազանց հեշտությամբ ճկվում են, կորանում (մի առանձնահատկություն, որ ընդհանուր է նաև չփաթաթվող բույսերի շատ մեծ թվի համար), և երկրորդ, այն բանից, որ նրանք հաջորդաբար կորանում են հորիզոնի բոլոր կողմերի վրա, կրկնելով այդ շարժումները միևնույն կարպով. այդ շարժման հետևանքով նրանք թեքվում են բոլոր կողմերը և նրանց գագաթները գծում են շրջաններ։ Հենց որ ցողունի ստորին մասը դիպչում է որևէ առարկայի և դադարում է շարժվելուց, իսկ վերինը շարունակում է թեքվել և պտտվել, ապա նա, ըստ անհրաժեշտության, բարձրանում է դեպի վերև և փաթաթվում նեցուկի շուրջը։ Պտտական շարժումը դադարում է յուրաքանչյուր ընձյուղի սկզբնական աճեցողության հետ միասին։ Քանի որ պտտելու այդ ընդունակությամբ օժտված են բույսերի միմյանցից հեռու կանգնած ընտանիքների առանձին սեռեր և առանձին տեսակներ, ուստի ակներև է, որ նրանք փափաթվող բույսեր են դառել մեկը մյուսից բոլորովին անկախ կերպով, այլ ոչ թե մի ընդհանուր նախահորից ժառանգելու շնորհիվ։ Սրանից ես կարող էի նախագուշակել, որ այդպիսի շարժումների նկատմամբ թույլ հակումը պետք է ամենևին էլ հազվադեպ չլինի նաև չփաթաթվող բույսերի մեջ և որ այդ հակումը ծառայել է որպես հիմք, որը վաղուց մշակվել ու կատարելագործվել է բնական ընտրության միջոցով։ Երբ ես արեցի այդ կանխագուշակումը, ինձ հայտնի էր միայն մի շատ անկատար դեպք, այն է՝ Maurandia-ի մատաղ ծաղկակոթերը, որոնք թեթևակի և շատ անկանոն կերպով պտտվում են փաթաթվող բույսերի ցողունների նման, բայց դրանից իրենց համար ոչ մի օգուտ չեն քաղում։ Շատ չանցած՝ Ֆրից Մյուլլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] նկատել է, որ Alisma-ի և կտավատի՝ բնական սիստեմում մեկը մյուսից հեռու տեղ գրավող այդ չփաթաթվող բույսերի մատաղ ցողունները ակնհայտորեն պտտվում են, թեև անկանոն կերպով. նա ավելացնում է, թե հիմք ունի կասկածելու, որ այդ երևույթը գոյություն ունի նաև մի քանի այլ բույսերի մոտ։ Այդ թույլ շարժումները, ըստ երևույթին, ոչ մի նշանակություն չունեն հիշված բույսերի համար, համենայն դեպս նրանք ոչ մի առնչություն չունեն բույսի վեր մագլցելու ընդունակության հետ, որը և միմիայն մեզ հետաքրքրում է։ Ինչ էլ որ լինի, մենք կարող ենք պատկերացնել, որ եթե այդ բույսերի ցողունները հեշտությամբ ճկվել են և եթե իրենց գոյության պայմաններում նրանց ձեռնտու է եղել դեպի վեր մագլցելը, ապա աննշան ու անկանոն պտտվելու այդ սովորությունը ևս կարող էր ուժեղացվել ու օգտագործվել բնական ընտրությամբ այն աստիճան, որ այդ տեսակները կփոխարկվեն իսկական փաթաթվող բույսերի։
Տերևակոթերի, ծաղկակոթերի և խիղբերի գրգռականության վերաբերմամբ կիրառենք կշռադատության գրեթե նույն եղանակը, ինչ որ փաթաթվող բույսերի պտտական շարժումների վերաբերմամբ։ Քանի որ այդքան մեծ թվով տեսակներ, որոնք պատկանում են մեկը մյուսից հեռու կանգնած խմբերի, ունեն այդպիսի գրգռականություն, ապա մենք պետք է սպասենք, որ կհանդիպենք նրանց սաղմնային վիճակում նաև այնպիսի բույսերի վրա, որոնք չեն դարձել փաթաթվող։ Այդ էլ նկատվում է իրականության մեջ։ Ես նկատել եմ, որ սրանից առաջ հիշատակած Maurandia-ի մատաղ ծաղկակոթերը թեթևակի ծռվում էին դեպի այն կողմը, որ կողմից նրանց հետ շփվում էին։ Մորրենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 166] նկատել է, որ թթվառվույտի մի քանի տեսակների տերևները և տերևակոթերը, հատկապես երբ նրանք կիզիչ արևի տակ են գտնվել, կրկնվող զգուշավոր հպումների կամ ցնցումների ժամանակ ցուցաբերել են շարժումներ։ Ես կրկնել եմ այդ փորձերը նույնպիսի հաջողությամբ թթվառվույտի այլ տեսակների վրա. մի քանիսի շարժումը շատ ակներև էր, նա ամենից լավ նկատվում էր մատաղ տերևների վրա. այլ տեսակների շարժումները չափազանց աննշան էին։ Շատ կարևոր է այն փաստը, որ հաստատել է այնպիսի բարձր հեղինակություն, ինչպիսին է Հոֆմեյստերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 167] որ բոլոր բույսերի մատաղ ընձյուղները և տերևները ցուցաբերում են շարժում՝ որպես ցնցումների հետևանք. և մենք գիտենք, որ փաթաթվող բույսերի տերևակոթերը և խիղբեըը գրգռականություն են ցուցաբերում միայն աճման վաղ ստադիաներում։
Հազիվ թե կարելի է ընդունել, որ նման հազիվ նշմարելի շարժումները, որոնք առաջանում են շփումից կամ ցնցումից մատաղ աճող օրգաններում, կարող են ունենալ որևէ ֆունկցիոնալ նշանակություն։ Բայց բույսը զանազան արտաքին գրգիռների ազդեցության տակ ընդունակություն ունի ցուցաբերելու շարժումներ, որոնց նշանակությունը ակներև է. այդպիսիներն են, օրինակ, շարժումները դեպի լույսը և ավելի հազվադեպ է լույսին հակառակ, շարժումները ծանրության ուժի ներգործության հակառակ ուղղությամբ, կամ ավելի հազվադեպ՝ նրա ներգործության ուղղությամբ։ Երբ կենդանու նյարդերը և մկանները գրգռվում են գալվանականությամբ կամ ստրիխնինի ներգործությամբ, դրանից առաջացող շարժումները կարող են անվանվել պատահական, որովհետև նյարդերը և մկանները, ի հարկե, հատկապես այդ գրգիռներին հարմարված չեն։ Այդպես են նաև բույսերը, որոշ գրգիռների ազդեցության տակ շարժվելու ընդունակությունն ունենալով՝ նրանք պատահականորեն գրգռվում են նաև շփումից կամ ցնցումից։ Հետևաբար, կարելի է առանց դժվարության ընդունել նաև տերևներով ու խիղբերով կառչող բույսերի վերաբերմամբ, որ նրանց մեջ ևս այդ ընղհանուր ընդունակությունը աջակցություն է ստացել ու զարգացել բնական ընտրության միջոցով։ Սակայն իմ հետազոտության մեջ շարադրածս պատճառների հիման վրա հավանական է թվում, որ այդ բոլորը կարող է տեղի ունենալ միայն բույսերի հետ, որոնք արդեն ունեն պտտվելու ընդունակություն և, հետևաբար, արդեն դարձել են փաթաթվող։
Ես արդեն փորձել եմ բացատրել, թե ինչպես բույսերը դարձել են փաթաթվող այն ընդունակության ուժեղացման շնորհիվ, որի միջուկով նրանք կատարել են թույլ և անկանոն, սկզբում ոչ մի օգուտ չներկայացնող պտտական շարժումներ, այդ շարժումները, ինչպես նաև շփումից ու ցնցումից առաջացողները, իրենց հերթին հանդիսացել են շարժվելու ընդհանուր ընդունակության պատահական հետևանք, որ ձեռք է բերվել բոլորովին այլ և օգտակար նպատակների համար։ Հանձն չեմ առնում լուծելու այն հարցը, թե փաթաթվող բույսերի աստիճանական զարգացումը բնական ընտրության միջոցով արդյո՞ք տեղի է ունեցել վարժության ժառանգական հետևանքների աջակցությամբ, բայց մեզ հայտնի է, որ մի քանի պարբերական շարժումներ, ինչպես, օրինակ, այսպես կոչված բույսերի քունը կախված է նրանց կառուցվածքից։
Ես քննարկեցի բավականաչափ, գուցե և նույնիսկ ավելի քան բավականաչափ դեպքեր, որոնք խնամքով ընտրվել են հմուտ բնախույզի կողմից որպես ապացույց, որ անհնարին է բնական ընտրության օգնությամբ բացատրել օգտակար հարմարումների սաղմնային ստադիաները, և ես հույսով եմ, թե ցույց տվեցի, որ այդ տեսակետից ոչ մի լուրջ դժվարություն չի հանդիպում։ Միաժամանակ լավ առիթ ներկայացավ մի քիչ ավելի մանրամասն կանգ առնելու կառուցվածքի աստիճանական այն տարբերությունների հարցի վրա, որ հաճախ կապված են գործունեությունների ձևափոխման հետ, մի կարևոր հարց, որը բավանակաչափ մանրամասնությամբ չի զարգացվել այս աշխատության նախորդ հրատարակություններում։ Ես այժմ համառոտակի կկրկնեմ բերված օրինակները։
Ընձուղտի վերաբերմամբ կասենք՝ շարունակ պահպանվել են մի ինչ որ անհետացած բարձրահասակ որոճողի անհատները, որոնք ունեցել են ամենաերկար վիզ, ոտներ և այլն և կրծել են միջին բարձրությունից քիչ ավելի բարձր բույսեր, ինչպես նաև շարունակ ոչնչացել են անհատներ, որոնք չեն կարողացել այդքան բարձր բույսեր կրծել, այս կարող է լիովին բացատրել այդ զարմանալի չորքոտանու գոյացումը։ Միասին վերցրած այղ բոլոր մասերի երկարատև վարժությունը՝ կապված ժառանգականության հետ, զգալի չափով կարող էր նպաստել նրանց կոորդինացիային։ Այն միջատների վերաբերմամբ, որոնք նմանվում են զանազան առարկաների, անհավանական չի թվա, որ բնական ընտրության ներգործության համար ելանյութ է հանդիսացել պատահական նմանությունը մի որևէ սովորական առարկայի հետ։ Այդ նմանությունը կարող էր շարունակ ավելի ու ավելի մեծանալ պահպանելով իրեն մեծացնող աննշան փոփոխությունները, ընդ որում այդ պրոցեսը շարունակվել է այնքան, մինչև որ միջատը փոփոխվել է, մինչև որ ավելի ու ավելի կատարյալ նմանությունը կարողացել է ծառայել որպես միջոց, որով նա պաշտպանվել է շատ սրատես թշնամիների հետապնդումից։ Կետերի մի քանի տեսակների մոտ երևան է գալիս քիմքի վրա անկանոն, կետերի ձևով դասավորված եղջերային կույտեր առաջացնելու ձգտում, միանգամայն հնարավոր է, որ բնական ընտրությունը պահպանել է բոլոր բարենպաստ փոփոխությունները, մինչև որ այդ կույտերը դարձել են սկզբում շերտավոր թմբիկներ կամ ատամիկներ, նման նրանց, որոնք հանդիպում ենք սագի կտուցի մեջ, հետո կարճ թիթեղիկներ, ինչպես ընտանի բադինն է, այնուհետև նույնքան կատարյալ թիթեղիկներ, որքան լայնակտուցինն է, և վերջապես, այսպես կոչված կետոսկրի հսկայական թիթեղներ գրենլանդական կետի բերանում։ Բադերի ընտանիքում այդ թիթեղիկները գործ են ածվում սկզբում որպես ատամներ, այնուհետև մասամբ որպես ատամներ, մասամբ որպես քամող ապարատ և, վերջապես, բացառապես միայն այս վերջին նպատակի համար։
Այդպիսի օրգանների՝ ինչպես հենց նոր հիշատակած եղջերային թիթեղիկի կամ կետոսկրի առաջացման ժամանակ, որքան մենք կարող ենք դատել, սովորությունները կամ վարժությունը միայն շատ քիչ կարող էին նպաստել կամ նույնիսկ բոլորովին չեն նպաստել կատարելագործման պրոցեսին։ Մյուս կողմից, ներքևի աչքի տեղափոխումը դեպի գլխի վերևի կողմը կամ կառչուն պոչի առաջացումը կարելի է գրեթե բացառապես վերագրել անընդհատ վարժությանը՝ շաղկապված ժառանգականության հետ։ Բարձրակարգ կենդանիների պտուկների վերաբերմամբ ամենահավանական ենթադրությունն այն է, որ սկզբում ենթամաշկային գեղձերը պարկավորների պարկի ամբողջ մակերեսի վրա արտաթորել են սննդարար հեղուկ և որ հետո այդ գեղձերը կատարելագործվել են բնական ընտրության միջոցով և կենտրոնացել են որոշ սահմանափակ մակերեսների վրա, որով և պտուկների առաջացման հիմք է դրվել։ Հին փշամորթների մոտ պաշտպանության համար ծառայող ճյուղավոր փշերի ժամանակակից եռաբաժան պեդիցելլյարիաների ձևափոխվելու բացատրությունը նույնպես քիչ դժվարություն է ներկայացնում, ինչպես որ խեցեմորթների բռնաչանչերի առաջացումը սկզբում բացառապես շարժման համար նախանշված վերջավորությունների վերջին ու նախավերջին հատվածներից։ Մամռակենդանիների ավիկուլարիաները և վիբրակուլները ներկայացնում են իրենց արտաքինով խիստ տարբերվող, բայց մեկ աղբյուրից առաջացած օրգաններ, և վիբրակուլների վերաբերմամբ մենք կարող ենք ըմբռնել, թե ինչ օգուտ են բերել զարգացման միջակա ստադիաները։ Ինչ վերաբերում է խոլորձիների ծաղկափոշու զանգվածներին, ապա կարելի է հետամուտ լինել, թե ինչպես թելիկները, որ սկզբում կապում են միմյանց հետ ծաղկափոշու հատիկները, կարող են իրար հետ միաձուլվել ու կազմել կոթուն, ճիշտ այդպիսի ստադիաների միջոցով կարելի է հետամտել, թե ինչպես կպչուն նյութը, որը նման է սովորական ծաղիկների սպիների վրա արտադրվող և գրեթե նույն նպատակին ծառայող նյութին, կապված է լինում կոթունների ազատ վերջավորությունների հետ, ընդ որում այդ բոլոր անցումային աստիճանները ակներևորեն օգտակար են նրանցով օժտված բույսերին։ Փաթաթվող բույսերի մասին ես ավելորդ եմ համարում կրկնել այն, ինչ որ ասվել է քիչ առաջ։
Հաճախ հարց է հարուցվել, թե՝ եթե բնական ընտրությունը այդպես հզոր ուժ է ներկայացնում, հապա ինչո՞ւ նա չի օժտել որոշ տեսակների այս կամ այն օրգանով, որ հավանորեն կարող էր նրանց օգտակար լինել։ Սակայն միանգամայն անխոհեմություն է նման հարցերին որոշակի պատասխան սպասելը, հիշելով, թե որքան խորն է մեր անգիտակությունը ամեն մի տեսակի անցյալ պատմության և այն պայմանների մասին, որոնք ներկայումս որոշում են նրա թիվը և տարածման սահմանները։ Մեծ մասամբ կարելի է նշել այդ երևույթների ընդհանուր և միմիայն հազվագյուտ դեպքերում մերձավոր պատճառները։ Մի որևէ տեսակ որպեսզի հարմարվի նոր կենսակերպի, անհրաժեշտ են բազմաթիվ միաժամանակ տեղի ունեցող փոփոխություններ, և հաճախ կարող էր պատահել, որ համապատասխան մասերում հարկավոր ուղղությամբ կամ հարկավոր չափով փոփոխություններ չեն դրսևորվել։ Շատ տեսակների բազմացումը կարող էր դադարել այն գործոնների քայքայիչ ներգործության հետևանքով, որ ոչ մի առնչություն չունեն այնպիսի օրգանների հետ, որոնք մեզ թվում են օգտակար և կարծես կարող էին առաջանալ բնական ընտրության միջոցով։ Այդպիսի դեպքերում, քանի որ գոյության կռիվը չէր պայմանավորվել այդ օրգաններով, նրանք չէին կարող մշակվել բնական ընտրությամբ։ Հաճախ որոշ օրգանի զարգացման համար կարող էին անհրաժեշտ լինել միանգամայն յուրահատուկ բարդ և երկարատև պայմաններ, իսկ այդպիսի պայմաններ կարող էին միայն շատ հազվագյուտ դեպքում իրագործվել։ Այն ենթադրությունը, թե տվյալ յուրաքանչյուր օրգան, որը մեր, հաճախ սխալ կարծիքով կարող էր օգտակար լինել տեսակի համար, պետք է բոլոր հանգամանքներում առաջանա ր բնական ընտրության շնորհիվ, ուղղակի հակասում է նրան, ինչ մեզ հայտնի է այդ սկզբունքի ներգործության եղանակի մասին։ Միստր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] չի ժխտում, որ բնական ընտրությունը որոշ բան արել է, բայց նա կարծում է, որ կարելի է ապացուցել նրա «ակնհայտ անբավարարությունը» ամբողջ մի շարք երևույթների բացատրության համար, որոնք ես բացատրում եմ բնական ընտրությամբ։ Նրա ամենագլխավոր փաստարկներն այստեղ քննարկվել են, իսկ մնացածները ուշադրության կառնվեն հետագայում։ Ինձ թվում է, որ նրանք ապացուցականություն չունեն և շատ քիչ կշիռ են ներկայացնում նրանց համեմատությամբ, որոնք առաջադրվում են հօգուտ բնական ընտրության ներգործության, որին օժանդակում են նաև այլ գործոններ, որոնց մասին հաճախ է հիշատակվել։ Իմ պարտքն եմ համարում նկատել, որ վերևում իմ շարադրած փաստերից ու ապացույցներից մի քանիսը նույն նպատակով արդեն բերվել են մի հիանալի հոդվածում, որը վերջերս լույս է տեսել «Medico-Chirurgical Review»[56]-ում։
Ներկայումս համարյա բոլոր բնախույզներն ընդունում են էվոլյուցիան այս կամ այն ձևով։ Մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] ենթադրում է, որ տեսակները փոփոխվում են և այդ փոփոխությունը առաջանում է ինչ որ «ներքին ուժով կամ ձգտմամբ», որի բնույթի մասին ոչինչ հայտնի չէ։ Որ տեսակները փոփոխվելու ընդունակություն ունեն, այդ ընդունում են բոլոր էվոլյուցիոնիստները, բայց, իմ կարծիքով, ոչ մի կարիք չկա հրավիրելու որևէ ներքին ուժի միջամտությունը, բացի սովորական փոփոխականության հակումից, որը արհեստական ընտրության աջակցությամբ, մարդու մասնակցությամբ սկիզբ է տվել հիանալի կերպով հարմարված ընտանի ցեղերին, իսկ բնական ընտրության աջակցությամբ նա նույնպես կարող էր աննկատելի անցումների միջոցով առաջացնել բնական ցեղեր, կամ տեսակներ։ Վերջնական հետևանքում, ինչպես արդեն բացատրվել է, ստացվել է կազմվածքի ընդհանուր առաջադիմություն, իսկ քիչ դեպքերում նաև հետադիմություն։
Մ-ր Մայվարտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] այնուհետև ըստ երևույթին հակվում է դեպի այն կարծիքը, որին համաձայն են նրա հետ նաև մի քանի բնախույզներ, որ նոր տեսակները երևան են եկել «նաև միանգամից առաջացած փոփոխությունների միջոցով»։ Այսպես, օրինակ, նա ենթադրում է, որ մահաջնջված եռամատ հիպարիոնի և ձիու միջև եղած տարբերությունը հանկարծակի է առաջացել։ Նա դժվարանում է ենթադրել, որ թռչունի թևը, «կարող էր առաջանալ այլ կերպ, քան խիստ արտահայտված ու կարևոր հատկության համեմատաբար հանկարծակի փոփոխությամբ»․ ըստ երևույթին, նույն տեսակետը նա պատրաստ է տարածելու նաև չղջիկի և պտերոդակտիլի թևի վրա։ Այդ եզրակացությունը, որ հարկադրում է ենթադրելու հսկայական ընդմիջումներ զարգացման վարընթաց շարքերում, ինձ վերին աստիճանի անհավանական է թվում։
Էվոլյուցիայի դանդաղության ու հաջորդականության մեջ համոզված յուրաքանչյուր ոք անշուշտ կընդունի, որ տեսակների փոփոխությունները կարող էին նույնքան ցայտուն և մեծ լինել, որքան այն եզակի փոփոխություններից ամեն մեկը, որ մենք հանդիպում ենք բնության մեջ կամ նույնիսկ արհեստական ցեղերում։ Բայց քանի որ ընտանեցված և մշակովի վիճակում տեսակներն ավելի փոփոխական են, քան բնական վիճակում, ուստի քիչ հավանական է թվում, որ բնության մեջ նույնպես հաճախ հանդիպեն այդպիսի ցայտուն և հանկարծական փոփոխություններ, ինչպիսիք երբեմն հանդիպում են ընտանեցված ու մշակովի ձևերի մեջ։ Այս վերջին փոփոխություններից շատերը պետք է վերադրել վերադարձին դեպի նախկին հատկանիշները, իսկ այս վերջինները, հավանորեն, սկզբում շատ դեպքերում ձեռք են բերվել աստիճանաբար։ Ավելի մեծ թվով փոփոխություններ պետք է դասել այլանդակությունների շարքը. այդպիսիներն են վեցմատնանի մարդիկ, խոզանանման մազերով մաշկ ունեցող մարդիկ, անկոնյան ոչխարները, նիատային եղջերավոր անասունները և այլն, և քանի որ նրանք խիստ տարբերվում են բնական տեսակներից, այդ պատճառով էլ շատ քիչ լույս են սփռում մեզ հետաքրքրող հարցի վրա։ Եթե հանկարծակի փոփոխության նման դեպքերը բացառենք, ապա մնացած դեպքերը, եթե նրանք գտնվել են բնության մեջ, կդասվեին կասկածելի տեսակների թվում, որոնք սերտ կերպով կապված են իրենց նախասկզբնական տիպերի հետ։
Այն ապացույցները, որոնք ինձ հարկագրում են կասկածել նրանում, որ բնական տեսակները կարող էին փոփոխվել այնպես հանկարծակի կերպով, ինչպես այդ պատահում է ընտանի ցեղերի հետ, և հարկադրում են ինձ վերջնականապես ժխտել նրանց փոփոխությունների այդ հրաշագործ եղանակը, որ ենթադրում է մ-ր Մայվարտը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] հետևյալներ են։ Մեր փորձի հիման վրա հանկարծական և խիստ արտահայտված փոփոխությունները մեր ընտանի ցեղերի մեջ երևան են գալիս որպես եզակի դեպքեր և ժամանակի երկար ընդմիջումներից հետո։ Եթե այդպիսի փոփոխությունները երևան են եկել բնական վիճակում, նրանք, ինչպես պարզաբանվել է առաջ, շատ հեշտությամբ կանհետանային պատահական քայքայիչ պատճառների և ձևերի հետագա խաչաձևման հետևանքով, նույնը ճիշտ է նաև ընտանի ցեղերի նկատմամբ, եթե միայն այդ հանկարծական փոփոխությունները առանձին խնամքով չեն պահպանվում և ընտրվում մարդու հոգատարությամբ։ Ուրեմն մ-ր Մայվարտի մատնանշած եղանակով տեսակների հանկարծական առաջացումն ընդունելու, համար պետք է հակառակ բոլոր անալոգիաներին ընդունենք, որ միաժամանակ միևնույն մարզում այդպիսի հրաշագործ եղանակով առաջացել են մի քանի փոփոխված անհատներ։ Այդ դժվարությունը, ինչպես որ մարդու կատարած անգիտակցական ընտրության դեպքում, վերացվում է աստիճանական էվոլյուցիայի տեսության տեսակետից այն ենթադրությամբ, որ մեծ թվով անհատներ ավելի կամ պակաս փոփոխվել են բարենպաստ ուղղությամբ և պահպանվել, իսկ հակառակ ուղղությամբ փոփոխված մեծ թվով անհատներ ենթարկվել են ոչնչացման։
Կասկած չի կարող լինել, որ զգալի թվով տեսակներ առաջացել են չափազանց դանդաղ և աստիճանական փոփոխության միջոցով։ Մի քանի բնական մեծ ընտանիքների տեսակները և նույնիսկ սեռերն ունեն ազգակցության այնպիսի սերտ աստիճան, որ նրանց մեջ դժվար զանազանելիները քիչ չեն։ Յուրաքանչյուր մայր ցամաքում, շարժվելով հյուսիսից դեպի հարավ կամ հարթավայրից դեպի բարձունքները, մենք հանդիպում ենք բազմաթիվ իրար մոտ ազգակից կամ այսպես կոչված փոխարինող տեսակների. նույնը մենք ունենք նաև որոշ մեկուսացած մայր ցամաքների վրա, որոնք, ինչպես մենք հիմք ունենք ենթադրելու, առաջ միացած են եղել միմյանց հետ։ Արտահայտելով այս և հետագա դիտողությունները, ես ստիպված եմ վկայակոչել որոշ տվյալներ, որոնք կքննարկվեն հետագայում[57]։ Մի հայացք գցեք որևէ մայր ցամաք եզրապատող և ափից հեռու գտնվող շատ կղզիների վրա և ուշադրություն դարձրեք, թե ինչպես նրանց բնակիչների զգալի քանակը կարելի է բարձրացնել միայն մինչև կասկածելի տեսակների աստիճանի։ Ճիշտ այդպես էլ է, երբ մենք աչքի ենք անցնում անցյալը և միևնույն մարզի սահմաններում ասպարեզից հենց նոր հեռացած տեսակները համեմատում ենք դեռ գոյություն ունեցող տեսակների հետ, կամ եթե մենք համեմատում ենք միմյանց հետ բրածո տեսակները, որոնք պարփակված են միևնույն երկրաբանական ֆորմացիայի հաջորդող կեղևաշերտերում։ Ակնհայտ է, որ բազմաթիվ տեսակներ սերտ կերպով կապված են դեռ գոյություն ունեցող կամ դեռ վերջերս գոյություն ունեցած տեսակների հետ, և դրանց վերաբերմամբ անշուշտ ոչ ոք չի պնդի, թե նրանք առաջացել են հանկարծակի կերպով։ Ինչպես և չպետք է մոռանալ, որ երբ մենք ուշադրություն ենք դարձնում միմյանց մերձավոր առանձին տեսակների կառուցվածքի առանձին մասերի վրա, այլ ոչ թե առանձին տեսակների վրա, այդ դեպքում մենք կարող ենք հետամուտ լինել բազմաթիվ և զարմանալի նուրբ անցումների, որոնք շաղկապում են միմյանց հետ խիստ տարբեր օրգաններ։
Փաստերի բազմաթիվ խմբերը հասկանալի են դառնում, եթե միայն ելնում ենք այն սկզբունքից, որ տեսակները փոփոխվել են շատ անզգալի աստիճաններով։ Այսպիսին է, օրինակ, այն փաստը, որ մեծ սեռերին պատկանող տեսակները ավելի մեծ չափով են նմանվում իրար և ունեն ավելի մեծ թվով տարատեսակներ, քան փոքր սեռերի տեսակները։ Առաջին կարգի տեսակները նույնպես կենտրոնացած են փոքրիկ խմբերում այնպես, ինչպես տարատեսակները խմբվում են տիպական տեսակային ձևերի շուրջը, նրանք ներկայացնում են նաև այլ համանմանություններ տարատեսակների հետ, ինչպես այդ ցույց է տրված մեր գրքի երկրորդ գլխում։ Այս սկզբունքի հիման վրա մենք կարող ենք բացատրել, թե ինչու տեսակային հատկանիշներն ավելի փոփոխական են, քան սեռային հատկանիշները, և ինչու անսովոր կերպով և անսովոր աստիճանի զարգացած մասերը նույնպես ավելի փոփոխական են, քան մնացած մասերը։ Այստեղ կարելի էր բերել անալոգ շատ փաստեր, որոնք սէալիս են նման ցուցումներ։
Թեև շատ բազմաթիվ տեսակներ գրեթե հաստատապես առաջացել են հաջորդական աստիճաններով, որոնց տարբերությունները ավելի մեծ չեն, քան ամենանուրբ տարատեսակների միջև եղած տարբերությունները, այնուամենայնիվ կարելի է ընդունել, որ այլ դեպքերում անցումը կարող էր կատարվել մի այլ, ավելի հանկարծական եղանակով, բայց այդպիսի ենթադրությունը չի կարելի ընդունել առանց պարզորոշ ակներևություն ներկայացնելու։ Հեռավոր և շատ դեպքում սխալ անալոգները, ինչպես այդ ցույց է տվել Չոնսեյ Ռայտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 146] (Chauncey Wright), որոնք ներկայացված են ի պաշտպանություն այդ տեսակետի, ինչպես, օրինակ, անօրգանական նյութերի բյուրեղացման հանկարծականությանը կամ մի բազմակողմանու փոխարկվելը մի ուրիշի՝ նրա կողմերի փոփոխության միջոցով, հազիվ թե արժանի են ուշադրության։ Փաստերի միայն մեկ կատեգորիան, այն է՝ կյանքի այն նոր ձևերի երևան գալը, որոնք տարբերվում են մնացածներից մեր երկրաբանական ֆորմացիաներում, առաջին հայացքից կարծես թե հաստատում է այդ ենթադրությունը հանկարծական փոփոխության մասին, բայց այդ վկայությունների համոզարարությունը կախված է բացառապես երկրի պատմության հեռավոր ժամանակաշրջանների վերաբերյալ երկրաբանական տարեգրության կատարելության աստիճանից։ Եթե այդ տարեգրությունն այդքան կցկտուր է, ինչպես հաստատում են շատ երկրաբաններ, ապա զարմանալի ոչինչ չկա, որ շատ ձևեր մեզ թվում են հանկարծակի կերպով զարգացած։
Եթե միայն մենք չընդունենք այն ձևափոխությունները, որ նույնքան հրաշալի են, ինչպես մ-ր Մայվարտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] պաշտպանածները, ինչպիսիք են թռչունի կամ չղջիկի թևի հանկարծակի երևան գալը, կամ Հիպարիոնի նույնպիսի հանկարծակի փոխարկվելը ձիու, ապա ես չեմ կարծում, որ հանկարծական ձևափոխությունների մասին եղած ենթադրությունը որևէ լույս կսփռեր մեր երկրաբանական տարեգրության մեջ շաղկապող օղակների բացակայության վրա։ Հանկարծական փոփոխությունների գոյությանը համոզվելու դեմ բողոքում է նաև սաղմնաբանությունը։ Հասկանալի է այն փաստի կարևորությունը, որ թռչունի կամ չղջիկի թևերը և ձիու կամ որևէ այլ չորքոտանու ոտքերը սաղմի զարգացման վաղ շրջանում միմյանցից չեն տարբերվում և, միայն ավելի ուշ դիֆերենցվում են աստիճանաբար համարյա անզգալի աստիճաններով։ Սաղմնաբանական նմանության բազմապիսի գծերը կարելի է, ինչպես մենք հետագայում կտեսնենք, բացատրել նրանով, որ գոյություն ունեցող տեսակների նախնիները փոփոխվել են ոչ թե իրենց զարգացման ամենավաղ ժամանակաշրջանում, այլ իրենց ձեռք բերած հատկանիշները փոխանցել են իրենց հետնորդներին համապատասխան հասակում։ Ուրեմն՝ ներգործությունը համարյա չի շոշափում սաղմի զարգացման վաղ ստադիաները, և սաղմը մնում է կարծես վկա այն անցյալ վիճակների, որոնցով անցել է տեսակը։ Դրանով է բացատրվում այն փաստը, որ գոյություն ունեցող տեսակները իրենց զարգացման վաղ ստադիաների ընթացքում այնքան հաճախ նմանվում են նույն դասի ավելի հին ու անհետացած ձևերին։ Սաղմնաբանական նմանության տեսակետից, բայց ըստ էության ամեն մի այլ տեսակետից էլ, անհավանական է թվում, որ կենդանին կարող էր ենթադրվել այնպիսի ակնթարթային ու հանկարծական ձևափոխությունների, որոնց մասին խոսվել է, և որ նրա զարգացման սաղմնային ստադիաները չպահպանեին այդ հանկարծական ձևափոխությունների հետքերը. կառուցվածքի յուրաքանչյուր ամենաաննշան գիծ զարգանում է աստիճանաբար, անզգալի աստիճաններով։
Նա, ով կարծում է, թե որևէ հին ձև ինչ որ ներքին ձգտման կամ ուժի շնորհիվ փոփոխվել է հանկարծակի կերպով, օրինակ՝ ստացել է թևեր, պետք է հակառակ ամեն մի անալոգիայի ընդունի, որ այդ փոփոխությունը միաժամանակ ընդգրկել է մեծ թվով անհատներ։ Անհնարին է ժխտել, որ կազմվածքի նման հանկարծակի և խոր փոփոխությունները խիստ կերպով հակասում են այն բանին, ինչ որ իրականում տեղի է ունեցել տեսակների մեծամասնության վերաբերմամբ։ Նա այնուհետև ստիպված կլիներ ընդունելու, որ կազմվածքի բազմաթիվ գծերը, որոնք հիանալի կերպով հարմարված են միևնույն էակի մյուս մասերին և նրան շրջապատող պայմաններին, նույնպես հանկարծակի են առաջացել, և այդ բարդ ու հրաշալի փոխադարձ հարմարումների համար նա ի վիճակի չի լինի առաջարկելու որևէ բացատրության գեթ նշույլն անգամ։ Նա ստիպված կլինի ընդունելու, որ այդ խոր հանկարծական փոփոխությունները իրենց ներգործության ամենափոքր հետքն անգամ չեն թողել սաղմի վրա։ Ընդունել այդ բոլորը, ինձ թվում է, կնշանակեր անցնել հրաշքների բնագավառը, լքելով գիտության բնագավառը։
ԳԼՈՒԽ VIII։ ԲՆԱԶԴ
ԲՆԱԶԴՆԵՐԸ ՆՄԱՆ ԵՆ ՍՈՎՈՐՈՒԹՅՈՒՆՆԵՐԻՆ, ԲԱՅՑ ՆՐԱՆՑԻՑ ՏԱՐԲԵՐՎՈՒՄ ԵՆ ԾԱԳՄԱՄԲ։— ԲՆԱԶԴՆԵՐԸ ԿԱՏԱՐԵԼԱԳՈՐԾՎՈՒՄ ԵՆ ԱՍՏԻՃԱՆԱԲԱՐ։— ԽՈՏԵՐԻ ԼՎԻՃՆԵՐԸ ԵՎ ՄՐՋՅՈՒՆՆԵՐԸ։— ԲՆԱԶԴՆԵՐԸ ՓՈՓՈԽԱԿԱՆ ԵՆ։ ԸՆՏԱՆԻ ԿԵՆԴԱՆԻՆԵՐԻ ԲՆԱԶԴՆԵՐԸ, ՆՐԱՆՑ ԾԱԳՈՒՄԸ։— ԿԿՎԻ, MOLOTHRUS-Ի, ՋԱՅԼԱՄԻ ԵՎ ՄԱԿԱԲՈՒԾԱՅԻՆ ՄԵՂՈՒՆԵՐԻ ԲՆԱԿԱՆ ԲՆԱԶԴՆԵՐԸ։ ՍՏՐԿԱՏԵՐ ՄՐՋՅՈՒՆՆԵՐ։— ՄԵՂՈՒՆ, ՆՐԱ ՇԻՆԱՐԱՐԱԿԱՆ ԲՆԱԶԴԸ։— ԲՆԱԶԴԻ ԵՎ ԿԱԶՄՎԱԾՔԻ ՓՈՓՈԽՈՒԹՅՈՒՆՆԵՐԸ ԱՆՀՐԱԺԵՇՏ ՉԷ, ՈՐ ՄԻԱԺԱՄԱՆԱԿ ԼԻՆԵՆ։ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՏԵՍՈՒԹՅԱՆ ԴԺՎԱՐՈՒԹՅՈՒՆՆԵՐԸ ԲՆԱԶԴՆԵՐԻ ՆԿԱՏՄԱՄԲ ԿԻՐԱՌԵԼՈՒ ԴԵՊՔՈԻՄ։— ԱՆՍԵՌ ԿԱՄ ԱՆՊՏՈՒՂ ՄԻՋԱՏՆԵՐ։— ԵԶՐԱԿԱՑՈՒԹՅՈՒՆ։
Շատ բնազդներ այնքան զարմանալի են, որ նրանց զարգացումը հավանորեն ընթերցողին կթվա մի դժվարություն, որը կբավարարի իմ ամբողջ տեսությունը խորտակելու համար։ Ես այստեղ կարող եմ կանխապես ասել, որ մտավոր ընդունակությունների ծագման հարցով չեմ զբաղվում, ինչպես որ չեմ զբաղվել նաև բուն կյանքի ծագման հարցով։ Մեզ վերաբերում է միայն միևնույն դասին պատկանող կենդանիների բնազդների և այլ մտավոր ընդունակությունների բազմազանությունը։
Ես չպիտի փորձեմ տալ բնազդի որևէ սահմանում։ Հեշտ կլիներ ցույց տալ, որ այդ անվան տակ սովորաբար հասկանում են տարբեր մտավոր գործողություններ, բայց յուրաքանչյուր ոք հասկանում է, թե ինչ են ուզում ասել, երբ խոսում են, որ բնազդն ստիպում է կկուին կատարելու իր թռիչքները և ձու ածելու այլ թռչունների բներում։ Մի այնպիսի գործողություն, որը մենք կարող ենք կատարել միայն որոշ փորձառությունից հետո, եթե կատարում է կենդանին, հատկապես շատ մատաղ հասակում՝ առանց նախնական փորձառության, կամ կատարում են միանման ձևով շատ անհատներ՝ առանց գիտենալու, թե ինչ նպատակի համար է կատարվում այն, սովորաբար անվանում են բնազդ։ Բայց ես կարող էի ցույց տալ, որ այդ սահմանումներից ոչ մեկն էլ ընդհանուր չէ։ Ըմբռնողության կամ դատողության մի փոքրիկ մաս, ինչպես արտահայտվում է Պ. Հուբերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 168] հաճախ նկատվում է բնության սանդուխքի նույնիսկ ցածր աստիճանների վրա կանգնած կենդանիների մոտ։
Ֆրեդերիկ Կյուվյեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 133] և հին մետաֆիզիկներից էլի մի քանիսը բնազդը համեմատել են սովորության հետ։ Ես կարծում եմ, որ այդ համեմատությունը ճիշտ պատկերացում է տալիս բնազդական գործողության մեջ արտահայտվող մտավոր վիճակի բնույթի մասին, բայց ոչ միշտ նրա ծագման մասին։ Սովորական շատ շարժումներ մենք կատարում ենք բոլորովին անգիտակցաբար և հաճախ նույնիսկ ուղղակի հակառակ գիտակցական ցանկության, բայց կարող են փոփոխվել կամքի կամ դատողության մասնակցությամբ։ Որոշ սովորություններ հաճախ զուգորդվում են մյուսների հետ, կապված են լինում մերթ որոշ ժամանակաշրջանի և մերթ օրգանիզմի որոշ դրության հետ։ Երբ ձեռք են բերված, նրանք մնում են անփոփոխ ամբողջ կյանքի ընթացքում։ Կարելի է մատնանշել նմանության նաև այլ գծեր բնազդների և սովորությունների միջև։ Ինչպես լավ ծանոթ եղանակի կրկնության ժամանակ, այնպես էլ բնազդական գործողության ժամանակ մի վարմունքը հետևում է մյուսին որոշ չափով ռիթմիկ կերպով, եթե որևէ մեկին ընդհատում են երգելիս կամ մի անգիր արած բան կրկնելիս, նա սովորաբար ստիպված է լինում ետ դառնալու, որպեսզի ձեռք բերի մտածողության սովորական ընթացքը. նույնն է դիտել Պ. Հուբերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 168] շատ բարդ բոժոժ հյուսող թրթուրի վերաբերմամբ։ Եթե նա վերցրել է բոժոժի կառուցումը, ասենք թե, մինչև վեցերորդ ստադիան հասցրած թրթուրին և տեղափոխել նրան միայն մինչև երրորդ ստադիան հասցրած բոժոժի մեջ, այդ դեպքում թրթուրը պարզապես նորից հյուսել է կառուցման չորրորդ, հինգերորդ և վեցերորդ ստադիաները։ Բայց երբ թրթուրը վերցրել է, օրինակ, երրորդ ստադիայի բոժոժից ու տեղափոխել վեցերորդ ստադիային հասցրած բոժոժի մեջ, այնպես որ նրա աշխատանքի մեծ մասն արդեն կատարված էր, այդ դեպքում նա փոխանակ դրանում որևէ թեթևություն գտնելու, ընկել է շատ մեծ դժվարության մեջ, և աշխատանքը վերջացնելու համար ստիպված է եղել սկսելու երրորդ ստադիայից, որի վրա նրան կանգնեցրել էին, և այդտեղից էլ փորձել է ավարտել գրեթե արդեն վերջացրած աշխատանքը։
Եթե մենք ենթադրենք, որ մի որևէ սովորական գործողություն դառնում է ժառանգական, իսկ ապացուցել կարելի է, որ երբեմն իրոք այդպես էլ լինում է, այդ դեպքում այն նմանությունը, որ սկզբում կար սովորության և բնազդի միջև, հասնում է դժվար տարբերելիության աստիճանին մոտ։ Եթե երեք տարեկան Մոցարտն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 169] սկսեր դաշնամուր նվագել ոչ թե զարմանալի փոքրիկ պրակտիկայից հետո, այլ բոլորովին առանց պրակտիկայի, այդ դեպքում իրավացի կլիներ ասել, որ նա այդ անում է բնազդաբար։ Բայց մեծ սխալ կլիներ կարծել, որ բնազդների մեծ մասը կարող է ծագում առնել մի սերնդի սովորությունից և ժառանգաբար փոխանցվել հաջորդ սերունդներին։ Առանց դժվարության կարելի է ապացուցել, որ այն բնազդներից ամենազարմանալիները, որոնց հետ մենք գործ ունենք, այն է՝ մեղուների և մրջյուններից շատերի բնազդները, չէին կարող ձեռք բերվել այդպիսի ճանապարհով։
Բոլորն ընդունում են, որ բնազդները տեսակի բարեկեցության համար նրա գոյության ներկա պայմաններում նույնքան կարևոր են, որքան որ նրա կառուցվածքի առանձնահատկությունները։ Կենսապայմանների փոփոխության դեպքում հնարավոր է, որ բնազդի մեջ տեղի ունեցող թույլ փոփոխությունները կարող են օգտակար լինել տեսակի համար և եթե կարելի է ապացուցել, որ բնազդները թեկուզ թույլ չափով փոփոխվում են, այդ դեպքում ես դժվարություն չեմ տեսնում ընդունելու այն, որ բնական ընտրությունը պահպանելով ու շարունակ կուտակելով բնազդների մեջ կատարվող շեղումները, կարող է դրանք զարգացնել մինչև ուզած աստիճանի օգտակարության։ Այսպես են ծագել, ինչպես ես կարծում եմ, ամենաբարդ և զարմանալի բնազդները։ Ինչպես որ կառուցվածքում փոփոխությունները առաջանում ու զարգանում են գործածությունից կամ սովորությունից և թուլանում կամ անհետանում են անգործածությունից, ճիշտ այդպես, չեմ կասկածում, եղել է նաև բնազդների հետ։ Բայց ես կարծում եմ, որ սովորության ներգործությունը շատ դեպքերում երկրորդական նշանակություն ունի համեմատած այն փոփոխությունների բնական ընտրության հետ, որոնք կարելի է անվանել բնազդների պատահական փոփոխություններ, այսինքն փոփոխություններ, որ առաջանում են հենց նույն անհայտ պատճառներից, որոնցով պայմանավորված են նաև թույլ շեղումները մարմնի կառուցվածքում։
Բարդ բնազդներից հազիվ թե որևէ մեկը կարող է զարգանալ բնական ազդեցությամբ այլ կերպ, քան թույլ, բայց օգտակար շեղումների դանդաղ ու աստիճանական կուտակման միջոցով։ Հետևաբար, ինչպես որ օրգանական առանձնահատկությունների դեպքում՝ մենք պետք է բնության մեջ գտնենք ոչ թե աստիճանական անցումներ, որոնց միջոցով զարգացել է յուրաքանչյուր բարդ բնազդ,— իսկ այդ կարելի էր հետազոտել յուրաքանչյուր տեսակի միայն ուղղակի նախնիների շարքի մեջ,— այլ պետք է գտնենք որոշ ցուցումներ այդ աստիճանականության նկատմամբ հետնորդների կողմնական գծերում կամ գոնե պետք է ապացուցենք, որ որոշ աստիճանականություն հնարավոր է, և այդ մենք իսկապես կարող ենք ապացուցել։ Ուշադրության առնելով այն, որ Եվրոպայից ու Հյուսիսային Ամերիկայից դուրս կենդանիների բնազդները շատ քիչ են ուսումնասիրված, իսկ մահաջնջված տեսակների բնազդները մեզ բոլորովին անհայտ են, ես զարմացել էի, թե ինչքան շատ կարելի է գտնել դեպի բարդ բնազդները տանող աստիճանական անցումներ։ Բնազդի մեջ փոփոխությունները երբեմն կարող են հեշտանալ նրանով, որ միևնույն տեսակն իր կյանքի զանազան շրջանների ընթացքում, տարվա զանազան եղանակներին, գտնվելով գոյության զանազան պայմաններում և այլն, ունենում է տարբեր բնազդներ, այդպիսի դեպքում բնական ընտրությունը կարող է պահպանել երբեմն այս, երբեմն էլ մի այլ բնազդ։ Եվ միևնույն տեսակի մոտ բնազդի բազմազանության նման դեպքեր, ինչպես կարելի է ապացուցել, բնության մեջ իրոք գոյություն ունեն։
Այնուհետև ճիշտ այնպես, ինչպես որ օրգանիզմի կառուցվածքի դեպքում և իմ տեսության համաձայն՝ յուրաքանչյուր տեսակի բնազդը օգտակար է հենց իրեն, բայց որքան մենք կարող ենք դատել, երբեք չի զարգանում ուրիշների համար նրանից առաջացող օգուտի պատճառով։ Ինձ հայտնի այդպիսի ամենաբնորոշ օրինակներից մեկը, երբ կենդանին կատարում է գործողություն, որն ըստ երևույթին օգտակար է միայն մի ուրիշի համար, ներկայացնում են խոտային լվիճները, որոնք, ինչպես այդ առաջին անգամ նկատել է Հուբերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 168] կամավոր կերպով քաղցր արտաթորանք են տալիս մրջյուններին, որ նրանք այդ անում են կամավոր կերպով, ապացուցվում է հետևյալ փաստերով։ Թրթնջուկի տերևի վրա նստած մոտավորապես մի դյուժին լվիճներից ես հեռացրի մրջյուններին և մի քանի ժամ շարունակ մրջյուններին չէի թողնում նրանց այցելելու։ Ես համոզվեցի, որ այդ ժամանակն անցնելուց հետո լվիճները նեղվում են, որ իրենց քաղցր պարունակությունը չեն արտաթորում։ Որոշ ժամանակ ես նրանց զննում էի խոշորացույցով, բայց նրանցից ոչ մեկը չէր արտաթորում. այնուհետև մազով դիպչում և խուտուտ էի ածում, աշխատելով ըստ հնարավորության անել այդ այնպես, ինչպես անում են մրջյուններն իրենց բողկուկներով, բայց և այնպես ոչ մեկը չարտաթորեց։ Դրանից հետո ես նրանց մոտ թողեցի մի մրջյուն և նրա արագ շարժումներից իսկույն երևաց, որ նա լավ գիտե, թե ինչ հարուստ նախիր է գտել․ այնուհետև նա սկսեց իր բողկուկներով շոյել նախ մեկ, հետո մյուս լվիճի որովայնը և նրանցից յուրաքանչյուրը հենց որ զգում էր բողկուկների հպումը, իսկույն բարձրացնում էր իր որովայնը և արտաթորում հեղուկ քաղցր նյութի կաթիլ, որը ագահաբար լափում էր մրջյունը։ Նույնիսկ բոլորովին մատղաշ լվիճները իրենց պահում էին ճիշտ նույն ձևով, ապացուցելով, որ այդ գործողությունը նրանց մոտ բնազդային է, այլ ոչ թե փորձի արդյունք։ Հուբերի դիտումներից հայտնի է, որ լվիճները անբարյացակամություն չեն տածում դեպի մրջյունները․ բայց եթե վերջիններս չկան, լվիճները, վերջապես, հարկադրված են լինում բաց թողնել իրենց արտաթորանքը առանց մրջյունների։ Քանի որ այդ արտաթորանքը չափազանց կպչուն է, ապա կասկած չկա, որ նրա հեռացումը լվիճների համար ձեռնտու է. իսկ այստեղից էլ հավանական է այն, որ նրանք այն արտաթորում են ոչ բացառապես մրջյունների օգտին։ Այսպիսով, թեև ապացույց չկա, որ մի որևէ կենդանի կատարի մի գործողություն, որը բացառապես օգտակար է մի այլ տեսակի համար, սակայն յուրաքանչյուրը ձգտում է ուրիշների բնազդներից օգուտ քաղել ճիշտ այնպես, ինչպես յուրաքանչյուրն օգտագործում են մի այլ տեսակի կառուցվածքի նույնիսկ մանր թերությունները։ Հետևաբար, որոշ բնազդներ բացարձակորեն կատարյալ չի կարելի համարել, բայց քանի որ այդ և նման մյուս կետերի վերաբերյալ մանրամասնություններն անհրաժեշտ չեն, ուստի այստեղ նրանց վրա կարելի է կանգ չառնել։
Քանի որ բնական վիճակում բնազդների փոփոխելիության որոշ աստիճանը և այդպիսի վարիացիաների ժառանգական փոխանցումը անհրաժեշտ են նրա համար, որպեսզի բնական ընտրությունը կարողանա ներգործել նրանց վրա, ուստի այդ հաստատելու համար պետք էր բերել հնարավորին չափ շատ օրինակներ, բայց տեղի սղությունը ինձ խանգարում է այդ անել։ Ես միայն կարող եմ վավերացնել, որ բնազդներն անշուշտ փոփոխվում են, օրինակ, թափառելու բնազդը կարող է փոփոխվել ինչպես թափառումների չափերի և ուղղությունների, այնպես էլ այդ ընդունակության կորստի տեսակետից։ Ճիշտ նույնն է նաև թռչունների բների հետ, որոնք մասամբ փոխվում են՝ կախված նրանք համար ընտրված տեղից, թռչունների ապրած երկրի բնույթից ու ջերմաստիճանից, մասամբ էլ մեզ բոլորովին անհայտ պատճառներից. Օդյուբոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 117] բերել է Հյուսիսային և Հարավային Միացյալ նահանգներում միևնույն տեսակի բների կառուցվածքում եղած տարբերության մի քանի ուշագրավ դեպքեր։ Սակայն եթե բնազդը կարող է փոփոխվել, այդ դեպքում կարելի է հարցնել, թե ինչո՞ւ նա մեղուներին չի տալիս «որևէ այլ նյութից օգտվելու հմտություն, երբ մեղրամոմ չկա»։ Բայց ուրիշ ի՞նչ բնական նյութ կարող են գործածել մեղուները։ Նրանք կարող են գործադրել, ինչպես ես տեսել եմ, պնդացած մեղրամոմը կինաբարի հետ խառը և փափուկը՝ ճարպի հետ խառը։ Անդրյու Նայտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 31] դիտել է, որ իր մեղուները փոխանակ աշխատասիրութամբ մոմ հավաքելու, գործ են ածել մեղրամոմի և սկիպիդարի խառնուրդ, որով քսված էին պլոկված կեղևով ծառերը։ Վերջերս ապացուցվել է, որ մեղուները փոխանակ ծաղկափոշի հավաքելու, սիրով օգտվում են բոլորովին այլ նյութերից, այն է՝ վարսակի ալյուրից։ Որևէ որոշակի թշնամուց վախենալը, ի հարկե, բնազդական հատկություն է, ինչպես այղ կարելի է տեսնել թռչունների ձագերի վրա, բայց այն պահպանվում է փորձով և նույն թշնամուց այլ կենդանիների վախը դիտելով։ Մարդուց վախենալը ձեռք է բերվում դանդաղորեն, ինչպես ես այդ ցույց եմ տվել մի այլ տեղում անմարդաբնակ կղզիների վրա բնակվող զանազան կենդանիների վերաբերմամբ, և դրա օրինակը մենք տեսնում ենք նույնիսկ Անգլիայում, որտեղ մեր բոլոր խոշոր թռչունները փոքրերի համեմատությամբ ավելի վայրենի են, որովհետև խոշորներին ուժեղ կերպով հալածում են մարդիկ։ Խոշոր թռչունների ավելի մեծ վայրենությունը մենք կարող ենք հանգիստ կերպով վերագրել այդ պատճառին, որովհետև անմարդաբնակ կղզիների վրա խոշոր թռչունները փոքրերից ավելի վախկոտ չեն, և կաչաղակը, որն այնքան զգույշ է Անգլիայում, նույնքան էլ վստահացող է Նորվեգիայում, ինչքան գորշ ագռավը Եգիպտոսում։
Որ միևնույն տեսակին պատկանող վայրի վիճակում ծնված կենդանիների մտավոր հատկությունները շատ ուժեղ կերպով տարբերվում են, կարելի է ապացուցել շատ փաստերով։ Ճիշտ այդպես էլ կարելի է վայրի կենդանիների պատահական ու զարմանալի սովորությունների շատ օրինակներ բերել, որոնք տեսակի համար օգտակար լինելու դեպքում կարող էին բնական ընտրության միջոցով նոր բնազդների սկիզբ տալ։ Բայց ես լավ գիտեմ, որ այդպիսի անապացույց պնդումները, առանց փաստերի մանրամասն շարադրման, քիչ տպավորություն են թողնում ընթերցողի մտքի վրա, և կարող եմ միայն կրկնել իմ հավաստիացումը, որ ես չեմ խոսում՝ չունենալով ծանրակշիռ ապացույցներ։
ՍՈՎՈՐՈՒԹՅԱՆ ԿԱՄ ԲՆԱԶԴԻ ԺԱՌԱՆԳՎԱԾ ՓՈՓՈԽՈՒԹՅՈՒՆՆԵՐՆ ԸՆՏԱՆԻ ԿԵՆԴԱՆԻՆԵՐԻ ՄՈՏ
Բնազդի ժառանգական փոփոխությունների հնարավորությունը կամ նույնիսկ հավանականությունը բնական պայմաններում կարող է հաստատվել ընտանի կենդանիներին վերաբերող մի քանի դեպքերի համառոտ ակնարկով։ Այսպիսով, մենք կկարողանանք ծանոթանալ այն մասնակցությանը, որ ունեցել են սովորությունը և այսպես կոչված պատահական շեղումների ընտրությունը մեր ընտանի կենդանիների մտավոր հատկությունների փոփոխության ժամանակ։ Հայտնի է, թե ինչքան ուժեղ կերպով են տարբերվում ընտանի կենդանիներն իրենց մտավոր հատկություններով։ Այսպես, օրինակ, որոշ կատուներ ծնվելու օրից բռնում են մեծամկներ, մյուսները՝ մկներ, և այդ հակումները ժառանգաբար փոխանցվում են։ Մի կատու, ըստ մ-ր Ս. Ջոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 170] վկայության, միշտ թռչուն էր բերում տուն, մյուսը՝ նապաստակներ կամ ճագարներ, երրորդը որս էր անում ճահճուտ հողամասում և համարյա ամեն գիշեր որսում էր անտառակտցարներ և մորեկտցարներ։ Կարելի է բերել որոշ տրամադրությունների կամ ժամանակաշրջանների հետ կապ ունեցող զանազան ձևի հակումների, ճաշակների և վերին աստիճանի զարմանալի տարօրինակությունների ժառանգական փոխանցման բազմաթիվ հետաքրքրական ու անկասկածելի դեպքեր։ Բայց կանգ առնենք շան ձագերի լավ հայտնի դեպքի վրա. ոչ մի կասկած չկա, որ երիտասարդ պոյնտերները (ես ինքս եմ տեսել այդ ապշեցուցիչ օրինակը) երբեմն կարողանում են կաց անել և նույնիսկ ավելի լավ, քան ուրիշ շներ հենց առաջին անգամից, երբ նրանք դուրս են գալիս որսի, որսորդական շները, անկասկած, որոշ չափով ժառանգում են վարժեցումները․ ոչխարապահ շները փոխանակ ոչխարներին դեմ հանդիման դուրս գալու, ժառանգում են հոտի շուրջը վազելու սովորություն։ Ես չեմ կարող ընդունել, որ այդ գործողությունները գրեթե միանման ձևով, առանց որևէ նախնական փորձի, ընդունակ են կատարելու բոլոր մատղաշ անհատները, ըստ որում յուրաքանչյուր շան ձագ դրանք կատարում է մեծ բավականությամբ և առանց գիտենալու, թե ինչի է հանգում այդ, որովհետև նույնքան քիչ է հասկանում, որ ինքը օգնում է տիրոջը, ինչքան որ կաղամբաթիթեռը գիտե, թե ինչու նա իր ձվերն ածում է կաղամբի տերևի վրա,— ես չեմ կարող ընդունել, որ այդ գործողությունները էապես տարբերվում են իսկական բնազդայիններից։ Եթե մենք տեսնենք, թե ինչպես մի երիտասարդ և անվարժ գայլ, որսի հոտն զգալով, կանգ է առնում մեխվածի պես և հետո դանդաղորեն առանձին քայլվածքով գաղտագողի շարժվում է առաջ, մինչդեռ մի ուրիշը փոխանակ հարձակվելու եղջերուների հոտի վրա, վազում է նրա շուրջը, որպեսզի քշի դեպի մեկուսի մի տեղ, մենք վստահությամբ կարող ենք այդ գործողություններն անվանել բնազդական։ Ընտանի բնազդները, ինչպես կարելի է անվանել նրանց, ի հարկե, անհամեմատ ավելի քիչ կայուն են, քան բնականները, բայց նրանք ենթակա են անհամեմատ ավելի պակաս դաժան ընտրության և փոխանցվել են անհամեմատ ավելի կարճ ժամանակաշրջանի ընթացքում, գոյության նվազ կայուն պայմաններում։
Թե ինչքան ուժեղ է այդ ընտանի բնազդների, սովորությունների և հակումների ժառանգական փոխանցումը և ինչքան հետաքրքրական ձևով խառնվում են նրանք միմյանց հետ, կարելի է տեսնել զանազան ցեղերի շները խաչաձևելիս։ Հայտնի է, որ բուլդոգի հետ խաչաձևումը զարգացնում է համարձակություն և համառություն բարակի շատ սերունդների մեջ. բարակի հետ խաչաձևումը զարգացրել է հովվական շների մի ամբողջ ընտանիքում նապաստակներ որսալու ձգտում։ Այդ ընտանի բնազդները, փոխանցվելով խաչաձևման ժամանակ, նմանվում են բնական բնազդներին, որոնք նույնպես հետաքրքիր ձևով խառնվում են միմյանց հետ և երկար ժամանակի ընթացքում պահպանում են ծնողներից յուրաքանչյուրի բնազդների հետքերը։ Օրինակ, Լերուան[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 171] նկարագրում է մի շուն, որի նախապապը գայլ է եղել և որը պահպանել էր իր վայրենի արյան հետքերը միայն նրանով, որ երբեք ուղիղ գծով չէր գնում տիրոջ մոտ, երբ նա կանչում էր նրան։
Ընտանի բնազդները երբեմն դիտվում են որպես այնպիսի գործողություններ, որոնք ժառանգական են դարձել միայն տևական բռնի կերպով պատվաստված սովորությունների հետևանքով, բայց այդ ճիշտ չէ։ Ոչ ոք չէր մտածի, ասենք, հավանորեն չէր էլ կարողանա սովորեցնել թուրմանին գլխկոնձի տալ օդում, մի շարժում, որը, ինչպես ես կարող եմ հաստատել, կատարում են նաև երիտասարդ թռչունները, որոնք երբեք չեն տեսել գլխկոնձի տվող աղավնի։ Մենք կարող ենք կարծել, որ որևէ աղավնի այդ տարօրինակ սովորության նկատմամբ հակում է ցուցաբերել, և լավագույն անհատների երկարատև ընտրությունը հաջորդական սերունդների շարքում թուրմաններին դարձրել է այնպես, ինչպես մենք նրանց տեսնում ենք այժմ։ Գլազգոյի մոտերքը կան թուրմաններ, որոնք, ինչպես ես լսել եմ մ-ր Բրենտից,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 172] առանց գլխկոնձի տալու 18 դյույմ չեն կարող բարձրանալ։ Կարելի է կասկածել, որ որևէ մեկը մտածեր սովորեցնել շանը կաց անելու, եթե մի որևէ շուն դրա նկատմամբ բնածին հակում չցուցաբերեր, բայց հայտնի է, որ առիթ ներկայանալիս այդ ընդունակությունն ինքն իրեն երևան է գալիս, ինչպես որ ինքս նկատել եմ մի զտարյուն տերյերի վրա։ Կացը, ինչպես կարծում են շատերը, հավանորեն ներկայացնում է որսի վրա հարձակվելու պատրաստվող կենդանու միայն տևական կանգառ։ Եթե մի անգամ երևան է եկել կացի հակումը, ապա, այդ ընդունակության ուժեղ զարգացման հետևանքների սիստեմատիկ ընտրությունը և ժառանգական փոխանցումը հաջորդ սերունդներում կարող էին արագ կերպով ավարտել գործը, անգիտակցական ընտրությունը միշտ մասնակցում է այդ գործին, որովհետև յուրաքանչյուրը ձգտում է, առանց հոգ տանելու ցեղի բարելավման մասին, ձեռք բերել այնպիսի շներ, որոնք դիմացկուն են և ամենալավ ձևով են անում կաց։ Մյուս կողմից, որոշ դեպքերում բավական էր միայն սովորությունը, հազիվ թե մի այլ կենդանու ավելի դժվար լինի վարժեցնել, քան երիտասարդ վայրենի ճագարին, և հազիվ թե մի այլ կենդանի ավելի ձեռնասուն կլինի, քան ընտելացրած երիտասարդ ճագարը. բայց ես դժվարությամբ կարող եմ ենթադրել, որ ընտանի ճագարները երբևիցե ընտրության նյութ են դարձել միայն իրենց հեզության համար և այդ պատճառով գոնե նրանց ժառանգական փոփոխության ավելի մեծ մասը՝ սկսած ծայրահեղ վայրենությունից մինչև ընտելացրած վիճակի ծայրահեղ աստիճանը մենք պետք է վերագրենք սովորությանը և անազատ դրության մեջ երկարատև պահվածքին։
Բնական բնազդները կորչում են ընտանեցման ազդեցության ներքո. դրա մի հիանալի օրինակը մենք կարող ենք տեսնել այն հավերի վրա, որոնք շատ հազվադեպ կամ նույնիսկ երբեք թուխս չեն նստում, այսինքն երբեք չեն ուզում նստել ձվերի վրա։ Շատ երևույթների միայն ամենօրեությունը մեզ խանգարում է նկատելու, թե ինչ աստիճանի և ինչքան խոր կերպով փոփոխվել են մեր ընտանի կենդանիների մտավոր ընդունակությունները։ Հազիվ թե կարելի է կասկածել այն բանում, որ մարդու հետ շների կապվածությունը դարձել է բնազդային։ Բոլոր գայլերը, աղվեսները, չախկալները, և կատվազգիների զանազան տեսակները, ընտելանալով, այնուամենայնիվ սիրում են հարձակվել ընտանի թռչունների, ոչխարների ու խոզերի վրա, և հենց այդ սովորությունը անուղղելի եղավ այն շների մեջ, որոնք մատաղ հասակում բերված են այնպիսի երկրներից, ինչպիսիներն են Հրո երկիրը և Ավստրալիան, որտեղ վայրենիները չեն պահում ընտանի կենդանիներ։ Մյուս կողմից, ինչքան հազվագյուտ դեպքում մեր ընտելացրած շներին նույնիսկ ամենավաղ հասակում պահանջվում է զսպել, որ նրանք չհարձակվեն ընտանի թռչունների, ոչխարների և խոզերի վրա։ Կասկած չկա, որ առիթ ներկայանալիս նրանք այդպիսի հարձակումներ գործում են, բայց այդ ժամանակ նրանց ծեծում են, իսկ եթե այդ չի օգնում, այդ դեպքում նրանց ոչնչացնում են. այսպիսով, սովորությունը և մի որոշ չափի ընտրությունը հավանորեն միասին ժառանգականության օգնությամբ մասնակցել են մեր շների ընտելացմանը։ Մյուս կողմից, հավի ձագերը սովորության ազդեցության տակ բոլորովին կորցրել են շնից ու կատվից վախ զգալը, որն անկասկած սկզբում նրանց մեջ եղել է բնազդային, որովհետև, ինչպես ինձ հաղորդել է կապիտան Հյոտոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 173] Gallus bankiva-ի ճտերը, որոնք Հնդկաստանում են դուրս եկել թխսկան հավի տակ, սկզբում չափազանց վայրենի են լինում։ Նույնն են նաև փասիանի ձագերը, որ դուրս են եկել Անգլիայում թխսկան հավի տակ։ Այդ չի նշանակում, որ ճտերն ընդհանրապես կորցրել են վախի զգացումը, այլ միայն շներից ու կատուներից վախենալու զգացումը, որովհետև երբ մայրը նրանց նախազգուշացնում է վտանգի մասին, նրանք (մանավանդ հնդուհավի ճտերը) մոր տակից փախչում են ու թաքնվում շուրջը բուսնող խոտերի կամ թփուտների մեջ. այդ հավանորեն արվում է բնազդաբար, որպեսզի մորը հնարավորություն տրվի թռչելու, որը մենք տեսնում ենք հավազգի ընտանիքի վայրի թռչունների մեջ։ Բայց այդ բնազդը, որ պահպանում են մեր թռչունների ճտերը, անօգուտ է դարձել ընտանեցման ազդեցության տակ, որովհետև հավն իր թևերը չգործածելու հետևանքով գրեթե իսպառ կորցրել է թռչելու ընդունակությունը։
Այստեղից մենք կարող ենք եզրակացնել, որ ընտանեցման ազդեցության տակ ձեռք են բերվել որոշ բնազդներ, իսկ բնականները կորցվել են, մասամբ սովորության հետևանքով, մասամբ շնորհիվ այն հանգամանքի, որ մարդը մի շարք հաջորդական սերունդներում ընտրել և ուժեղացրել է այնպիսի յուրահատուկ մտավոր հակումներ ու շարժումներ, որոնք զարգանում են այն պատճառներից, որոնց մենք մեր անտեղյակության պատճառով անվանում ենք պատահական։ Որոշ դեպքերում միայն առաջացրած սովորությունը բավական էր նրա համար, որպեսզի կատարվեն մտավոր ընդունակությունների ժառանգական փոփոխություններ, այլ դեպքերում առաջացրած սովորությունը նշանակություն չի ունեցել և բոլորը եղել է ինչպես սիստեմատիկ, այնպես էլ անգիտակցական ընտրության արդյունք, բայց շատ դեպքերում սովորությունը և ընտրությունը հավանորեն ընթացել են ձեռք ձեռքի տված։
ԱՌԱՆՁՆԱՀԱՏՈՒԿ ԲՆԱԶԴՆԵՐ
Ծանոթանալով մի քանի օրինակների, գուցե մենք ավելի լավ կըմբռնենք բնազդների փոփոխությունը բնական վիճակում ընտրության ազդեցության տակ։ Ես կընտրեմ միայն երեքը, այն է՝ այն բնազդը, որ կկուին ստիպում է ուրիշների բներում ձու ածել, որոշ մրջյունների ստրկատիրական բնազդը և մեղուների շինարարական ընդունակությունները։ Երկու վերջին բնազդները սովորաբար բնախույզները միանգամայն արդարացիորեն համարում են մեզ հայտնի բնազդներից ամենազարմանալիները։
Կկուի բնազդները։ Մի քանի բնախույզներ կարծում են, թե կկուի բնազդի անմիջական պատճառն այն է, որ նա ամեն օր ձու չի ածում, այլ միշտ երկու կամ երեքօրյա ընդմիջումներով. ուստի՝ եթե նա շիներ իր սեփական բունը և ինքը թուխս նստեր, այդ դեպքում կամ առաջին ածած ձվերը մի որոշ ժամանակ կմնային չթխսված, կամ միևնույն բնում կլինեին թե ձվեր և թե զանազան հասակի ձագեր։ Այդ դեպքում ձու ածելու և թխսելու ժամանակաշրջանը անհարիր կերպով մեծ կլիներ, մանավանդ որ կկուն շատ վաղ է չվում, և առաջին դուրս եկած ձագին հավանորեն միայն արուն պիտի կերակրեր։ Սակայն ամերիկական կկուն գտնվում է հենց այդ դրության մեջ, որովհետև նա իր սեփական բունն է շինում և ունենում է միևնույն ժամանակ թե ձվեր և թե ձվից հաջորդականորեն դուրս եկած ձագեր։ Ոմանք պնդել են, մյուսները ժխտել են, որ ամերիկական կկուն ուրիշների բներում պատահաբար է ածում ձվեր, բայց ես վերջերս լսել եմ այովացի դ-ր Մերրելից,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 174] որ նա մի անգամ Իլլինոիսում գտել է կապույտ ճայի (Garrulus cristatus) բնում մատղաշ ճայի հետ միասին մատղաշ կկու, և որովհետև երկու թռչնակներն էլ համարյա փետրավորված էին, ուստի այս դեպքում թռչուններին որոշելու մեջ սխալ չէր կարող լինել։ Ես կարող էի այդպիսի թռչունների մի քանի օրինակներ ևս բերել, որոնք պատահականորեն ածում են իրենց ձվերն ուրիշների բներում։ Այժմ ենթադրենք, թե մեր եվրոպական կկուի հեռավոր նախահայրը ունեցել է ամերիկական կկուի սովորությունը և պատահականորեն ձու է ածել ուրիշի բնում։ Եթե այդ պատահական հանգամանքի շնորհիվ ծեր թռչունը օգուտ է ձեռք բերել նրանով, որ կարողացել է ավելի վաղ չվել կամ օգտվել է մի այլ ձևով, կամ եթե մատղաշ թռչնակը դարձել է ավելի ամրակազմ, մի այլ թռչնատեսակի խաբված բնազդի հետևանքով գտնվելով ավելի բարենպաստ պայմաններում, քան եթե սնուցվեր իր մոր կողմից, որը չափազանց ծանրաբեռնված է միաժամանակյա հոգսերով ձվերի և զանազան հասակի ձագերի մասին, այդ դեպքում թե ծեր թռչունները և թե սնուցված ձագերը կգտնվեին բարենպաստ դրության մեջ։ Իսկ անալոգիան մեգ հարկադրում է ընդունել, որ այդ ձևով սնուցված ձագերը ժառանգականության միջոցով կարողացել են յուրացնել իրենց մոր պատահական ու շեղվող սովորությունը, կարողացել են իրենց հերթին ձվեր ածել ուրիշների բներում և դրա շնորհիվ հաջողությամբ մեծացնել ձագեր։ Այդպիսի երկարատև պրոցեսի միջոցով, ես կարծում եմ, մեր կկուի յուրահատուկ բնազդը կարող էր դառնալ բնածին։ Վերջերս Ադոլֆ Մյուլլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 175] նույնպես բավարար ապացույց է բերել, որ կկուն երբեմն ձվերն ածում է լերկ գետնի վրա, թուխս է նստում և սնուցում է ձագերին։ Այդպիսի հազվագյուտ դեպքը հավանորեն ներկայացնում է վերադարձ դեպի բնակալության վաղուց կորցրած բնազդը։
Ինձ դիտողություն են արել, որ ես ուշադրություն չեմ դարձրել կկուի մի քանի բնազդների և նրա կառուցվածքներում եղած հարմարումների վրա, որոնք, ինչպես ասում են, անպայման գտնվում են որոշ հարաբերակցության մեջ։ Սակայն տեսական կշռադատությունները մի բնազդի առթիվ, որը մեզ հայտնի է բացառապես մի տեսակի մոտ, համենայն դեպս անօգուտ են, որովհետև մենք դրա համար ղեկավարող տվյալներ չենք գտնում։ Մինչև վերջին ժամանակները հայտնի էին միայն եվրոպական և ամերիկական ոչ պարազիտային կկուի բնազդները. այժմ, ըստ մ-ր Ռեմզեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 176] դիտումների, մենք տեղեկություններ ունենք ավստրալիական երեք տեսակների մասին, որոնք ձու են ածում այլ թռչունների բներում։ Պետք է ուշադրություն դարձնել երեք հանգամանքի վրա։ Առաջինն այն է, որ սովորական կկուն, հազվագյուտ բացառությամբ, մի բնում մեկական ձու է ածում, որի հետևանքով նրա խոշոր և շատակեր ձագը առատ կեր է ստանում։ Երկրորդը՝ որ նրա ձվերը զարմանալի կերպով փոքր են, դաշտային արտուտի ձվերից ոչ ավելի մեծ, մի թռչունի, որը չորս անգամ ավելի փոքր է կկուից։ Որ ձվերի փոքրությունը իրոք ներկայացնում է հարմարեցման երևույթ, մենք կարող ենք տեսնել նրանից, որ ամերիկական ոչ պարազիտային կկուն ածում է խոշոր ձվեր։ Երրորդը, որ մատղաշ կկուն ձվից դուրս գալուց հետո շատ չանցած ունենում է բնազդ, ուժ և յուրահատուկ կառուցված մեջք, որպեսզի դուրս նետի իր խորթ եղբայրներին, որոնք հետո ոչնչանում են սովից ու ցրտից։ Եվ այդ անվերապահորեն կոչվում է բարերար հարմարում, որի հետևանքով մատղաշ կկուն ստանում է բավարար կեր, իսկ նրա խորթ եղբայրները դեռ զգայունակություն ձեռք չբերած ոչնչանում են։
Այժմ դառնանք ավստրալիական տեսակներին, թեև այս թռչունները սովորաբար մեկական ձու են ածում բնում, բայց հազվադեպ չէ միևնույն բնում երկու և նույնիսկ երեք ձու գտնելը։ Բրոնզագույն կկուի ձվերն իրենց մեծությամբ զգալիորեն տատանվում են, ունենալով ութից մինչև տասը գծաչափ երկարություն։ Եթե տեսակի համար ձեռնտու լիներ ավելի ևս փոքր չափերի ձվեր ածել, քան նա ածում է այժմ նրա համար, որպեսզի ձագերն ավելի շուտ դուրս գան ձվից (որովհետև ապացուցված է, որ ձվերի մեծության և նրանց թխսելու տևողության միջև գոյություն ունի հարաբերակցություն), այդ դեպքում դժվարություն չկա ընդունելու այնպիսի ցեղի կամ տեսակի առաջացման հնարավորությունը, որը կածեր ավելի ու ավելի մանր ձվեր, որովհետև ավելի հուսալի կլիներ նրանց վրա թուխս նստել և խնամել։ Մ-ր Ռեմզեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 176] նկատում է, որ ավստրալիական երկու կկուներ, ձվերը բաց բնում ածելիս, բացահայտ գերադասություն են տալիս իրենց սեփական ձվերին գույնով նմանվող ձվեր ունեցող բներին։ Եվրոպական տեսակը նկատելի կերպով հակում է ցուցաբերում դեպի այդպիսի բնազդը, բայց հաճախ նաև շեղվում է դրանից, ինչպես այդ երևում է նրանից, որ նա իր բաց գույնի, դժգույն ձվերն ածում է կանաչակապույտ գույնով ձվեր ունեցող շիկահավի բներում։ Եթե մեր կկուն անպայման դրսևորեր մատնանշված բնազդը, ապա այդ բնազդը անշուշտ կդասվեր նրանց շարքը, որոնց վերաբերմամբ ընդունվում է, որ նրանք բոլորը միաժամանակ են ձեռք բերվել։ Ավստրալիական բրոնզագույն կկուի ձվերը, ըստ մ-ր Ռեմզեյի խոսքերի, գույնով արտասովոր կերպով փոփոխական են. այդ պատճառով բնական ընտրությունը կարող էր զարգացնել և ամրացնել նրանց մեջ ամեն մի բարենպաստ փոփոխություն ինչպես այդ ուղղությամբ, այնպես էլ մեծության տեսակետից։
Եվրոպական կկուի ձագերն իրենց ընդունող ծնողների ձագերին բնից դուրս են նետում ձվից դուրս գալուց հետո երեք օրվա ընթացքում, իսկ քանի որ կկուի ձագն այդ հասակում խիստ անօգնական է, ուստի մ-ր Գուլդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 89] սկզբում հակվել է կարծելու, որ ձագերին դուրս նետելու գործողությունը կատարում են հենց իրենց ընդունող ծնողները։ Բայց այժմ նա վստահելի վկայություն է ստացել, որ իրոք տեսել են, թե ինչպես մատղաշ կկուն, որը թեև կույր է և գլուխը բարձրացնելու ընդունակ չէ, ինքն է դուրս նետում իր խորթ եղբայրներին։ Դիտողը նրանցից մեկին նորից է դրել բնի մեջ, և նրան այնտեղից նորից դուրս են նետել։ Ինչ վերաբերում է այն բանին, թե ինչպես կարող էր զարգանալ այդպիսի տարօրինակ ու զզվելի մի բնազդ, ապա այն դեպքում, եթե մատղաշ կկուի համար շատ կարևոր էր,— ինչպես որ հավանորեն կա իրականում,— ծնվելուց հետո շուտով ստանալ հնարավորին չափ ավելի շատ սնունդ, ես առանձին դժվարություն չեմ տեսնում այն բանում, որ կկուն աստիճանաբար մի շարք հաջորդական սերունդներում ձեռք է բերել կույր ցանկություն, ուժ և կառուցվածք, որոնք անհրաժեշտ են ձագերին դուրս նետելու գործողությունը կատարելու համար, որովհետև այդպիսի սովորությունների և անհրաժեշտ կառուցվածքի ամենալավ զարգացում ունեցող մատղաշ կկուները կարող էին ամենից ավելի հեշտությամբ մեծանալ։ Այդ բնածին բնազդը ձեռք բերելու համար առաջին քայլը կարող էր լինել մատղաշ թռչունների մի մասի բոլորովին չնախամտածված անհանգստությունը, երբ նրանք մեծացել և ավելի ամրակազմ էին դարձել, իսկ հետո այդ սովորությունը կարող էր զարգանալ և անցնել ավելի վաղ հասակին։ Դա ինձ համար ավելի մեծ դժվարություն չի ներկայացնում, քան այն, որ ձվից դեռ դուրս չեկած ճուտը ձեռք է բերել կեղևը ծակելու բնազդ, կամ այն, որ օձի ձագը իր վերին ծնոտի վրա, ըստ Օուենի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] դիտողության, ունի ժամանակավոր սուր ատամ ձվի ամուր կեղևը պատռելու համար։ Ուստի՝ եթե օրգանիզմի յուրաքանչյուր մասը ենթակա է անհատական տատանումների ամեն մի հասակում և վարիացիաները հակում ունեն համապատասխան կամ ավելի վաղ հասակում ժառանգաբար փոխանցվելու,— սրանք դրույթներ են, որոնք չի կարելի ժխտել,— այդ դեպքում երիտասարդ անհատի բնազդները և կառուցվածքը կարող են աստիճանաբար փոփոխվել, ինչպես չափահաս անհատինը, այս երկու բացատրությունները պետք է կամ պահպանվեն կամ ընկնեն բնական ընտրության ամբողջ տեսության հետ միասին։
Ամերիկական թռչունների մի շատ յուրահատուկ սեռի Molothrus-ի մի քանի տեսակները, որոնք մոտ են մեր սարյակներին, կկուի նման ունեն պարազիտային սովորություններ և իրենց բնազդների կատարելության աստիճանի մեջ ներկայացնում են հետաքրքրական հաջորդականություն։ Molothrus badius-ի երկու սեռն էլ, ինչպես այդ հաստատում է գերազանց դիտող մ-ր Հուդզոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 115] երբեմն ապրում են խառը երամներով, երբեմն էլ զույգերով։ Երբեմն նրանք սեփական բուն են շինում, երբեմն էր գրավում են որևէ այլ թռչունի պատկանող բուն և այդ դեպքում դուրս են նետում օտար ձագերին։ Որոշ դեպքերում նրանք ձու են ածում իրենց գրաված բնում, այլ դեպքերում դրա համար բնի գագաթին սարքում են մի բավական տարօրինակ կառուցում։ Սովորաբար այդ թռչուններն իրենց ձվերի վրա իրենք են թուխս նստում և իրենք էլ սնուցում են իրենց ձագերին, բայց մ-ր Հուդզոնն ասում է, որ ամենայն հավանականությամբ, նրանք երբեմն են պարազիտություն անում, որովհետև նա տեսել է այդ տեսակի երիտասարդ թռչուններ, որոնք գնում են այլ տեսակների չափահաս թռչունների հետևից, ըստ որում նրանք ճիչերով պահանջում էին իրենց համար կեր։ Molothrus-ի մի այլ տեսակի՝ M. bonariensis-ի մոտ պարազիտային սովորությունները անհամեմատ ավելի մեծ չափով են զարգացած, քան առաջինի մոտ, բայց դեռ ամենևին կատարելագործված չեն։ Որքան ինձ հայտնի է, այդ թռչունը միշտ ուրիշների բներում է ձու ածում, բայց ուշագրավ է այն, որ երբեմն մի քանի թռչուններ միասին սկսում են շինել կոպիտ անկանոն մի բուն, տեղավորելով այն զարմանալի մի անհաջող տեղում, օրինակ մեծ տատասկափշի վրա։ Սակայն, որքան մ-ր Հուդզոնը կարողացել է համոզվել, նրանք երբեք չեն ավարտում իրենց այդպիսի բունը։ Հաճախ նրանք օտար բնում այնքան շատ (15-ից մինչև 20) ձու են ածում, որ այդ ձվերից հավանորեն միայն շատ քչերի կամ նույնիսկ ոչ մեկի վրա թուխս չեն նստում։ Բացի դրանից, նրանք սովորություն ունեն գրաված բնի ձվերը կտցահարելով ծակել անկախ այն բանից, թե նրանք պատկանում են իրենց սեփական տեսակին կամ ընդունող ծնողներին։ Վերջապես, նրանք գետին են նետում շատ ձվեր, որոնք այնտեղ էլ ոչնչանում են։ Երրորդ տեսակը՝ հյուսիս-ամերիկական M. pecoris-ը ձեռք է բերել նույնքան կատարելագործված բնազդներ, ինչքան կկուն, և օտար բնում մեկ ձվից ավելի երբեք չի ածում, որի հետևանքով մատղաշ թռչունի սնուցումը բոլորովին ապահովված է լինում։ Հուդզոնը էվոլյուցիայի վճռական հակառակորդ է, բայց թվում է, թե նա այնքան զարմացել է Molothus bonariensis-ի անկատար բնազդներից, որ բերում է իմ խոսքերը, հարցնելով. «Արդյոք չպե՞տք է մենք այդ սովորությունները համարենք ոչ թե որպես հատկապես ի վերուստ առաքված կամ ստեղծված բնազդներ, այլ որպես փոքրիկ հետևանքներ, որոնք բխում են մի ընդհանուր օրենքից, այն է՝ աստիճանական անցման օրենքից»։
Ինչպես արդեն նկատված է, զանազան թռչուններ երբեմն ձվեր են ածում այլ թռչունների բներում։ Այդ սովորությունը բավական սովորական է հավազգիների մեջ և որոշ լույս է սփռում ջայլամի յուրահատուկ բնազդի վրա։ Այդ ընտանիքում մի քանի էգեր, միանալով միասին, սկզբում քիչ ձու են ածում մի բնում, հետո՝ մյուսում, իսկ ձվերի վրա նստում են արուները։ Այդ բնազդը հավանորեն առաջացել է նրանից, որ էգերը շատ ձու են ածել, բայց կկուի նման երկու կամ երեք օրվա ընդմիջումներով։ Սակայն ամերիկական ջայլամի մոտ, ինչպես և Molothrus bonariensis-ի մոտ, այդ բնազդը կատարելության դեռ չի հասել, և այդ պատճառով այդ թռչունի ձվերը զարմանալի մեծ քանակությամբ ցրված են հարթությունների վրա, այնպես որ ես մի օրվա ընթացքում հավաքելիս գտել եմ քսանից ոչ պակաս կորցված ու փչացած ձվեր։
Շատ մեղուներ պարազիտություն անողներ են, շարունակ իրենց ձվերը դնում են այլ մեղուների բներում։ Այս դեպքն ավելի ևս ուշագրավ է, քան կկուի պարազիտությունը, որովհետև այդ մեղուների մոտ, նրանց պարազիտային սովորություններից կախված, փոփոխվել է ոչ միայն բնազդը, այլև կառուցվածքը, նրանք չունեն ծաղկափոշի հավաքելու ապարատ, մինչդեռ այն անհրաժեշտ կլիներ, եթե նրանք իրենք կերի պաշար հավաքեին իրենց սերնդի համար։ Sphegidae-ի (պիծակների նման միջատներ) մի քանի տեսակներ նմանապես պարազիտություն են անում, և մ-ր Ֆաբրը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 78] վերջերս բերել է լավ ապացույց հաստատելու համար այն, որ թեև Tachites nigra-ն սովորաբար իր սեփական բներն է շինում, դրանք օժտելով կաթվածահար արված որսով իր թրթուրների համար, սակայն եթե գտնում է մի ուրիշ սֆեքսի համարյա պատրաստի, պաշարներ ունեցող բնիկ, այդ դեպքում օգտվում է ավարից և դառնում է պարազիտ։ Այս դեպքում էլ, ինչպես որ Molothrus-ի կամ կկուի վերաբերմամբ, ես դժվարություն չեմ տեսնում, որպեսզի բնական ընտրությունը պատահական սովորությունը դարձներ մշտական, եթե նա օգտակար է տեսակի համար և եթե այն մի շատը, որի բունը և կերի պաշարը այդպիսի դավաճանական ձևով բռնագրավված են, չի մահաջնջվի դրանից։
Ստրկատիրական բնազդ։ Այս ուշագրավ բնազդն առաջին անգամ հայտնագործել է Formica (Polyerges) rufescens-ի մոտ Պյոտր Հուբերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 168] որը էլ ավելի լավ դիտող էր, քան նրա հռչակավոր հայրը։ Մրջյունը, որի մասին խոսվում է, լիակատար կախման մեջ է գտնվում իր ստրուկներից. առանց նրանց օգնության այս տեսակը հավանորեն կմահաջնջվեր մի տարվա ընթացքում։ Արուները և պտղաբերող էգերը բոլորովին չեն աշխատում, իսկ աշխատավորները կամ ամուլ էգերը, լինելով շատ եռանդուն և արի ստրուկներ բռնելիս, ուրիշ ոչինչ չեն անում։ Նրանք անընդունակ են իրենց համար մրջնանոց կառուցելու կամ կերակրելու իրենց թրթուրներին։ Երբ գտնում են, որ հին բունը անբավարար է, և նրանք հարկադրված են տեղափոխվելու, ապա այդ տեղափոխությունը կատարում են ստրուկները, որոնք իրենց տերերին տանում են ծնոտների մեջ բռնած։ Տերերն այնքան անօգնական են, որ երբ Հուբերը նրանցից մոտավորապես 30 հատ փակել է առանց ստրուկների, բայց առատությամբ տալով սիրած կերը և նրանց մոտ դնելով թրթուրներ և բոժոժներ, որպեսզի նրանք ստիպվեն գործելու, նրանք չեն արել, չեն կարողացել նույնիսկ կերակրվել և նրանցից շատերը սովամահ են եղել։ Այն ժամանակ Հուբերը նրանց մոտ թողել է մի ստրուկ (F. fusca), վերջինս իսկույն անցել է գործի, կերակրել ու փրկել ողջ մնացածներին, պատրաստել է մի քանի բջիջներ, հոգ է տարել թրթուրների մասին և ամեն ինչ կարգի է բերել։ Ի՞նչը կարող է այդ ստույգ փաստերից ավելի զարմանալի լինել։ Եթե մենք չգիտենայինք ոչ մի ուրիշ ստրկատեր մրջյուն, ապա բոլորովին անհուսալի կլիներ քննարկել, թե ինչպես կարող էր զարգանալ մի այդքան զարմանալի բնազդ։
Մի այլ տեսակ՝ Formica sanguinea-ն որպես ստրկատեր մրջյուն առաջին անգամ հայտնի է դարձել նույնպես շնորհիվ Պ. Հուբերի։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 168] Այս տեսակը հանդիպում է Անգլիայի հարավային մասերում, և նրա կենսակերպը ուսումնասիրել է Բրիտանական թանգարանի կողմից մ-ր Ֆ. Սմիտը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 177] որին ես շատ պարտական եմ ինչպես այդ, այնպես էլ այլ փաստերի մասին ինձ հաղորդածի համար։ Լիովին վստահելով Հուբերի և Սմիտի վկայություններին, ես այնուամենայնիվ թերահավատությամբ վերաբերվեցի տվյալ դեպքին, որովհետև հասկանալի է, որ յուրաքանչյուրին ներելի են նրա կասկածները մի այդպիսի անսովոր բնազդի գոյության մասին, ինչպիսին է ստրկատիրական բնազդը։ Այդ պատճառով ես այստեղ կհաղորդեմ իմ կատարած դիտումները որոշ մանրամասնություններով։ Ես բաց եմ արել Formica sanguinea-ի տասնևչորս բուն և բոլորի մեջ գտել եմ մի քանի ստրուկներ։ Ստրկացրած տեսակի (F. fusca) արուներն ու պտղաբերող էգերը գտնվել են միայն իրենց սեփական բներում և երբեք չեն նկատվել F. Sanguinea-ի բներում։ Ստրուկները սև են և երկու անգամ ավելի մանր են իրենց կարմրագույն տերերից, այնպես որ արտաքին տեսքով մեկի և մյուսի տարբերությունը շատ մեծ է։ Եթե մրջնանոցն անհանգստացված է թույլ լափով, ապա որոշ դեպքերում ստրուկները դուրս են վազում և նույնպես, ինչպես և տերերը, խիստ գրգռված են լինում, պաշտպանելով բունը, իսկ եթե մրջնանոցը շատ խիստ է վնասված, այնպես որ թրթուրներն ու բոժոժները դուրս են մնում, այդ դեպքում ստրուկները տերերի հետ միասին եռանդուն կերպով աշխատում են, որպեսզի նրանց տեղափոխեն անվտանգ տեղ։ Այստեղից պարզ է, որ ստրուկներն իրենց զգում են ինչպես բոլորովին սեփական տանը։ Երեք տարի անընդհատ հունիսի և հուլիսի ընթացքում ես ժամերով դիտել եմ զանազան բներ Սուրրեյում և Սուսսեքսում և երբեք ստրուկներին չեմ տեսել ել ու մուտ անելիս։ Որովհետև այդ ամիսներին ստրուկները շատ սակավաթիվ են, ուստի ես կարծում էի, որ նրանք կարող են այլ կերպ վարվել, երբ ավելի բազմաթիվ են լինում, բայց մ-ր Սմիտը ինձ հաղորդում է, որ մայիսի, հունիսի և օգոստոսի ընթացքում, ինչպես Սուրրեյում, այնպես էլ Հեմպշիրում նա դիտել է բները օրվա տարբեր ժամերին և երբեք չի տեսել, որ ստրուկները մտնեին բները կամ նրանցից դուրս գային, թեև օգոստոսին նրանք խիստ բազմաթիվ են։ Այստեղից էլ նա նրանց համարում է բացառապես տնային ստրուկներ։ Իսկ տերերին, ընդհակառակը, շարունակ կարելի է տեսնել, երբ նրանք բնի համար բերում են նյութեր կամ բազմապիսի կեր։ Սակայն 1860 թվականի հուլիսին հանդիպել եմ անսովոր կերպով մեծ քանակությամբ ստրուկներ ունեցող մի համայնքի և դիտել եմ, թե ինչպես նրանցից մի քանիսը տերերի հետ միասին թողեցին բունը և գնացին միասին մի ճանապարհի երկարությամբ դեպի 25 յարդ հեռավորության վրա կանգնած շոտլանդական սոճին, որի վրա նրանք բարձրանում էին հավանորեն լվիճներ ու զատիկներ որոնելու համար։ Ըստ Հուբերի, որը հնարավորություն է ունեցել երկարատև դիտումներ կատարելու, ստրուկները Շվեյցարիայում սովորաբար տերերի հետ միասին են աշխատում բունը շինելու ժամանակ, բայց առավոտյան ու երեկոյան միայն նրանք են բաց անում և փակում մուտքի անցքերը, նրանց գլխավոր պարտականությունը, ինչպես հաստատում է Հուբերը, լվիճներ որոնելն է։ Տերերի և ստրուկների կենսակերպի միջև եղած տարբերությունը երկու երկրներում հավանորեն կախված նրանից, որ Շվեյցարիայում ավելի մեծ քանակությամբ ստրուկներ են որսվում, քան Անգլիայում։
Մի անգամ ինձ հաջողվեց դիտել F. sanguinea-ի տեղափոխությունը մի մրջնանոցից մյուսը, ընդ որում չափազանց հետաքրքրական էր տեսնել, թե ինչպես տերերը հոգատարությամբ տեղափոխում էին ստրուկներին իրենց ծնոտներում բռնած՝ փոխանակ իրենք փոխադրված լինելու ստրուկների կողմից, ինչպես որ F. rufescens-ի մոտ է։ Մի ուրիշ անգամ իմ ուշադրությունը գրավեցին երկու տասնյակ ստրկատերեր, որոնք միևնույն տեղում վազվզում էին ակնհայտորեն ոչ կեր որոնելու համար, նրանք հարձակվում էին, իսկ նրանց վռնդում էին ստրկացրած տեսակի (F. fusca) անկախ գաղութից։ Այդ ժամանակ երբեմն F. fusca-ի երեք անհատներ միանգամից կպչում էին F. sanguinea-ի մի ստրկատիրոջ ոտքերին, իսկ վերջինս անողոք կերպով սպանում էր իր փոքրիկ հակառակորդներին և նրանց դիակները որպես կեր տանում էր իր բունը, որ գտնվում էր քսանևինը յարդ հեռավորության վրա. բայց նրանց թույլ չտվեցին տիրանալու ոչ մի հարսնյակի ստրուկներ դարձնելու համար։ Այն ժամանակ ես վերցրեցի F. fusca-ի մի ուրիշ մրջնանոցից փոքր քանակությամբ հարսնյակներ և դրեցի մարտի տեղին մոտ մի հարթակի վրա. բռնակալները ագահաբար վրա պրծան և հափշտակեցին նրանց, հավանորեն երևակայելով, որ իրենք այսպես թե այնպես հաղթեցին վերջին մարտում։
Միևնույն ժամանակ ես նույն տեղում դրեցի մի այլ տեսակի՝ F. flava-ի մի քանի հարսնյակներ այդ փոքրիկ դեղնագույն մրջյունի մի քանի անհատների հետ միասին, որոնք դեռ կպած էին իրենց բնից պոկած մասնիկներին։ Այս տեսակը, թեև հազվադեպ, երբեմն նույնպես ենթարկվում է ստրկացման, ինչպես այդ նկարագրել է Սմիտը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 177] Չնայած իր չնչին մեծությանը, դեղին մրջյուններն աչքի են ընկնում արիությամբ, և ես տեսել եմ, որ նրանք կատաղի կերպով հարձակվել են այլ մրջյունների վրա։ Մի անգամ, ի զարմանս իմ, ես գտա F. flava-ի մի անկախ գաղութ ստրկատիրական F. sanguinea տեսակի բնից ցած, քարի տակ, և երբ ես պատահաբար վնասեցի երկու բները, փոքրիկ մրջյունները զարմանալի արիությամբ հարձակվեցին իրենց խոշոր հարևանների վրա։ Ինձ հետաքրքիր է համոզվել, թե արդյոք F. sanguinea-ի տարբերում է F. fusca-ի հարսնյակները, որոնց սովորաբար նրանք ստրկացնում են, փոքրիկ ու կատաղի F. flava-ի հարսնյակներից, որոնց նրանք միայն հազվադեպ են ստրկացնում, և ակնհայտ եղավ, որ նրանք իսկույն տարբերեցին նրանց։ Մենք տեսանք, որ նրանք եռանդուն ու արագ կերպով հափշտակեցին F. fusca-ի հարսնյակներին, բայց շատ վախեցան, երբ հանդիպեցին F. flava-ի հարսնյակներին կամ բնից պոկված հողակտորներին և փութով փախչում էին հեռու, սակայն մոտավորապես քառորդ ժամից հետո քիչ ժամանակ անց, երբ բոլոր դեղին մրջյունները հեռացել էին, նրանք արիությամբ զինվեցին ու հարսնյակներին տիրացան։
Մի անգամ երեկոյան ես այցելեցի F. sanguinea-ի մի այլ համայնք և գտա, որ մեծ քանակությամբ այդ մրջյունները տուն են վերադառնում և մտնում բունը, իրենց հետ բերելով F. fusca-ի դիակներ (դա ցույց էր տալիս, որ իմ դիտածս տեղափոխություն չէր) և բազմաթիվ հարսնյակներ։ Ես հետևեցի ավարով բեռնավորված այդ մրջյունների երկար շղթային համարյա 40 յարդի վրա՝ մինչև հավամրգենու խիտ թփուտը, որտեղից դուրս եկավ F. sanguinea-ի վերջին մրջյունը, որը մի հարսնյակ էր տանում, բայց ինձ չհաջողվեց հավամրգենու խիտ թփուտում գտնել ավերված բունը, թեև նա պետք է բոլորովին մոտ լիներ, որովհետև F. fusca-ի երկու կամ երեք անհատներ մեծ հուզմունքով վազվզում էին շուրջը, և նրանցից մեկը այդ տեսակի մի հարսնյակ բերանում անշարժ կերպով տեղավորվել էր հավամրգենու ճյուղի գագաթին, իրենով մարմնավորվելով այն հուսահատությունը, որ առաջացրել է նրանց տան ավերումը։
Այդպիսիք են ստրկատիրական զարմանալի բնազդին վերաբերող փաստերը, որոնք, ասենք, իմ կողմից հաստատման կարիք չունեն։ Ուշադրություն դարձրեք այն բանի վրա, թե ինչպիսի հակադրություն են ներկայացնում F. sanguinea-ի բնազդական սովորությունները մայր ցամաքի F. rufescens-ի սովորությունների համեմատությամբ։ Վերջինս ինքը բուն չի շինում, չի տեղափոխվում, կեր չի հավաքում ոչ իրեն համար, ոչ էլ մատղաշների համար և չի կարող ինքը կերակրվել, այլ կախված է իր բազմաթիվ ստրուկներից։ Մյուս կողմից, Formica sanguinea-ն ունի անհամեմատ ավելի քիչ ստրուկներ, իսկ ամառվա սկզբին նույնիսկ չափազանց քիչ. տերերն իրենք են որոշում, թե երբ և որտեղ պետք է կառուցվի նոր բնակարանը, և տեղափոխվելու ժամանակ տանում են իրենց ստրուկներին։ Ինչպես Շվեյցարիայում, այնպես էլ Անգլիայում բացառապես ստրուկների վրա է ընկնում, ինչպես ինձ թվում է, թրթուրների հոգսը, իսկ տերերը միայն արշավանքներ են կատարում ստրուկներ բռնելու համար։ Շվեյցարիայում ստրուկներն ու տերերը միասին են աշխատում, հավաքելով ու բերելով նյութեր բնի համար, թե՛ ստրուկները և թե՛ տերերը, բայց գերադասորեն առաջինները, պահպանում ու կթում են, եթե կարելի է այդպես ասել, լվիճներին. նրանք երկուսն էլ հավաքում են կեր համայնքի համար։ Անգլիայում սովորաբար միայն տերերն են դուրս գալիս բնից շինարարական նյութ և ինչպես իրենց կերը, այնպես էլ ստրուկների ու թրթուրների կերը հավաքելու համար։ Այսպիսով, Անգլիայում տերերն իրենց ստրուկներից ավելի քիչ են սպասարկություն ստանում, քան Շվեյցարիայում։
Ես կռահումներով չեմ տարվի, թե ինչպես է զարգացել F. sanguinea-ի բնազդը։ Բայց քանի որ ստրկատիրական բնազդ չունեցող մրջյունները, ինչպես ես նկատել եմ, հափշտակում են այլ տեսակի հարսնյակներին, եթե վերջիններս թափված են իրենց բների մոտ, ապա հնարավոր է, որ սկզբում կերի համար հափշտակված այդպիսի հարսնյակները կարող էին զարգանալ ու դառնալ չափահաս միջատներ, իսկ այդ ձևով առանց դիտավորության սնուցված օտար մրջյունները հետևեին իրենց սեփական բնազդներին և անեին այն, ինչ կարող էին։ Եթե նրանց ներկայությունը օգտակար էր հափշտակող տեսակի համար, եթե նրա համար ավելի ձեռնտու էր աշխատավորներ հափշտակելը, քան նրանց սնուցելով ստանալը, այդ դեպքում հարսնյակները սկզբում կերի համար հավաքելու սովորությունը բնական ընտրության ազդեցության տակ կարող էր ուժեղանալ ու դառնալ զանազան նպատակներով ստրուկներ դաստիարակելու մշտական սովորություն։ Մի անգամ ձեռք բերված բնազդը, որը եթե նույնիսկ անհամեմատ ավելի քիչ չափով է զարգացած, քան մեր բրիտանական F. sanguinea տեսակինը, իսկ այդ տեսակը, ինչպես մենք տեսանք, իր ստրուկների ավելի քիչ ծառայություններից է օգտվում, քան հենց միևնույն տեսակը Շվեյցարիայում, կարող է բնական ընտրության ազդեցության տակ թե՛ ավելի ևս զարգանալ և թե՛ փոփոխվել, ի հարկե այն պայմանով, որ նրա փոփոխություններից յուրաքանչյուրը օգտակար է տեսակի համար, մինչև որ վերջապես կստացվի մի մրջյուն, որ այնքան ամոթալիորեն կախված է իր ստրուկներից, ինչպես Formica ruescens-ն է։
Մեղուների շինարարական բնազդը։ Ես այստեղ այդ հարցը չեմ քննարկի իր ամբողջ մանրամասնություններով, բայց կհաղորդեմ ընդհանուր գծերով այն եզրակացությունները, որոնց ես հանգել եմ։ Միայն տխմարը կարող է դիտել մեղրահացի զարմանալի կառուցվածքը, որ այնքան կատարյալ կերպով հարմարված է որոշ նպատակների համար, և չընկնել ծայրահեղ հիացմունքի մեջ։ Մաթեմատիկոսների վկայությամբ մեղուները գործնականում լուծել են մի դժվարին խնդիր՝ կառուցել բջիջներ, որոնք հնարավորին չափ մեծ քանակությամբ մեղր են պարունակում, ծախսելով նրանց կառուցման վրա հնարավորին չափ քիչ քանակությամբ թանկարժեք մեղրամոմ։ Նկատված է նաև, որ նույնիսկ հմուտ բանվորի համար, որն ունի անհրաժեշտ գործիքներն ու չափերը, շատ դժվարին կլիներ շինել մեղրամոմից պատշաճ ձևի բջիջներ, մինչդեռ աշխատավոր մեղուները դրանք պատրաստում են, աշխատելով մութ փեթակում։ Կարելի է բնազդներին վերաբերվել ինչպես ցանկալի է, բայց սկզբում բոլորովին անհասկանալի է թվում ոչ միայն այն, թե ինչպես մեղուները կարողանում են կառուցել բոլոր անհրաժեշտ անկյուններն ու կողքերը, այլև նրանք ընդունակություն ունեն իմանալու, որ ամեն ինչ ճիշտ է արված։ Սակայն դժվարությունն արդեն այնքան էլ մեծ չէ, ինչպես թվում է առաջին անգամ, ես կարծում եմ, կարելի է ապացուցել, որ այդ հիանալի շինվածքը կառուցվել է պարզ բնազդների միջոցով։
Այդ հարցն ուսումնասիրելու ինձ դրդել է մ-ր Ուոտերհաուզը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 87] որը ցույց է տվել, որ բջջի ձևը սերտ կախման մեջ է հարևան բջիջների ներկայությունից, և այդ պատճառով ստորև արտահայտված տեսակետը գուցե պետք է դիտել միայն որպես նրա տեսության մի ձևափոխություն։ Որպես ելակետ ընդունենք հաջորդական զարգացման մեծ սկզբունքը և տեսնենք, արդյոք բնությունն ինքը մեզ չի՞ բացատրի, թե ինչպես է կատարվում նրա աշխատանքը։ Կարճ շարքի մի ծայրում մենք ունենք իշամեղուներին, որոնք մեղր պահելու համար օգտագործում են իրենց հին բոժոժները, որոնց վրա նրանք երբեմն շինում են մեղրամոմե կարճ խողովակներ, իսկ երբեմն էլ պատրաստում են առանձին և չափազանց անկանոն մեղրամոմե կլորիկ բջիջներ։ Այդ շարքի մյուս ծայրում են գտնվում մեղուների բջիջները, որոնք դասավորված են երկու շարքով, յուրաքանչյուր բջիջ, ինչպես հայտնի է, ներկայացնում է վեցանիստ հատվածակողմ, որի հիմքը այնպես է հատված, որ կազմվում է երեք շեղանկյուններից բաղկացած բուրգ։ Այդ շեղանկյուններն ունեն որոշակի անկյուններ, և մեղրահացի մի կողմի վրա գտնվող առանձին բջիջի բրգաձև հիմքը առաջացնող երեք շեղանկյունիները մտնում են հակառակ կողմի երեք հարակից բջիջների կազմի մեջ։ Այդ շարքում, մեղուների չափազանց կատարեգործված և իշամեղուների պարզ բջիջների միջև, մենք ունենք մեքսիկական Melipona domestica-ի բջիջները, որոնք մանրազնին կերպով նկարագրել ու նկարել է Պյոտր Հուբերը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 168] Melipona-ն ինքն իր կառուցվածքով միջակա տեղ է գրավում մեղվի և իշամեղվի միջև, բայց վերջինիս ավելի մոտ կանգնած։ Նա կառուցում է գլանաձև բջիջներից բաղկացած բավական կանոնավոր մեղրամոմի մեղրահացեր, որոնց մեջ մեծանում են նրա թրթուրները և, բացի ղրանից, մի քանի խոշոր բջիջներ մեղր պահելու համար։ Վերջինները համարյա գնդաձև են, մոտավորապես հավասար մեծությամբ և հավաքված են, կազմելով անկանոն զանգված։ Առանձնապես պետք է ուշադրություն դարձնել այն հանգամանքի վրա, որ այդպիսի բջիջները կառուցվում են մեկը մյուսին այնքան մոտիկ, որ եթե գնդերը լինեին լրիվ, նրանք կհատեին մեկը մյուսին կամ էլ իրար մեջ կմտնեին, այդպես, սակայն, երբեք չի լինում, որովհետև այն տեղերում, որտեղ գնդերը պետք է իրար հետ հատվեն, մեղուները նրանց միջև կառուցում են բոլորովին տափակ մեղրամոմե պատեր։ Դրա հետևանքով յուրաքանչյուր բջիջ կազմված է արտաքին գնդաձև մասից և երկու, երեք կամ ավելի տափակ մակերեսներից, նայած այն բանին, թե բջիջը հարում է երկու, երեք կամ ավելի թվով այլ բջիջների։ Երբ մի բջիջ կռթնում է ուրիշ երեքի վրա, որ անհրաժեշտորեն հաճախ է լինում բջիջների մոտավորապես միանման մեծության դեպքում, երեք տափակ մակերեսները միանալով՝ կազմում են բուրգ, որը, ինչպես նկատում է Հուբերը, ներկայացնում է մեղվի բջջի բրգաձև եռանիստ հիմքի կոպիտ նմանություն։ Ճիշտ այնպես, ինչպես մեղվի բջիջներինն է, մի բջջի երեք տափակ մակերեսները անհրաժեշտորեն մտնում են հարակից երեք բջիջների կազմի մեջ։ Ակներև է, որ կառուցման այդ եղանակով Melipona-ն խնայում է մեղրամոմ և, որն ավելի ես կարևոր է, աշխատանք, որովհետև հարևան բջիջների միջև եղած տափակ պատերը ոչ թե կրկնակի, այլ միևնույն հաստությունն ունեն, ինչ որ արտաքին գնդաձև մասերը և յուրաքանչյուր տափակ պատ կազմում է երկու բջջի մաս։
Խորհելով դրա վրա, ես եկել եմ այն եզրակացության, որ եթե Melipona-ն կառուցեր իր գնդերը մեկը մյուսից որոշակի հեռավորությամբ, շիներ դրանք միևնույն մեծությամբ և դասավորեր համաչափորեն երկու շարքով, այդ դեպքում կառուցումը կլիներ նույնքան կատարյալ, ինչքան մեղվի մեղրահացերն են։ Այդ հիման վրա ես գրել եմ Քեմբրիջ՝ պրոֆեսոր Միլլերին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 178] և այդ երկրաչափը սիրալիր կերպով ստուգել է իր ցուցումների հիման վրա արված հետևյալ կշռադատությունները և գտել է, որ նրանք միանգամայն ճիշտ են։
Արտագծենք որոշ թվով հավասար գնդեր, որոնց կենտրոնները տեղավորված են երկու զուգահեռ շարքում, յուրաքանչյուր գնդի կենտրոնը գտնվում է միևնույն չարքի շրջապատող վեց գնդերի կենտրոններից շառավիղը 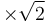 , կամ շառավիղը
, կամ շառավիղը  1,41421 (կամ ավելի ևս փոքր) հեռավորության վրա և մյուս զուգահեռ շարքի հարակից գնդերի կենտրոններից միևնույն հեռավորության վրա. այն ժամանակ, կառուցելով երկու շարքերի փոխադարձ հատման հարթությունները, մենք կստանանք վեցանիստ հատվածակողմերի կրկնակի շարք, որոնք միմյանց հետ միացած են երեք շեղանկյունիներից կազմված բրգաձև հիմքերով. և ինչպես շեղանկյունիները, այնպես էլ վեցանիստ հատվածակողմերի կողմերը, ամենամանրազնին չափումների համաձայն, կունենան այն բոլոր անկյունները, որոնք համապատասխանում են մեղվի մեղրահացի բջիջների նույնական անկյուններին։ Բայց ես իմացել եմ պրոֆ. Ուայմենից,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 36] որը կատարել է բազմաթիվ մանրազնին չափումներ, որ մեղվի աշխատանքի ճշտակատարությունը չափազանցված է, որովհետև ինչպիսին էլ որ լինի բջջի տիպական ձևը, այն շատ հազվագյուտ դեպքում է իրագործված լինում, եթե միայն առհասարակ իրագործվում է այն։
1,41421 (կամ ավելի ևս փոքր) հեռավորության վրա և մյուս զուգահեռ շարքի հարակից գնդերի կենտրոններից միևնույն հեռավորության վրա. այն ժամանակ, կառուցելով երկու շարքերի փոխադարձ հատման հարթությունները, մենք կստանանք վեցանիստ հատվածակողմերի կրկնակի շարք, որոնք միմյանց հետ միացած են երեք շեղանկյունիներից կազմված բրգաձև հիմքերով. և ինչպես շեղանկյունիները, այնպես էլ վեցանիստ հատվածակողմերի կողմերը, ամենամանրազնին չափումների համաձայն, կունենան այն բոլոր անկյունները, որոնք համապատասխանում են մեղվի մեղրահացի բջիջների նույնական անկյուններին։ Բայց ես իմացել եմ պրոֆ. Ուայմենից,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 36] որը կատարել է բազմաթիվ մանրազնին չափումներ, որ մեղվի աշխատանքի ճշտակատարությունը չափազանցված է, որովհետև ինչպիսին էլ որ լինի բջջի տիպական ձևը, այն շատ հազվագյուտ դեպքում է իրագործված լինում, եթե միայն առհասարակ իրագործվում է այն։
Ուրեմն մենք կարող ենք վստահությամբ եզրակացնել, որ եթե հնարավոր լիներ գոնե մի քիչ փոփոխելու Melipona-ի բնազդները, որ ամենևին առանձնապես զարմանալի չեն իրենց այժմյան տեսքով, ապա այդ միջատը կկառուցեր նույնպիսի հիանալի կատարելագործված մեղրահաց, ինչպիսին է մեղվինը։ Ենթադրենք, թե Melipona-ն ընդունակություն ունի կառուցելու միանման մեծությամբ կանոնավոր գնդաձև բջիջներ, որը նույնիսկ մեծ անսպասելիություն չէր լինի, քանի որ նա որոշ չափով այդ անում է, իսկ շատ այլ միջատներ ծառի մեջ շինում են միանգամայն գլանաձև անցքեր, հավանորեն պտտվելով որոշ կետի շուրջը։ Ենթադրենք նաև, թե Melipona-ն իր բջիջները դասավորում է կանոնավոր շարքերով, ինչպես նա արդեն անում է իր գլանաձև բջիջների նկատմամբ, և այնուհետև մենք պետք է ենթադրենք, հենց որի մեջ է ամենամեծ դժվարությունը, որ միջատը կարողանում է որոշ չափով ճիշտ դատել այն մասին, թե ինչ հեռավորության վրա պետք է կանգնել հարևան աշխատավորներից, երբ մի քանի անհատներ իրար կողքի կառուցում են իրենց բջիջները, բայց չէ որ նա արդեն այնքան հեռու է գնացել հեռավորության մասին դատելու ընդունակությամբ, որ գնդերը միշտ գծում է այնպես, որ նրանք պետք է միմյանց հատեն որոշ տարածության վրա և, բացի դրանից, հատման կետերը միացնում է բոլորովին տափակ մակերեսներով։ Ես կարծում եմ, որ մեղուն բնազդների այդպիսի փոփոխությամբ բնական ընտրության ազդեցության տակ ձեռք է բերել իր աննման շինարարական ընդունակությունները, իսկ այդ բնազդներն իրենց պարզ ձևով ամենևին առանձնապես զարմանալի չեն և հազիվ թե ավելի զարմանալի են, քան այն, որն ստիպում է թռչունին բուն շինել։
Բայց այդ տեսությունը կարելի է ստուգել փորձով։ Հետևելով Տեգետմեյերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 179] օրինակին, ես բաժանեցի երկու մեղրահաց և նրանց միջև դրեցի մի երկար, հաստ ուղղանկյուն մոմաշերտ, մեղուներն անմիջապես սկսեցին նրա մեջ փորել փոքրիկ կլոր փոսիկներ և դրանք խորացնելու հետ զուգընթաց նաև լայնացնում էին, մինչև որ նրանք դարձան ոչ խոր փոսեր, որոնք աչքաչափով թվում էին կանոնավոր գնդի մի մասը, որի տրամագիծը մոտավորապես հավասար էր բջջի տրամագծին։ Չափազանց հետաքրքիր էր դիտել, երբ մի քանի մեղուներ սկսում էին փորել իրենց փոսիկները մեկը մյուսի մոտ, աշխատելով այնպիսի հեռավորության վրա, որ ժամանակին այդ փոսիկներն ստանում էին վերոհիշյալ լայնությունը (այսինքն մոտավորապես սովորական բջջի լայնությունը), ունենալով այն գնդի տրամագծի շուրջ 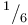 -ը, որի մի մասն էին կազմում նրանք, ըստ որում փոսիկների եզրերը հատում էին միմյանց և անցնում մեկը մյուսին։ Հենց որ այդ տեղի էր ունենում, մեղուները դադարում էին փոսիկներ խորացնել և սկսում էին մեղրամոմից տափակ պատեր կառուցել փոսիկների հատման գծերի ուղղությամբ. այսպիսով, յուրաքանչյուր վեցանիստ հատվածակազմ կառուցվում էր ոչ խոր փոսիկի ատամնավոր եզրի վրա, այլ ոչ թե եռանիստ բուրգի ուղիղ եզրերի վրա, ինչպես այդ լինում է սովորական բջիջների կառուցման դեպքում։
-ը, որի մի մասն էին կազմում նրանք, ըստ որում փոսիկների եզրերը հատում էին միմյանց և անցնում մեկը մյուսին։ Հենց որ այդ տեղի էր ունենում, մեղուները դադարում էին փոսիկներ խորացնել և սկսում էին մեղրամոմից տափակ պատեր կառուցել փոսիկների հատման գծերի ուղղությամբ. այսպիսով, յուրաքանչյուր վեցանիստ հատվածակազմ կառուցվում էր ոչ խոր փոսիկի ատամնավոր եզրի վրա, այլ ոչ թե եռանիստ բուրգի ուղիղ եզրերի վրա, ինչպես այդ լինում է սովորական բջիջների կառուցման դեպքում։
Դրանից հետո հասա ուղղանկյուն մեղրամոմի կտորի փոխարեն ես փեթակում տեղավորեցի բարակ նեղ սուր եզրով և կինաբարով ներկված մոմաշերտ։ Մեղուներն իսկույնևեթ սկսեցին նրա մեջ երկու կողմից փորել մանր փոսիկներ, մեկը մյուսին մոտիկ, ինչպես որ առաջ, բայց մեղրամոմը այնքան բարակ էր, որ հակադիր կողմերի փոսիկների հիմքերը պիտի մեկը մյուսի դիմաց ծակվեին, եթե փոսիկները այնչափ խորացվեին, որչափ որ նախորդ փորձում։ Սակայն մեղուները մինչև այդտեղ չհասցրին և ժամանակին դադարեցին փորել փոսիկները, վերջինների մեջ շինելով տափակ հատակ, հենց որ քիչ խորացել էին նրանք, և այդ տափակ հատակը, որ կազմվում էր կարմիր մեղրամոմի բարակ թիթեղիկից, որը մնացել էր չկրծոտված, ինչքան այդ մասին կարելի էր դատել աչքաչափով, ընկնում էր ճիշտ մոմեթիթեղիկի հակադիր կողմերի փոսերի երևակայական հատման տեղում։ Տեղ-տեղ հակադիր փոսիկների միջև մնում էին շեղանկյունի թիթեղիկների կտորներ, տեղ-տեղ էլ մեծ մասեր։ բայց անբնական պայմանների պատճառով աշխատանքը մաքուր չէր կատարվել։ Որպեսզի հատման տեղում աշխատանքը դադարեցնելիս փոսիկների միջև ստացվեն տափակ պատեր, մեղուները պետք է, կրծոտելով ու խորացնելով, կարմիր մեղրամոմի երկու կողմից էլ աշխատեին համարյա միանման արագությամբ։
Ի նկատի առնելով մեղրամոմի բարակ թիթեղիկի ճկունությունը, ես գրավարություն չեմ տեսնում նրանում, որ մեղուները նույնիսկ աշխատելով թիթեղիկի երկու կողմերի վրա, կարող են իմանալ, թե երբ են կրծոտել այն մինչև հարկ եղած սահմանը, որպեսզի դադարեցնեին աշխատանքը։ Ինձ թվում է, որ սովորական մեղրահացի կառուցման դեպքում մեղուները երկու կողմից միշտ բոլորովին միանման արագությամբ չեն աշխատում, գոնե ես նկատել եմ դեռ նոր սկսած բջջի հիմքի մոտ կիսավարտ շեղանկյունիները, որոնք քիչ գոգավոր էին մի կողմի վրա, որտեղ ես կարծում եմ, խորացումն ավելի արագ էր կատարվել և ուռուցիկ էր մյուս կողմում, որտեղ մեղուներն ավելի դանդաղ էին աշխատել։ Մի շատ հետաքրքրական դեպքում ես մեղրահացը կրկին դրեցի փեթակի մեջ, թողնելով, որ մեղուները նրա վրա մի որոշ ժամանակ դարձյալ աշխատեն, հետո նորից հետազոտեցի բջիջը և գտա, որ շեղանկյունաձև թիթեղիկն ավարտվել է և դարձել բոլորովին տափակ, ուշադրության առնելով այն, որ այդ թիթեղիկը չափազանց բարակ էր, միանգամայն անհնարին է ընդունել, որ մեղուներն են նրան տափակեցրել կրծելով ուռուցիկ կողմը. ես կասկածում եմ, որ այդպիսի դեպքերում նրանք կանգնում են հակադիր կողմերում, հրում ու ճնշում փափուկ տաքացրած մեղրամոմը (որը, ինչպես ես փորձեցի, հեշտությամբ է ստացվում) հարկ եղած տեղում և այսպիսով նրան տափակեցնում են։
Կինաբարով ներկված մեղրամոմի թիթեղի վրա կատարված փորձից կարելի է տեսնել, որ եթե մեղուներն իրենք կառուցեին իրենց համար բարակ մեղրամոմե պատ, նրանք կկարողանային պատրաստել հարկավոր մեծության բջիջներ, տեղավորվելով մեկը մյուսից պատշաճ հեռավորության վրա, աշխատելով միևնույն արագությամբ և ջանալով կառուցել միանման գնդաձև փոսիկներ, բայց թույլ չտալով գնդերին մտնելու մեկը մյուսի մեջ։ Զննելով կառուցվող մեղրահացի եզրը, կարելի է պարզ կերպով տեսնել, որ մեղուները մեղրահացի շուրջը շինում են հաստ եզրակ կամ շրջանակ և հետո կրծոտում են այն հակառակ երեսից, միշտ պատրաստելով կլոր փոսիկներ յուրաքանչյուր բջջի համար։ Նրանք միանգամից չեն անում բջջի եռանիստ բրգաձև հիմքը, այլ պատրաստում են երբեմն մեկ, երբեմն էլ երկու շեղանկյունի թիթեղիկներ, նայած թե նրանցից մեկը, թե երկուսն է ընկնում կառուցվող մեղրահացի ամենածայրին, և երբեք չեն ավարտում շեղանկյունի թիթեղիկների վերին եզրերը, քանի դեռ չեն սկսել հատվածակողմային պատերի կառուցումը։ Այս դիտումներից մի քանիսը տարբերվում են ըստ արժանվույն հռչակված Հուբեր ավագի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 168] կատարած դիտումներից, բայց ես համոզված եմ նրանց ճշտության միջև, եթե տեղը թույլ տար, ես կարող էի ապացուցել, որ նրանք համապատասխանում են իմ տեսությանը։
Հուբերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 168] պնդումը, թե ամենաառատին բջիջը փորվում է զուգահեռ պատերով, փոքրիկ մոմաթիթեղի մեջ, որքան ես կարող եմ դատել, այնքան էլ ճիշտ չէ. մեղրահացի սկիզբը միշտ ներկայացնում է մեղրամոմի թասակ, բայց ես այստեղ չեմ մտնի մանրամասնությունների մեջ։ Մենք տեսանք, թե բջիջների կառուցման մեջ ինչպիսի կարևոր ղեր է կատարում փոսիկներ պատրաստելը, բայց մեծ սխալ կլիներ ենթադրել, թե մեղուները չեն կարող մեղրահացի անհարթ եզրին կառուցում կատարել հարկ եղած ձևով, այսինքն պատրաստելով միջնապատեր հարևան գնդերի հատման հարթություններում։ Ես ունեմ մի քանի նմուշներ, որոնք անկասկած ապացուցում են, որ մեղուները կարող են աշխատել այդ ձևով։
Կառուցվող մեղրահացի նույնիսկ կոպիտ կերպով պատրաստված մեղրամոմե եզրակի կամ մեղրամոմե պատի մեջ երբեմն կարելի է տեսնել ծռվածքներ, որոնք իրենց դիրքով համապատասխանում են ապագա բջիջների հիմնական շեղանկյունաձև թիթեղիկների հարթություններին։ Բայց հաստ մոմե շրջանակի մեջ կառուցումը համենայն դեպս ավարտվում է մոմի թե մեկ և թե մյուս կողմից լայն չափերով գործադրվող կրծոտման միջոցով։ Մեղուների ընտրած կառուցման եղանակը շատ հետաքրքրական է. նրանք միշտ շինում են առաջին կոպիտ պատը 10—20 անգամ ավելի հաստ, քան բջիջի չափազանց բարակ, բոլորովին պատրաստի պատը, որը վերջ ի վերջո մնում է նրանից։ Այդ աշխատանքի կատարումը մեզ հասկանալի կդառնա, եթե մենք ենթադրենք, թե որմնադիրները սկզբում պատրաստում են ցեմենտից հաստ պատ, իսկ հետո սկսում են երկու կողմից հիմքի մոտ տաշել այն, մինչև որ մեջտեղում կմնա շատ բարակ, հարթ պատ։ Միաժամանակ որմնադիրները շարունակ հավաքում են տաշված ցեմենտը և նորից ավելացնում են պատի վերին եզրի վրա։ Այսպիսով, մեր առջև կլինի մի բարակ պատ, որը աստիճանաբար աճում է դեպի վեր, բայց միշտ պսակված է հսկայական քիվով։ Քանի որ բոլոր բջիջները, ինչպես հենց նոր սկսվածները, այնպես էլ բոլորովին ավարտվածները, ծածկված են մեղրամոմի մեծ զանգվածով, ուստի մեղուները կարող են խմբվել և սողալ մեղրահացի վրայով, չվախենալով, որ կարող են վնասել հատվածակողմերի բարակ պատերը։ Այդ պատերն իրենց հաստությամբ, համաձայն պրոֆ. Միլլերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 178] հաշվումների, որ նա սիրալիր կերպով արել է ինձ համար, խիստ տատանվում են, մեղրահացի եզրին մոտիկ արված 12 չափումներով այստեղ նրանց հաստությունը միջին հաշվով 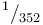 դյույմ է, մինչդեռ հիմքային շեղանկյունաձև թիթեղիկները համարյա մեկ երրորդականով ավելի հաստ են, ունենալով մոտավորապես
դյույմ է, մինչդեռ հիմքային շեղանկյունաձև թիթեղիկները համարյա մեկ երրորդականով ավելի հաստ են, ունենալով մոտավորապես 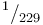 դյույմ միջին հաստություն, որը վերցված է 21 չափումներից։ Կառուցման յուրահատուկ եղանակի շնորհիվ շարունակ պահպանվում է մեղրահացի ամրությունը` պահպանելով մեղրամոմի խիստ խնայողություն։
դյույմ միջին հաստություն, որը վերցված է 21 չափումներից։ Կառուցման յուրահատուկ եղանակի շնորհիվ շարունակ պահպանվում է մեղրահացի ամրությունը` պահպանելով մեղրամոմի խիստ խնայողություն։
Առաջին հայացքից թվում է, թե բջիջների կառուցման ձևն ըմբռնելու դժվարությունը միայն ավելանում է կառուցմանը մասնակցող մեղուների քանակությունից, մեղուն կարճ ժամանակ աշխատելով բջջի վրա, անցնում է մյուսին, այնպես, որ, ինչպես ասում է Հուբերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 168] տասնյակ անհատներ աշխատում են առաջին բջջի սկզբնավորման վրա։ Ինձ հաջողվել է գործնականում ապացուցել այդ, ծածկելով կարմիր մեղրամոմի շատ բարակ շերտով միակ հատվածակողմաձև բջջի պատերի եզրերը կամ մեղրամոմի եզրակի արտաքին եզրերը. այդպիսի դեպքում ես միշտ տեսել եմ, որ մեղուներն այդ գույնը շատ համաչափորեն տարածում էին ամբողջ մեղրահացի վրա, այնքան համաչափ կերպով, ինչպես նկարիչն այդ կաներ վրձինով. ներկված մեղրամոմի մասնիկները վերցվում էին այն տեղերից, որտեղ նրանք կպցրած էին և զետեղվում էին կառուցվող բջիջների եզրերին։ Կառուցման ամբողջ աշխատանքն իրենից ներկայացնում է յուրահատուկ հավասարակշռություն, որը պահպանում են բազմաթիվ աշխատող մեղուները, նրանք բոլորը բնազդաբար կանգնում են մեկը մյուսից հավասար հեռավորության վրա, աշխատում են պատրաստել միանման գնդեր և կամ վերևում կառուցումը շարունակելով, կամ թողնելով չկրծոտված, շինում են հատման հարթությունները գնդերի միջև։ Իրոք հետաքրքրական էր դիտել, թե ինչպես դժվարին դեպքերում, օրինակ, երբ երկու մեղրահացեր հանդիպում են՝ կազմելով այս կամ այն անկյունը, մեղուները ստիպված էին լինում մի քանի անգամ քանդել և ամենատարբեր ձևով վերակառուցել միևնույն բջիջը, երբեմն վերադառնալով այն ձևին, որն սկզբից իրենք խոտանել էին։
Երբ մեղուները տեղ ունեն, որտեղ նրանք կարող են աշխատելու համար պատշաճ ձևով տեղավորվել, օրինակ, փայտի մի կտորի վրա՝ դեպի ներքև կառուցվող մեղրահացի ուղղակի մեջտեղում, այնպես որ մեղրահացը պետք է կառուցվի այդ փայտի կտորի մեկ կողմից, այդպիսի դեպքում մեղուները կարող են նոր բջջի մեկ պատի հիմքը դնել հենց այնտեղ, որտեղ հարկավոր է, և մյուս ավարտված բջիջների սահմաններից դուրս։ Բավական է, որ մեղուները կարողանան անհրաժեշտ դիրք գրավել ինչպես մեկը մյուսի, այնպես էլ վերջին ավարտված բջիջների պատերի վերաբերմամբ, այն ժամանակ նրանք, գծելով երևակայական գնդերը, կարողանում են կառուցել միջնապատեր երկու հարևան գնդերի միջև. բայց, որքան ես առիթ եմ ունեցել դիտելու, նրանք երբեք չեն կրծոտում ու չեն վերջացնում բջջի անկյունները, քանի դեռ չի կառուցված ինչպես այդ, այնպես էլ հարևան բջիջների մեծ մասը։ Մեղուների այդ ընդունակությունը, որ նրանք որոշ պայմաններում միջնապատերը հիմնում են հարկ եղած տեղերում երկու հենց նոր սկսած բջիջների միջև, շատ կարևոր է, որովհետև հիմնված է այն փաստի վրա՝ որն առաջին հայացքից կործանարար է թվում նախորդ տեսության համար, այն է՝ որ պիծակների մեղրահացի արտաքին եզրի վրա երբեմն լինում են կատարյալ վեցանիստ բջիջներ. սակայն ես այստեղ չեմ կարող կանգ առնել այդ հարցի վրա։ Նմանապես ես մեծ դժվարություն չեմ տեսնում նաև նրանում, որ եզակի մի միջատ (օրինակ՝ մայր պիծակը) կառուցում է վեցանիստ բջիջներ, եթե նա փոփոխակի կերպով աշխատել է երկու կամ երեք միաժամանակ սկսած բջիջների մերթ ներսի, մերթ դրսի կողքից, շարունակ գտնվելով հենց նոր սկսված բջիջների մասերից որոշակի հարաբերական հեռավորության վրա գծելով գնդեր կամ գլաններ և կառուցելով միջակա հարթություններ։
Քանի որ բնական ընտրությունը ներգործում է բացառապես անհատի գոյության տվյալ պայմաններում նրան օգտակար բնազդների կամ կառուցվածքի թույլ փոփոխությունների կուտակման միջոցով, ուստի մենք լիակատար հիմք ունենք հարցնելու, ի՞նչ օգուտ են ձեռք բերել մեղվի նախնիները շինարարական բնազդի այդ հաջորդական փոփոխությունների երկար շարքից, որոնք հանգեցրել են կառուցման այժմյան կատարելագործված եղանակին։ Ես կարծում եմ, որ այդ հարցին դժվար չէ պատասխանել. բջիջները, որոնք կառուցված են ինչպես մեղուների և պիծակների բջիջները, շահել են ամրության տեսակետից և դրա հետ միասին խնայվել են, մի կողմից, շատ աշխատանք ու տեղ, մյուս կողմից՝ շինանյութ, որից կառուցվում են նրանք։ Ինչ վերաբերում է մեղրամոմի առաջացմանը, ապա մենք գիտենք, որ մեղուները հաճախ բավարար քանակությամբ նեկտարի շատ մեծ կարիք են զգում, և մ-ր Տեգետմեյերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 179] ինձ հաղորդում է, որ ինչպես փորձով ապացուցված է, մեկ ֆունտ մեղրամոմ արտաթորելու համար փեթակը գործ է ածում 12-ից մինչև 15 ֆունտ չոր շաքար. սրանից հետևում է, որ փեթակում մեղրահացերի կառուցման վրա ծախսվող անհրաժեշտ մեղրամոմն արտաթորելու համար մեղուները պետք է հավաքեն ու սպառեն ահագին քանակությամբ հեղուկ նեկտար։ Բացի դրանից, մոմ արտաթորելու պրոցեսի ժամանակ բազմաթիվ մեղուներ պետք է շատ օրեր անգործ մնան։ Մեծ քանակությամբ մեղուներ ձմեռվա ընթացքում պահելու համար անհրաժեշտ է մեղրի մեծ պաշար, իսկ ինչպես հայտնի է, փեթակի գոյության ապահովությունը կախված է առավելապես պահպանվող մեղուների մեծ քանակությունից։ Ուրեմն, մեղրի, ինչպես նաև նրա հավաքման վրա ծախսված աշխատանքի ուժեղ տնտեսման միջոցով մոմի խնայողությունը պետք է հանդիսանա ամեն մի մեղվաընտանիքի բարգավաճման էական բաղադրիչը։ Իհարկե, տեսակի բարգավաճումը կարող է կախված լինել նրա թշնամիների, մակաբույծների և բոլորովին առանձնահատուկ պատճառների թվից և այդ իմաստով կախում չունի մեղրի այն քանակությունից, որ կարող են հավաքել մեղուները։ Բայց ենթադրենք, թե վերջին հանգամանքը որոշում է, ինչպես հավանորեն իրականում հաճախ և որոշել է, թե կարո՞ղ է արդյոք մեր իշամեղվին մոտիկ մի մեղու գոյություն ունենալ մեծ քանակությամբ որևէ երկրում. այնուհետև ենթադրենք, թե իշամեղուների համայնքը գոյություն է ունեցել ձմեռվա ընթացքում և, հետևաբար, մեղրի պաշարի կարիք է ունեցել, այդ դեպքում կասկած չի կարող լինել, որ մեր երևակայած իշամեղվի համար ձեռնտու էր նրա բնազդներում այդպիսի փոփոխությունը, որի ուղղությունն է՝ կառուցել մոմե բջիջներ այնքան մոտ, որպեսզի նրանք մի քիչ փոխադարձաբար հատվեն, որովհետև երկու կողք-կողքի գտնվող բջիջների ընդհանուր պատն անգամ կարող է խնայել որոշ քանակությամբ թե՛ աշխատանք և թե՛ մեղրամոմ։ Այս հիման վրա մեր իշամեղուների համար ավելի ու ավելի ձեռնտու կլիներ, եթե նրանք իրենց բջիջները պատրաստեին ավելի կանոնավոր, մեկը մյուսին ավելի մոտիկ և նրանց հավաքեին կույտերում՝ Melipona-ի բջիջների նման, այդ դեպքում յուրաքանչյուր բջջի սահմանազատող մակերեսի մեծ մասը միաժամանակ կծառայեր հարևան բջիջների սահմանազատման համար, և դրանով իսկ կխնայվեր շատ աշխատանք և մոմ։ Բայց հետո, դարձյալ նույն հիման վրա, Melipona-ի համար ձեռնտու կլիներ բջիջներ պատրաստել մեկը մյուսից ավելի մոտիկ և ընդհանրապես ավելի կանոնավոր, քան անում է նա այժմ, որովհետև այդ դեպքում, ինչպես մենք տեսանք, գնդաձև մակերեսները կարող էին իսպառ անհետանալ, իրենց տեղը զիջելով տափակ մակերեսներին, և Melipona-ն կկարողանար նույնքան կատարելագործված մեղրահաց պատրաստել, ինչքան այն, որ կառուցում է մեր մեղուն։ Ճարտարապետության մեջ դրանից ավելի կատարելության աստիճանի չէր կարող հասցնել բնական ընտրությունը, որովհետև, ինչպես մենք տեսանք, մեղվի մեղրահացերը բացարձակապես կատարյալ են աշխատանքի ու մոմի տնտեսման տեսակետից։
Այսպիսով, ինչպես ես կարծում եմ, բոլոր հայտնի բնազդներից ամենազարմանալին՝ մեղվի շինարարական բնազդը կարելի է բացատրել բնական ընտրության ներգործությամբ, որն օգտագործել է պարզ բնազդների բազմաթիվ հաջորդական ու թույլ փոփոխությունների առավելությունները և հետզհետե մեղուներին հասցրել է այնտեղ, որ նրանք ավելի ու ավելի կատարյալ ձևով գծեն հավասար գնդեր մեկը մյուսից որոշ հեռավորության վրա, դրանք դասավորելով երկու շարքով և կառուցեն թե՛ մոմե պատերը և թե խորացնեն մեղրամոմը գնդերի հատման հարթությունների ուղղությամբ։ Մեղուներն իհարկե նույնքան քիչ բան գիտեն այն մասին, որ իրենք գնդեր են շինում մեկը մյուսից որոշակի հեռավորության վրա, ինչքան այն մասին, թե ինչպիսիք են վեցանիստ հատվածակողմերի անկյուններն ու նրանց հիմքային շեղանկյունիները. քանի որ բնական ընտրությունը հարուցող հանգամանքների էությունն այն է, որ կառուցվեն թրթուրների համար բջիջներ, որոնք աշխատանքի և մոմի հնարավորին չափ տնտեսման հետ լինեն բավականաչափ ամուր, անհրաժեշտ մեծության և ձևի, ապա այն պարսը, որը կառուցում է ամենալավ բջիջները աշխատանքի նվազագույն ծախսումով և նվազագույն քանակությամբ մեղր է կորցնում մեղրամոմ արտաթորելու վրա, ունենում է առավելագույն հաջողություն և փոխանցում է իր նոր ձեռք բերած տնտեսողական բնազդները նոր ձագերի, որոնք իրենց հերթին գոյության կռվում հաջողության համար ունենում են լավագույն հնարավորությունները։
ԱՌԱՐԿՈՒԹՅՈՒՆՆԵՐ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՏԵՍՈՒԹՅՈՒՆԸ ԲՆԱԶԴՆԵՐԻ ՎԵՐԱԲԵՐՄԱՄԲ ԿԻՐԱՌԵԼՈՒ ԴԵՄ. ԱՆՍԵՌ ԵՎ ԱՆՊՏՈՒՂ ՄԻՋԱՏՆԵՐ
Բնազդների ծագման մասին վերևում շարադրված տեսակետի դեմ է առարկություն, թե «կառուցվածքի և բնազդի փոփոխությունները պետք է տեղի ունենան միաժամանակ և ճիշտ կերպով համապատասխանեն մեկը մյուսին, որովհետև մի ուղղությամբ փոփոխությունը, առանց մյուս ուղղությամբ անհապաղ կատարվող համապատասխան փոփոխության կունենար ճակատագրական հետևանքներ»։ Այս առարկության ծանրակշռությունը ամբողջապես կախված է այն ենթադրությունից, որ բնազդի և կառուցվածքի փոփոխությունները գալիս են հանկարծակիորեն։ Վերցնենք, օրինակի համար, նախորդ գլխում բերված դեպքը մեծ երաշտահավի (Parus major) մասին. այս թռչունը, հաճախ ոտների մեջ ճյուղի վրա սեղմելով կենու սերմերը, այնքան է փորում կտուցով, մինչև որ հասնում է միջուկին։ Բայց ի՞նչ առանձին դժվարություն կարող է ներկայացնել այն, որ բնական ընտրությունը պահպաներ թույլ անհատական փոփոխությունները կտուցի կառուցվածքում, որպեսզի նա ավելի ու ավելի լավ հարմարվեր սերմերը բաց անելու համար, մինչև նա կդառնար այդ նպատակի համար նույնքան լավ պիտանի, որքան սիտտայի կտուցն է, և որպեսզի միևնույն ժամանակ սովորությունը, ճաշակի հարկադրական կամ քմահաճ փոփոխությունը թռչունին կդարձնեն ավելի ու ավելի հատիկակեր։ Այս դեպքում մենք ընդունում ենք, որ կտուցը դանդաղորեն և աստիճանաբար փոփոխվում է բնական ընտրությամբ, համապատասխանելով սովորությունների ու ճաշակի աստիճանական փոփոխություններին. բայց թող երաշտահավի ոտները կամ կտուցի փոփոխության կապակցությամբ, կամ որևէ անհայտ պատճառից կախված նույնպես փոփոխվեն և դառնան ուժեղ, այդ դեպքում բավական հավանական կլինի, որ ոտները կստիպեն թռչունին ավելի ու ավելի շատ մագլցել, մինչև որ նա ձեռք կբերի սիտտայի զարմանալի բնազդը և մագլցելու ընդունակությունը։ Այդ դեպքում ենթադրվում է, որ կառուցվածքի աստիճանական փոփոխությունը ուղեկցում է բնազդական սովորությունների փոփոխությանը։ Վերցնենք էլի մի օրինակ։ Քիչ բնազդներ կան ավելի ուշագրավ, քան այն բնազդը, որը հարկադրում է Խաղաղօվկիանոսյան կղզիների ծիծեռնակին բներ կառուցել բացառապես թանձրացած թքից։ Որոշ թռչուններ բները կառուցում են, ինչպես կարծում են, թքով ցեմենտացրած տիղմից, հյուսիսամերիկական մի ծիծեռնակ, ինչպես ես տեսել եմ, իր բունը շինում է թքով սոսնձված ջախերից և նույնիսկ այդ նյութի գնդիկներից։ Ուրեմն մի՞թե շատ անհավանական է, որ շարունակ ավելի ու ավելի շատ թուք արտադրող ծիծեռնակների անհատների բնական ընտրությունը կարող էր վերջապես առաջացնել այնպիսի բնազդներ ունեցող տեսակ, որոնք կհարկադրեին ծիծեռնակներին արհամարհել այլ նյութերը և բուն շինել միայն թանձրացած թքից։ Այդպես էլ այլ դեպքերում։ Սակայն պետք է խոստովանել, որ շատ դեպքերում մենք չենք կարող որոշել, թե ինչի՞ց է սկսվել փոփոխությունը՝ բնազդի՞ց, թե կառուցվածքից։
Ոչ մի կասկած չկա, որ դժվար բացատրելի շատ բնազդներ կարող են հակադրվել բնական ընտրության տեսությանը. դրանք կամ այնպիսի դեպքեր են, երբ մենք չենք կարող հետամուտ լինել, թե ինչպես է զարգացել բնազդը, կամ այնպիսիներն են, երբ անհայտ են բնազդի աստիճանական բարդացման անցումային աստիճանները, կամ այնպիսիներն են, երբ բնազդի դերն այնքան աննշան է, որ հազիվ թե նա կարող էր զարգանալ բնական ընտրության ազդեցության տակ․ կամ, վերջապես համարյա նույն բնազդների՝ երևալու դեպքերը այնպիսի կենդանիների մոտ, որոնք բնության սիստեմում մեկը մյուսից այնքան հեռու են կանգնած, որ մենք չենք կարող նրանց նմանությունը բացատրել մեկ ընդհանուր նախահորից ժառանգական փոխանցման միջոցով և, հետևաբար պետք է ընդունենք, որ նրանք ձեռք են բերվել մեկը մյուսից անկախ կերպով՝ բնական ընտրության ազդեցության ներքո։ Ես այստեղ կանգ չեմ առնի բոլոր նման դեպքերի վրա և կսահմանափակվեմ ամենակարևոր դժվարության քննարկությամբ, որը սկզբում ինձ թվում էր անհաղթահարելի և, իսկապես, ամբողջ տեսությանս համար ճակատագրական։ Ես խոսում եմ միջատների համայնքների մեջ եղած անսեռ կամ անպտուղ էգերի մասին, որոնք հաճախ շատ ուժեղ կերպով տարբերվում են արուներից և սովորական էգերից ինչպես բնազդով, այնպես էլ կառուցվածքով և, լինելով անպտուղ, չեն կարող արտադրել իրենց նմաններ։
Այս հարցը արժանի է բազմակողմանի քննարկման, բայց ես այստեղ կվերցնեմ միմիայն աշխատավոր կամ անպտուղ մրջյունների դեպքը։ Թե ինչպես են աշխատավորները դարձել անպտուղ, դժվար է բացատրել, բայց ավելի դժվար չէ, քան կառուցվածքում ամեն մի այլ խիստ փոփոխություն, որովհետև կարելի է ապացուցել, որ մի քանի միջատներ և այլ հոդվածավոր կենդանիներ երբեմն դառնում են անպտուղ բնական պայմաններում, իսկ եթե այդպիսիները պատկանում են հասարակական միջատների թվին և հասարակության համար ձեռնտու է, որ ամեն տարի ծնվում են որոշ թվով անհատներ, որոնք աշխատունակ են, բայց բազմացման համար անընդունակ են, ապա ես առանձին դժվարություն չեմ տեսնում այն բանում, որ նրանք կարող էին ընկնել բնական ընտրության ազդեցության տակ։ Բայց այդ սկզբնական դժվարության վրա կարելի է կանգ չառնել։ Մեծ դժվարություն են ներկայացնում աշխատավոր մրջյունները, որոնք խիստ տարբերվում են ինչպես արուներից, այնպես էլ էգերից իրենց կառուցվածքով, կրծքի ձևով, թևերի, երբեմն էլ աչքերի բացակայությամբ և բնազդով։ Ինչ վերաբերում է միայն բնազդին, ապա այն զարմանալի տարբերությունը, որ այդ տեսակետից գոյություն ունի աշխատավոր և նորմալ էգերի միջև, կարող է ամենից ավելի լավ քննվել մեղուների վրա։ Եթե աշխատավոր մրջյունը կամ մի այլ անսեռ միջատ լիներ սովորական կենդանի, ես առանց վարանելու կարող էի ընդունել, որ նրա բոլոր առանձնահատկությունները աստիճանաբար ձեռք են բերվել բնական ընտրության ազդեցության տակ. այսինքն՝ սկզբում կարող էին ծնվել թույլ չափով օգտակար առանձնահատկություններ ունեցող անհատներ, որոնք իրենց այդ հատկությունները փոխանցել են իրենց հետնորդներին. սրանք էլ փոփոխվել ու իրենց հերթին ընտրվել են և այլն։ Բայց աշխատավոր մրջյունը շատ խիստ տարբերվում է իր ծնողներից, միևնույն ժամանակ բոլորովին անպտուղ է. հետևաբար նա ոչ մի կերպ չի կարող փոխանցել կառուցվածքի կամ բնազդի հաջորդականորեն ձեռք բերված փոփոխություններն իր ժառանգներին, և այդ պատճառով կարելի է ամենայն իրավամբ հարցնել, հնարավո՞ր է արդյոք այս դեպքը համաձայնեցնել բնական ընտրության տեսության հետ։
Նախ և առաջ հիշենք, որ ինչպես ընտանի, այնպես էլ վայրենի կենդանիները հանդես են բերում կառուցվածքի մեջ ժառանգական բազմազան փոփոխությունների անհամար դեպքեր, որոնք կապված են կամ հասակի, կամ սեռի հետ։ Կան փոփոխություններ, որոնք կապված են ոչ միայն սեռի, այլև, բացի դրանից, վերարտադրողական օրգանների գործունեության կարճատև շրջանի հետ, ինչպես, օրինակ, ներկայացնում են շատ թռչունների հարսանեկան զարդարանքները կամ արու սալմոնի ծնոտի ծռվելը։ Մենք տեսնում ենք, որ ընտանի անասունների զանազան ցեղերի եղջյուրների նույնիսկ թույլ փոփոխությունները կախման մեջ են գտնվում արհեստական եղանակով արուների մեջ առաջացրած անկատար վիճակի հետ, որովհետև մի քանի ցեղերի եզների եղջյուրներն ավելի երկար են, քան մյուսների եղջյուրները՝ համեմատած միևնույն ցեղերի ցուլերի և կովերի եղջյուրների երկարության հետ։ Ուստի ես մեծ դժվարություն չեմ կարող տեսնել այն բանում, որ որևէ առանձնահատկություն կապ է ունենում միջատների համայնքի որոշ անդամների անպտղության հետ. դժվարությունը այն հասկանալն է, թե ինչպես կառուցվածքի այդպիսի աննշան հարաբերակցական փոփոխությունները կարող էին աստիճանաբար ուժեղանալ բնական ընտրության ազդեցության տակ։
Սակայն այդ դժվարությունը, թեև անհաղթահարելի է թվում, նվազում է և, իմ կարծիքով, նույնիսկ իսպառ անհետանում է, եթե մենք հիշենք, որ ընտրությունը կարող է ընտանիքին վերաբերել նույնպես, ինչպես որ անհատին և ինչպես մի, այնպես էլ մյուս դեպքում հասցնել որոշ նպատակի։ Անասնաբույծները ցանկանում են, որ միսն ու ճարպը միացած լինեն որոշ ձևով, և թեպետ այդ հատկություններն ունեցող կենդանին սպանդանոց է տարվում, սակայն անասնաբույծը ամենայն վստահությամբ առաջացնում է այդ ցեղը, և նրան այդ հաջողվում է։ Որքան մենք պիտի հավատանք ընտրության ուժին, որպեսզի ընդունենք, թե եղջերավոր անասունների այն ցեղը, որը միշտ առաջացնում է շատ երկար եղջյուրներով եզներ, ամենայն հավանականությամբ ստեղծվել է ցուլերի և կովերի այնպիսի անհատների մանրազնին ընտրությամբ, որոնք զուգավորվելով՝ ծնել են ամենից ավելի երկար եղջյուրներով եզներ, թեև եզն ինքը չի կարող շարունակել իր ցեղը։ Բայց այդ տեսակետից կարելի է բերել ավելի ևս լավ ու ավելի համոզիչ օրինակ։ Միամյա շահպրակների մի քանի տարատեսակները, ինչպես ասում է Վերլոն,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 180] երկարատև ու մանրազնին ընտրության միջոցով հասած լինելով որոշ աստիճանի կատարելության, միշտ տալիս են զգալի քանակությամբ սերմնաբույսեր, որոնք ունենում են բոլորովին անպտուղ բազմաթերթ ծաղիկներ, բայց նմանապես տալիս են նաև որոշ քանակությամբ պարզ, պտղատու բույսեր։ Վերջինները, որոնց շնորհիվ միայն տարատեսակը կարող է բազմանալ, կարելի է համեմատել մրջյունների պտղատու արուների և էգերի հետ, իսկ բազմաթերթ անպտուղ բույսերը համապատասխանում են նույն համայնքի անպտուղ անհատներին։ Ինչպես շահպրակի տարատեսակի, այնպես էլ հասարակական միջատների մոտ որոշ նպատակի հասնելու ձգտող ընտրությունը կիրառվել է ընտանիքի, այլ ոչ թե անհատի նկատմամբ։ Սրանից մենք պետք է հետևեցնենք, որ կառուցվածքում և բնազդի մեջ տեղի ունեցող փոքրիկ փոփոխությունները, որոնք կապված են համայնքի որոշ անդամների անպտղության հետ, օգտակար են եղել, դրա շնորհիվ պտղաբերող արուներն ու էգերը բարգավաճել են և իրենց հերթին փոխանցել են իրենց պտղաբերող հետնորդներին միևնույն փոփոխություններն ունեցող անպտուղ անհատներ առաջացնելու հակումը։ Այդ պրոցեսը պիտի կրկնվեր բազմիցս անգամ, մինչև որ միևնույն տեսակի պտղաբերող ու անպտուղ էգերի միջև տարբերությունը կհասներ այն չափերին, որ մենք նկատում ենք զանազան հասարակական միջատների մոտ։
Բայց մենք դեռ չենք շոշափել քննարկվող դժվարության գագաթնակետը, այսինքն այն, որ մի քանի մրջյունների անսեռ անհատները տարբերվում են ոչ միայն պտղաբերող արուներից և էգերից, այլև տարբերվում են միմյանցից, երբեմն անհավատալի չափով՝ բաժանվելով երկու և մինչև անգամ երեք կատեգորիայի։ Բացի դրանից, այդ կաստաները աստիճանաբար չեն անցնում ու փոխարկվում մեկը մյուսի, այլ խիստ կտրուկ կերպով սահմանազատված են և այնքան են տարբերվում իրարից, որքան տարբերվում են մի սեռի երկու տեսակներ կամ նույնիսկ մի ընտանիքի երկու սեռեր մեկը մյուսից։ Այսպես, Eciton-ը ունի անսեռ աշխատավորներ և զինվորներ շատ տարբեր ծնոտներով ու բնազդներով․ Cryptocerus-ի մի կաստայի աշխատավորներն ունեն գլխի վրա յուրահատուկ վահան, որի գործածությունը միանգամայն անհայտ է. մեքսիկական Myrmecocystus-ի մի կաստայի աշխատավորները երբեք չեն հեռանում բնից, կերակրվում են մի այլ կաստայի աշխատավորների օգնությամբ, բայց ունեն չափազանց զարգացած որովայն և արտաթորում են մեղրանման մի հյութ, որը փոխարինում է այն օշարակին, որ արտաթորում են լվիճները կամ ընտանի անասունները, ինչպես նրանց կարելի է անվանել, որոնց արածեցնում և պահում են գերության մեջ մեր եվրոպական մրջյունները։
Գուցե կարելի է կարծել, թե ես չափազանցրած հավատ ունեմ դեպի բնական ընտրության սկզբունքը, եթե ընդունում եմ, որ այդքան զարմանալի փաստերը, ապացուցված լինելով, միանգամից չեն խորտակում տեսությունս։ Այն ավելի պարզ դեպքից, որ մի կաստայի անսեռ միջատները առաջացել են, ինչպես ես կարծում եմ, պտղաբերող արուներից և էգերից բնական ընտրության միջոցով սովորական փոփոխությունների անալոգիայով, կարելի է եզրակացնել, որ հաջորդական, թույլ բարենպաստ փոփոխությունները միանգամից չեն առաջացել մի բնի բոլոր անսեռ անհատների մեջ, այլ սկզբում երևացել են քչերի մեջ, և որ այն համայնքները, որոնց էգերն ամենամեծ քանակությամբ են առաջացրել բարենպաստ փոփոխություններ ունեցող անսեռ անհատներ, գերապրել են, որի հետևանքով բոլոր անսեռ անհատները ձեռք են բերել վերջապես այդ առանձնահատկությունները։ Այս տեսակետի համաձայն, մենք պետք է հույս ունենանք միևնույն բնում երբեմն գտնելու անսեռ միջատներ, որոնք ներկայացնում են կառուցվածքի հաջորդական փոփոխություն. և մենք իրոք գտնում ենք դրանց և նույնիսկ հաճախ, եթե ուշադրության առնենք այն, թե Եվրոպայի սահմաններից դուրս ինչքան քիչ անսեռ միջատներ ենթարկվել են մանրազնին հետազոտման։ Մ-ր Ֆ. Սմիտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 177] ցույց է տվել, որ բրիտանական զանազան մրջյունների անսեռ անհատները զարմանալի կերպով տարբերվում են միմյանցից մեծությամբ և երբեմն էլ գույնով, և որ այդպիսի ծայրային ձևերը կարող են միացած լինել մեկը մյուսի հետ միևնույն բնից վերցրած անհատներով. ես ինքս համեմատել եմ այդպիսի հիանալի աստիճանական անցումները։ Երբեմն պատահում է, որ առանձնապես բազմաթիվ են ավելի խոշոր կամ ավելի մանր աշխատավորները կամ թե մեկը և մյուսը բազմաթիվ են, իսկ միջակա անհատները՝ սակավաթիվ։ Formica flava-ն ունի խոշոր և մանր աշխատավորներ միջին մեծությամբ սակավաթիվ անհատների հետ միասին, և այդ տեսակի խոշոր աշխատավորները, մ-ր Ֆ. Սմիտի դիտումների համաձայն, ունենում են պարզ աչքեր (ocelli) թեև փոքրիկ, բայց պարզ կերպով նկատելի, մինչդեռ մանրերի մոտ նրանք թերաճ են։ Այս աշխատավորներից մի քանի անհատների մանրազնին հերձման հիման վրա ես կարող եմ հաստատել, որ մանր աշխատավորների աչքերը անհամեմատ ավելի թերաճ են, քան այդ կարելի էր սպասել միայն նրանց փոքրիկ մեծության հիման վրա. և ես լիովին համոզված եմ, թեև չեմ կարող այդ նույնքան էլ վճռականորեն հաստատել, որ միջակ մեծություն ունեցող աշխատավորների պարզ աչքերն իրենց զարգացմամբ հենց միջին տեղ են գրավում։ Ուրեմն՝ այս դեպքում մենք ունենք մեկ բնում անպտուղ աշխատավորների երկու խումբ, որոնք տարբերվում են իրարից ոչ միայն մեծությամբ, այլև տեսողության օրգաններով, ինչպես նաև կապված են միմյանց հետ միջին դրություն գրավող սակավաթիվ անհատներով։ Ես կարող եմ սրանով վերջացնել, ավելացնելով, որ եթե մանր աշխատավորները շատ օգտակար լինեին համայնքի համար և ընտրությունը շարունակաբար տարածվեր այն արուների և էգերի վրա, որոնք արտադրում են ավելի ու ավելի մեծ քանակությամբ մանր աշխատավորներ, մինչև որ բոլոր աշխատավորները կդառնային այդպիսիներ, այդ դեպքում մենք կունենայինք մրջյունի մի տեսակ, որի անսեռ անհատները մոտավորապես նման են նրանց, որոնք մենք տեսնում ենք Myrmica-ի մոտ։ Myrmica-ի աշխատավորները չունեն նույնիսկ թերաճ պարզ աչքեր, թեև այդ մրջյունի արուների ու էգերի աչքերը լավ զարգացած են։
Բերեմ էլի մի օրինակ։ Խորապես վստահ լինելով, որ միևնույն տեսակի անսեռ ձևի զանազան կաստաների միջև հնարավոր է գտնել աստիճանական անցումներ կառուցվածքի էական գծերում, ես մեծ գոհունակությամբ օգտագործել եմ արևմտյան Աֆրիկայից մ-ր Սմիտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 177] բերած Anomma մրջյունի բազմաթիվ անհատները, որոնք վերցված են մի բնից։ Գուցե ընթերցողն ավելի լավ պատկերացնի այդ աշխատավորների մեծությունների տարբերությունը, եթե ես պարզ չափերի փոխարեն ակներևության համար բերեմ այսպիսի համեմատություն՝ պատկերացնենք, որ մենք տեսնում ենք տուն կառուցող բանվորներ, որոնցից ոմանք ունեն հինգ ոտնաչափ և չորս դյույմ բարձրություն, իսկ մյուսները՝ տասնը վեց ոտնաչափ, դրան ավելացնենք, որ մեծ բանվորների գլուխը ոչ թե երեք, այլ չորս անգամ մեծ է փոքրերի գլխից, ծնոտներն էլ հինգ անգամ մեծ են։ Զանազան մեծություն ունեցող աշխատավոր մրջյունների ծնոտները առանձնապես զարմանալի բազմազանություն են ներկայացնում իրենց տեսքով, ձևով և ատամիկների թվով։ Բայց մեզ համար կարևոր է այն, որ թեև աշխատավորներն իրենց մեծությամբ կարող են բաժանվել մի քանի կաստաների, սակայն նրանց միջև գոյություն ունեն մի շարք աստիճանական անցումներ, ինչպես որ անցումներ կան նաև նրանց զարմանալիորեն բազմազան ծնոտների միջև։ Ես համարձակորեն եմ ասում վերջինի մասին, որովհետև սըր Ջ. Լեբբոկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 60] լյուցիդա-խցիկի միջոցով ինձ համար նկարել է զանազան աշխատավոր մրջյունների ծնոտներ, որոնք կտրված էին իմ ձեռքով։ Մ-ր Բետսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 181] իր «Բնախույզը Ամազոն գետի վրա» հետաքրքրական գրքում բերել է համանման տվյալներ։
Ունենալով իմ առջև այսպիսի փաստեր, ես չեմ կասկածում, որ բնական ընտրությունը պտղաբեր մրջյունների կամ մրջյուն ծնողների վրա ներգործելով, կարող էր առաջ բերել այնպիսի տեսակ, որը միշտ կառաջացներ մի ձևի ծնոտներով ավելի ու ավելի մեծ անսեռ անհատներ, կամ բոլորն էլ խիստ տարբեր ծնոտներով փոքր մեծությամբ անհատներ, կամ, վերջապես, որն ամենամեծ դժվարությունն է, մեկ խումբը մի մեծությամբ ու կառուցվածքով և միաժամանակ մյուս խումբը՝ այլ մեծությամբ ու կառուցվածքով. ըստ որում սկզբում կառաջանային մի շարք հաջորդական անցումներ, ինչպես մենք այդ տեսնում ենք աֆրիկական մրջյունի մոտ, իսկ հետո ծայրային ձևերը երևան կգային ավելի ու ավելի մեծ թվով, որպես նրանց արտադրող ծնողների գերապրելու հետևանք, մինչև որ միջակա անհատները բոլորովին կդադարեին երևան գալուց։
Համանման բացատրություն է առաջարկել մ-ր Ուոլլեսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] մալայան թիթեռների մի քանի տեսակների մեջ երկու կամ նույնիսկ երեք տարբեր ձևի էգերի երևան գալու նույնքան բարդ դեպքի համար և Ֆրից Մյուլլերը՝[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] արուների երկու շատ տարբեր ձևեր ունեցող բրազիլական մի քանի խեցեմուրթների համար։ Բայց կարիք չկա այստեղ քննարկել այդ երևույթները։
Այսպիսով, ես կարծում եմ, ինձ հաջողվեց բացատրել անպտուղ աշխատավորների երկու խիստ տարբեր կաստաների ծագումը, որոնք գոյություն ունեն միևնույն բնում, բայց խիստ տարբերվում են թե մեկը մյուսից և թե իրենց ծնողներից։ Թե որքան այդ օգտակար է մրջնային համայնքների համար, կարելի է հասկանալ ելնելով աշխատանքի բաժանման հենց այն սկզբունքից, որն այնքան մեծ օգուտ է բերում քաղաքակրթված մարդուն։ Սակայն մրջյուններն աշխատում են ժառանգված բնազդների և ժառանգված օրգանների կամ գործիքների միջոցով, մինչդեռ մարդն աշխատում է ձեռք բերված գիտելիքի և հատկապես պատրաստված գործիքների միջոցով։ Բայց ես պիտի խոստովանեմ, որ չնայած իմ ամբողջ հավատին դեպի ընտրությունը՝ ես երբեք չէի կարող ենթադրել, թե այդ սկզբունքը կարող է այդքան մեծ չափով արտահայտվել, եթե ինձ այդ եզրակացության չբերեին անսեռ միջատների ներկայացրած օրինակները։ Դրա համար էլ ես այդ հարցի վրա քիչ ավելի կանգ առա, թեև ընդհանրապես ոչ բավականաչափ մանրամասնորեն, ինչպես բնական ընտրության նշանակությունն ապացուցելու համար, այնպես էլ այն պատճառով, որ այդ դեպքերը հանդիսանում են իմ տեսության ամենամեծ դժվարություններից մեկը։ Այդ դեպքերը հետաքրքրական են նաև որպես ապացույց այն բանի, որ կենդանին, ինչպես նաև բույսը, կարող է ենթարկվել ամեն մի փոփոխության բազմաթիվ թույլ պատահական փոփոխությունների կուտակման միջոցով, միայն թե նրանք որևէ բանով օգտակար լինեն՝ առանց վարժեցման կամ սովորության որևէ մասնակցության։ Յուրահատուկ սովորությունները, որ հատուկ են աշխատավոր կամ անպտուղ էգերին, ինչքան ժամանակ էլ գոյություն ունենային, իհարկե, չէին կարողանա ներգործել արուների և պտղաբերող էգերի վրա, որոնք միայն սերունդ են տալիս։ Եվ ինձ զարմացնում է այն, որ մինչև այժմ ոչ ոք չի օգտագործել անսեռ միջատների այդ ցուցադրական օրինակը ժառանգված սովորությունների մասին՝ Լամարկի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 2] պաշտպանած լավ հայտնի ուսմունքի դեմ։
ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ
Ես այս գլուխը նվիրեցի մեր ընտանի կենդանիների մտավոր ընդունակությունների փոփոխելիությունը և այդ փոփոխությունների ժառանգական փոխանցումը համառոտակի ապացուցելուն։ Ավելի ևս կարճառոտ կերպով ես աշխատեցի ապացուցել, որ բնազդները բնական պայմաններում թույլ չափով են փոփոխվում։ Ոչ ոք չի վիճարկի այն, որ բնազդները հսկայական նշանակություն ունեն յուրաքանչյուր կենդանու համար։ Այդ պատճառով էլ իրականում ոչ մի դժվարություն չկա նրանում, որպեսզի գոյության փոփոխված պայմաններում բնական ընտրությունը ուժեղացներ բնազդների թույլ փոփոխությունները մինչև որոշ աստիճան, եթե միայն նրանք օգտակար են որևէ ուղղությամբ։ Հնարավոր է, որ որոշ դեպքերում իրենց ազդեցությունն են թողնում թե սովորությունները և թե օրգանի վարժեցումը կամ ոչ վարժեցումը։ Ես չեմ հավակնում, որ այս գլխում բերված փաստերը գոնե որոշ չափով ամրապնդում են իմ տեսությունը, բայց բերված դժվարին դեպքերից ոչ մեկը, որքան ես կարող եմ դատել, չի հերքում այն։ Մյուս կողմից, այնպիսի փաստերը, ինչպիսին է այն, որ բնազդը միշտ չէ, որ կատարյալ է լինում և կարող է սխալների տեղիք տալ, որ ոչ մի բնազդի համար չի կարելի ապացուցել, թե նա առաջացել է այլ կենդանիների օգտին, թեև որոշ կենդանիներ օգտվում են ուրիշների բնազդներից, որ «Natura non facit saltum»[58]. բնապատմական օրենքը բնազդների նկատմամբ նույնքան կիրառելի է, որքան օրգանիզմի կառուցվածքի նկատմամբ, և միանգամայն բացատրելի է վերոհիշյալ տեսակների հիման վրա, բայց անբացատրելի է որևէ այլ եղանակով, այդ բոլորն ավելի շուտ ծառայում են բնական ընտրության տեսությունն ամրացնելու համար։
Այս տեսությունը հաստատվում է նաև բնազդներին վերաբերող մի քանի այլ փաստերով, օրինակ, այն սովորական դեպքով, երբ երկու մերձավոր, բայց տարբեր տեսակներ, որոնք ապրում են աշխարհի տարբեր երկրներում և գտնվում են բոլորովին տարբեր պայմաններում, հաճախ պահպանում են համարյա միևնույն բնազդները։ Օրինակ, ելնելով ժառանգականության սկզբունքից, մենք կարող ենք հասկանալ, թե ինչու Հարավային Ամերիկայի արևադարձային մասերի կեռնեխը իր բունը ծեփում է կավով, բոլորովին այնպես, ինչպես այդ անում է մեր բրիտանական կեռնեխը, թե ինչու Աֆրիկայի և Հնդկաստանի ռնգեղջյուր թռչուններն ունեն միևնույն յուրահատուկ բնազդը, նրանք իրենց էգերին փակում են ծառերի փչակներում, ծեփվածքի մեջ թողնելով միայն մի փոքրիկ անցք, որի միջոցով արուն կերակրում է էգին և ձագերին, երբ վերջինները դուրս են գալիս. թե ինչու հյուսիսամերիկական ցախսարեկի (Troglodytes) արուները շինում են այսպես կոչված «աքլորաբներ», որոնք ծառայում են որպես թառ, բոլորովին այնպես, ինչպես անում են մեր ցախսարեկի արուները, և այդ սովորությունը բոլորովին նման չէ մեզ հայտնի որևէ այլ թռչունի սովորությանը։ Վերջապես, թեև գուցե թե ոչ ամբողջապես այդ տրամաբանական է, բայց ես մեծ չափով բավարարություն եմ զգում, համարելով, որ այդպիսի բնազդները, ինչպիսիք են իր խորթ եղբայրներին բնից դուրս նետող մատղաշ կկուի բնազդը, ստրուկներ բռնող մրջյունների բնազդը, կենդանի թրթուրի ներսում սնվող իխնևմոնների թրթուրների բնազդը, ոչ թե դիտմամբ շնորհված կամ ստեղծված բնազդներ են, այլ միայն բոլոր օրգանական էակների զարգացումը պայմանավորող մի ընդհանուր օրենքի, այն է, բազմացման, փոփոխության, ամենաուժեղների գերապրման և թույլերի ոչնչացման փոքրիկ հետևանքներն են։
ԳԼՈՒԽ IX։ ՀԻԲՐԻԴԱՑՈՒՄ
ԱՌԱՋԻՆ ԽԱՉԱՁԵՎՈՒՄՆԵՐԻ ԵՎ ՀԻԲՐԻԴՆԵՐԻ ԱՆՊՏՂՈՒԹՅԱՆ ՄԻՋԵՎ ԵՂԱԾ ՏԱՐԲԵՐՈՒԹՅՈՒՆԸ։— ԱՆՊՏՂՈՒԹՅՈՒՆԸ ՏԱՐԲԵՐ է ԸՍՏ ԱՍՏԻՃԱՆԻ. ՆԱ ԸՆԴՀԱՆՐԱԿԱՆ ՉԷ. ՆԱ ՈՒԺԵՂԱՆՈՒՄ է ԱԶԳԱԿՑՈՒԹՅԱՆ ՄՈՏ ԱՍՏԻՃԱՆՆԵՐՈՒՄ ԽԱՉԱՁԵՎՄԱՆ ԱԶԴԵՑՈՒԹՅԱՆ ՏԱԿ Եվ ՎԵՐԱՆՈՒՄ Է ԸՆՏԱՆԵՑՄԱՄԲ ԵՎ ՄՇԱԿՈՒԹՅԱՄԲ։— ՀԻԲՐԻԴՆԵՐԻ ԱՆՊՏՂՈՒԹՅՈՒՆԸ ՈՐՈՇՈՂ ՕՐԵՆՔՆԵՐԸ։— ԱՆՊՏՂՈՒԹՅՈՒՆԸ ԻՆՔՆՈՒՐՈՒՅՆ ԱՌԱՆՁՆԱՀԱՏԿՈՒԹՅՈՒՆ ՉԷ, ԱՅԼ ԿԱՊՎԱԾ է ԱՅԼ ՏԱՐԲԵՐՈՒԹՅՈՒՆՆԵՐԻ ՀԵՏ ԵՎ ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՄԻՋՈՑՈՎ ՉԻ ԿՈՒՏԱԿՎԵԼ։— ԱՌԱՋԻՆ ԽԱՉԱՁԵՎՈՒՄՆԵՐԻ ԵՎ ՀԻԲՐԻԴՆԵՐԻ ԱՆՊՏՂՈՒԹՅԱՆ ՊԱՏՃԱՌՆԵՐԸ։— ԿԵՆՍԱՊԱՅՄԱՆՆԵՐԻ ՓՈՓՈԽՈՒԹՅԱՆ ԵՎ ԽԱՉԱՁԵՎՄԱՆ ՀԵՏԵՎԱՆՔՆԵՐԻ ՄԻՋԵՎ ԵՂԱԾ ՆՄԱՆՈՒԹՅՈՒՆԸ։— ԴԻՄՈՐՖԻԶՄ ԵՎ ՏՐԻՄՈՐՖԻԶՄ։— ԽԱՉԱՁԵՎՎՈՂ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԵՎ ՆՐԱՆՑ ԽԱՌՆԱՑԵՂԵՐԻ ՊՏՂԱԲԵՐՈՒԹՅՈՒՆԸ ՀԱՄԸՆԴՀԱՆՈՒՐ ԵՐԵՎՈՒՅԹ ՉԻ ՆԵՐԿԱՅԱՑՆՈՒՄ։— ՀԻԲՐԻԴՆԵՐԻ ԵՎ ԽԱՌՆԱՑԵՂԵՐԻ ՀԱՄԵՄԱՏՈՒԹՅՈՒՆԸ՝ ԱՆԿԱԽ ՆՐԱՆՑ ՊՏՂԱԲԵՐՈՒԹՅՈՒՆԻՑ։— ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ
Բնախույզների մեծամասնությունը այն կարծիքին է, թե տեսակները խաչաձևման դեպքում օժտված են անպտղությամբ նրա համար, որպեսզի կանխվի նրանց խառնածնությունը։ Այղ կարծիքը, իհարկե, առաջին հայացքից շատ ճշմարտացի է թվում, որովհետև միասին ապրող տեսակները հազիվ թե կարողանային անխառն մնալ, եթե նրանք ընդունակ լինեին ազատ կերպով խաչաձևվելու։ Այդ հարցը մեզ համար կարևոր է շատ տեսակետներից, մանավանդ այն պատճառով, որ առաջին խաչաձևման ենթարկվող տեսակների և նրանցից առաջացող հիբրիդների անպտղաբերությունը չի կարող ձեռք բերվել, ինչպես ես այդ ցույց կտամ, անպտղության հաջորդական օգտակար աստիճանների պահպանման ճանապարհով։ Նա ծնողական տեսակների արտադրողական սիստեմում եղած տարբերությունների կողմնակի հետևանքն է։
Այս հարցը քննարկելիս սովորաբար խառնվել են երկու կատեգորիայի փաստեր, որոնք ըստ էության շատ տարբեր են միմյանցից, այն է՝ տեսակների անպտղությունը առաջին խաչաձևման ժամանակ և նրանցից առաջացած հիբրիդների անպտղությունը։
Մաքուր տեսակների վերարտադրողական օրգանները, իհարկե գտնվում են կատարյալ վիճակում, սակայն այդ տեսակները խաչաձևման ժամանակ կամ քիչ սերունդ են արտադրում կամ թե բոլորովին չեն արտադրում։ Մյուս կողմից, հիբրիդների վերարտադրողական օրգաններն անընդունակ են կատարելու իրենց ֆունկցիան, ինչպես այդ կարելի է պարզ կերպով տեսնել ինչպես բույսերի, այնպես էլ կենդանիների արական էլեմենտի վիճակից, թեև օրգաններն իրենք, որքան մանրադիտակը թույլ է տալիս դատելու, կառուցվածքով միանգամայն կատարյալ են։ Առաջին դեպքում սեռական երկու տարրերը, որոնցից գոյանում է ապագա սաղմը, միանգամայն կատարյալ են. երկրորդ դեպքում նրանք կամ բոլորովին զարգացած չեն, կամ զարգացած են ոչ լիովին։ Այդ տարբերությունը կարևոր է, երբ քննարկելու կլինենք անպտղության պատճառը, որն ընդհանուր է երկու դեպքերի համար։ Այդ տարբերությունը հավանորեն աչքաթող է արվել այն պատճառով, որ երկու դեպքում էլ անպտղության վրա նայել են որպես մի հատկապես շնորհված բանի վրա, որը մեր ըմբռնողության սահմաններից դուրս է գտնվում։
Տարատեսակների, այսինքն այն ձևերի, որոնց մասին մենք գիտենք կամ ենթադրում ենք, որ նրանք ծագել են ընդհանուր ծնողներից խաչաձևման ժամանակ, ինչպես նաև նրանցից առաջացած խառնացեղերի պտղաբերությունը նույնպես կարևոր է իմ տեսության տեսակետից, ինչպես որ տեսակների անպտղությունը, որովհետև նա, ըստ երևույթին, խոշոր ու պարզորոշ տարբերություն է սահմանում տարատեսակների և տեսակների միջև։
ԱՆՊՏՂՈՒԹՅԱՆ ԱՍՏԻՃԱՆՆԵՐ
Ամենից առաջ դառնանք խաչաձևման ժամանակ տեսակների և նրանից առաջացած հիբրիդների անպտղությանը։ Անկարելի է ուսումնասիրել երկու բարեխիղճ և զարմանալի հետազոտողների և գրեթե իրենց ամբողջ կյանքը այդ հարցին նվիրած Կյոլռեյտերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] և Գերտների[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] զանազան մեմուարները և աշխատությունները, և խոր տպավորություն չստանալ այն բանից, որ միջտեսակային խաչաձևումները մինչև որոշ աստիճանը անպտուղ են։ Կյոլռեյտերն այդ կանոնից բացառություն չի ընդունում, բայց այդ դեպքում կտրում է հանգույցը, որովհետև տասը դեպքում, երբ հեղինակների մեծամասնության կողմից որպես առանձին տեսակներ ընդունված երկու ձևերը եղել են լիովին պտղաբեր, նա առանց տատանվելու այդ ձևերը համարում է որպես տարատեսակներ։ Գերտները նույնպես դրանից ընդհանուր կանոն է հանում և վիճարկում է Կյոլռեյտերի նշած տասը դեպքի լրիվ պտղաբերությունը։ Բայց այդ և շատ այլ դեպքերում Գերտներն ստիպված է խնամքով հաշվել սերմերը, որպեսզի ցույց տա որոշ աստիճանի անպտղության գոյությունը։ Նա միշտ համեմատում է սերմերի ամենամեծ թիվը, որ տալիս են երկու տեսակները խաչաձևման ժամանակ և նրանց հիբրիդային սերունդները, այն սերմերի միջին թվի հետ, որը տալիս են երկու ծնողական տեսակները բնական դրության մեջ։ Բայց այստեղ լուրջ սխալի առիթ է ներկայանում. բույսը խաչաձևելու համար հարկավոր է նրան նախապես ենթարկել կաստրացիայի, և, որ հաճախ ավելի ևս կարևոր է, նրան պետք է մեկուսացնել, որպեսզի արգելվի այլ բույսերի ծաղկափոշու տեղափոխությունը նրա վրա միջատների միջոցով։ Համարյա բոլոր բույսերը, որոնց վրա Կյոլռեյտերը կատարել է իր փորձերը, տնկված են եղել ծաղկամաններում և պահվել են իր տանը՝ սենյակում։ Որ այդ եղանակները հաճախ վնասակար ազդեցություն են ունենում բույսերի պտղաբերության վրա, դրանում կասկած չի կարող լինել։ Գերտներն ինքը իր աղյուսակում բերում է երկու տասնյակ դեպքեր, երբ նա բույսերին կաստրացիայի է ենթարկել և արհեստական եղանակով բեղմնավորել իրենց սեփական ծաղկափոշիով, ըստ որում այդ քսան բույսերից կեսի պտղաբերությունը (չհաշված այնպիսի բույսերի վրա կատարված փորձերը, ինչպես լոբազգիներն են, որոնց հետ վարվելը ակնհայտորեն շատ դժվար է), եղել է քիչ թուլացած։ Բացի դրանից, Գերտները բազմիցս խաչաձևել է մի քանի ձևեր, ինչպես օրինակ, սովորական կարմիր և կապույտ տզրուկախոտը (Anagallis arvensis և coerolea), որոնց լավագույն բուսաբանները համարում են տարատեսակներ, և գտել է, որ նրանք միանգամայն անպտուղ են. այդ պատճառով մենք իրավունք ունենք կասկածելու, թե շատ տեսակներ արդյո՞ք իսկապես այդպես անպտուղ են խաչաձևման ժամանակ, ինչպես նա կարծել է։
Մի կողմից, անկասկածելի է, որ տարբեր տեսակների խաչաձևման ժամանակ անպտղությունը այնքան տարբեր է ըստ աստիճանի և ներկայացնում է այնքան աննկատելի անցումներ, իսկ մյուս կողմից, որ մաքուր տեսակների պտղաբերության վրա այնքան հեշտությամբ են ազդում զանազան հանգամանքներ, որ բոլոր գործնական նպատակների համար շատ դժվար է ասել, թե որտեղ է վերջանում լրիվ պտղաբերությունը և որտեղ է սկսվում անպտղությունը։ Իմ կարծիքով, դրան ավելի լավ ապացույց չի կարելի պահանջել, քան այն փաստը, որ երբևիցե ապրածներից երկու ամենափորձված դիտողները, այսինքն՝ Կյոլռեյտերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] և Գերտները,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] միևնույն ձևերի վերաբերող մի քանի դեպքերում եկել են տրամագծորեն իրար հակառակ եզրակացությունների։ Չափազանց ուսանելի է համեմատել,— բայց տեղի սղության պատճառով ես չեմ կարող մանրամասնությունների մեջ մտնել,— նաև մեր լավագույն բուսաբանների ներկայացրած ապացույցները այն հարցի վերաբերյալ, թե կարո՞ղ են արդյոք որոշ կասկածելի ձևեր համարվել տեսակներ կամ տարատեսակներ, այն ապացույցների հետ, որոնք վերցվում են պտղաբերության փաստերից և բերվում են ձևերի խաչաձևմամբ զբաղվող տարբեր դիտողների կողմից, կամ տարբեր տարիների ընթացքում միևնույն դիտողի կատարած փորձերի հիման վրա։ Ուրեմն, կարելի է ցույց տալ, որ ոչ անպտղությունը և ոչ էլ պտղաբերությունը վստահելի հատկանիշներ չեն, որոնցով կարելի է տարբերել տեսակները տարատեսակներից։ Այս աղբյուրից բխող մատնանշումները ներկայացնում են նույնպիսի աննկատելի աստիճաններ և նույնքան կասկածելի են, որքան կազմվածքի ու կառուցվածքի այլ տարբերությունների վրա հիմնված ապացույցները։
Ինչ վերաբերում է հիբրիդների անպտղությանը հաջորդական սերունդներում, ապա թեև Գերտներին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] հաջողվել է վեց, յոթ և մի դեպքում նույնիսկ տասը սերունդների ընթացքում ստանալ մի քանի հիբրիդներ՝ խիստ կերպով թույլ չտալով նրանց խաչաձևումը մաքուր ծնողական ձևերի հետ, սակայն նա դրականապես հաստատում է, որ նրանց պտղաբերությունը երբեք չի բարձրանում, այլ սովորաբար խիստ և հանկարծակի կերպով նվազում է։ Պտղաբերության այդ նվազման վերաբերմամբ, ամենից առաջ կարելի է նկատել, որ կառուցվածքում կամ կազմվածքում երբ որևէ շեղում երկու ծնողների համար էլ ընդհանուր է, նա փոխանցվում է ժառանգներին ուժեղացած աստիճանով, մինչդեռ հիբրիդային բույսերի մոտ երկու սեռական տարրերը արդեն մի քիչ տուժած են լինում։ Բայց ես կարծում եմ, որ նրանց պտղաբերությունը գրեթե բոլոր դեպքերում նվազել է մի անկախ պատճառից, այսինքն նրա հետևանքով, որ խաչաձևումը կատարվել է միմյանց հետ չափազանց մոտ ազգակից անհատների միջև։ Ես այնքան փորձեր եմ կատարել և հավաքել այնքան փաստեր, որոնք ցույց են տալիս, մի կողմից, որ մի ուրիշ անհատի կամ ուրիշ տարատեսակի հետ պատահական խաչաձևումը մեծացնում է սերնդի ուժն ու պտղաբերությունը և, մյուս կողմից, որ չափազանց մոտ ազգականների խաչաձևումը նվազեցնում է նրանց ուժն ու պտղաբերությունը, որ ես չեմ կարող կասկածել այդ եզրակացության ճշտության վերաբերմամբ։ Փորձարարները հազվագյուտ դեպքում են առաջ բերում մեծ քանակությամբ հիբրիդներ, իսկ քանի որ ծնողական տեսակները կամ այլ ազգակից հիբրիդները սովորաբար աճում են միևնույն պարտեզում, ուստի ծաղկման շրջանում պետք է նրանց խնամքով պաշտպանել միջատների այցելությունից. հետևապես եթե նրանք թողնվեն իրենք իրենց, այդ դեպքում սովորաբար նրանք յուրաքանչյուր սերնդի ընթացքում կբեղմնավորվեն միևնույն ծաղկի ծաղկափոշիով, իսկ այդ հավանորեն կարող է վատ անդրադառնալ նրանց պտղաբերության վրա, որն արդեն թուլացած է նրանց հիբրիդային ծագման հետևանքով։ Այդ համոզմունքն իմ մեջ ամրացնում է այն ուշագրավ ցուցումը, որ բազմիցս արել է Գերտներր, այն է՝ որ եթե արհեստականորեն բեղմնավորենք նույնիսկ ամենաքիչ պտղաբերող հիբրիդներին միանման հիբրիդների ծաղկափոշիով, այդ դեպքում նրանց պտղաբերությունը, չնայած այդ գործողության հաճախ նկատվող վնասակար հետևանքներին, հաճախ վճռականորեն բարձրանում է և շարունակում է բարձրանալ։ Բայց քանի որ արհեստական բեղմնավորման ժամանակ ծաղկափոշին նույնքան էլ հաճախ վերցվում է պատահականորեն (ինչպես այդ ինձ հայտնի է իմ սեփական վարձերից) մի այլ ծաղկի փոշանոթներից, ինչպես նաև հենց բեղմնավորման ենթարկվող նույն ծաղկի փոշանոթներից, ուստի այդ դեպքում տեղի է ունենում խաչաձևում երկու, թեկուզև միևնույն բույսին պատկանող, ծաղիկների միջև։ Բացի դրանից, ամեն անգամ, երբ կատարվել են նման բարդ փորձեր, այնպիսի մանրազնին դիտողը, ինչպիսին է Գերտները, հավանորեն կաստրացիայի ենթարկել է իր հիբրիդներին և դրանով ապահովել է յուրաքանչյուր սերնդում խաչաձև բեղմնավորումը այլ ծաղկի ծաղկափոշու միջոցով, որը պատկանել է նույն բույսին կամ նույնպիսի հիբրիդային հատկություններ ունեցող մի այլ բույսի։ Ուրեմն՝ ինքնաբեղմնավորվող հիբրիդներին հակառակ արհեստականորեն բեղմնավորվող հիբրիդների հաջորդական սերունդների շարքում պտղաբերության բարձրացման տարօրինակ փաստը, ինչպես ինձ թվում է, կարելի է բացատրել նրանով, որ այդ դեպքում վերացվել է խաչաձևումը այնպիսի անհատների միջև, որոնք չափազանց մոտ ազգակցության մեջ են գտնվում։
Այժմ դառնանք այն արդյունքներին, որ ստացել է խաչաձևումների գործում չափազանց փորձված մի երրորդ դիտող, այսինքն՝ արժանահարգո Վ. Հերբերտը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 7] Նա պնդում է, որ որոշ հիբրիդներ միանգամայն պտղաբեր են՝ նույնքան պտղաբեր են, ինչպես որ մաքուր ծնողական տեսակները, և պնդում է այդ այնպիսի հաստատամտությամբ, ինչպես Կյոլռեյտերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] և Գերտները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] հավատացնում են, որ տեսակների խաչաձևումների ժամանակ նրանց որոշ աստիճանի անպտղաբերությունը բնական օրենք է, որը բացառություններ չի ընդունում։ Նա փորձերը կատարել է նույն բույսերից մի քանիսի վրա, որոնց վերցրել էր նաև Գերտները։ Արդյունքների տարբերությունը, ես կարծում եմ, կարելի է մասամբ բացատրել Հերբերտի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 7] որպես այգեգործի, մեծ հմտությամբ, մասամբ էլ նրանով, որ նա իր տրամադրության տակ ունեցել է ջերմատներ։ Նրա կարևոր շատ վկայություններից, օրինակի համար, ես կբերեմ միայն մեկը, այն է՝ որ «Crinum capense-ի ծաղկափոշիով բեղմնավորված Crinum revolutum-ի սերմնարանում յուրաքանչյուր սերմնաբողբոջ առաջ է բերել բույս, որը ես երբեք չեմ նկատել նրա բնական բեղմնավորման դեպքերում»։ Հետևաբար, մենք այստեղ ունենք լիակատար կամ նույնիսկ սովորականից ավելի քան լիակատար պտղաբերություն երկու տարբեր տեսակների առաջին խաչաձևման ժամանակ։
Crinum-ի այդ դեպքը ինձ առիթ է տալիս հիշելու մի ուշագրավ փաստի մասին, այն է, որ Lobelia-ի Yerbascum-ի և Passiflora-ի մի քանի տեսակների առանձին բույսեր հեշտությամբ բեղմնավորվում են այլ տեսակների ծաղկափոշիով, բայց ոչ նույն բույսի ծաղկափոշիով, թեև վերջինս միանգամայն առողջ է, որը կարելի է ապացուցել, նրանով բեղմնավորելով այլ բույսեր կամ տեսակներ։ Hippeastrum սեռի մեջ Corydalis-ի, ինչպես ցույց է տվել պրոֆ. Հիլդեբրանդը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 84] զանազան խոլորձիների մոտ, ինչպես ցույց են տվել Սկոտտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 182] և Ֆրից Մյուլլերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] բոլոր անհատներն ունեն այդ հատկությունը։ Այսպիսով, մի քանի տեսակների միայն նորմայից շեղվող անհատները, իսկ մյուսների բոլոր անհատները իսկապես ավելի հեշտությամբ կարող են հիբրիդային սերունդ տալ, քան թե բեղմնավորվել հենց միևնույն անհատից վերցված ծաղկափոշիով։ Բերենք մի օրինակ, Hippeastrum aulicum-ի կոճղեզը տվել է չորս ծաղիկ, Հերբերտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 7] նրանցից երեքը բեղմնավորել է սեփական ծաղկափոշիով, իսկ հետո չորրորդը բեղմնավորել է երեք զանազան տեսակներից առաջացած բարդ հիբրիդի ծաղկափոշիով. վերջում ստացվել է այն, որ «առաջին ծաղիկների սերմնարանները շուտով դադարել են աճելուց և մի քանի օրից հետո բոլորովին ոչնչացել են, մինչդեռ տուփիկը, որ փոշոտված էր հիբրիդի ծաղկափոշիով, սկսել է ուժեղ կերպով աճել, հասունացել է և տվել սերմեր, որոնք լավ ծլել են»։ Հերբերտն այդպիսի վարձեր կատարել է մի քանի տարիների ընթացքում և միշտ միանման արդյունքով, այս դեպքերը ցույց են տալիս, թե ինչպիսի չնչին ու խորհրդավոր պատճառներից երբեմն կախված է տեսակի ավելի մեծ կամ ավելի փոքր պտղաբերությունը։
Պարտիզագործների գործնական փորձերը, թեկուզև նրանք գիտական ճշտությամբ կատարված չլինեն էլ, արժանի են որոշ ուշադրության։ Բոլորին հայտնի է, թե ինչպիսի բարդ խաչաձևումների են ենթարկվել Pelargonium-ի Fuchsia-ի, Cabceolaria-ի, Petunia-ի, Rhododendron-ի և այլ տեսակները, բայց և այնպես այդ հիբրիդներից շատերը առատությամբ սերմ են տալիս։ Այսպես, օրինակ, Հերբերտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 7] պնդում է, որ Calceolaria integrifolia և C. plantaginea-ի արտաքին տեսքով որքան կարելի է միմյանց չնմանվող այդ տեսակների հիբրիդը «բազմանում է այնքան հիանալի կերպով, կարծեք թե դա չիլիական լեռների մի բնական տեսակ լինի»։ Ես քիչ ջանք չեմ գործադրել, որպեսզի ստույգ իմանամ ռոդոդենդրոնների մի քանի բարդ խաչաձևումների պտղաբերության աստիճանը և համոզվել եմ, որ նրանցից շատերը միանգամայն պտղաբեր են։ Մ-ր Ս. Նոբլը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 183] օրինակ, ինձ հաղորդել է, որ նա վայրակներ է բազմացնում պատվաստելու համար Rhododendron ponticum-ի և Rh. catawbiense-ի հիբրիդը և որ այդ հիբրիդը «այնպիսի առատությամբ սերմեր է տալիս, ինչպիսին միայն կարելի է պատկերացնել»։ Եթե հիբրիդների պտղաբերությունը լավ խնամքի դեպքում շարունակ նվազեր յուրաքանչյուր հաջորդ սերնդում, ինչպես այդ կարծում է Գերտները,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] ապա այդ փաստը քաջ ծանոթ կլիներ պարտիզագործներին, որոնք գործ ունեն տնկարանների հետ։ Պարտիզագործները միևնույն հիբրիդները բազմացնում են ամբողջ մեծ մարգերով, իսկ չէ՞ որ այստեղ միայն հանդիպում են հարկ եղած իրադրությանը, որովհետև այդ դեպքում առանձին անհատներ միջատների միջոցով հնարավորություն են ունենում ազատ կերպով խաչաձևվելու ամեն մի այլ անհատի հետ, և այսպիսով կանխվում է մոտ ազգակցական փոշոտման վնասակար ազդեցությունը։ Յուրաքանչյուր ոք կարող է հեշտությամբ համոզվել միջատների միջնորդության իրական լինելու մեջ, հետազոտելով հիբրիդային ռոդոդենդրոնների ամենաանպտղաբեր ցեղերի ծաղիկները, որոնք երբեք ծաղկափոշի չեն առաջացնում, նա այդ ծաղիկների վրա կգտնի առատ ծաղկափոշի, որը բերված է այլ ծաղիկներից։
Ինչ վերաբերում է կենդանիներին, ապա նրանց վրա անհամեմատ ավելի քիչ մանրազնին փորձեր են կատարվել, քան բույսերի վրա։ Եթե կարելի է վստահել մեր սիստեմատիկական ստորաբաժանումներին, այսինքն եթե կենդանիների սեռերը նույնքան տարբերվում են իրարից, որքան բույսերի սեռերը, այդ դեպքում մենք կարող ենք եզրակացնել, որ բնական սանդուխքի վրա միմյանցից ավելի հեռու կանգնած կենդանիները կարող են ավելի հեշտությամբ խաչաձևվել իրար հետ, քան նման պայմաններում գտնվող բույսերը. բայց հիբրիդներն իրենք, ինչպես ինձ թվում է, ավելի անպտղաբեր են։ Սակայն չպետք է մոռանալ, որ կենդանիների վրա քիչ փորձեր են կատարված հարկ եղած պայմաններում, որովհետև միայն քիչ կենդանիներ կան, որ բազմանում են գերության մեջ։ Այսպես, օրինակ, դեղձանիկը խաչաձևվել է սերինոսների ընտանիքից ինը զանազան տեսակների հետ, բայց քանի որ նրանցից ոչ մեկը ազատ կերպով չի բազմանում գերության մեջ, ուստի մենք իրավունք չունենք սպասելու, որ նրանց և դեղձանիկի միջև առաջին խաչաձևումները կամ դրանցից առաջացած հիբրիդները լինեն լիովին պտղաբեր։ Այնուհետև, ինչ վերաբերում է ամենապտղաբեր կենդանական հիբրիդների պտղաբերությանը հաջորդական սերունդների մեջ, ապա ինձ հազիվ թե հայտնի է գոնե մի դեպք, երբ միևնույն հիբրիդի երկու ընտանիք առաջ բերված լինեին միաժամանակ տարբեր ծնողներից, որպեսզի կանխվեին ազգակցության մոտ աստիճաններում կատարվող խաչաձևումների վնասակար հետևանքները։ Ընդհակառակը, յուրաքանչյուր հաջորդ սերնդում սովորաբար խաչաձևվել են եղբայրները քույրերի հետ՝ հակառակ այն նախազգուշացմանը, որ շարունակ կրկնում են բոլոր անասնաբույծները։ Իսկ այդպիսի դեպքում բոլորովին զարմանալի չէ, որ հիբրիդների հատուկ անպտղաբերությունը ավելի ու ավելի աճել է։
Թեև ինձ հայտնի չէ կենդանական հիբրիդների կատարյալ պտղաբերության գրեթե ոչ մի լիովին ստույգ դեպք, սակայն ես հիմք ունեմ կարծելու, որ Cervulus[59] vaginalis-ից և G. Reevesii-ից, ինչպես Phasianus colchicus-ից և Ph. torquatus-ից ստացված հիբրիդները լիովին պտղաբեր են։ Կատրֆաժը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 184] նշում է, որ երկու մետաքսագործների (Bombyx cynthia և B. arrindia) հիբրիդները Փարիզում inter se[60] պտղաբեր են եղել ութ սերունդների ընթացքում։ Վերջերս հավատացնում էին, որ երկու այնքան տարբեր տեսակներ, ինչպես նապաստակը և ճագարը, եթե նրանց հարկադրեն բազմանալու, կառաջացնեն սերունդ, որը վերին աստիճանի պտղաբեր է ծնողական տեսակներից մեկն ու մեկի հետ խաչաձևվելու դեպքում։ Սովորական սագի և չինական սագի (Anser cygnoides), որոնք այնքան տարբեր տեսակներ են, որ նրանց սովորաբար դասում են տարբեր սեռերում, խաչաձևումից ստացված հիբրիդները հաճախ Անգլիայում սերունդ են տվել, երբ ենթարկվել են կրկնվող խաչաձևման երկու մաքուր ծնողական տեսակների հետ, իսկ մի անգամ էլ՝ նույնիսկ inter se։ Այս վերջինը հաջողվել է ստանալ մ-ր Էյտոնին,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 113] որը ստացել է երկու հիբրիդներ միևնույն ծնողներից, բայց տարբեր ձագահանումներից. իսկ այդ երկու թռչուններից նա ստացել է մի բնում ութից ոչ պակաս հիբրիդներ (մաքուր սագերի թոռներ)։ Սակայն Հնդկաստանում խաչաձևման միջոցով ստացված այդ սագերը պետք է անհամեմատ ավելի պտղաբեր լինեն, որովհետև, ինչպես ինձ հավատացնում են այնպիսի ամենաձեռնհաս դատավորներ, ինչպես մ-ր Բլայտն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 39] ու կապիտան Հյոտոն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 173] են, այդ երկրի զանազան վայրերում բուծվում են այդպիսի սագերի ամբողջ հոտեր, որոնք ստացված են խաչաձևման միջոցով. բայց քանի որ նրանք բուծվում են օգուտ ստանալու համար նույնիսկ այնտեղ, որտեղ չի հանդիպում ծնողական ոչ մեկ և ոչ էլ մյուս տեսակը, ուստի նրանք պետք է շատ կամ լիովին պտղաբեր լինեն։
Մեր ընտանի կենդանիների զանազան ցեղերը փոխադարձ խաչաձևման ժամանակ կատարելապես պտղաբեր են, իսկ մինչդեռ շատ դեպքերում նրանք առաջացել են երկու կամ երեք վայրի տեսակներից։ Այս փաստից մենք պետք է եզրակացնենք երկուսից մեկը՝ կամ նախասկզբնական ծնողական տեսակները հենց սկզբից արտադրել են միանգամ այն պտղաբեր հիբրիդներ, կամ ընտանեցման վիճակում հետո դուրս բերված հիբրիդները դարձել են բոլորովին պտղաբեր։ Այս վերջին երկընտրանքը, որ առաջին անգամ առաջարկել է Պալլասը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 104] շատ ավելի ճշմարտացի է թվում և, իսկապես, հազիվ թե կարող է որևէ կասկած հարուցել։ Այսպես, օրինակ, գրեթե անկասկածելի է, որ մեր շներն առաջացել են մի քանի վայրի ցեղից, բայց և այնպես, թերևս բացառությամբ Հարավային Ամերիկայի մի քանի տեղական ընտանի շների, նրանք բոլորն էլ միմյանց հետ զուգավորվելիս միանգամայն պտղաբեր են. սակայն անալոգիան ինձ հարկադրում է խիստ կասկածել այն բանում, որ սկզբնական տեսակները կարող էին ազատ կերպով զուգավորվել և տալ լիովին պտղաբեր հիբրիդներ։ Ճիշտ այդպես էլ ես վերջերս կարողացա լիովին համոզվել, որ եղջերավոր անասունների հնդկական սապատավորի և սովորական ցեղի խաչաձևման միջոցով ստացված սերունդը inter se միանգամայն պտղաբեր է. բայց հիմնվելով մեկ և մյուս ձևերի կարևոր ոսկրաբանական տարբերությունների վերաբերմամբ Ռյուտիմեյերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 40] կատարած դիտումների, ինչպես նաև նրանց արտաքին տեսքի, ձայնի, կազմության և այլ առանձնահատկությունների վերաբերմամբ մ-ր Բլայտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 39] կատարած դիտումների վրա, այդ երկու ձևերը պետք է դիտել որպես լավ, առանձին տեսակներ։ Նույնը կարելի է նշել նաև խոզերի երկու գլխավոր ցեղերի մասին։ Հետևաբար, մենք պետք է կամ հրաժարվենք այն համոզմունքից, որ տեսակները խաչաձևման ժամանակ ցուցաբերում են ընդհանուր անպտղություն, կամ կենդանիների անպտղության վրա նայել ոչ թե որպես մի անշնչելի հատկանիշի, այլ այնպիսին, որը կարելի է ընտանեցման միջոցով վերացնել։
Վերջապես, ուշադրության առնելով բոլոր վավերացված փաստերը բույսերի և կենդանիների խաչաձևման մասին, կարելի է եզրակացնել, որ ինչպես առաջին սերնդում, այնպես էլ հիբրիդների մոտ որոշ աստիճանի ամլություն չափազանց հաճախ է տեղի ունենում. սակայն մեր արդի գիտելիքների վիճակում չի կարելի համարել, որ նա անպայման տարածված է ամենուրեք։
ՕՐԵՆՔՆԵՐ, ՈՐՈՆՔ ՂԵԿԱՎԱՐՈՒՄ ԵՆ ԱՌԱՋԻՆ ԽԱՉԱՁԵՎՈՒՄՆԵՐԻ ԵՎ ՀԻԲՐԻԴՆԵՐԻ ԱՆՊՏՂՈՒԹՅՈՒՆԸ
Այժմ փոքր ինչ ավելի մանրամասնորեն քննարկենք այն օրենքները, որոնք ղեկավարում են առաջին խաչաձևումների և հիբրիդների անպտղությունը։ Գլխավորապես տեսնենք, թե այդ օրենքները չե՞ն նշում արդյոք այն, որ տեսակները դիտավորյալ կերպով օժտվել են այդ հատկությամբ, որպեսզի կանխեն նրանց խաչաձևումը և լիակատար խառնաշփոթումը իրար հետ։ Ստորև բերված եզրակացությունները փոխ են առնված գլխավորապես Գերտների[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] հիանալի աշխատությունից՝ բույսերի մոտ հիբրիդների առաջացման մասին։ Ես քիչ ջանք չեմ գործադրել, որպեսզի հավաստիանամ, թե ինչ աստիճանի նրանք կիրառելի են կենդանիների վերաբերմամբ, և նկատի առնելով մեր տեղեկությունների աղքատիկությունը կենդանական հիբրիդների առաջացման մասին, ես զարմացել եմ, երբ տեսել եմ թե որքան միևնույն օրենքներն ընդհանուր են բնության երկու թագավորությունների համար։
Արդեն նշվել է, որ ինչպես առաջին խաչաձևումների, այնպես էլ հիբրիդների պտղաբերության աստիճանը ներկայացնում է աստիճանաբար անցումներ' սկսած զրոյից մինչև լրիվ պտղաբերությունը։ Զարմանալի է, թե որքան հետաքրքիր եղանակներով կարելի է ցույց տալ այդ աստիճանականությունը, բայց ես այստեղ կարող եմ այդ փաստերը շարադրել միայն ամենաընդհանուր գծերով։ Եթե մի ընտանիքի պատկանող բույսից վերցրած ծաղկափոշին տեղափոխենք մի այլ ընտանիքի բույսի սպիի վրա, նա նույնքան քիչ ներգործություն է առաջացնում, ինչքան և անօրգանական փոշին։ Սկսելով պտղաբերության այդ բացարձակ զրոյից, երբ զանազան տեսակների ծաղկափոշին տեղափոխում ենք նույն սեռի որևէ տեսակի սպիի վրա, մենք նկատում ենք արտադրվող սերմերի քանակի լիակատար աստիճանականություն՝ ընդհուպ մինչև համարյա լրիվ կամ միանգամայն լրիվ պտղաբերություն, իսկ նորմայից շեղվող որոշ դեպքերում, ինչպես մենք տեսանք, մինչև անգամ լիառատ պտղաբերություն, այսինքն անցնում է նրանից, որ առաջ է բերում բույսի սեփական ծաղկափոշիով բեղմնավորվելը։ Հիբրիդների մեջ պատահում են նաև այնպիսիները, որոնք երբեք չեն արտադրել և հավանորեն չեն էլ կարողանում արտադրել ոչ մի հատ ծլունակ սերմ նույնիսկ մաքուր ծնողական ձևերի ծաղկափոշիով բեղմնավորելու դեպքում, բայց այդ դեպքերից մի քանիսում կարելի է պտղաբերության առաջին հետքերը հայտնաբերել այն բանում, որ մաքուր ծնողական տեսակներից մեկի ծաղկափոշին առաջ է բերում հիբրիդի ծաղկի ավելի վաղ թառամում, քան այդ կարող էր պատահել առանց նրա ազդեցության, իսկ ինչպես հայտնի է, ծաղկի ավելի վաղ թառամումը ծառայում է որպես սկսվող բեղմնավորման նշան։ Անպտղաբերության այդ ծայրահեղ աստիճանից հեռանալով՝ մենք հանդիպում ենք այլ ինքնաբեղմնավորվող հիբրիդների, որոնք արտադրում են ավելի ու ավելի մեծ թվով սերմեր, հասնում են մինչև լիակատար պտղաբերության։
Այն հիբրիդները, որոնք ստացվում են դժվար խաչաձևվող և հազվագյուտ դեպքում սերունդ արտադրող երկու տեսակներից, սովորաբար չափազանց անպտղաբեր են։ Սակայն առաջին խաչաձևման դժվարության և սրանից առաջացող հիբրիդների անպտղության (երկու կատեգորիայի փաստեր, որ սովորաբար խառնվում են) միջև եղած զուգահեռությունը երբեք խիստ չէ։ Կան շատ դեպքեր, երբ երկու մաքուր տեսակներ, օրինակ, Verbascum սեռի մեջ, կարող են չափազանց հեշտությամբ խաչաձևվել և տալ բազմաթիվ հիբրիդային ժառանգներ, իսկ մինչդեռ հիբրիդներն իրենք չափազանց անպտղաբեր են։ Մյուս կողմից կան տեսակներ, որոնց հաջողվում է խաչաձևել շատ հազվագյուտ դեպքում կամ ծայրահեղ դժվարությամբ, բայց նրանց հիբրիդները, եթե միայն նրանք առաջացել են, խիստ պտղաբեր են։ Մինչև անգամ միևնույն սեռի սահմաններում, օրինակ Dianthus-ի մոտ, հանդիպում են այս երկու հակադիր դեպքերը։
Ինչպես առաջին խաչաձևումների, այնպես էլ հիբրիդների պտղաբերությունը ավելի հեշտությամբ է տուժում անբարենպաստ պայմանների ներգործությունից, քան մաքուր տեսակների պտղաբերությունը։ Բայց առաջին խաչաձևումների պտղաբերությունը նույնպես բնածին կերպով փոփոխական է, որովհետև նա իր աստիճանով միշտ միանման չի լինում. երբ միևնույն երկու տեսակները խաչաձևվում են միանման հանգամանքներում, նա մասամբ կախված է այն անհատների, կառուցվածքից, որոնք պատահաբար ընտրվել են փորձի համար։ նույնն է նկատվում նաև հիբրիդների մոտ. նրանց պտղաբերության աստիճանը հաճախ խիստ տարբեր է լինում զանազան անհատների մոտ, որոնք աճել են միևնույն տուփիկի սերմերից և ենթարկվել են միևնույն պայմանների ազդեցությանը։
«Սիստեմատիկական ազգակցություն» արտահայտությամբ հասկանում են տեսակների միջև եղած ընդհանուր նմանությունը կառուցվածքով և կերտվածքով։ Բայց առաջին խաչաձևումների և նրանցից առաջացած հիբրիդների պտղաբերությունը մեծ չափով կախված է նրանց սիստեմատիկական ազգակցությունից։ Դա պարզ երևում է նրանից, որ հիբրիդներ երբեք չեն ստացվել երկու տեսակներից, որոնք սիստեմատիկների կողմից դասվում են տարբեր ընտանիքներում, և որ մյուս կողմից, շատ մերձավոր տեսակները սովորաբար հեշտությամբ խաչաձևվում են միմյանց հետ։ Բայց սիստեմատիկական ազգակցության և խաչաձևման հեշտության միջև եղած համապատասխանությունը բնավ խիստ չէ։ Կարելի է բերել բազմաթիվ օրինակներ, երբ շատ մերձավոր տեսակներ հրաժարվում են միմյանց հետ խաչաձևվելուց կամ խաչաձևվում են ծայրահեղ դժվարությամբ, իսկ մյուս կողմից, կարելի է բերել չափազանց հեշտությամբ խաչաձևվող տարբեր տեսակների խիստ շատ օրինակներ։ Միևնույն ընտանիքում կարող է լինել այնպիսի սեռ, ինչպես Dianthus-ն է, որի մեջ մտնող շատ տեսակներ չափից դուրս հեշտությամբ են խաչաձևվում, և մի այլ սեռ, ինչպես Silene-ն է, որի մեջ մտնող նույնիսկ ծայրահեղ մերձավոր տեսակներից, չնայած գործադրված ամենահամառ ջանքերին, ոչ մի հիբրիդ չի հաջողվել ստանալ։ Նույն տարբերությանը մենք հանդիպում ենք մինչև անգամ միևնույն սեռի սահմաններում։ Օրինակ, Nicotiana-ի շատ տեսակներ խաչաձևվում են շատ ավելի հաճախ, քան որևէ այլ սեռի տեսակներ, այնինչ Գերտները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] գտել է, որ N. acuminata տեսակը, որն առանձնապես աչքի չի ընկնում մյուսների մեջ, համառորեն հրաժարվել է բեղմնավորել կամ բեղմնավորվել Nicotiana-ի ութից ոչ պակաս այլ տեսակների հետ խաչաձևումների ժամանակ։ Կարելի էր էլի շատ համանման օրինակներ բերել։
Ոչ ոքին չի հաջողվել պարզ կերպով ցույց տալ, թե նկատելի հատկանիշներից որևէ մեկում ինչպիսի տարբերությունը և նրա ինչ աստիճանը բավական են երկու տեսակների միջև խաչաձևումը կանխելու համար։ Կարելի է ցույց տալ, որ կերտվածքով ու ընդհանուր տեսքով միմյանցից չափազանց տարբեր և ծաղկի յուրաքանչյուր մասում, նույնիսկ ծաղկափոշու, պտղի և շաքիլների մեջ խիստ արտահայտված տարբերություններ հանդես բերող բույսերը կարող են խաչաձևվել։ Միամյա և բազմամյա բույսերը, տերևաթափ ունեցող ծառերը և մշտադալար բույսերը, որոնք բնակվում են զանազան վայրերում և հարմարված են շատ տարբեր կլիմաների, հաճախ և հեշտ կերպով կարող են խաչաձևվել միմյանց հետ։
Երկու տեսակների միջև փոխադարձ խաչաձևում ասելով ես հասկանում եմ այն դեպքը, երբ, օրինակ, սկզբում էգ էշը խաչաձևվում է հովատակի հետ, իսկ հետո՝ զամբիկը էշի հետ. այդ դեպքում կարելի է ասել, որ այդ երկու տեսակները փոխադարձաբար խաչաձևվել են։ Հաճախ մեծագույն տարբերություն է նկատվում փոխադարձ խաչաձևումներ ստանալու հեշտության մեջ։ Այդպիսի դեպքերը չափազանց կարևոր են, որովհետև նրանք ապացուցում են, որ երկու որևէ տեսակների խաչաձևվելու ընդունակությունը հաճախ բոլորովին կախում չունի նրանց սիստեմատիկական ազգակցությունից, այսինքն կառուցվածքում կամ կերտվածքում եղած որևէ տարբերությունից, բացառությամբ նրանց վերարտադրողական սիստեմում եղած տարբերության։ Միևնույն տեսակների միջև փոխադարձ խաչաձևումների ժամանակ ստացված արդյունքների մեջ եղած տարբերությունը արդեն վաղուց նկատել է Կյոլռեյտերը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] Բերենք մի օրինակ. Mirabilis jalapa-ն հեշտությամբ կարող է բեղմնավորվել M. longiflora-ի ծաղկափոշով և այդ ձևով ստացվող հիբրիդները բավականաչափ պտղաբեր են. բայց Կյոլռեյտերը ութ տարվա ընթացքում շարունակ երկու հարյուրից ավելի փորձեր է կատարել M. longiflora-ն բեղմնավորել M. jalapa-ի ծաղկափոշով և բոլորովին ապարդյուն։ Կարելի է բերել մի քանի այլ զարմանալի դեպքեր։ Տյուրեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 185] նույն փաստը նկատել է ծովային ջրիմուռների՝ ֆուկուսների մոտ։ Բացի դրանից, Գերտները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] գտել է, որ փոխադարձ խաչաձևումներ ստանալու հեշտության մեջ եղած տարբերությունը, ճիշտ է, ոչ այնքան խիստ աստիճանի, չափազանց ընդհանուր երևույթ է։ Նա այդ նկատել է նույնիսկ մոտ ազգակից ձևերի մոտ (օրինակ, Matthiola annua-ի և M. glabra-ի մոտ), որոնց շատ բուսաբաններ համարում են տարատեսակներ։ Ուշագրավ է նաև այն փաստը, որ թեև փոխադարձ խաչաձևումներից առաջացած հիբրիդները, իհարկե, ներկայացնում են միևնույն երկու տեսակների զուգակցում (ըստ որում մի տեսակը սկզբում հոր դեր է կատարել, իսկ հետո՝ մոր) և հաճախ մեկը մյուսից տարբերվում են արտաքին հատկանիշներով, սակայն նրանց պտղաբերությունը սովորաբար տարբերվում է աննշան, իսկ երբեմն էլ մեծ չափով։
Գերտների[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] դիտումներից կարելի է բերել մի քանի այլ յուրահատուկ օրենքներ․ այսպես, օրինակ, մի քանի տեսակներ այլ տեսակների հետ խաչաձևվելու հիանալի ընդունակություն ունեն, իսկ նույն սեռի այլ տեսակներ աչքի են ընկնում իրենց արտաքին առանձնահատկությունները հիբրիդային սերնդին փոխանցելու ընդունակությամբ։ Բայց այդ երկու ընդունակությունների միջև անհրաժեշտ զուգադիպություն գոյություն չունի։ Որոշ հիբրիդներ փոխանակ ներկայացնելու, ինչպես այդ սովորաբար լինում է, միջակա հատկանիշներ երկու ծնողների միջև, միշտ ցուցաբերում են նմանություն ծնողներից մեկն ու մեկին և այդպիսի հիբրիդները, չնայած մաքուր ծնողական տեսակի հետ նրանց ունեցած խիստ նմանությանը, հազվադեպ բացառությամբ լինում են չափազանց անպտղաբեր։ Ճիշտ այդպես էլ այն հիբրիդների մեջ, որոնք իրենց կառուցվածքով միջին տեղ են գրավում ծնողների միջև, երբեմն ծնվում են բացառիկ, նորմայից շեղվող անհատներ, օժտված մոտ նմանությամբ մաքուր ծնողական ձևերից մեկի հետ, և այդ հիբրիդները համարյա միշտ չափազանց անպտղաբեր են նույնիսկ այն դեպքում, երբ մյուս հիբրիդները, որ աճեցված են հենց միևնույն տուփիկից վերցված սերմերից, դրսևորում են մեծ պտղաբերություն։ Այս փաստերը ցույց են տալիս, թե հիբրիդի պտղաբերությունը որքան կարող է անկախ լինել ծնողներից մեկի հետ նրա ունեցած արտաքին նմանությունից։
Քննարկելով վերևում բերված զանազան օրենքները, որոնք որոշում են առաջին խաչաձևումների և հիբրիդների պտղաբերությունը, մենք տեսնում ենք հետևյալը. երբ խաչաձևվում են այնպիսի ձևեր, որոնք պետք է համարվեն որպես լավ, առանձին տեսակներ, նրանց պտղաբերությունը ներկայացնում է աստիճանական անցումներ՝ սկսած զրոյից մինչև լրիվ պտղաբերությունը կամ նույնիսկ որոշ պայմաններում մինչև լիառատ պտղաբերությունը։ Նրանց պտղաբերությունը ոչ միայն չափազանց զգայուն է բարենպաստ ու անբարենպաստ պայմանների ազդեցության նկատմամբ, այլև ունի բնածին փոփոխականություն. նա իր աստիճանով երբեք միշտ միանման չի լինում առաջին խաչաձևումներում և նրանցից ծագում առած հիբրիդների մոտ, հիբրիդների պտղաբերությունը առնչություն չունի ծնողներից մեկին արտաքին հատկանիշներով նրանց նմանվելու աստիճանի հետ, և վերջապես, երկու տեսակների միջև տեղի ունեցող առաջին խաչաձևման հեշտությունը միշտ չէ, որ կախված է նրանց սիստեմատիկական ազգակցությունից կամ միմյանց հետ ունեցած նմանության աստիճանից։ Այս վերջին դրույթը պարզ կերպով ապացուցվում է աջն տարբերությամբ, որ գոյություն ունի միևնույն տեսակների միջև փոխադարձ խաչաձևումներից ստացված արդյունքների մեջ։ Իրոք, նայած այն բանին, թե արդյոք մեկ կամ մյուս տեսակն է կատարում հոր կամ մոր ղերը, սովորաբար նկատվում է որոշ, երբեմն էլ շատ մեծ տարբերություն այն հեշտության մեջ, որով տեղի է ունենում խաչաձևումը։ Բացի դրանից, փոխադարձ խաչաձևումներից առաջացած հիբրիդները հաճախ տարբերվում են պտղաբերությամբ։
Արդյո՞ք այս բարդ և յուրահատուկ օրենքները ցույց են տալիս, որ տեսակներն օժտված են անպտղաբերությամբ, որպեսզի կանխեն նրանց խառնածնությունը բնության մեջ։ Չեմ կարծում, որովհետև ինչո՞ւ տարբեր տեսակների պտղաբերությունը իր աստիճանով այնքան խիստ բազմազան պիտի լիներ, մինչդեռ, պետք է կարծել, միանման կարևոր է, որ նրանք բոլորն էլ պահպանվեն խառնածությունից։ Ինչո՞ւ անպտղության աստիճանը պետք է բնածին կերպով փոփոխական լիներ միևնույն տեսակի անհատների մոտ։ Ինչո՞ւ որոշ տեսակներ պետք է հեշտությամբ խաչաձևվեն և միևնույն ժամանակ առաջացնեն խիստ անպտղաբեր հիբրիդներ, իսկ մյուս տեսակները պետք է խաչաձևվեն չափազանց դժվարությամբ, բայց և այնպես առաջացնեն բավական պտղաբեր հիբրիդներ։ Ինչո՞ւ հենց միևնույն երկու տեսակների միջև փոխադարձ խաչաձևումների արդյունքը պետք է հաճախ նույնպես ներկայացներ մեծ տարբերություն։ Ինչո՞ւ, կարելի է մինչև անգամ հարցնել, բնությունը թույլ է տվել հենց հիբրիդների առաջացումը։ Օժտել տեսակները հիբրիդներ առաջացնելու ընդունակությամբ և հետո դադարեցնել նրանց հետագա բազմացումը անպտղության տարբեր աստիճանների միջոցով, որոնք սերտ առնչություն չունեն նրանց ծնողների առաջին խաչաձևման հեշտության հետ, դա հարցի տարօրինակ լուծում է։
Մյուս կողմից, նախորդ օրենքները և փաստերը, ինչպես ինձ թվում է, պարզ կերպով ցույց են տալիս, որ ինչպես առաջին խաչաձևումների, այնպես էլ հիբրիդների անպտղությունը մի երևույթ է, որը ուղեկցում է կամ կախման մեջ է գտնվում նրանց վերարտադրողական սիստեմներում եղած անհայտ առանձնահատկություններից. այդ տարբերություններն այնքան յուրահատուկ ու սահմանափակ են իրենց բնույթով, որ հաճախ միևնույն երկու տեսակների փոխադարձ խաչաձևումների ժամանակ մեկի արական սեռական տարրը ազատ կերպով ներգործում է մյուսի իգական սեռական տարրի վրա, բայց ոչ ընդհակառակը։ Ավելորդ չի լինի փոքր ինչ ավելի լրիվ պարզաբանել մի օրինակով, թե ես ինչ եմ հասկանում, ասելով, որ անպտղաբերությունը մյուս տարբերություններին ուղեկցող մի ինչ-որ բան է, այլ ոչ թե ժառանգված հատկություն։ Քանի որ բույսերի մեկը մյուսի վրա պատվաստվելու ընդունակությունը կարևոր չէ բնական դրության մեջ նրանց բարեկեցության համար, ապա ոչ ոք, ես կարծում եմ, չի ենթադրի, որ այդ ընդունակությունը առանձնահատուկ ժառանգված հատկություն է, և կհամաձայնի, որ նա երկու բույսերի աճման օրենքների մեջ եղած տարբերություններին ուղեկցող հետևանքն է։ Թե ինչու մի ծառ չի պատվաստվում մյուսի վրա, մենք երբեմն կարող ենք պատճառը գտնել նրանց աճեցողության արագության, բնափայտի ամրության, հյութի շարժման ժամանակաշրջանի և նրա հատկությունների և այլ տարբերությունների մեջ, բայց շատ դեպքերում մենք չենք կարող ոչ մի պատճառ նշել։ Երկու բույսերի մեծությունների տարբերությունը կամ այն տարբերությունը, որ նրանցից մեկը, դիցուք, ծառատեսակ է, իսկ մյուսը խոտանման, մեկը՝ մշտադալար է, իսկ մյուսը տերևաթափ ունեցող և որ նրանք հարմարված են շատ տարբեր կլիմաների, միշտ չի կարող խանգարել երկու բույսերի միմյանց հետ պատվաստվելուն։ Ինչպես խաչաձևման, այնպես էլ պատվաստման դեպքում բույսերի ընկալունակությունը սահմանափակված է սիստեմատիկական ազգակցությամբ, որովհետև դեռ ոչ ոքի չի հաջողվել բոլորովին տարբեր ընտանիքների պատկանող ծառեր պատվաստել մեկը մյուսի վրա։ Իսկ մյուս կողմից, շատ մերձավոր տեսակներ և միևնույն տեսակների տարատեսակները սովորաբար, թեև ոչ միշտ, հեշտությամբ պատվաստվում են իրար հետ։ Սակայն, ինչպես որ խաչաձևման ժամանակ, այդ ընկալունակությունը ամենևին անպայմանորեն չի որոշվում սիստեմատիկական ազգակցությամբ։ Չնայած նրան, որ միևնույն ընտանիքի զանազան շատ սեռեր պատվաստվում են մեկը մյուսի վրա, այլ դեպքերում միևնույն սեռի տեսակները հրաժարվում են իրար հետ միանալուց։ Տանձենին անհամեմատ ավելի հեշտությամբ պատվաստվում է սերկևլենու վրա, որը պատկանում է այլ սեռի, քան խնձորենու վրա, որը տանձենու հետ միասին պատկանում է միևնույն սեռին։[61] Նույնիսկ նրա զանազան տարատեսակները միանման հեշտությամբ չեն պատվաստվում սերկևլենու վրա. նույնը նշվում է ծիրանենու և դեղձենու զանազան տարատեսակները սալորենու մի քանի տարատեսակների վրա պատվաստելու վերաբերմամբ։
Ինչպես մենք տեսանք, Գերտները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] գտել է, որ երկու տեսակների պատկանող տարբեր անհատների խաչաձևման ընդունակության մեջ նկատվում է բնածին տարբերություն։ Սաժերեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 186] կարծիքով նույնը պատահում է նան զանազան անհատներ մեկը մյուսի վրա պատվաստելու դեպքում։ Մենք տեսանք, որ փոխադարձ խաչաձևման ժամանակ այն հեշտությունը, որով նա ձեռք է բերվում, հաճախ ամենևին միանման չէ. նույնն է նկատվում նաև պատվաստման վերաբերմամբ։ Օրինակ, սովորական կոկռոշենին չի կարելի պատվաստել հաղարջենու վրա, մինչդեռ հաղարջենին, թեպետև դժվարությամբ, պատվաստվում է կոկռոշենու վրա։
Մենք տեսանք, որ անկատարյալ վերարտադրողական օրգաններ ունեցող հիբրիդների անպտղությունը և կատարյալ վերարտադրողական սիստեմով օժտված երկու մաքուր տեսակների խաչաձևման դժվարությունը երկու տարբեր բաներ են, սակայն այս երկու կատեգորիայի փաստերը զգալի չափով զուգահեռ են։ Ինչ որ նման բան նկատվում է նաև պատվաստման վերաբերմամբ։ Այսպես, օրինակ, Տուենը (Thoin)[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 187] գտել է, որ Robinia-ի[62] երեք տեսակները, որոնք առատորեն սերմեր էին տալիս իրենց սեփական արմատի վրա և առանց առանձին դժվարության պատվաստվում էին չորրորդ տեսակի վրա, այդպիսի պատվաստումից հետո դառնում էին բոլորովին անպտղաբեր։ Մյուս կողմից, Sorbus սեռի որոշ տեսակներ, որոնք պատվաստվել են այլ տեսակների վրա, տալիս էին երկու անգամ ավելի պտուղներ, քան սեփական արմատի վրա։ Այս վերջին փաստը մեզ հիշեցնում է արտասովոր դեպքերը Hippeastrum-ի, Passiflora-ի և այլ բույսերի մոտ, որոնք այլ տեսակի ծաղկափոշիով բեղմնավորելիս անհամեմատ ավելի առատ սերմեր են տալիս, քան միևնույն բույսից վերցված ծաղկափոշիով բեղմնավորելու դեպքում։
Այսպիսով, մենք տեսնում ենք, որ թեև պարզորոշ և խոշոր տարբերություն կա պատվաստման միջոցով կատարվող պարզ սերտաճման և վերարտադրության գործողությամբ արական և իգական տարրերի միավորման միջև, սակայն որոշ կոպիտ զուգահեռություն է նկատվում զանազան տեսակների պատվաստման ու խաչաձևման հետևանքներում։ Եվ ինչպես որ մի ծառը մյուսի վրա պատվաստման մեծ կամ փոքր հեշտությունը որոշող հետաքրքրական ու բարդ օրենքները մենք պետք է համարենք բույսերի վեգետատիվ սիստեմում եղած մեզ անհայտ առանձնահատկությունների կողմնակի հետևանքներ, ճիշտ այդպես էլ, իմ կարծիքով, մենք պետք է առաջին խաչաձևումների հեշտությունը որոշող ավելի ևս բարդ օրենքները դիտենք որպես վերարտադրողական սիստեմի մեզ անհայտ առանձնահատկությունների կողմնակի հետևանքներ։ Այդ առանձնահատկությունները երկու դեպքում էլ, ինչպես որ պետք էր սպասել, որոշ չափով կապված են սիստեմատիկական ազգակցության հետ՝ մի տերմին, որի միջոցով աշխատում են արտահայտել օրգանական էակների միմյանց նմանությունը և ոչ նմանությունը։ Այդ փաստերը, ինչպես ինձ թվում է, ոչ մի չափով չեն ցույց տալիս, որ զանազան տեսակների միջև պատվաստման կամ խաչաձևման ավելի կամ պակաս դժվարությունը մի հատուկ առանձնահատկություն լիներ, թեև խաչաձևման դեպքում այդ դժվարությունը նույնքան կարևոր է տեսակային ձևերի պահպանման ու կայունության համար, որքան որ պատվաստման դեպքում այն կարևոր չէ նրանց բարեկեցության համար։
ԱՌԱՋԻՆ ԽԱՉԱՁԵՎՈԻՄՆԵՐԻ ԵՎ ՀԻԲՐԻԴՆԵՐԻ ԱՆՊՏՂՈՒԹՅԱՆ ԾԱԳՈՒՄԸ ԵՎ ՊԱՏՃԱՌՆԵՐԸ
Մի ժամանակ ինձ, ինչպես և ուրիշներին, հավանական էր թվում, որ առաջին խաչաձևումների և հիբրիդների անպտղությունը կարող էր դանդաղորեն ձեռք բերվել պտղաբերության թեթևակի նվազող աստիճանների բնական ընտրության միջոցով, որոնք ամեն մի այլ փոփոխության նման իրենք իրենց երևան են գալիս մի տարատեսակի առանձին անհատների մոտ, երբ նրանք խաչաձևվում են մի այլ տարատեսակի անհատների հետ։ Եվ իրոք, երկու տարատեսակների կամ սկզբնավորվող տեսակների համար հավանորեն ձեռնտու կլիներ, եթե նրանց կարելի լիներ հեռու պահել խառնածնությունից, ձեռնտու կլիներ նույն պատճառով, ինչ պատճառով երկու տարատեսակներ միաժամանակ ընտրող մարդը անհրաժեշտորեն պետք է նրանց պահի մեկը մյուսից առանձին։ Նախ, կարելի է նկատել, որ տարբեր վայրերում ապրող տեսակները խաչաձևվելու դեպքում հաճախ անպտղաբեր են. մինչդեռ պարզ է, որ այդպիսի մեկը մյուսից հեռավոր տեսակներին ոչ մի օգուտ չէր բերի այն, եթե նրանք փոխադարձաբար անպտղաբեր դառնային, և, հետևաբար, այդ չէր կարող տեղի ունենալ բնական ընտրության միջոցով։ Սակայն թերևս կարելի էր բերել այն փաստարկը, որ եթե տեսակն անպտղաբեր է դարձել իր որևէ հայրենակցի հետ խաչաձևվելիս, ապա այլ տեսակների հետ անպտղաբերությունը կլինի որպես մի անհրաժեշտ հետևանք։ Երկրորդ, բնական ընտրության տեսությունը գրեթե նույն չափով, ինչպես որ հատուկ ստեղծագործության տեսությունը, հակասում է փոխադարձ խաչաձևումների ժամանակ դիտվող փաստերին, այն է՝ անհասկանալի է թվում, թե ինչպես մի ձևի արական տարրը կարող էր միանգամայն անզոր դառնալ երկրորդ ձևի վերաբերմամբ, այնինչ միևնույն ժամանակ այդ երկրորդ ձևի արական տարրը հեշտությամբ կարողանում է բեղմնավորել առաջին ձևին, և իրոք, վերարտադրողական սիստեմում եղած այդպիսի յուրահատուկ վիճակը հազիվ թե կարող էր օգտակար լինել երկու տեսակներից որևէ մեկի համար։
Այն հավանականությունը, որ բնական ընտրությունն է առաջացրել նաև անպտղությունը միջտեսակային խաչաձևման դեպքում, շատ մեծ դժվարությունների է հանդիպում այն փաստի մեջ, որ գոյություն ունեն բազմաթիվ հաջորդական աստիճաններ՝ սկսած թեթևակի թուլացած պտղաբերությունից մինչև կատարյալ անպտղաբերությունը։ Կարելի է ընդունել, որ սկզբնավորվող տեսակին ձեռնտու կլիներ, եթե նա որոշ թույլ չավ։ով անպտուղ դառնար իր ծնողական ձևերի կամ որևէ այլ տարատեսակի հետ խաչաձևվելիս, որովհետև այդ կերպով ավելի քիչ կառաջանար խառնածին վատթարացած սերունդ, որը կարող էր իր արյունը խառնել նոր առաջացող տեսակի հետ։ Բայց նա, ով նեղություն հանձն կառնի խորհելու այն աստիճանների մասին, որոնցով պետք է անցներ անպտղաբերության առաջին աստիճանը, որպեսզի բնական ընտրության ճանապարհով հասներ մինչև այն բարձր աստիճանին, որն ընդհանուր է այնքան շատ տեսակների և նույնիսկ ընդհանրապես տեսակների համար, որոնք դիֆերենցման միջոցով բարձրացել են մինչև սեռերի կամ ընտանիքների աստիճանի վրա, նա, ով այդ նեղությունը հանձն կառնի, կգա այն եզրակացության, որ հարցը արտասովոր չափով բարդ է։ Հասուն մտորմունքից հետո ինձ թվում է, որ այդ չէր կարող տեղի ունենալ բնական ընտրության միջոցով։ Վերցնենք այն դեպքը, երբ երկու տեսակ խաչաձևվելիս առաջացնում են բազմաթիվ և անպտղաբեր սերունդ։ Այդ դեպքում ի՞նչը կարող էր նպաստել այդ անհատների գերապրելուն, որոնք պատահմամբ օժտված են եղել փոքր ինչ ավելի մեծ աստիճանի փոխադարձ անպտղաբերությամբ և որոնք այդպիսով մի փոքրիկ աստիճանով կմոտենային կատարյալ անպտղաբերության։ Մինչդեռ, եթե բնական ընտրության տեսությունը այստեղ կիրառելի է, ապա այդ ուղղությամբ քայլեր պետք է միշտ հանդիպեն շատ տեսակների մոտ, որովհետև նրանցից շատ տեսակներ փոխադարձաբար միանգամայն անպտղաբեր են։ Անպտղաբեր, անսեռ միջատների համար մենք հիմք ունենք ենթադրելու, որ նրանց կառուցվածքի և պտղաբերության մեջ կատարված ձևափոխությունները դանդաղորեն կուտակվել են բնական ընտրության միջոցով այն բանի շնորհիվ, որ դրա հետ միասին այն համայնքը, որին պատկանում են այդ միջատները, անուղղակի կերպով առավելություն է ստացել նույն տեսակի մյուս համայնքների հանդեպ, բայց եթե սոցիալական համայնքի չպատկանող մի անհատական կենդանի թեթևակի անպտղաբեր դառնար որևէ այլ տարատեսակի հետ խաչաձևվելու դեպքում, նա ինքը դրանից ոչ մի առավելություն չէր ստանա և չէր էլ կարողանա անուղղակի կերպով որևէ առավելություն տալ նույն տարատեսակին պատկանող մյուս անհատներին և այդպիսով օժանդակել նրանց պահպանմանը։
Բայց ավելորդ է այս հարցն իր մանրամասնություններով քննարկել, որովհետև բույսերի վերաբերմամբ մենք ունենք համոզիչ ապացույց այն մասին, որ խաչաձևվող տեսակների անպտղաբերությունը պետք է վերագրել մի որևէ սկզբունքի, որը բոլորովին անկախ է բնական ընտրությունից։ Ինչպես Գերտները,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] այնպես էլ Կյոլռեյտերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] ապացուցել են, որ բազմաթիվ տեսակներ պարփակող սեռերում կարելի է կազմել հաջորդական շարք՝ սկսելով այն տեսակներից, որոնք խաչաձևվելիս տալիս են հետզհետե ավելի ու ավելի քիչ թվով սերմեր, և վերջացրած այն տեսակներով, որոնք երբեք ոչ մի հատ սերմ չեն տալիս, բայց և այնպես ենթարկվում են մի քանի այլ տեսակների ծաղկափոշու ազդեցությանը, ինչպես այդ երևում է սերմնարանի ուռչելուց։ Ակներև է, որ այստեղ անկարելի է ընտրել ավելի անպտղաբեր անհատներ, որոնք արդեն դադարել են սերմեր տալուց, այնպես որ անպտղաբերության այդ ծայրահեղ աստիճանը, որի առկայությամբ միմիայն սերմնարանն է կրում ծաղկափոշու ազդեցությունը, չէր կարող ձեռք բերվել ընտրության միջոցով, իսկ այն հանգամանքից, որ անպտղաբերության տարբեր աստիճանները որոշող օրենքները այնքան միանման են կենդանական ու բուսական թագավորություններում, մենք կարող ենք եզրակացնել, որ պատճառն էլ, նա ինչ էլ ուզում է լինի, բոլոր դեպքերում միանման է կամ գրեթե միանման է։
Այժմ մենք փոքր ինչ ավելի ուշադիր կերպով քննարկենք այն տարբերությունների հավանական բնույթը, որոնք հարուցում են առաջին խաչաձևումների և հիբրիդների անպտղաբերություն։ Առաջին խաչաձևումների դեպքում զուգավորվելու և սերունդ ստանալու ավելի մեծ կամ փոքր դժվարությունը ակներևորեն կախված է մի քանի տարբեր պատճառներից։ Երբեմն արական տարրի համար երևի ֆիզիկապես անկարելի է հասնել ձվիկին, ինչպես, օրինակ, այն բույսի մեջ, որի վարսանդն այնքան երկար է, որ ծաղկափոշու խողովակները չեն կարողանում թափանցել սերմնարանի խոռոչը։ Նկատվել է նաև, որ երբ մի տեսակի ծաղկափոշին ընկնում է նրա հետ հեռավոր ազգակցություն ունեցող մի այլ տեսակի սպիի վրա, այդ դեպքում թեև ծաղկափոշին ծլում է, բայց նրա խողովակները սպիի մակերեսի միջով ներս չեն թափանցում։ Այնուհետև, արական տարրը կարող է հասնել իգական տարրին, բայց անընդունակ է լինում առաջացնելու սաղմի զարգացումը, որը, ըստ երևույթին, տեղի է ունեցել ֆուկուսների վրա՝ Տյուրեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 185] կատարած մի քանի փորձերի ժամանակ։ Այս փաստերը նույնքան քիչ բացատրելի են, որքան այն պատճառը, թե ինչու որոշ ծառատեսակներ չեն կարող պատվաստվել մյուսների վրա։ Վերջապես սաղմը կարող է գոյանալ, բայց հետո՝ ոչնչանալ վաղ ստադիայում։ Այս վերջին երկընտրանքի վրա բավականաչափ ուշադրություն չի դարձվել, բայց ես կարծում եմ, հիմնվելով այն դիտումների վրա, որ ինձ հաղորդել է մ-ր Հյուետը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 188] որը մեծ փորձառություն է ունեցել փասիաններին ընտանի թռչունների հետ խաչաձևելու գործում, որ սաղմի վաղաժամ մահը շատ հաճախ առաջին խաչաձևումների անպղաբերության պատճառ է ծառայում։ Վերջերս մ-ր Սոլտերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 189] հաղորդել է մոտավորապես 500 ձվի վրա կատարած իր հետազոտությունների արդյունքը. ձվերն ստացված էին Gallus սեռի երեք տեսակների և նրանց հիբրիդների միջև տեղի ունեցած զանազան խաչաձևումներից։ Այդ ձվերի մեծ մասը բեղմնավորված է եղել և բեղմնավորված ձվերի մեծամասնության մեջ սաղմերը կամ մասամբ զարգացել են և հետո ոչնչացել, կամ հասել են գրեթե լիակատար հասունության, բայց մատղաշ ձագերը անընդունակ են եղել կեղևը ծակելու։ Լույս աշխարհ եկած ճուտիկներից ավելի քան 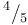 -ը ոչնչացել է առաջին օրերի կամ ամենաուշը մի շաբաթվա ընթացքում «առանց որևէ ակնհայտ պատճառի, ըստ երևույթին պարզապես կենսունակություն չունենալու պատճառով», այնպես որ 500 ձվից ստացվել են միայն 12 ճուտեր։ Բույսերի խաչաձևումների հետևանք հանդիսացող սաղմերը հավանորեն հաճախ ոչնչանում են նույն ձևով։ Համենայն դեպս հայտնի է, որ խիստ տարբեր տեսակներից ստացված հիբրիդները երբեմն շատ նվազ են, ցածրահասակ և ոչնչանում են վաղ հասակում։ Մաքս Վիխուրան[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 190] հաղորդել է մի քանի դրա նման զարմանալի դեպքեր հիբրիդային ուռենիների վրա իր կատարած դիտումներից։ Պետք է այստեղ նշել, որ կուսածնության որոշ դեպքերում մետաքսագործ-թիթեռի չբեղմնավորված ձվերի մեջ սաղմերն անցնում են զարգացման վաղ ստադիաները, հետո ոչնչանում են, նման այն սաղմերին, որոնք առաջացել են տարբեր տեսակների միջև տեղի ունեցած խաչաձևման միջոցով։ Քանի դեռ ծանոթ չէի այս փաստերի հետ, ես դժկամորեն էի հավատում, որ հիբրիդային սաղմերի վաղաժամ մահը հաճախակի երևույթ է, որովհետև հիբրիդները, քանի որ արդեն ծնվել են, սովորաբար լինում են առողջ և երկարակյաց, ինչպես այդ մենք տեսնում ենք սովորական ջորու օրինակի վրա։ Սակայն հիբրիդները մինչև ծնվելը և ծնվելուց հետո գտնվում են տարբեր պայմաններում։ Ծնվելով և ապրելով այն երկրում, որտեղ ապրում են երկու ծնողները, նրանք սովորաբար ընկնում են հարմար կենսապայմանների մեջ։ Սակայն հիբրիդը միայն կիսով չափ ունենում է մոր բնույթն ու կերտվածքը, ուստի մինչև ծնվելը, քանի դեռ նա սնվում է մոր արգանդում, կամ մոր արտադրած ձվի կամ սերմի մեջ, նա կարող է գտնվել որոշ տեսակետներից անհարմար պայմաններում, և, հետևաբար, վաղ հասակում ոչնչանալու վտանգին է ենթարկվում, մանավանդ որ բոլոր շատ մատղաշ օրգանիզմները արտասովոր կերպով զգայուն են դեպի գոյության վնասակար կամ անբնական պայմանները, վերջ ի վերջո ամենից ավելի հավանական է, որ պատճառը գտնվում է բեղմնավորման սկզբնական գործողության որևէ անկատարելության մեջ, որից առաջանում է սաղմի անկատար զարգացումը, և ոչ թե այն պայմանների մեջ, որոնք նրա վրա ներգործում են հետագայում։
-ը ոչնչացել է առաջին օրերի կամ ամենաուշը մի շաբաթվա ընթացքում «առանց որևէ ակնհայտ պատճառի, ըստ երևույթին պարզապես կենսունակություն չունենալու պատճառով», այնպես որ 500 ձվից ստացվել են միայն 12 ճուտեր։ Բույսերի խաչաձևումների հետևանք հանդիսացող սաղմերը հավանորեն հաճախ ոչնչանում են նույն ձևով։ Համենայն դեպս հայտնի է, որ խիստ տարբեր տեսակներից ստացված հիբրիդները երբեմն շատ նվազ են, ցածրահասակ և ոչնչանում են վաղ հասակում։ Մաքս Վիխուրան[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 190] հաղորդել է մի քանի դրա նման զարմանալի դեպքեր հիբրիդային ուռենիների վրա իր կատարած դիտումներից։ Պետք է այստեղ նշել, որ կուսածնության որոշ դեպքերում մետաքսագործ-թիթեռի չբեղմնավորված ձվերի մեջ սաղմերն անցնում են զարգացման վաղ ստադիաները, հետո ոչնչանում են, նման այն սաղմերին, որոնք առաջացել են տարբեր տեսակների միջև տեղի ունեցած խաչաձևման միջոցով։ Քանի դեռ ծանոթ չէի այս փաստերի հետ, ես դժկամորեն էի հավատում, որ հիբրիդային սաղմերի վաղաժամ մահը հաճախակի երևույթ է, որովհետև հիբրիդները, քանի որ արդեն ծնվել են, սովորաբար լինում են առողջ և երկարակյաց, ինչպես այդ մենք տեսնում ենք սովորական ջորու օրինակի վրա։ Սակայն հիբրիդները մինչև ծնվելը և ծնվելուց հետո գտնվում են տարբեր պայմաններում։ Ծնվելով և ապրելով այն երկրում, որտեղ ապրում են երկու ծնողները, նրանք սովորաբար ընկնում են հարմար կենսապայմանների մեջ։ Սակայն հիբրիդը միայն կիսով չափ ունենում է մոր բնույթն ու կերտվածքը, ուստի մինչև ծնվելը, քանի դեռ նա սնվում է մոր արգանդում, կամ մոր արտադրած ձվի կամ սերմի մեջ, նա կարող է գտնվել որոշ տեսակետներից անհարմար պայմաններում, և, հետևաբար, վաղ հասակում ոչնչանալու վտանգին է ենթարկվում, մանավանդ որ բոլոր շատ մատղաշ օրգանիզմները արտասովոր կերպով զգայուն են դեպի գոյության վնասակար կամ անբնական պայմանները, վերջ ի վերջո ամենից ավելի հավանական է, որ պատճառը գտնվում է բեղմնավորման սկզբնական գործողության որևէ անկատարելության մեջ, որից առաջանում է սաղմի անկատար զարգացումը, և ոչ թե այն պայմանների մեջ, որոնք նրա վրա ներգործում են հետագայում։
Ինչ վերաբերում է այն հիբրիդներին, որոնց սեռական տարրերը ոչ բոլորովին զարգացած են, ապա այդ դեպքում բանը մի քիչ այլ է։ Ես շատ անգամ վկայակոչել եմ փաստերի ընդարձակ մի խումբ, որոնք ցույց են տալիս, որ կենդանին և բույսը, երբ տեղափոխվում են անբնական պայմանների մեջ, այդ դեպքում նրանք շատ հակված են լինում ունենալու վերարտադրողական սիստեմի լուրջ խանգարումներ։ Եվ իսկապես, դա մեծ խոչընդոտ է հանդիսանում կենդանիների ընտանեցմանը։ Այդ հողի վրա առաջացած անպտղաբերության և հիբրիդների անպտղաբերության միջև նմանության շատ գծեր կան։ Այսպես, երկու դեպքում էլ անպտղաբերությունը կախված չէ առողջության ընդհանուր վիճակից և հաճախ հանդիպում է հասակի բարձրացման կամ խիստ փարթամ զարգացման հետ միասին։ Այնուհետև, թե՛ մեկ և թե՛ մյուս դեպքում անպտղաբերությունն արտահայտվում է տարթեր աստիճանով։ Երկու դեպքում էլ արական տարրը ավելի շատ է տուժում. բայց երբեմն էլ լինում է հակառակը։ Երկու դեպքում էլ այդ հակումը որոշ չափով կապ ունի սիստեմատիկական ազգակցության հետ, ինչպես այդ երևում է նրանից, որ կենդանիների և բույսերի ամբողջ խմբեր սեռական տեսակետից անզոր են դառնում միևնույն անբնական պայմանների ազդեցության տակ, և տեսակների ամբողջ խմբեր ձեռք են բերում անպտղաբեր հիբրիդներ առաջացնելու հակում։ Մյուս կողմից, ամբողջ խմբի մեջ մի տեսակ երբեմն դիմանում է պայմաններում տեղի ունեցած խոր փոփոխություններին, չկորցնելով իր պտղաբերությունը, իսկ որոշ տեսակներ տվյալ խմբում առաջացնում են արտասովոր պտղաբեր հիբրիդներ։ Ոչ ոք չի կարող առանց նախնական փորձի ասել, թե տվյալ կենդանին արդյոք կպտղաբերի գերության մեջ և որևէ օտարերկրյա բույս արդյոք ազատ կերպով սերմեր կտա մշակովի վիճակում. չի կարելի նաև ասել, թե միևնույն սեռի որևէ երկու տեսակներ կտա՞ն արդյոք ավելի կամ պակաս անպտղաբեր հիբրիդներ։ Վերջապես, եթե օրգանական էակները մի քանի սերունդների ընթացքում շրջապատված են լինում իրենց համար անհատուկ պայմաններով, այդ դեպքում նրանք դառնում են չափազանց փոփոխական, որն, ըստ երևույթին, մասամբ կախված է նրանց վեր արտադրողական սիստեմի յուրահատուկ խանգարումից, թեև ավելի փոքր չափով, քան այն դեպքում, երբ սկսվում է անպտղաբերությունը։ Նույնն է նկատվում նաև հիբրիդների մեջ, որովհետև նրանց ժառանգները հաջորդական սերունդներում փոփոխականության մեծ հակում ունեն, որը նկատել են բոլոր փորձարարները։
Այսպիսով, մենք տեսնում ենք, որ երբ օրգանական էակները դրվում են նոր ու անբնական պայմանների մեջ և երբ երկու տեսակների անբնական խաչաձևման միջոցով ստացվում են հիբրիդներ, երկու դեպքում էլ վերարտադրողական սիստեմը, անկախ առողջության ընդհանուր դրությունից, խանգարվում է խիստ նման ձևով։ Մի դեպքում խախտվում են կյանքի բուն պայմանները, թեև հաճախ այնքան թույլ չափով, որ մենք այդ չենք նկատում, հիբրիդներին վերաբերող մյուս դեպքում արտաքին պայմանները մնում են նախկինը, բայց կազմվածքն արդեն ցնցված է կառուցվածքի և կերտվածքի երկու տարբեր տիպերի միաձուլվելուց, հաշվելով այդ թվում նաև վերարտադրողական սիստեմը։ Իրոք, հազիվ թե հնարավոր լինի, որ երկու կազմվածքներ միանան իրար հետ առանց որոշ խանգարման իրենց զարգացման մեջ կամ զանազան մասերի և օրգանների պարբերական գործունեության մեջ կամ մեկը մյուսի հետ և կյանքի պայմանների նկատմամբ ունեցած փոխհարաբերությունների մեջ։ Երբ հիբրիդները միմյանց հետ խաչաձևվելիս ընդունակ են պտղաբերելու, նրանք իրենց ժառանգներին սերնդե-սերունդ փոխանցում են միևնույն խառը կազմվածքը, հետևապես մենք չպետք է զարմանանք, որ նրա պտղաբերությունը, որը թեև որոշ չափով փոփոխական է, չի նվազում, նա ընդունակ է նույնիսկ ավելանալու, որը սովորաբար հանդիսանում է ինչպես մենք արդեն բացատրել ենք, շատ մոտ ազգակցության սահմաններում տեղի ունեցող խաչաձևման արդյունք։ Վերևում բերված տեսակետը, ըստ որի հիբրիդների անպտղաբերության պատճառը երկու կազմվածքների միավորումն է, ուժեղ կերպով պաշտպանել է Մաքս Վիխուրան։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 190]
Սակայն պետք է խոստովանել, որ մենք ոչ վերոհիշյալ, ոչ էլ որևէ այլ տեսակետով չենք կարող ընդունել մի քանի փաստեր, որոնք վերաբերում են հիբրիդների անպտղաբերությանը. այդպիսիներն են, օրինակ, փոխադարձ խաչաձևումից առաջացած հիբրիդների ոչ միանման պտղաբերության երևույթները, կամ այն հիբրիդների ուժեղացած անպտղաբերությունը, որոնք պատահաբար ունեն բացառիկ նմանություն մաքուր ծնողներից մեկն ու մեկի հետ։ Ես բոլորովին չեմ պնդում նաև, թե նախորդ դիտողությունները թափանցում են հարցի բուն էության մեջ, միանգամայն գիտակցելով, որ նրանք ոչ մի բացատրություն չեն տալիս, թե ինչու անբնական պայմանների մեջ ընկած օրգանիզմը դառնում է անպտղաբեր։ Ես աշխատել եմ միայն ցույց տալ, որ երկու դեպքում էլ, որոնք որոշ չափով նման են իրար, ընդհանուր արդյունքը լինում է անպտղաբերությունը, որը մի դեպքում առաջանում է կենսապայմանների խախտվելուց, մյուսը՝ երկու կազմվածքների միավորվելու հետևանքով օրգանիզմի խանգարումից։
Նման զուգահեռությունը կիրառելի է նաև մի այլ նմանօրինակ, բայց և այնպես շատ տարբեր կատեգորիայի փաստերի նկատմամբ։ Գոյություն ունի մի հին և համարյա ընդհանրական համոզմունք, որը հիմնված է զգալի թվով վկայությունների վրա, որոնք ես բերել եմ մի այլ տեղում, այն է՝ որ կենսապայմանների մեջ տեղի ունեցող թույլ փոփոխությունները բարերար ազդեցություն են գործում բոլոր կենդանի էակների վրա։ Մենք տեսնում ենք, որ գործնականում այդ կատարում են բոլոր ֆերմերները և պարտիզպանները, որոնք միմյանց հետ շարունակ փոխանակում են սերմեր, պալարներ և այլն, ստանալով և ուղարկելով դրանք մի տեսակ հող ու կլիմա ունեցող վայրից մի ուրիշ վայր։ Կենդանիների առողջացման ժամանակ կենսակերպի մեջ տեղի ունեցող ամեն մի փոփոխություն շատ բարենպաստ ազդեցություն է գործում։ Այնուհետև, ինչպես բույսերի, այնպես էլ կենդանիների վերաբերմամբ կան ամենաակնհայտ ապացույցներ, որ միևնույն տեսակին պատկանող, բայց որոշ չափով մեկը մյուսից տարբերվող երկու անհատների միջև խաչաձևումը ժառանգներին հաղորդում է ուժ և պտղաբերություն, և որ մյուս կողմից, մշտական, մի քանի սերունդների ընթացքում շարունակվող խաչաձևումները մոտ ազգականների միջև, եթե նրանք պահվում են միևնույն կենսապայմաններում, գրեթե միշտ տանում են դեպի հասակի փոքրացում, թուլություն կամ անպտղաբերություն։
Այդ պատճառով էլ ինձ թվում է, մի կողմից, որ կենսապայմանների թույլ փոփոխություններն օգուտ են բերում օրգանական էակներին, իսկ մյուս կողմից որ թեթև խաչաձևումները, այսինքն խաչաձևումները միևնույն տեսակի արուների և էգերի միջև, որոնք շրջապատված են եղել փոքր ինչ տարբեր պայմաններով կամ թեթևակի տարբերվել են, ուժ և պտղաբերություն են հաղորդում սերունդներին։ Բայց ինչպես մենք տեսանք, եթե երկար ժամանակ բնական վիճակում իրենց շրջապատող որոշ միապաղաղ պայմանների ընտելացած օրգանիզմների, կենսապայմանները ենթարկենք զգալի փոփոխությունների, ինչպես այդ լինում է գերության մեջ, ապա շատ հաճախ նրանք դառնում են ավելի կամ պակաս չափով անպտղաբեր, և մենք գիտենք, որ խոշոր կամ տեսակային տարբերություններ ձեռք բերած երկու ձևերի միջև խաչաձևումը առաջացնում է հիբրիդներ, որոնք համարյա միշտ որոշ չափով անպտղաբեր են։ Ես լիովին համոզված եմ, որ այդ կրկնակի զուգահեռությունը ամենևին պատահական կամ թվացող չէ։ Ով որ կարող է բացատրել, թե ինչու փիղը և այլ շատ կենդանիներ անընդունակ են իրենց հայրենիքում նույնիսկ ոչ լրիվ գերության մեջ պտղաբերելու, նա կկարողանա բացատրել այն սկզբնական պատճառը, թե ինչու հիբրիդները ընդհանրապես այնքան անպտղաբեր են։ Միևնույն ժամանակ նա կկարողանա բացատրել, թե ինչպես է պատահել, որ մեր ընտանի կենդանիներից մի քանիսների ցեղերը, որոնք հաճախ են ենթարկվել նոր և այն էլ ոչ միանման պայմանների ներգործությանը, իրար միջև խաշաձևվելիս միանգամայն պտղաբեր են, թեև նրանք առաջացել են տարբեր տեսակներից, որոնք հավանորեն անպտղաբեր կլինեին, եթե նրանց խաչաձևեինք նախնական վայրի վիճակում։ Փաստերի վերոհիշյալ երկու զուգահեռ շարքերը, ըստ երևույթին, իրար հետ միացած են որոշ ընդհանուր, բայց անհայտ կապով, որ էական առնչություն ունի կենսական սկզբունքի հետ. այդ սկզբունքը, մ-ր Հերբերտ Սպենսերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 18] որոշման համաձայն այն է, որ կյանքը կախված է կամ նույնիսկ կազմված է զանազան ուժերի մշտական ազդեցությունից և հակազդեցությունից, որոնք ինչպես որ բնության մեջ ամենուրեք, մշտապես ձգտում են հավասարակշռության, իսկ երբ այդ ձգտումը թեթևակի, խախտված է որևէ փոփոխությամբ, այդ դեպքում կենսական ուժերը ձեռք են բերում մեծ հզորություն։
ՓՈԽԱԴԱՐՁ ԴԻՄՈՐՖԻԶՄ ԵՎ ՏՐԻՄՈՐՖԻԶՄ
Մենք այստեղ համառոտակի կքննարկենք այն հարցը որը, ինչպես մենք, կտեսնենք, որոշ լույս է սփռում հիբրիդացման վրա։ Զանազան ընտանիքների պատկանող որոշ բույսեր հանդես են գալիս երկու ձևերով, որոնք գոյություն ունեն համարյա հավասար թվով և ոչնչով չեն տարբերվում իրարից, բացառությամբ բազմացման օրգանների, այն է՝ մի ձևն ունենում է երկար վարսանդ կարճ առէջքներով միասին, մյուսը կարճ վարսանդ և երկար առէջքներ, միաժամանակ երկուսն էլ ունենում են ոչ միանման մեծության փոշեհատիկներ։ Տրիմորֆ բույսերն ունենում են երեք ձևեր, որոնք նման ձևով տարբերվում են իրենց վարսանդների և առէջքների երկարությամբ, փոշեհատիկների մեծությամբ ու գույնով և մի քանի այլ հատկանիշներով, քանի որ այդ երեք ձևերից ամեն մեկն ունի երկու խումբ առէջքներ, ուստի երեք ձևերն ընդհանուր առմամբ ունեն վեց խումբ առէջքներ և երեք տիպի վարսանդներ։ Այդ օրգանները մեկը մյուսի նկատմամբ երկարությամբ այնքան չափակից են, որ երկու ձևերի առէջքների կեսը գտնվում է երրորդ ձևի սպիի հետ միևնույն մակարդակի վրա։ Մինչդեռ ես ցույց եմ տվել, և այդ հաստատել են ուրիշ դիտողներ, որ այդ բույսերից լրիվ պտղաբերություն ստանալու համար անհրաժեշտ է, որպեսզի մի ձևի սպին փոշոտվի մյուս ձևի համապատասխան բարձրություն ունեցող առէջքներից վերցված ծաղկափոշիով։ Այնպես որ դիմորֆ տեսակների մոտ երկու սեռական միավորումներ, որոնց կարելի է անվանել օրինական, միանգամայն պտղաբեր են, և երկուսը, որոնց կարելի է անվանել ապօրինի, ավելի կամ պակաս չափով անպտղաբեր են։ Տրիմորֆ տեսակների մոտ վեց սեռական միավորումներ օրինական են, կամ լիովին պտղաբեր են, և տասներկուսը ապօրինի են, կամ ավելի կամ պակաս չափով անպտղաբեր են։
Այն անպտղաբերությունը, որ նկատվում է զանազան դիմորֆ և տրիմորֆ բույսերի մոտ, երբ նրանք բեղմնավորվում են ապօրինի ձևով, այսինքն վարսանդի բարձրությանը չհամապատասխանող առէջքների ծաղկափոշով, իր աստիճանով խիստ տարբեր է՝ ընդհուպ մինչև կատարյալ ու լրիվ անպտղաբերությունը, մի խոսքով՝ միանգամայն այնպես, ինչպես որ տարբեր տեսակների խաչաձևման դեպքում։ Եվ ինչպես որ այս վերջին դեպքում անպտղաբերության աստիճանը մեծ չափով կախված է կյանքի առավել կամ պակաս բարենպաստ պայմաններից, բանն այդպես է, ըստ իմ դիտումների, նաև այդ ապօրինի միավորումների հետ։ Լավ հայտնի է, որ եթե որևէ ծաղկի սպիի վրա ընկնում է մի այլ տեսակի ծաղկափոշի, և հետո նույն սպիի վրա կընկնի, թեկուզ զգալի ժամանակամիջոցից հետո, ծաղկի սեփական ծաղկափոշին, ապա նա այնպիսի գերակշռող ազդեցություն է գործում, որ սովորաբար ոչնչացնում է օտար ծաղկափոշու ներգործությունը[63]։ Նույնը կարելի է ասել նաև միևնույն տեսակի զանազան ձևերի ծաղկափոշու մասին, որովհետև օրինական ծաղկափոշին ուժեղ կերպով գերիշխում է ապօրինի ծաղկափոշու վրա, երբ նրանք երկուսն էլ ընկնում են միևնույն սպիի վրա։ Ես դրանում համոզվել եմ, բեղմնավորելով զանազան ծաղիկներ նախ ապօրինի ձևով և հետո, 24 ժամ անցած՝ օրինական ձևով այն ծաղկափոշով, որ վերցված էր յուրահատուկ գունավորված մի տարատեսակից, և այդ ձևով ստացված սերմերից աճած բոլոր բույսերը նման ձևով գունավոր էին։ Այդ ցույց է տալիս, որ օրինական ծաղկափոշին, թեև նա զետեղվել էր 24 ժամ անցած, բոլորովին ոչնչացրել կամ կանխել է նախքան այդ զետեղված ապօրինի ծաղկափոշու ներգործոլթյունը։ Այնուհետև, ինչպես երկու տեսակների փոխադարձ խաչաձևման ժամանակ, որոշ դեպքերում արդյունքի մեջ նկատվում է մեծ տարբերություն, նույնն էլ նկատվում է տրիմորֆ բույսերի վրա, այսպես, օրինակ, միջակ երկարությամբ վարսանդներ ունեցող Lythcum salicaria ձևը ապօրինի եղանակով մեծ հեշտությամբ բեղմնավորվելով ավելի երկար առէջքներ և կարճ վարսանդ ունեցող ձևի ծաղկափոշու միջոցով, շատ սերմ է տվել, մինչդեռ այս վերջին ձևը ոչ մի սերմ չի տվել, երբ բեղմնավորվել է միջին երկարության վարսանդներ ունեցող ձևի ավելի երկար առէջքների ծաղկափոշիով։
Այս բոլոր, ինչպես և այլ տեսակետներից, որ կարելի էր դրանից զատ մատնանշել, միևնույն անկասկածելի տեսակի ձևերը միանալով ապօրինի ձևով, իրենց այնպես են պահում, ինչպես երկու տարբեր տեսակներ խաչաձևվելիս։ Սա հարկադրել է ինձ չորս տարի շարունակ մանրազնին կերպով դիտել այն բույսերը, որոնք աճել էին զանազան ապօրինի միավորումներից ստացված սերմերից։ Գլխավոր արդյունքն այն է, որ այդ, ինչպես նրանց կարելի է անվանել, ապօրինածին բույսերը ոչ լրիվ պտղաբեր են։ Դիմորֆ տեսակներից հնարավոր է ստանալ ինչպես երկար, այնպես էլ կարճ վարսանդներով ապօրինի բույսեր, և տրիմորֆներից էլ բոլոր երեք ապօրինի ձևերը։ Այս վերջինները կարելի է հետո միացնել սովորական օրինական ձևով։ Երբ այդ արվում է, ապա ոչ մի ակնհայտ պատճառ չկա, որ նրանք չտային նույնքան սերմ, որքան տվել են նրանց ծնողները օրինական բեղմնավորման դեպքում։ Բայց այդպես չի լինում։ Նրանք բոլորն էլ տարբեր աստիճաններով անպտղաբեր են, ըստ որում մի քանիսն այնքան անբուժելիորեն ու կատարելապես անպտղաբեր են, որ չորս տարվա ընթացքում չեն տվել ոչ մի սերմ և նույնիսկ ոչ մի խակ պտղիկ։ Այդ ապօրինի բույսերի անպտղաբերությունը, երբ նրանք բեղմնավորում են մեկը մյուսին օրինական եղանակով, կարելի է խիստ իմաստով համեմատել հիբրիդների անպտղաբերության հետ, երբ նրանք inter se խաչաձևվում են։ Մյուս կողմից, եթե հիբրիդը խաչաձևվում է երկու մաքուր ծնողական տեսակներից մեկի հետ, այդ դեպքում անպտղաբերությունը զգալիորեն նվազում է. նույնը տեղի է ունենում նաև այն ժամանակ, երբ ապօրինի բույսերը բեղմնավորվում են օրինականներով։ Ճիշտ այնպես, ինչպես հիբրիդների անպտղաբերությունը միշտ չէ, որ ընթանում է երկու ծնողական տեսակների միջև առաջին խաչաձևման դժվարության հետ միասին, այդպես էլ որոշ ապօրինածին բույսերի անպտղաբերությունը արտասովոր կերպով մեծ է եղել, մինչդեռ այն միավորման անպտղաբերությունը, որից առաջացել են նրանք, բոլորովին մեծ չի եղել։ Մի սերմնատուփիկից վերցրած սերմերից աճեցրած հիբրիդների անպտղաբերության աստիճանը բնածին կերպով փոփոխական է. նույնը նկատելի կերպով երևան է գալիս նաև ապօրինի բույսերի մոտ։ Վերջապես, շատ հիբրիդներ ծաղկում են խիստ առատ ու տևական կերպով, մինչդեռ, մյուսները, ավելի անպտղաբերները, շատ քիչ ծաղիկ են տալիս և ներկայացնում են թույլ, խղճալի թզուկներ, բոլորովին նման դեպքեր հանդիպում են նաև զանազան դիմորֆ և տրիմորֆ բույսերի ապօրինի ժառանգների մեջ։
Ընդհանրապես ապօրինածին բույսերի և հիբրիդների միջև բնույթի և վարքի տեսակետից ամենասերտ նմանություն կա։ Հազիվ թե չափազանցություն լինի, եթե ասենք, որ ապօրինածին բույսերը հիբրիդներ են, որոնք ստացված են միևնույն տեսակի սահմաններում որոշ ձևերի անբնական միավորման հետևանքով, մինչդեռ սովորական հիբրիդներն առաջանում են այսպես կոչված տարբեր տեսակների միջև կատարված անբնական միավորումից։ Մենք արդեն տեսանք նաև, որ բոլոր տեսակետներից շատ մոտ նմանություն կա առաջին ապօրինի միավորումների և առանձին տեսակների առաջին խաչաձևումների միջև։ Գուցե այդ ավելի լրիվ պարզաբանվի հետևյալ օրինակով։ Դիցուք, թե որևէ բուսաբան գտել է տրիմորֆ Lythrum salicaria-ի երկար վարսանդներ ունեցող երկու խիստ տարբերվող ձևեր (իսկ այդպիսիներ հանդիպում են) և նա որոշել է խաչաձևման միջոցով ստուգել, թե արդյոք նրանք առանձին տեսակներ են, թե ոչ։ Նա կգտներ, որ այդ դեպքում ստացվում է սերմերի նորմալ թվի մոտավորապես մեկ հինգերորդ մասը միայն և որ ընդհանրապես այդ երկու բույսերը բոլոր վերոհիշյալ տեսակետներից իրենց պահում են ինչպես երկու առանձին տեսակներ։ Բայց այդ հարցը վերջնականապես պարզելու համար նա կաճեցներ բույսեր այդ ենթադրյալ հիբրիդային սերմերից և կգտներ, որ նրանք խղճուկ գաճաճներ են, միանգամայն անպտղաբեր և բոլոր տեսակետներից նման են սովորական հիբրիդներին։ Նա այն ժամանակ կարող էր պնդել, որ ինքն իսկապես ապացուցել է, բոլորի կողմից ընդունված տեսակետի համաձայն, որ իր երկու տարատեսակները նույնպիսի լավ զատելի տեսակներ են, ինչպես որ յուրաքանչյուր այլ տեսակ, սակայն դա նրա կողմից լիակատար սխալ կլիներ։
Դիմորֆ և տրիմորֆ բույսերի մասին վերոհիշյալ փաստերը շատ կարևոր են, որովհետև, առաջին, նրանք մեզ ապացուցում են, որ ինչպես առաջին խաչաձևումների, այնպես էլ հիբրիդների ֆիզիոլոգիական հատկանիշը՝ նվազած պտղաբերությունը չի կարող վստահելի չափանիշ ծառայել տեսակները տարբերելու համար․ երկրորդ, մենք կարող ենք եզրակացնել, որ գոյություն ունի մի ինչ որ անհայտ կապ ապօրինի միավորումների անպտղաբերության և նրանց ապօրինի հաջորդների անպտղաբերության միջև, և դա մեզ հանգեցնում է նույն տեսակետը տարածելու առաջին խաչաձևումների և հիբրիդների վրա․ երրորդ, մենք գտնում ենք, և ինձ թվում է սա առանձնապես կարևոր է, որ կարող են գոյություն ունենալ միևնույն տեսակի երկու կամ երեք ձևեր և ոչնչով չտարբերվել միմյանցից իրենց կառուցվածքով և կերտվածքով արտաքին պայմանների նկատմամբ և այնուամենայնիվ լինել անպտղաբեր, եթե նրանց միավորենք որոշ եղանակով։ Չէ որ մենք պիտի հիշենք, որ տվյալ դեպքում անպտղաբերություն է առաջացնում միևնույն ձևի անհատների երկու սեռական տարրերի միավորումը, օրինակ, երկար վարսանդներ ունեցող անհատների միացումը, մինչդեռ երկու տարբեր ձևերին հատուկ սեռական տարրերի միավորումը պտղաբեր է լինում։ Հետևաբար, այս օրինակը առաջին հայացքից թվում է միանգամայն հակառակը այն բանի, ինչ որ տեղի է ունենում մի տեսակի անհատների սովորական միավորումների և տարբեր տեսակների խաչաձևման ժամանակ։ Սակայն կասկածելի է, որ այդ իրոք այդպես է, բայց ես երկար չեմ խոսի այդ մութ հարցի մասին։
Ինչ էլ որ լինի, դիմորֆ և տրիմորֆ բույսերի քննարկումից մենք կարող ենք անել այն հավանական եզրակացությունը, որ զանազան տեսակների խաչաձևման դեպքում առաջացող անպտղաբերությունը և նրանց հիբրիդային հաջորդների անպտղաբերությունը կախված է բացառապես սեռական տարրերի բնույթից, այլ ոչ թե նրանց կառուցվածքի կամ ընդհանուր կերտվածքի մեջ եղած որևէ տարբերությունից։ Նույնպիսի եզրակացության է հանգեցնում մեզ փոխադարձ խաչաձևումների քննարկությունը, երբ մի տեսակի արական անհատը չի կարող միավորվել կամ միավորվում է միայն մեծ դժվարությամբ մի այլ տեսակի իգական անհատի հետ, մինչդեռ հակառակ խաչաձևումը կարող է ստացվել զարմանալի հեշտությամբ։ Այնպիսի գերազանց դիտող, ինչպես Գերտներն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] է, նույնպես եկել է այն եզրակացության, որ տեսակները խաչաձևվելիս անպտղաբեր են լինում այն տարբերությունների հետևանքով, որոնք սահմանավորվում են նրանց վերարտադրողական սիստեմով։
ԽԱՉԱՁԵՎՎՈՂ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԵՎ ՆՐԱՆՑ ԽԱՌՆԱՑԵՂԵՐԻ ՊՏՂԱԲԵՐՈՒԹՅՈԻՆՆ ԸՆԴՀԱՆՈՒՐ ԵՐԵՎՈՒՅԹ ՉԷ
Տեսակների և տարատեսակների միջև հիմնական տարբերության գոյության մասին որպես ճնշող փաստարկ կարելի է բերել այն, որ տարատեսակներն արտաքին տեսքով ինչքան էլ տարբերվեն մեկը մյուսից, բոլորովին հեշտ կերպով խաչաձևվում են միմյանց հետ և տալիս են միանգամայն պտղաբեր հաջորդներ։ Բացառությամբ մի քանիսների, որոնք այժմ կմատնանշվեն, ես լիովին ընդունում եմ, որ ընդհանուր կանոնն այդպես է։ Բայց այդ հարցը շրջապատված է դժվարություններով, որովհետև դառնալով բնության մեջ առաջացած տարատեսակներին, մենք տեսնում ենք, որ երկու ձևեր, որոնք մինչ այդ համարվել են տարատեսակներ, հենց որ խաչաձևվելիս թեկուզ մի քիչ անպտղաբեր են լինում, իսկույն բնախույզների մեծամասնությունը սկսում է նրանց համարել տեսակներ։ Այսպես, օրինակ, կարմիր և կապույտ տզրուկախոտերը[64], որոնց բուսաբանների մեծամասնությունը համարում է որպես տարատեսակներ, Գերտների[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] փորձերում խաչաձևման ժամանակ բոլորովին անպտղաբեր են եղել, և այդ պատճառով նա նրանց դասում է անկասկածելի տեսակների շարքը։ Եթե մենք կշռադատենք՝ դուրս չգալով շրջանակից (այսինքն՝ այդ ենթադրությունից), ապա բնական վիճակում գտնվող բոլոր տարատեսակների պտղաբերությունը, իհարկե, պետք է ընդունենք։
Եթե մենք դառնանք այն տարատեսակներին, որոնք առաջացել են ընտանի վիճակում կամ համարվում են որպես այդպիսիներ, ապա մենք այստեղ ևս չենք ազատվում որոշ կասկածներից։ Իրոք, եթե մեզ հաղորդում են, օրինակ, որ հարավ-ամերիկական մի քանի տեղական շներ դժկամությամբ են խաչաձևվում եվրոպական շների հետ, ապա ինքնըստինքյան յուրաքանչյուրի մտքով անցնում է մի բացատրություն, և այն, հավանորեն, արդարացի է, որ նրանք ծագել են սկզբնապես տարբեր տեսակներից։ Այնուամենայնիվ, արտաքին տեսքով մեկը մյուսից խիստ տարբերվող այդքան շատ ընտանի կենդանիների, օրինակ՝ կաղամբի կամ աղավնիների զանազան ցեղերի պտղաբերությունը շատ ուշագրավ փաստ է, մանավանդ, երբ մենք ուշադրության ենք առնում այն, թե ինչքան շատ կան տեսակներ, որոնք, չնայած միմյանց հետ ունեցած շատ մոտ նմանությանը, այնուամենայնիվ խաչաձևվելիս բոլորովին անպտղաբեր են։ Սակայն զանազան նկատառումներ մեզ հարկադրում են ընտանի կենդանիների այդ պտղաբերությունը այնքան էլ զարմանալի չհամարել։ Նախ կարելի է նկատել, որ երկու տեսակների միջև եղած արտաքին տարբերությունների աստիճանը ամենևին վստահելի ուղեցույց չի ծառայում նրանց փոխադարձ անպտղաբերության աստիճանը որոշելու համար, ուստի և տարատեսակների նման տարբերությունները նույնպես չեն կարող վստահելի ցուցում լինել։ Կարելի է արժանահավատ համարել, որ տեսակների համար պատճառը թաքնված է բացառապես նրանց սեռական կերտվածքի տարբերություններում։ Փոփոխական պայմանների ազդեցության ներքո, որոնց ենթարկվել են ընտանեցված կենդանիներն ու մշակովի բույսերը, նրանք այնքան քիչ հակում են ձեռք բերել փոփոխելու վերարտադրողական սիստեմը փոխադարձ անպտղաբերության հանգեցնող ուղղությամբ, որ մենք բավարար հիմք ունենք ընդունելու Պալլասի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 104] ուղղակի հակառակ տեսակետը, այն է, որ նման պայմաններն ընդհանրապես վերացնում են այդ հակումը, այնպես որ այն տեսակների ընտանեցված հետնորդները, որոնք բնական դրության մեջ հավանորեն եղել են որոշ չափով անպտղաբեր, միմյանց հետ խաչաձևվելիս դառնում են միանգամայն պտղաբեր։ Ինչ վերաբերում է բույսերին, ապա մշակությունը նրանց մեջ ամենից ավելի քիչ է հարուցում անպտղաբերության հակում զանազան տեսակների խաչաձևման դեպքում. և իրոք, մի քանի արժանահավատ դեպքերում, որոնց մասին մենք արդեն թեթևակիորեն խոսել ենք, որոշ բույսեր մշակության ազդեցության տակ փոփոխվել են ուղղակի հակառակ ձևով. նրանք դարձել են ինքնաբեղմնավորման համար անընդունակ, միաժամանակ պահպանելով այլ տեսակներին բեղմնավորելու և նրանցից բեղմնավորվելու ընդունակությունը։ Եթե ընդունենք Պալլասի ուսմունքը՝ երկարատև ընտանեցման միջոցով անպտղաբերությունը վերացնելու մասին, իսկ այդ հազիվ թե կարելի է ժխտել, ապա չափազանց անհավանական է դառնում, որ նման պայմանները երկար տևելով, կարողանային առաջացնել այդ հակումը, թեև առանձին դեպքերում նրանք կարող են յուրահատուկ կերտվածք ունեցող տեսակների անպտղաբերության պատճառ դառնալ։ Իմ կարծիքով, դրանով է բացատրվում այն, թե ինչու ընտանի կենդանիներից չեն ստացվում փոխադարձաբար անպտղաբեր տարատեսակներ և թե ինչու բույսերի մեջ այդ նկատվել է միայն քիչ դեպքերում, որոնք հիմա ցույց կտրվեն։
Մեր քննարկելի հարցի իսկական դժվարությունը, ինչպես ինձ թվում է, այն չէ, թե ինչու ընտանի տարատեսակները խաչաձևվելիս փոխադարձաբար անպտղաբեր չեն դարձել, այլ այն է, թե ինչու անպտղաբերությունը դարձել է այնքան սովորական երևույթ բնական տարատեսակների մոտ, հենց որ նրանք հաստատուն կերպով փոփոխվել են բավարար չափով, որպեսզի բարձրացվեին մինչև տեսակների աստիճանի։ Մենք հեռու ենք պատճառները Ճիշտ կերպով գիտենալուց, և այդ զարմանալի չէ, եթե ուշադրության առնենք այն, թե ինչպես խորն է մեր անտեղյակությունը վերարտադրողական սիստեմի նորմալ և աննորմալ գործունեության մասին։ Բայց մենք հասկանում ենք, որ տեսակներն իրենց բազմաթիվ մրցակիցների դեմ մղած գոյության կռվում պետք է երկար ժամանակաշրջանների ընթացքում ենթարկվեին ավելի միանման պայմանների ներգործությանը, քան այն պայմանները, որոնց մեջ գտնում են ընտանի տարատեսակները, իսկ դա ի հարկե, կարող է հետևանքի մեջ առաջացնել մեծ տարբերություն։ Եվ իրոք, մենք գիտենք, թե ինչպես հաճախ վայրի կենդանիներն ու բույսերը, եթե նրանք վերցվում են իրենց բնական իրադրությունից և ենթարկվում են գերության, անպտղաբեր են դառնում, բնական իրադրության մեջ մշտապես ապրող օրգանական էակների արտադրողական ֆունկցիաները հավանորեն ճիշտ նույն ձևով շատ զգայուն կլինեն անբնական խաչաձևման ազդեցության նկատմամբ։ Մյուս կողմից, կարելի է նախապես սպասել, որ ընտանի ցեղերը, որոնք, ինչպես արդեն ապացուցում է նրանց ընտելացվածության բուն փաստը, հենց սկզբից շատ զգայուն չեն եղել դեպի իրենց կենսապայմանների փոփոխությունները և որոնք այժմ էլ առանց նվազեցնելու իրենց պտղաբերությունը, կարողանում են դիմանալ պայմանների կրկնական փոփոխություններին, կառաջացնեն տարատեսակներ, որոնց արտադրողական սիստեմը քիչ է ենթարկվում վտանգավոր խանգարման, երբ նրանք խաչաձևվում են նույն ձևով առաջացած այլ տարատեսակների հետ։
Մինչև այժմ ես այնպես եմ արտահայտվել, կարծես թե միևնույն տեսակին պատկանող տարատեսակները միմյանց հետ խաչաձևվելիքս անպայման պտղաբեր են։ Բայց անկարելի է ժխտել այն փաստերը, որոնք վկայում են անպտղաբերության որոշ աստիճանի գոյության մասին հետևյալ սակավաթիվ դեպքերում, որ ես համառոտակի կշարադրեմ։ Այդ փաստերը համենայն դեպս ոչնչով վատ չեն նրանցից, որոնց հիման վրա մենք հավատում ենք բազմաթիվ տեսակների անպտղաբերությանը։ Բացի դրանից, նրանք վերցված են մեր հակառակորդների հաղորդումներից, որոնք բոլոր դեպքերում պտղաբերության և անպտղաբերության վրա նայում են որպես տեսակային տարբերությունների մի վստահելի չափանիշի։ Գերտները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] մի քանի տարիների ընթացքում բուծել է դեղին սերմեր ունեցող եգիպտացորենի գաճաճ ցեղը և կարմիր սերմեր ունեցող բարձրահասակ տարատեսակը, որոնք մեկը մյուսի կողքին աճել են նրա պարտեզում, և թեև այդ բույսերը բաժանասեռ են, նրանք երբեք չեն խաչաձևվել բնական եղանակով։ Այնուհետև նա բեղմնավորել է մի ցեղի տասներեք ծաղիկները մյուսի ծաղկափոշով. բայց ստացվել է այն, որ միայն մեկ կողրը տվել է սերմեր, և այն էլ ընդամենը միայն հինգ հատիկ։ Այս դեպքում փոշոտման կատարումն ինքը չէր կարող վնասակար ներգործություն ունենալ, որովհետև եգիպտացորենը բաժանասեռ է։ Ես կարծում եմ, որ ոչ ոք չի կասկածի, թե եգիպտացորենի այդ տարատեսակները առանձին տեսակներ են եղել, և կարևոր է նշել այն, որ այդ եղանակով ստացված հիբրիդային բույսերը կատարելապես պտղաբեր են եղել, այնպես որ նույնիսկ Գերտները չի վստահել այդ երկու տարատեսակները համարել զանազան տեսակներ։
Ժիրու-դը-Բյուզարենգը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 191] խաչաձևել է եգիպտացորենի նման բաժանասեռ դդումի երեք տարատեսակներ և պնդում է, որ նրանց փոխադարձ բեղմնավորումը այնքան ավելի հեշտ է ընթանում, որքան մեծ է լինում նրանց միջև տարբերությունը։ Թե որքան կարելի է վստահել այդ փորձերին, ես չգիտեմ, բայց այն ձևերը, որոնց վրա նրանք կատարվել են, Սաժերեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 186] համարում է տարատեսակներ, իսկ նա իր դասակարգումը հիմնում է գլխավորապես անպտղաբերության հատկանիշի վրա. նույն եզրակացության է եկել նաև Նոդենը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 19]
Հետևյալ դեպքը շատ ավելի ուշագրավ է և առաջին հայացքից անհավատալի է թվում, բայց նա ներկայացնում է զարմանալի մեծ թվով փորձերի արդյունք, որոնք մի քանի տարիների ընթացքում կատարվել են խռնդատի (Verbascum) ինը տեսակների վրա և այն էլ այնպիսի լավ դիտողի և մեզ թշնամի այնպիսի վկայի կողմից, ինչպիսին է Գերտները.[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] այսինքն այն, որ մի տեսակի պատկանող դեղին ու սպիտակ տարատեսակները միմյանց հետ խաչաձևվելիս առաջացնում են ավելի քիչ սերմեր, քան միևնույն տեսակի միանման գույն ունեցող տարատեսակները։ Բացի դրանից, նա պնդում է, որ երբ մի տեսակի պատկանող դեղին և սպիտակ տարատեսակները խաչաձևվում են մի այլ տեսակի դեղին ու սպիտակ տարատեսակների հետ, ապա միանման գույն ունեցող ծաղիկների միջև կատարվող խաչաձևումից ավելի շատ սերմ է ստացվում, քան տարբեր գունավորումներ ունեցողների խաչաձևումից։ Մ-ր Սկոտտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 182] նույնպես փորձեր է կատարել Verbascum-ի տեսակների և տարատեսակների վրա, և թեև նա գտնում է, որ հնարավորդ չէ հաստատել Գերտների ստացած արդյունքները տարբեր տեսակների խաչաձևման մասին, սակայն նա գալիս է այն եզրակացության, որ մի տեսակի պատկանող ոչ միանման գունավորված տարատեսակները ավելի քիչ սերմ են տալիս՝ 86:100 հարաբերությամբ, քան միանման գունավորված տարատեսակները։ Սակայն այդ տարատեսակները ոչնչով չեն տարբերվում, բացառությամբ ծաղիկների գույնի, և մի տարատեսակը երբեմն կարող է ստացվել մյուսի սերմերից։
Կյոլռեյտերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] որի դիտումների ճշտությունը հաստատել են հետագա բոլոր դիտողները, ապացուցել է մի ուշագրավ փաստ, որ սովորական ծխախոտի մի առանձին տարատեսակ ավելի պտղաբեր է, քան մյուս տարատեսակները, երբ նա խաչաձևվում է իրենից խիստ տարբերվող տեսակի հետ։ Նա փորձեր է կատարել հինգ ձևերի վրա, որոնք սովորաբար համ արվում են տարատեսակներ, և որոնց նա ենթարկել է խստապահանջ ստուգման, այն է՝ փոխադարձ խաչաձևման միջոցով, ընդ որում եկել է այն եզրակացության, որ խառնացեղերը միանգամայն պտղաբեր են։ Բայց այդ հինգ տարատեսակներից մեկը, միևնույնն է թե հոր դեր է խաղացել կամ մոր դեր Nicotiana glutinosa-ի հետ խաչաձևվելիս, միշտ առաջացրել է ավելի քիչ անպտղաբեր հիբրիդներ, քան նրանք, որոնք ստացվել են N. glutinosa-ի հետ մյուս չորս տարատեսակների խաչաձևումից։ Ինչպես երևում է, այդ մի տարատեսակի վերարտադրողական սիստեմը որոշ չափով և որոշ կերպով ձևափոխված է։
Նկատի առնելով այս փաստերը, չի կարելի այլևս պնդել, որ տարատեսակները խաչաձևվելիս անպայման միանգամայն պտղաբեր են լինում, այնուհետև ուշադրության առնելով այն, թե ինչքան դժվար է հավաստիանալ, որ տարատեսակները բնական վիճակում անպտղաբեր են, որովհետև մի քիչ անպտղաբերություն ցուցաբերած ենթադրյալ տարատեսակը գրեթե միշտ բարձրացվում է տեսակի աստիճանի. այնուհետև ուշադրության առնելով այն հանգամանքը, որ մարդը հոգ է տանում իր ընտանի տարատեսակների արտաքին հատկանիշների մասին և որ այս օրգանիզմները երկար ժամանակաշրջանների ընթացքում չեն ենթարկվում միանման կենսապայմանների, նկատի առնելով այս ամենը, մենք կարող ենք եզրակացնել, որ խաչաձևումների դեպքում պտղաբերությունը հիմնական տարբերություն չի հանդիսանում տեսակների և տարատեսակների միջև։ Խաչաձևվող տեսակների սովորական անպտղաբերությունը կարելի է վստահորեն դիտել ոչ թե որպես հատուկ ձեռք բերված կամ շնորհված հատկություն, այլ որպես նրանց սեռական տարրերի փոփոխությունների կողմնակի հետևանք, փոփոխությունների, որոնց բնույթը անհայտ է։
ՀԻԲՐԻԴՆԵՐԻ ԵՎ ԽԱՌՆԱՑԵՂԵՐԻ ՀԱՄԵՄԱՏՈՒԹՅՈՒՆԸ՝ ԱՆԿԱԽ ՆՐԱՆՑ ՊՏՂԱԲԵՐՈՒԹՅՈՒՆԻՑ
Պտղաբերության հարցից անկախ կերպով կարելի է տեսակների և տարատեսակների խաչաձևումից առաջացած սերունդները համեմատել իրար հետ շատ տեսակետներից։ Գերտները,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] որ ուժեղ ցանկություն ուներ սահմանագիծ անցկացնել տեսակի և տարատեսակի միջև, կարող էր միայն շատ քիչ, և ինչպես ինձ թվում է, չափազանց անկարևոր տարբերություններ գտնել տեսակների սերունդների, կամ այսպես կոչված հիբրիդների, և տարատեսակների սերունդների, կամ այսպես կոչված խառնացեղերի միջև։ Իսկ մյուս կողմից, նրանք շատ կարևոր տեսակետներից չափազանց նման են լինում միմյանց։
Ես այստեղ այս հարցը կքննարկեմ շատ համառոտ կերպով։ Ամենակարևոր տարբերություն է ծառայում այն, որ խառնացեղերն առաջին սերնդում հիբրիդներից ավելի փոփոխական են։ Բայց Գերտներն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] ընդունում է, որ կուլտուրական վիճակում երկար ժամանակ գտնվող տեսակներից առաջացած հիբրիդները հաճախ փոփոխական են արդեն առաջին սերնդում, և ես ինքս տեսել եմ այդպիսի զարմանալի օրինակներ։ Գերտներն այնուհետև ընդունում է, որ երկու չափազանց մոտ ազգակից տեսակներից ստացված հիբրիդները ավելի փոփոխական են, քան միմյանցից խիստ տարբեր տեսակներից առաջացածները, իսկ այդ ցույց է տալիս, որ փոփոխականության աստիճանի մեջ եղած տարբերությունն աստիճանաբար հարթվում է։ Երբ խառնացեղերը և ավելի պտղաբեր հիբրիդները բազմանում են մի քանի սերունդների ընթացքում, թե մեկ և թե մյուս դեպքում նրանց հաջորդների ծայրահեղ փոփոխականությունը հանրածանոթ փաստ է. սակայն կարելի է բերել մի քանի առանձին օրինակներ, երբ թե՛ հիբրիդները և թե՛ խառնացեղերը երկար ժամանակ պահպանում են միևնույն հատկանիշները։ Համենայն դեպս խառնացեղերի փոփոխականությունը հաջորդական սերունդների մեջ ըստ երևույթին ավելի մեծ է, քան հիբրիդների փոփոխականությունը։
Հիբրիդների համեմատությամբ խառնացեղերի ունեցած ավելի մեծ փոփոխականությունը զարմանալի չի թվում։ Եվ իրոք, խառնացեղերի ծնողները տարատեսակներն են և այն էլ շատ դեպքերում ընտանի (միայն սակավաթիվ փորձեր են կատարվել բնական տարատեսակների վրա), իսկ դրանից արդեն հասկացվում է, որ հենց ծնողների փոփոխականությունը վաղուցվա ծագում չունի և որ շատ դեպքերում նա կշարունակվի և կուժեղացնի փոփոխականությունը, որը հանդիսանում է հենց խաչաձևման ակտի արդյունքը։ Հիբրիդի առաջին սերնդի մեջ աննշան փոփոխականությունը, հակառակ հաջորդ սերունդներում արտահայտվող ուժեղ փոփոխականության, հետաքրքրական և ուշադրության արժանի փաստ է, որովհետև նա ամրապնդում է իմ տեսակետը, որի համաձայն սովորական փոփոխականության պատճառներից մեկն այն է, որ վերարտադրողական սիստեմը լինելով չափազանց զգայուն դեպի կենսապայմանների փոփոխությունը, հրաժարվում է այդ դեպքում կատարել իր նորմալ ֆունկցիան՝ արտադրել սերունդ, որը բոլոր տեսակետներից շատ նման է ծնողական ձևերին. ինչ վերաբերում է առաջին սերնդի հիբրիդներին, ապա նրանք առաջացել են տեսակներից, որոնց վերարտադրողական սիստեմը (սակայն բացառությամբ վաղուց մշակվող տեսակների) չի ենթարկվել որևէ խանգարումների և, որոնք փոփոխական չեն. բայց դրա փոխարեն հիբրիդների իրենց իսկ վեր արտադրողական սիստեմը լուրջ կերպով խանգարված է, և նրանց սերունդները շատ փոփոխական են։
Բայց վերադառնանք խառնացեղերի և հիբրիդների մեր համեմատությանը։ Գերտները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] մատնանշում է, որ խառնացեղերը ավելի հակված են վերադառնալու ծնողներից մեկի տիպին, քան հիբրիդները, բայց եթե նույնիսկ այդ ճիշտ է, ապա այստեղ տարբերությունը միայն աստիճանի մեջ է։ Բացի դրանից, Գերտները վճռականորեն պնդում է, որ երկար ժամանակ մշակման ենթարկվող բույսերից առաջացած հիբրիդներն ավելի հակում ունեն վերադառնալու ծնողական տիպին, քան վայրի կերպով աճող տեսակների հիբրիդները։ Հավանորեն դրանով էլ բացատրվում է զանազան դիտողների ստացած արդյունքներում եղած հետաքրքիր տարբերությունը. այսպես, օրինակ, Մաքս Վիխուրան[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 190] կասկածում է, որ հիբրիդները երբևիցե վերադառնում են ծնողական տիպին, բայց նա փորձեր կատարել է վայրի տեսակների վրա։ Մյուս կողմից, Նոդենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 19] խիստ վճռական արտահայտություններով պնդում է, որ հիբրիդների մեջ ծնողներին վերադառնալու հակումը գրեթե ընդհանուր կանոն է, բայց նա էլ փորձերը կատարել է գլխավորապես կուլտուրական բույսերի վրա։ Այնուհետև Գերտները պնդում է, որ եթե երկու, թեկուզ և միմյանց շատ մոտ տեսակներ խաչաձևվում են մի երրորդ տեսակի հետ, այդ դեպքում ստացվող հիբրիդները խիստ կերպով տարբերվում են միմյանցից. մինչդեռ եթե միևնույն տեսակին պատկանող մեկը մյուսից հեռավոր երկու տարատեսակներ խաչաձևվում են մի այլ տեսակի հետ, այդ դեպքում հիբրիդները միմյանցից խիստ կերպով չեն տարբերվում։ Բայց այդ եզրակացությունը, որքան ինձ հայտնի է, հիմնվում է միայն մի փորձի վրա, և թվում է, թե ուղղակի հակասում է Կյոլռեյտերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] մի քանի փորձերի արդյունքներին։
Ահա և այն բոլոր անկարևոր տարբերությունները բուսական հիբրիդների և խառնացեղերի միջև, որ կարողացել է մատնանշել Գերտները։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] Մյուս կողմից, իրենց ծնողների հետ խառնացեղերի և հիբրիդների, մանավանդ մոտ տեսակներից առաջացած հիբրիդների ունեցած նմանության աստիճանը և բնույթը ենթարկվում են միանման օրենքների։ Երկու տեսակների խաչաձևման դեպքում նրանցից մեկը երբեմն ունենում է հիբրիդին իր նմանությունը տալու ավելի մեծ ընդունակություն․ նույնը, ես կարծում եմ, հատուկ է նաև բուսական տարատեսակներին։ Կենդանիների մեջ մեկ տարատեսակի այղ գերակշռող ընդունակությունը մյուսի հանդեպ, անկասկած, նույնպես հաճախ է հանդիպում։ Փոխադարձ խաչաձևումից առաջացած հիբրիդային բույսերը սովորաբար միմյանց հետ շատ մոտ նմանություն են ունենում։ Նույնը կարելի է ասել նաև փոխադարձ խաչաձևումից ստացված խառնացեղ բույսերի մասին։ Ինչպես հիբրիդներին, այնպես էլ խառնացեղերին կարելի է վերադարձնել մաքուր ծնողական տիպին, նրանց հաջորդող սերունդները ենթարկելով կրկնողական խաչաձևումների ծնողական ձևերից մեկի հետ։
Այս բոլոր դիտողություններն անշուշտ կիրառելի են նաև կենդանիների նկատմամբ, բայց այդ դեպքում հարցը զգալիորեն բարդանում է մասամբ երկրորդային սեռական հատկանիշների գոյության հետևանքով, առանձնապես այն հանգամանքի հետևանքով, որ մի սեռ մյուսի հանդեպ առավելապես ընդունակ է իր նմանությանը ժառանգաբար փոխանցելու ինչպես մի տեսակը մի այլ տեսակի հետ խաչաձևման, այնպես էլ երկու տարատեսակների խաչաձևման դեպքում։ Այսպես, օրինակ, ես կարծում եմ, որ իրավացի են այն հեղինակները, որոնք պնդում են, թե էշը ձիու հանդեպ գերակշռող ուժ ունի, որի հետևանքով թե էշաջորին և թե ձիաջորին ավելի շատ նման են էշին, քան ձիուն. բայց այդ գերակշռությունն ավելի ուժեղ է արտահայտված արու էշի մեջ, քան էգ էշի, այնպես որ արու էշից և զամբիկից առաջացած էշաջորին ավելի շատ նման է էշին, քան էգ էշից և հովատակից առաջացած ձիաջորին։
Որոշ հեղինակներ մեծ նշանակություն են տվել այն ենթադրյալ փաստին, թե խառնացեղերը հաճախ մոտ նման են լինում ծնողներից միայն մեկին, այլ ոչ թե ունենում են միջակա հատկանիշներ երկուսի միջև։ Բայց այդ երբեմն պատահում է նաև հիբրիդների հետ, թեև, ես համաձայն եմ, շատ ավելի հազվադեպ, քան խառնացեղերի հետ։ Եթե մոտիկից ուշադրությամբ նայենք իմ հավաքած օրինակներին, երբ փոխադարձ խաչաձևումից առաջացած կենդանիները մոտ նմանություն են ունենում ծնողներից մեկի հետ, ապա դուրս է գալիս որ նմանությունը սահմանափակվում է գլխավորապես իրենց բնույթով համարյա այլանդակ և հանկարծակի երևան եկող հատկանիշներով, ինչպիսիք են ալբինիզմը, պոչի կամ եղջյուրների բացակայությունը, ավելորդ մատները առջևի և ետևի վերջավորությունների վրա և չի վերաբերում ընտրության միջոցով աստիճանաբար ձեռք բերված հատկանիշներին։ Ծնողներից մեկի մաքուր տիպին հանկարծակի կերպով վերադառնալու հակումը նույնպես պետք է անհամեմատ ավելի հաճախ հանդիպի խառնացեղերի մոտ, որոնք ծագում են հաճախ հանկարծակի առաջացող և կիսաայլանդակ բնույթ ունեցող տարատեսակներից, քան հիբրիդների մոտ, որոնք ծագում են առել դանդաղ ու բնական ճանապարհով առաջացած տեսակներից։ Ընդհանուր առմամբ ես լիովին համաձայն եմ դ-ր Պրոսպեր Լյուկի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 37] հետ, որը վերլուծելով կենդանիներին վերաբերող փաստերի ահագին քանակություն, գալիս է այն եզրակացության, որ ձագի և ծնողների նմանության օրենքները նույնն են, անկախ նրանից՝ թե ծնողները ավելի շատ կտարբերվեն միմյանցից, թե քիչ, այսինքն արդյոք կխաչաձևվե՞ն միևնույն տարատեսակի անհատներ, թե զանազան տարատեսակների կամ տարբեր տեսակների անհատներ։
Բացի պտղաբերության և անպտղաբերության հարցից, մյուս բոլոր տեսակետներից, ըստ երևույթին, գոյություն ունի ընդհանուր և մոտ նմանություն խաչաձևվող տեսակներից և խաչաձևվող տարատեսակներից առաջացած սերունդների միջև։ Եթե տեսակները դիտենք որպես ձևեր, որոնք ծագել են առանձին ստեղծագործական ակտերի հետևանքով, իսկ տարատեսակները որպես երկրորդային օրենքների շնորհիվ առաջացած ձևեր, ապա այդ նմանությունը կարող է զարմանք առաջացնել։ Բայց նա լիակատար ներդաշնակության մեջ է գտնվում այն տեսակետի հետ, որի համաձայն էական տարբերություն չկա տեսակների և տարատեսակների միջև։
ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ
Առանձին տեսակներ համարվելու համար միմյանցից բավականաչափ տարբերվող ձևերի միջև առաջին խաչաձևումները և նրանց հիբրիդները շատ հաճախ, բայց ոչ միշտ անպտղաբեր են լինում։ Այդ անպտղաբերությունը ներկայացնում է բազմապիսի աստիճաններ, և հաճախ այնքան աննշան է, որ ամենամանրազնին դիտողները եկել են տրամագծորեն հակառակ եզրակացությունների, դասակարգելով ձևերն այդ հատկանիշի հիման վրա։ Միևնույն տեսակին պատկանող տարբեր անհատների անպտղաբերությունը բնածին կերպով փոփոխական է և խիստ զգայուն է բարենպաստ և անբարենպաստ պայմանների նկատմամբ։ Պտղաբերության աստիճանը սիստեմատիկական ազգակցությունից սերտ կախման մեջ չի գտնվում, բայց որոշվում է զանազան հետաքրքրական ու բարդ օրենքներով։ Նա սովորաբար տարբեր է, իսկ երբեմն էլ շատ տարբեր է միևնույն տեսակների միջև կատարվող փոխադարձ խաչաձևումների դեպքում։ Նա իր աստիճանով միշտ միանման չէ առաջին խաչաձևման ժամանակ և նրանից առաջացած հիբրիդների մեջ։
Ճիշտ այնպես, ինչպես որ ծառերի պատվաստման դեպքում մի տեսակի կամ տարատեսակի մի ուրիշի հետ պատվաստելու ընդունակությունը հանդիսանում է նրանց վեգետատիվ սիստեմում եղած իրենց բնույթով անհայտ տարբերությունների կողմնակի արդյունք, այնպես էլ խաչաձևման ժամանակ մի տեսակի մի այլ տեսակի հետ միանալու ընդունակությունը հանդիսանում է նրանց վերարտադրողական սիստեմում եղած անհայտ տարբերություններին ուղեկցող արդյունք։ Ենթադրելու համար, թե տեսակներն օժտված են անպտղաբերության տարբեր աստիճաններով հատկապես նրա համար, որպեսզի կանխեն նրանց խաչաձևումը և միաձուլումը բնության մեջ, նույնքան քիչ հիմք կա, որքան որ կարծելու, թե ծառերն օժտված են զանազան և որոշ տեսակետներից փոխադարձաբար պատվաստելու անընդունակության համանման աստիճաններով նրա համար, որպեսզի կանխեն նրանց միակցումը մեր անտառներում։
Առաջին խաչաձևումների և նրանց հիբրիդային սերունդների անպտղաբերությունը ձեռք չի բերվել բնական ընտրության միջոցով։ Առաջին խաչաձևումների դեպքում նա ըստ երևույթին կախված է զանազան հանգամանքներից. որոշ դեպքերում պատճառ է ծառայում գլխավորապես սաղմի վաղաժամ մահը։ Հիբրիդների մոտ նա հավանորեն կախված է նրանից, որ նրանց ամբողջ կազմվածքը խանգարվել է երկու տարբեր ձևերի միացման հետևանքով։ Ըստ երևույթին նա մոտ է այն անպտղաբերությանը, որից այնքան հաճախ տուժում են մաքուր տեսակները, երբ նրանք ենթարկվում են նոր ու անբնական կենսապայմանների ներգործությանը։ Նա, ով կարող է բացատրել այս վերջին դեպքերը, կկարողանա բացատրել նաև հիբրիդների անպտղաբերությունը։ Այդ տեսակետն իրեն ուժեղ հենարան է գտնում նաև մի այլ համադրության մեջ, այն է՝ մի կողմից, կենսապայմանների թեթև փոփոխություններն ուժեղացնում են բոլոր օրգանական էակների ամրակազմությունը և պտղաբերությունը, իսկ մյուս կողմից, այն ձևերի խաչաձևումը, որոնք գտնվել են թեթևակի տարբերվող կենսապայմաններում կամ ձևափոխվեք են, բարենպաստ կերպով է ներգործում նրանց սերունդների մեծության, ուժի և պտղաբերության վրա։ Վերոհիշյալ փաստերը, որոնք վերաբերում են դիմորֆ և տրիմորֆ բուրերի ապօրինի միավորումների և նրանց ապօրինի սերունդների անպտղաբերությանը, գուցե հավանական են դարձնում այն, որ այդ բոլոր դեպքերում մի ինչ որ անհայտ կապ միացնում է առաջին միավորումների անպտղաբերության աստիճանը նրանց սերունդների անպտղաբերության աստիճանի հետ։ Դիմորֆիզմի այս փաստերի, ինչպես նաև փոխադարձ խաչաձևումների արդյունքների քննարկությունը պարզորոշ կերպով բերում են մեզ այն եզրակացության, որ խաչաձևվող տեսակների անպտղաբերության սկզբնական պատճառը գտնվում է միայն նրանց սեռական տարրերի տարբերություններում։ Մենք չգիտենք, երբ գործը վերաբերում է տարբեր տեuակներին, թե ինչու սեռական տարրերի այդ ավելի մեծ կամ փոքր ձևափոխությունները, որոնք առաջացնում են փոխադարձ անպտղաբերություն, այնքան սովորական երևույթ են դարձել, սակայն թվում է, թե դա բավական սերտ կապ ունի այն բանի հետ, որ տեսակները երկար ժամանակաշրջանների ընթացքում ենթարկվել են գրեթե միանման կենսապայմանների ազդեցության։
Զարմանալի չէ, որ որևէ երկու տեսակների խաչաձևման դժվարությունը և նրանց հիբրիդային սերունդների անպտղաբերությունը շատ դեպքերում համապատասխանում են միմյանց, թեև պայմանավորվում են զանազան պատճառներով, որովհետև թե մեկը և մյուսը կախված են խաչաձևվող տեսակների միջև եղած տարբերությունների ամբողջ գումարից։ Զարմանալի չէ նաև այն, որ առաջին խաչաձևման հեշտությունը, դրանից ստացվող հիբրիդների պտղաբերությունը և միմյանց վրա պատվաստվելու ընդունակությունը, թեև այս վերջին ընդունակությունը ակներևորեն կախված է խիստ բազմազան հանգամանքներից, մինչև մի որոշ սահման ընթանում են փորձի ենթարկվող ձևերի սիստեմատիկական ազգակցության հետ կողք-կողքի, որովհետև սիստեմատիկական ազգակցության հասկացողության մեջ մենք մտցնում ենք նմանության բազմապիսի գծեր։
Առաջին խաչաձևումները այնպիսի ձևերի միջև, որոնք հայտնապես պատկանում են տարատեսակների թվին կամ բավականաչափ նման են, որպեսզի համարվեն այդպիսիներ, և այդ խաչաձևումներից առաջացած խառնացեղերը մեծ մասամբ պտղաբեր են, սակայն ոչ միշտ, ինչպես այդքան հաճախ պնդել են։ Այդ համարյա ընդհանուր և լիակատար պտղաբերությունը նույնպես զարմանալի չէ, եթե հիշենք, թե ինչպես հեշտությամբ մենք ընկնում ենք կեղծ ապացույցների շրջանի մեջ բնական վիճակում գտնվող տարատեսակների հարցում, և եթե մենք հիշենք, որ տարատեսակների մեծ մասն ընտանի վիճակում առաջացել է զուտ արտաքին տարբերությունների ընտրության միջոցով և որ նրանք երկար ժամանակի ընթացքում չեն ենթարկվել միանման կենսապայմանների ազդեցությանը։ Առանձնապես չպետք է մոռանալ նաև այն, որ երկարատև ընտանի վիճակը առաջ է բերում անպտղաբերության վերացում և այդ պատճառով էլ քիչ հավանական է, որ նա է ծնել այդ իսկ հատկությունը։ Պտղաբերության հարցից անկախ, մյուս բոլոր տեսակետներից՝ թե նրանց փոփոխականության, թե հաճախակի խաչաձևումների միջոցով միմյանց կլանելու ընդունակության և թե երկու ծնողների ձևերի հատկանիշները ժառանգելու ընդունակության մեջ նկատվում է շատ մոտ ընդհանուր նմանություն հիբրիդների և խառնացեղերի միջև։ Վերջին հաշվով, թեև մենք շատ քիչ գիտենք նաև առաջին խաչաձևումների և հիբրիդների անպտղաբերության ճշգրիտ պատճառը, ինչպես նաև այն, թե ինչու իրենց բնական շրջադրությունից հեռացված կենդանիներն ու բույսերը դառնում են անպտղաբեր, սակայն այս գլխում բերված փաստերը, ինչպես ինձ թվում է, չեն հակասում այն համոզմանը, որ տեսակները սկզբնապես գոյություն են ունեցել որպես տարատեսակներ։
ԳԼՈՒԽ X։ ԵՐԿՐԱԲԱՆԱԿԱՆ ՏԱՐԵԳՐՈՒԹՅԱՆ ԱՆԼՐԻՎՈՒԹՅԱՆ ՄԱՍԻՆ
ՆԵՐԿԱՅՈՒՄՍ ՄԻՋԱԿԱ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԲԱՑԱԿԱՅՈՒԹՅԱՆ ՄԱՍԻՆ։— ՄԱՀԱՋՆՋՎԱԾ ՄԻՋԱԿԱ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԲՆՈՒՅԹԻ ՄԱՍԻՆ։— ՆՐԱՆՑ ԹՎԻ ՄԱՍԻՆ։— ԴԵՆՈՒԴԱՑԻԱՅԻ ԵՎ ՆՍՏՎԱԾՔՆԵՐԻ ԿՈՒՏԱԿՄԱՆ ԱՐԱԳՈՒԹՅԱՄԲ ԵՐԿՐԱԲԱՆԱԿԱՆ ԺԱՄԱՆԱԿԻ ՏԵՎՈՂՈՒԹՅՈՒՆԸ ՈՐՈՇԵԼՈՒ ՄԱՍԻՆ։— ԵՐԿՐԱԲԱՆԱԿԱՆ ԺԱՄԱՆԱԿԻ ՏԵՎՈՂՈՒԹՅՈՒՆԸ ՏԱՐԻՆԵՐՈՎ ՈՐՈՇԵԼՈՒ ՄԱՍԻՆ։— ՄԵՐ ՀՆԷԱԲԱՆԱԿԱՆ ՀԱՎԱՔԱԾՈՒՆԵՐԻ ԱՂՔԱՏՈՒԹՅԱՆ ՄԱՍԻՆ։— ԵՐԿՐԱԲԱՆԱԿԱՆ ՖՈՐՄԱՑԻԱՆԵՐՈՒՄ ԸՆԴՀԱՏՈՒՄՆԵՐԻ ՄԱՍԻՆ։— ԳՐԱՆԻՏԱՅԻՆ ՄԱՐԶԵՐԻ ՈՂՈՂՄԱՆ ՄԱՍԻՆ։— ՅՈՒՐԱՔԱՆՉՅՈՒՐ ՖՈՐՄԱՑԻԱՅՈՒՄ ՄԻՋԱԿԱ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԲԱՑԱԿԱՅՈՒԹՅԱՆ ՄԱՍԻՆ։— ՏԵՍԱԿԱՅԻՆ ԽՄԲԵՐԻ ՀԱՆԿԱՐԾԱԿԻ ԵՐԵՎԱՆ ԳԱԼՈՒ ՄԱՍԻՆ։— ԲՐԱԾՈՆԵՐ ՈՒՆԵՑՈՂ ՄԵԶ ՀԱՅՏՆԻ ՇԵՐՏԵՐԻՑ ԱՄԵՆԱՍՏՈՐԻՆՆԵՐՈՒՄ ՏԵՍԱԿԱՅԻՆ ԽՄԲԵՐԻ ՀԱՆԿԱՐԾԱԿԻ ԵՐԵՎԱՆ ԳԱԼՈՒ ՄԱՍԻՆ։— ԵՐԿՐԻ ՎՐԱ ԿՅԱՆՔԻ ՀՆՈՒԹՅԱՆ ՄԱՍԻՆ։
Վեցերորդ գլխում ես թվեցի գլխավոր առարկությունները, որոնք միանգամայն հիմնավորապես կարող էին արվել այս գրքում պաշտպանված տեսակետների դեմ։ Նրանց մեծամասնությունը մենք քննարկել ենք։ Այդ առարկություններից մեկը, այն է՝ տեսակային ձևերի առանձնացածության և նրանց միջև անթիվ շաղկապող օղակների բացակայության վրա մատնանշումը ակնհայտ դժվարություններ է ներկայացնում։ Ես նշել եմ, թե ինչու այդպիսի կապող օղակները սովորաբար չեն հանդիպում ներկայումս նրանց գոյության համար ըստ երևույթին շատ նպաստավոր հանգամանքներում, այսինքն՝ աստիճանաբար փոփոխվող ֆիզիկական պայմաններով մի լայնածավալ ու չընդհատվող մարզում։ Ես աշխատել եմ ցույց տալ, որ յուրաքանչյուր տեսակի կյանք ավելի մեծ կախման մեջ է արդեն հաստատված այլ օրգանական ձևերի ներկայությունից, քան կլիմայից, և որ այդ պատճառով, իսկապես ասած, կյանքի ղեկավարող պայմանները միանգամայն աստիճանաբար չեն փոփոխվում, այնպես, ինչպես փոփոխվում է ջերմաստիճանը կամ խոնավությունը։ Ես աշխատել են ցույց տալ նաև, որ միջակա տարատեսակները, որոնք սկզբում գոյություն են ունեցել ավելի քիչ թվով, քան այդ ձևերը, որոնց նրանք շաղկապում ենք հետզհետե դուրս են մղվում և բնաջնջվում, զուգընթաց տեղի ունեցող հետագա փոփոխության ու կատարելագործության։ Սակայն դրա պատճառը, որ այժմ բնության մեջ ամենուրեք չեն հանդիպում անհամար միջակա օղակներ, գտնվում է բնական ընտրության բուն պրոցեսում, որի ընթացքում նոր տարատեսակները միշտ դուրս են մղում իրենց նախածնողական ձևերին և գրավում նրանց տեղը։ Բայց չէ որ այդ դեպքում մի ժամանակ գոյություն ունեցած միջակա տարատեսակների քանակությունը պետք է իսկապես ահագին լինի և համապատասխանի այն հսկայական մասշտաբին, որով կատարվել է բնաջնջման պրոցեսը։ Հապա ինչո՞ւ այդ դեպքում յուրաքանչյուր երկրաբանական ֆորմացիա և յուրաքանչյուր շերտ լեփ լեցուն չէ այդպիսի միջակա օղակներով։ Իրոք, երկրաբանությունը մեր առաջ չի բացում օրգանիզմների մի այդպիսի լրիվ անընդմեջ շղթա, և գուցե դա է ամենաակնհայտ և լուրջ առարկությունը, որ կարող է արվել իմ տեսության դեմ։ Այդ հանգամանքի բացատրությունը, ինչպես ես կարծում եմ, երկրաբանական տարեգրության չափազանց անլրիվության մեջ է։
Ամենից առաջ հարկավոր է միշտ պարզորոշ կերպով պատկերացնել, թե ինչպիսի միջակա ձևեր, տեսության համաձայն, պետք է անցյալում գոյություն ունենային։ Երբ քննարկում ես որևէ երկու տեսակ, դժվարությամբ ես զսպում քեզ, որ երևակայությանդ մեջ չստեղծվեն միջակա ձևեր, որոնք անմիջականորեն շաղկապում են այդ երկու տեսակները իրար հետ։ Բայց դա բոլորովին սխալ տեսակետ է. մենք պետք է միշտ պատկերացնենք այնպիսի ձևեր, որոնք միջակա են երկու տեսակներից յուրաքանչյուրի և նրա ընդհանուր, բայց անհայտ նախահոր միջև, իսկ նախահայրը, իհարկե, պետք է որևէ բանով տարբերվի իր բոլոր փոփոխված հետնորդներից։ Ահա դրա պարզ լուսաբանությունը. սիրամարգաղավնին և փքունը՝ երկուսն էլ ծագում են առել ժայռաղավնուց․ եթե մենք ունենայինք մի ժամանակ գոյություն ունեցած բոլոր միջակա տարատեսակները, մենք կունենայինք անցումների մի բոլորովին անընդմիջվող շարք այդ ձևերից յուրաքանչյուրի և ժայռաղավնու միջև, բայց մենք չէինք ունենա տարատեսակներ, որոնք ուղղակի միջակա են սիրամարգաղավնու և փքունի միջև, չէինք ունենա, օրինակ, այնպիսի ձև, որն իր մեջ համատեղում է փոքր ինչ բացված պոչ և մի քիչ փքված ուռույց, այսինքն հենց նոր հիշված երկու ցեղերի բնորոշ հատկանիշները։ Բացի դրանից, այդ երկու ցեղերն այնքան են ձևափոխվել, որ եթե մենք չունենայինք պատմական կամ որևէ անուղղակի ապացույցներ նրանց ծագման մասին, մենք չէինք կարողանա որոշել, համեմատելով նրանց հատկանիշները ժայռաղավնու՝ C. livia-ի հատկանիշների հետ, թե արդյոք այդ ցեղերը նրանից են, թե մի որևէ մերձավոր այլ ձևից, օրինակ C. oenas-ից։
Նույնն է տեղի ունենում նաև բնական տեսակների հետ. եթե մենք վերցնենք երկու ձև, մեկը մյուսից հեռու կանգնած, օրինակ ձին և տապիրը, ոչ մի հիմք չունենք ենթադրելու, որ երբևիցե նրանց միջև միջակա օղակներ են եղել, բայց կարող ենք մտածել, որ միջակա օղակները գոյություն ունեցել են այդ երկու ձևերից յուրաքանչյուրի և նրանց ընդհանուր, բայց մեզ անհայտ նախահոր միջև։ Այդ ընդհանուր նախահայրը պետք է իր ամբողջ կազմվածքում ունենար շատ նմանություն թե տապիրի և թե ձիու հետ, բայց այղ կազմվածքի որոշ գծերով նա կարող էր զգալիորեն տարբերվել այդ երկու կենդանիներից, գուցե նույնիսկ ավելի շատ, քան նրանք տարբերվում են մեկը մյուսից։ Այդ պատճառով մենք բոլոր նման դեպքերում ի վիճակի չենք որոշելու որևէ երկու կամ ավելի մեծ թվով տեսակների նախածնողական ձևը, եթե նույնիսկ մանրամասնորեն համեմատենք նախածնողական ձևի և նրա փոփոխված հետնորդների կազմվածքը, մեզ այդ կհաջողվեր միմիայն այն դեպքում, եթե մենք հենց այդ նույն ժամանակ ունենայինք միջակա օղակների համարյա լրիվ շղթան։
Համաձայն իմ տեսության միանգամայն հնարավոր է, որ այժմ ապրող երկու ձևերից մեկն առաջացել է մյուսից, օրինակ՝ ձին տապիրից. այդ դեպքում նրանց միջև պետք է գոյություն ունենային ուղղակի միջակա օղակներ. բայց այդպիսի դեպքը ենթադրում է, որ ձևերից մեկը շատ երկար ժամանակաշրջանի ընթացքում մնացել է անփոփոխ, մինչդեռ նրա սերունդները փոփոխվել են շատ մեծ չափով, որը օրգանիզմների միջև, ծնողների և զավակների միջև տեղի ունեցող մրցության հետևանքով չափազանց հազվադեպ երևույթ կլիներ, որովհետև միշտ կյանքի նոր և ավելի կատարելագործված ձևերը ձգտում են դուրս մղել հին ու պակաս կատարելագործված ձևերին։
Բնական ընտրության տեսության համաձայն այժմ ապրող բոլոր տեսակները կապված են եղել յուրաքանչյուր սեռի նախածնողական տեսակի հետ այնպիսի ձևերի միջոցով, որոնք միմյանցից ավելի շատ չեն տարբերվել, քան ներկայումս տարբերվում են միևնույն տեսակի բնական ու ընտանի տարատեսակները, իսկ այդ նախածնողական տեսակները, որ այժմ շատ դեպքում մահաջնջված են, իրենց հերթին նույնպես կապված են եղել ավելի հին ձևերի հետ. ուրեմն, որքան հին են ձևերը, այնքան ավելի շատ կապված են եղել միմյանց հետ, շարունակ իրար հանդիպելով և մերձենալով յուրաքանչյուր մեծ դասի ընդհանուր նախահորը։ Այսպիսով, այժմ ապրող և հանգած բոլոր տեսակների միջև միջակա և անցումային օղակների թիվը պետք է անըմբռնելի չափով մեծ լիներ։ Եվ իհարկե, եթե միայն այս տեսությունը ճիշտ է, նրանք բոլորն էլ գոյություն են ունեցել երկրի վրա։
ԴԵՆՈՒԴԱՑԻԱՅԻ ՏԱՐԱԾԱԿԱՆՈՒԹՅԱՄԲ ԵՎ ՆՍՏՎԱԾՔՆԵՐԻ ԿՈՒՏԱԿՄԱՆ ԱՐԱԳՈՒԹՅԱՄԲ ԵՐԿՐԱԲԱՆԱԿԱՆ ԺԱՄԱՆԱԿԻ ՏԵՎՈՂՈՒԹՅՈՒՆԸ ՈՐՈՇԵԼՈՒ ՄԱՍԻՆ
Անկախ այն բանից, որ մենք չենք գտնում այդ անսահմանորեն բազմաթիվ շաղկապող ձևերի բրածո մնացորդները, թերևս կարելի է առարկել, թե ժամանակը չէր բավականացնի այդպիսի չափերով օրգանական աշխարհի փոփոխման համար, եթե ընդունենք, որ փոփոխությունները դանդաղորեն են կատարվել։ Ես գրեթե հնարավորություն չունեմ գործնականորեն երկրաբանությամբ չզբաղվող ընթերցողի ուշադրությունը դարձնելու այն փաստերի վրա, որոնք թույլ են տալիս գեթ թույլ գաղափար կազմելու անցած ժամանակի տևողության մասին։ Նա, ով կարդալով Չարլզ Լյաելի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] մեծ երկը երկրաբանության հիմունքների մասին, որն ապագա պատմաբանը կհամարի բնագիտության մեջ հեղաշրջում կատարած մի ստեղծագործություն, չի կամենա իրեն պատկերացնել անցած դարաշրջանների ժամանակի վիթխարիությունը, նա թող իսկույն փակի այս գիրքը։ Բայց ամենևին դեռ բավական չէ ուսումնասիրել երկրաբանության հիմունքները կամ կարդալ զանազան դիտողների հատուկ տրակտատները առանձին ֆորմացիաների մասին և տեսնել թե ինչպես յուրաքանչյուր հեղինակ ձգտում է մոտավոր հասկացողություն տալ յուրաքանչյուր ֆորմացիայի կամ նույնիսկ յուրաքանչյուր շերտի նստվածքի տևողության մասին, անցած ժամանակների տևողության մասին, որոշ հասկացողություն ստանալու համար հարկավոր է դիտել երկրաբանական գործոնների աշխատանքի բուն պրոցեսը, պետք է լավ իմանալ, թե ինչպիսի հզոր ստվարություններ քայքայվել ու ջրատարվել են ցամաքի մակերեսից և նստվածքների ինչպիսի զանգվածներ նստել։ Մեր նստվածքային ֆորմացիաների տարածությունը և հզորությունը ներկայացնում են, ինչպես իրավացիորեն նկատել է Լյաելը, այն ջրաողողման հետևանքն ու չափը, որին որևէ տեղ ենթարկվել է երկրի կեղևը։ Դրա համար էլ հարկավոր է սեփական աչքերով տեսնել մեկը մյուսի վրա կիտված շերտերի հսկայական ստվարությունը, դիտել պղտոր տիղմը դեպի ցած տանող առվակները, ծովեզրյա դարափերը մաշող ալիքները, եթե կուզենք որևէ գաղափար կազմել անցյալ ժամանակների տևողության մասին, որոնց հուշարձանները մենք տեսնում ենք ամենուրեք մեր շուրջը։
Ուսանելի է թափառել ոչ այնքան կարծր ապարներից կազմված ծովափի երկարությամբ և դիտել քայքայման պրոցեսը։ Մակընթացությունը շատ դեպքերում օրը երկու անգամ միայն կարճ ժամանակով հասնում է մինչև ժայռերը և ալիքները միայն այն ժամանակ են քանդում նրանց տակը, երբ իրենց հետ բերում են ավազ ու խճաքար, որովհետև մաքուր ջուրը, իհարկե, չի կարող մաշել ապարը։ Երբ վերջապես ժայռի հիմքը քանդված է, հսկայական բեկորները գլորվում են դեպի ցած. այստեղ, մնալով անշարժ, նրանք քայքայվում են ատոմ առ ատոմ, մինչև որ այնքան են փոքրանում, որ սկսում են ալիքների միջոցով գլորվել և այսպիսով ավելի արագ մանրանալ, վերածվելով խճաքարի, ավազի կամ տղմի։ Բայց ինչքան հաճախ ենք մենք տեսնում ժայռոտ դարափերի ստորոտում կլորացած քարաբեկորներ, որոնք ամեն կողմից պատած են ծովային օրգանիզմներից կազմված համատարած ծածկոցով, որը ցույց է տալիս, թե ինչքան դանդաղ է ընթանում մակընթացության քայքայիչ աշխատանքը և թե ինչքան հազվագյուտ դեպքում այդ բեկորները գլորվում են ալիքների ուժով։ Բացի դրանից, դիտելով քայքայման ենթարկվող ժայռոտ դարափերի գիծը մի քանի մղոն տարածությամբ, մենք կհամոզվենք, որ միայն տեղ-տեղ կարճ տարածության վրա, կամ որևէ կարկառի շուրջը կարելի է ներկայումս տեսնել առափնյա ժայռերի քայքայման պրոցեսը. իսկ մյուս տեղերում մակերևույթի տեսքից ու բուսականության բնույթից կարելի է եզրակացնել, որ շատ տարիներ են անցել այն ժամանակից, երբ ջուրն սկսել է քանդել ժայռերի հիմքը։
Սակայն վերջերս բազմաթիվ հիանալի դիտողների՝ Ռեմզեյի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 176] Ջյուքսի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 192] Գեյկիի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 193] Կրոլլի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 194] և այլոց հետազոտությունների շնորհիվ մենք իմացանք, որ մթնոլորտային գործոնների միջոցով ցամաքի մակերեսի քայքայումը պետք է համարել շատ ավելի կարևոր, քան ծովափի քայքայումը ծովի ալիքներից։ Ցամաքի ամբողջ մակերևույթը ենթարկվում է մթնոլորտի և լուծված ածխաթթու պարունակող անձրևաջրի քիմիական ներգործությանը, իսկ ցուրտ երկրներում նաև սառնամանիքի ազդեցությանը, այդ աշխատանքի հետևանքով քայքայված նյութը ուժեղ անձրևի միջոցով ցած է տարվում նույնիսկ թույլ թեքված լանջերով, ինչպես նաև քամու միջոցով, մանավանդ չոր մարգերում, քշվում է ավելի մեծ չափով, քան սովորաբար ենթադրում են. այնուհետև քայքայված նյութը տեղափոխվում է հեղեղների և գետերի միջոցով, որոնք, եթե արագահոս են, խորացնում են իրենց հունը և մանրացնում բեկորները։ Անձրևային օրը, նույնիսկ թույլ ալիքաձև մակերևույթ ունեցող վայրում, մենք տեսնում ենք յուրաքանչյուր լանջից հոսող պղտոր առվակներում մթնոլորտի քայքայիչ աշխատանքը։ Ռեմզեյը և Ույտիկերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 195] ցույց են տվել,— և դա շատ ուշագրավ դիտողություն է,— որ Վելդյան մարզում, ինչպես նաև Անգլիայի լայնքով անցնող ժայռերի շղթաները հին ծովափ չեն եղել, ինչպես առաջները համարում էին դրանց, որովհետև ժայռերի շղթաներից յուրաքանչյուրը կազմված է միևնույն ֆորմացիայից, մինչդեռ մեր առափնյա ժայռերը ամենուրեք կազմված են մեկը մյուսին հաջորդող զանազան ֆորմացիաներից։ Նկատի առնելով այդ, մենք պետք է ընդունենք, որ հիշված ժայռերն իրենց ծագմամբ գլխավորապես պարտական են այն հանգամանքին, որ ժայռերը կազմող ապարները ավելի լավ են դիմադրել մթնոլորտի քայքայիչ աշխատանքին, քան հարակից մակերեսը, դրա հետևանքով վերջինս հետզհետե ցածրացել է, թողնելով ավելի կարծր ապարներից կազմված շղթաների ցցվածքները։ Ոչ մի բան մեր գիտակցության վրա ավելի խոր տպավորություն չի թողնում անցած ժամանակի տևողության մասին, եթե այդ ժամանակը հարմարեցնենք ժամանակի մասին մեր ունեցած հասկացողությանը, ինչպես այս եղանակով կազմված համոզմունքը, որ մթնոլորտի ազդեցությունը, որն ուժով այնքան չնչին է թվում և գործում է այնպես դանդաղ, տվել է այնքան խոշոր հետևանքներ։
Այսպիսով, ստանալով հասկացողություն այն դանդաղության մասին, որով ցամաքը քայքայվում է մթնոլորտի աշխատանքներից և ալիքների ափակոծությունից, անցած ժամանակի տևողությունը գնահատելու համար ուսանելի է մի կողմից՝ խորհել ապարի այն զանգվածների մասին, որոնք հեռացվել են շատ ընդարձակ մարզերի մակերեսից և մյուս կողմից՝ մեր նստվածքային ֆորմացիաների հաստության մասին։ Ես հիշում եմ, թե ինչպես ուժեղ կերպով զարմացել էի ես հրաբխային կղզիների տեսքից, որոնց ափերը, քայքայվելով ալեբախությունից, ընդհատվում էին ուղղաբերձ ժայռերով հազար կամ երկու հազար ոտնաչափ բարձրությամբ. այդ ավելի ևս զարմանալի էր, որ լավայի հեղեղների թույլ թեքությունը, որ պայմանավորված է նրա նախկին հեղուկ վիճակով, առաջին հայացքից ցույց էր տալիս, թե կարծր քարային շերտերը մի ժամանակ որքան խորն են մտած եղել բաց օվկիանոսի մեջ։ Նույնպիսի պատմություն, բայց ավելի ևս հասկանալի լեզվով, մեզ պատմում են սահքերը՝ այդ մեծ բեկվածքները, որոնց երկարությամբ շերտերը բարձրացել են մի կողմի վրա և իջել մյուս կողմի վրա հազարավոր ոտնաչափ բարձրությամբ կամ խորությամբ. որովհետև այն ժամանակից սկսած, երբ ճեղքվել է երկրի կեղևը և տեղի է ունեցել շերտերի այդ տեղաշարժը կամ հանկարծակի կամ, ինչպես այժմ կարծում է երկրաբանների մեծամասնությունը, դանդաղ կերպով և կատարվել է մի քանի նվագներով, երկրի մակերևույթը այն աստիճան է հարթվել, որ այժմ արտաքուստ այդ հսկայական տեղաշարժերի ոչ մի հետք չի երևում։ Պեննինյան սահքը, օրինակ, ունի 30 մղոն երկարություն, և այդ տարածության վրա շերտերի ուղղաձիգ տեղաշարժը տատանվում է 600-ից մինչև 3000 ոտնաչափի սահմաններում։ Պրոֆ. Ռեմզեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 176] նկարագրել է Էնգլեսիի սահքը, որն ունի շերտերի 2300 ոտնաչափ տեղաշարժ, և նա էլ հաղորդում է ինձ, որ ինքը միանգամայն համոզված է, որ Մերիոնետշիրում կա 12000 ոտնաչափ բարձրությամբ տեղաշարժ. սակայն հիշատակված դեպքերում երկրի մակերեսի վրա չկա ոչ մի բան, որ ցույց տար այդքան վիթխարի տեղաշարժերը, որովհետև բեկվածքի երկու կողմում ապարների շերտերը հետագայում հարթվել են, մակարդակով հավասարվել։
Մյուս կողմից աշխարհի բոլոր մասերում նստվածքային ապարների շերտերն ունեն զարմանալի մեծ հաստություն։ Կորդիլերներում ես տեսել եմ կոնգլոմերատի մի զանգված 10000 ոտնաչափ հաստությամբ, թեև կոնգլոմերատները ամենայն հավանականությամբ կուտակվել են ավելի արագ, քան ավելի մանրահատիկ մյուս նստվածքները, սակայն կազմված լինելով մաշված ու կլորացած ճալաքարերից, որոնցից յուրաքանչյուրն իր վրա կրում է ժամանակի կնիքը, նրանք կարող են ակնառու ապացույց ծառայել, թե ինչքան դանդաղորեն պետք է կուտակված լինի նրանց զանգվածը։ Պրոֆ. Ռեմզեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 176] հաղորդել է ինձ, թե ինչպիսի առավելագույն հաստություն ունեն հաջորդական ֆորմացիաները Մեծ Բրիտանիայի զանազան մասերում, որ որոշվել է մեծ մասամբ անմիջական չափումով, ահա այդ չափումների արդյունքները.
| ոտնաչափ | |
| Պալեոզոյան շերտեր (չհաշված հրաբխային ապարները) | 57 154 |
| Երկրորդային շերտեր | 13 190 |
| Երրորդային շերտեր | 2 140 |
Այդ բոլորը միասին կազմում է 72584 ոտնաչափ, այսինքն մոտավորապես տասներեք և երեք քառորդ անգլիական մղոն [23,6 կմ]։ Անգլիայում բարակ շերտերի ձևով զարգացած որոշ ֆորմացիաներ մայր ցամաքի վրա ներկայացնում են հազարավոր ոտնաչափ հզորությամբ շերտեր։ Բացի դրանից, երկրաբանների մեծամասնության կարծիքի համաձայն՝ յուրաքանչյուր հաջորդական ֆորմացիայի միջև գոյություն ունեն հսկայական տևողությամբ ընդմիջումներ, այնպես որ Անգլիայի նստվածքային կուտակումների հսկայական շերտերի հաստությունը միայն մոտավոր գաղափար է տալիս այն ժամանակի մասին, որի ընթացքում նրանք կուտակվել են։ Այդ բոլորի մասին խորհելը մտքի վրա նույնպիսի տպավորություն է թողնում, ինչպես որ հավիտենականության մասին պարզ պատկերացում կազմելու ապարդյուն փորձերը։
Սակայն այդ տպավորությունն այնքան էլ ճիշտ չէ։ Մ-ր Կրոլլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 194] մի հետաքրքրական հոդվածում նշում է, որ մենք սխալվում ենք ոչ այն ժամանակ, «երբ շատ չափազանցված հասկացողություն ենք կազմում երկրաբանական ժամանակաշրջանների տևողության մասին», այլ այն ժամանակ, երբ նրանց գնահատում ենք տարիներով։ Երբ երկրաբանը իր ուշադրությունը սևեռում է ընդարձակ ու բարդ երևույթների վրա և հետո նայում է մի քանի միլիոն տարի արտահայտող թվերի վրա, երկու դեպքերը բոլորովին տարբեր տպավորություն են առաջացնում, և թվերն առաջին հայացքից չափազանց անբավարար են թվում։ Հաշվելով մի քանի գետերի բերած նստվածքի որոշ քանակությունը, մ-ր Կրոլլը ապացուցում է, որ 1000 ոտնաչափ հաստությամբ միջին մակարդակով ամբողջ տարածությունից պինդ ապարի շերտի աստիճանական փխրեցման ու հեռացման համար հարկավոր է վեց միլիոն տարի։ Այդ եզրակացությունը զարմանալի է թվում, և որոշ նկատառումներ մեզ դրդում են ենթադրելու, թե արդյոք շատ չի չափազանցված այդ թիվը, բայց եթե նույնիսկ այն պակասեցնենք երկու կամ չորս անգամ, այնուամենայնիվ ապշեցուցիչ հետևանք է մնում։ Մեզանից քչերը թերևս գիտեն, թե իրականում ինչ է ներկայացնում միլիոնը, մ-ր Կրոլլը դրան տալիս է հետևյալ լուսաբանությունը. վերցրեք թղթի մի նեղ շերտ 83 ոտնաչափ և 4 դյույմ [25,7 մ] երկարությամբ և ձգեք մի մեծ դահլիճի պատի երկարությամբ, դրանից հետո չափեք այդ ժապավենի մի ծայրից դյույմի մի տասներորդ մասը [0,4 մմ]։ Դյույմի այդ մեկ տասներորդ մասը կլինի հարյուր տարի, իսկ ամբողջ ժապավենը՝ միլիոն տարի։ Բայց պետք է մեզ լավ պատկերացնենք, թե այս աշխատության խնդրի տեսակետից ինչ են պարփակում իրենց մեջ այդ հարյուր տարիները, որոնք պատկերացված են վերոհիշյալ մեծության դահլիճի մեջ այնքան ծայրահեղ չնչին չափով։ Նոր ցեղեր առաջացնելու մի քանի ականավոր սիրահարներ իրենց կյանքի ընթացքում այն աստիճան են փոփոխել որոշ բարձրակարգ կենդանիների, որոնք բազմանում են ավելի դանդաղ, քան ստորակարգ կենդանիների մեծ մասը, որ նրանք առաջացրել են այն, ինչ կարելի է անվանել նոր ենթացեղ։ Քչերը, հարկ եղած ջանասիրությամբ զբաղվելով որևէ ցեղի ստեղծմամբ, դրա վրա ծախսել են կես դարից ավելի, այնպես որ մեկ դարն արտահայտում է նոր ցեղերի երկու այդպիսի սիրահարների հաջորդական աշխատանքը։ Չի կարելի ենթադրել, որ տեսակները բնական վիճակում կարող էին փոփոխվել այնպես արագ, ինչպես փոփոխվում են ընտանի կենդանիները մեթոդական ընտրության միջոցով։ Համենայն դեպս այդ պրոցեսը կարելի է ավելի մոտ նմանեցնել անգիտակցական ընտրության ներգործությանը, այսինքն առանց ցեղը փոփոխելու մտադրության ամենաօգտակար կամ ամենագեղեցիկ կենդանիներ պահելուն, իսկ այդպիսի անգիտակցական ընտրության միջոցով որոշ տեսակներ նկատելի չափով փոփոխվել են երկու կամ երեք դարերի ընթացքում։
Տեսակները հավանորեն անհամեմատ ավելի դանդաղորեն են փոփոխվում, և միևնույն երկրում միայն քչերն են փոփոխվում միաժամանակ։ Այդ դանդաղությունը նրանից է առաջանում, որ միևնույն երկրի բոլոր բնակիչներն արդեն այնպես լավ հարմարված են միմյանց հետ, որ բնության տնտեսության մեջ նոր տեղեր կարող են բացվել միայն երկար ժամանակամիջոցներ անցնելուց հետո՝ որևէ ֆիզիկական փոփոխությունների ազդեցության տակ կամ երկրի մեջ նոր ձևերի ներգաղթի հետևանքով։ Բացի դրանից, ճիշտ ուղղությամբ փոփոխությունները կամ շեղումները, որոնց շնորհիվ բնակիչներից մի քանիսը կարող էին ավելի լավ, հարմարված լինել փոփոխված պայմաններում իրենց նոր տեղերին, կարող են անմիջապես չառաջանալ։ Դժբախտաբար մենք միջոցներ չունենք տարիների թվով որոշելու, թե ինչ ժամանակաշրջան է հարկավոր, որպեսզի տեսակը կարողանար փոփոխվել։ Բայց մենք հետո կրկին կվերադառնանք ժամանակի հարցին։
ՄԵՐ ՀՆԷԱԲԱՆԱԿԱՆ ՀԱՎԱՔԱԾՈՒՆԵՐԻ ԱՂՔԱՏՈՒԹՅԱՆ ՄԱՍԻՆ
Այժմ դառնանք մենք ամենահարուստ երկրաբանական թանգարաններին։ Ի՜նչ խղճուկ պատկեր են ներկայացնում նրանք։ Որ մեր հավաքածուները թերի են, դրան համաձայն են բոլորը։ Երբեք չպետք է մոռանալ Էդվարդ Ֆորբսի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] այդ զարմանալի հնէաբանի դիտողությունները, որ բրածո շատ տեսակներ որոշվել ու անվանվել են միակ և հաճախ ոչ լրիվ օրինակով կամ սակավաթիվ օրինակներով, որոնք հավաքված են փոքր տարածությունից։ Երկրի մակերեսի միայն մի փոքր մասն է ենթարկվել երկրաբանական հետազոտության, և ոչ մի վայր բավականաչափ լրիվությամբ հետազոտված չէ, որն ապացուցվում է այն կարևոր հայտնագործություններով, որոնք արվում են ամեն տարի Եվրոպայում։ Բոլորովին կակուղ օրգանիզմները ամենևին չեն կարող պահպանվել։ Խեցիներն ու ոսկորները քայքայվում են ու անհետանում, եթե մնում են ծովի հատակին այնպիսի տեղերում, որտեղ նստվածքներ չեն կուտակվում։ Մենք հավանորեն խիստ սխալվում ենք, եթե կարծում ենք, թե նստվածքները նստում են ծովի համարյա ամբողջ հատակի վրա և այնքան արագ կերպով, որ բրածո մնացորդները կարող էին ծածկվել ու պահպանվել։ Օվկիանոսի հսկայական տարածության վրա ջրի վառ կապույտ գույնը վկայում է նրա մաքրության մասին։ Հայտնի են շատ դեպքեր, որ ցույց են տալիս այնպիսի երկրաբանական ֆորմացիաներ, որոնք հսկայական տևողությամբ ընդմիջումից հետո համապատասխանորեն ծածկվել են մի այլ, ավելի ուշ ֆորմացիայով, ըստ որում ստորին շերտը այդ ժամանակամիջոցում քայքայման չի ենթարկվել. ըստ երևույթին, այդ կարելի է բացատրել միայն նրանով, որ ծովի հատակը հաճախ երկար ժամանակ մնում է անփոփոխ դրության մեջ։ Ավազի կամ ծովակոպիճի վրա ընկնող մնացորդները ծովի մակարդակից շերտերի դուրս գալուց հետո սովորաբար լուծվում են ածխաթթու պարունակող անձրևաջրի ներթափանցման շնորհիվ։ Այն բազմաթիվ կենդանիներից, որոնք բնակվում են ծովափերում մակընթացության և տեղատվության մակարդակի միջև, քչերն, ըստ երևույթին, պահպանվում են միայն հազվագյուտ դեպքում։ Օրինակ Chthamalinae (նստակյաց փնջոտնյա խեցգետինների ենթաընտանիքը) շատ տեսակները ամենուրեք անհամար բազմությամբ ծածկում են առափնյա ժայռերը. նրանք բոլորն էլ իսկական ծովափնյա կենդանիներ են, բացի մի միջերկրածովային տեսակից, որն ապրում է մեծ խորություններում և այդ տեսակը գտնվել է Սիցիլիայում բրածո վիճակում, մինչդեռ մինչև այժմ ոչ մի այլ տեսակ գտնված չէ երրորդային սիստեմի նստվածքներում, թեև հայտնի է, որ Chthamalus սեռը գոյություն է ունեցել կավճային ժամանակաշրջանում։ Վերջապես, նստվածքների հզոր շերտերից շատերը, որոնք իրենց կուտակման համար պահանջել են շատ երկար ժամանակ, բոլորովին զուրկ են օրգանական մնացորդներից, և այդ երեվույթի պատճառը մեզ համար անհասկանալի է մնում. դրա ամենազարմանալի օրինակներից մեկն է ներկայացնում ֆլիշ-ֆորմացիան, որը կազմված է մի քանի հազար, տեղ-տեղ մինչև վեց հազար ոտնաչափ հաստությամբ թերթաքարերից ու ավազաքարերից և ձգվում է առնվազն 300 մղոն տարածության վրա՝ Վիենայից մինչև Շվեյցիարիա. այդ ֆորմացիայում, չնայած ամենամանրազնին որոնումներին, ոչ մի բրածո գտնված չէ, բացի բուսական փոքրաթիվ մնացորդներից։
Ինչ վերաբերում է այն օրգանիզմներին, որոնք ապրել են մայր ցամաքների վրա մեզոզոյան և պալեոզոյան ժամանակաշրջաններում, խոսք չկա, որ մեր տեղեկությունները նրանց մասին վերին աստիճանի կցկտուր են։ Բավական է ասել, որ մինչև վերջին ժամանակները գտնված չէ ոչ մի ցամաքային խեցի, որը պատկաներ այդ հսկայական ժամանակաշրջաններից որևէ մեկին, եթե չհաշվենք մի տեսակը, որ հայտնաբերել են Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] և Դոուսոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 196] Հյուսիսային Ամերիկայի քարածխային շերտերում. բայց այժմ խեցիներ գտնված են լեյասում[65]։ Կաթնասունների մնացորդների վերաբերմամբ Լյաելի ձեռնարկում բերված պատմական աղյուսակի վրա մի հայացք գցելն ավելի լավ կապացուցի, քան մանրամասնությունների ամբողջ էջեր, թե որքան պատահական ու հազվագյուտ դեպքերում են պահպանվում նրանք։ Եվ նրանց հազվագյուտ լինելը զարմանալի չէ, եթե հիշենք այն մասին, թե երրորդային կաթնասունների ինչքան շատ ոսկորներ են գտնվել կամ այրերում և կամ լճային նստվածքներում, և այն մասին, որ ոչ մեզոզոյան և ոչ էլ պալեոզոյան գոյացումների մեջ մեզ հայտնի չէ ոչ մի քարայր և ոչ մի լճային նստվածք։
Սակայն երկրաբանական տարեգրության անլրիվությունը մեծ չափով կախված է մի այլ և ավելի կարևոր պատճառից, քան բոլոր վերոհիշյալները, այսինքն նրանից, որ տարբեր ֆորմացիաները մեկը մյուսից բաժանված են մեծ ժամանակամիջոցներով։ Այդ կարծիքը ջերմ կերպով պաշտպանել են շատ երկրաբաններ ու հնէաբաններ, որոնք Է. Ֆորբսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] նման բոլորովին չեն հավատում տեսակների փոփոխելիությանը։ Երբ մենք դիտում ենք երկրաբանական աշխատություններում աղյուսակների ձևով դասավորված ֆորմացիաների շարքը, կամ երբ նրանց հետազոտում ենք բնության մեջ, մեզ դժվար է հրաժարվել այն մտքից, որ նրանք առանց ընդհատման հաջորդում են մեկը մյուսի հետևից։ Բայց մենք գիտենք, օրինակ, Ռուսաստանի մասին Մուրչիսոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 197] նշանավոր աշխատությունից, թե ինչպիսի ազատ տեղեր գոյություն ունեն այդ երկրում մեկը մյուսի վրա ընկած ֆորմացիաների միջև, նույնը հայտնի է Հյուսիսային Ամերիկայի և շատ այլ երկրների մասին։ Եվ ամենափորձված երկրաբանը, եթե կենտրոնացներ իր ուշադրությունը բացառապես այդ մեծ մարզերի վրա, երբեք չէր կռահի, որ այն ժամանակաշրջանները, որոնցից ոչ մի հուշարձան չի մնացել իր սեփական երկրում, կուտակել են այլ վայրերում նստվածքների հզոր շերտեր, որոնք իրենց մեջ պարփակում են կյանքի նոր և ինքնօրինակ ձևեր։ Եվ եթե յուրաքանչյուր առանձին երկրում համարյա ոչ մի գաղափար չի կարելի կազմել միմյանց հաջորդող ֆորմացիաների միջև անցած ժամանակի տևողության մասին, ապա դրանից կարելի է եզրակացնել, որ ոչ մի տեղ էլ չի կարելի որոշել այդ։ Մեկը մյուսին հաջորդող ֆորմացիաների հանքաբանական կազմում հաճախակի և մեծ փոփոխությունները սովորաբար ցույց են տալիս, որ զգալի փոփոխություններ են տեղի ունեցել նստվածքների համար նյութ մատակարարող շրջապատի երկրների աշխարհագրության մեջ, և հաստատում են այն կարծիքը, որ ամեն մի ֆորմացիայի միջև մեծ ժամանակամիջոցներ են անցել։
Ինձ թվում է, որ դժվար չէ ըմբռնել, թե ինչու յուրաքանչյուր երկրի երկրաբանական ֆորմացիաները գրեթե միշտ լինում են ընդմիջվող, այսինքն մեկը մյուսին չեն հաջորդել առանց ընդհատման։ Երբ ես հետազոտում էի Հարավային Ամերիկայի ափը բազմահարյուր մղոններ տարածությամբ, որը արդի ժամանակաշրջանի ընթացքում մի քանի հարյուր ոտնաչափ բարձրացել է, հազիվ թե որևէ փաստ ինձ ավելի զարմացներ, քան որևէ նոր, այնքան խոշոր նստվածքի բացակայությունը, որից մի բան պահպանված մնար նույնիսկ կարճատև երկրաբանական ժամանակաշրջանում։ Արևմտյան ամբողջ ափի երկարությամբ, որտեղ ապրում են յուրահատուկ ծովային կենդանիներ, երրորդային շերտերն այնքան թույլ են զարգացած, որ այստեղ մեկը մյուսին հաջորդող զանազան յուրահատուկ կենդանիներից մինչև հեռավոր ժամանակները չէր պահպանվի ոչ մի հուշարձան։ Մի փոքր խորհրդածումը կբացատրի մեզ, թե ինչու Հարավային Ամերիկայի բարձրացող արևմտյան ափի երկարությամբ չի կարելի գտնել նոր կամ երրորդային մնացորդներ ունեցող զգալի ֆորմացիաներ, չնայած այն բանին, որ դարերի ընթացքում բերվող նստվածքի քանակությունը պետք է շատ մեծ լիներ՝ առափնյա ժայռերի արագ քայքայման և ծովը թափվող պղտոր հեղեղների շնորհիվ։ Անկասկած այդ բացատրվում է նրանով, որ առափնյա և ենթաառափնյա գոտու նստվածքները շարունակ ողողվում են, հենց որ նրանք երկրի դանդաղ ու աստիճանական բարձրացման հետևանքով մտնում են ափակոծության քայքայիչ ներգործության շրջանի մեջ։
Թվում է, թե այստեղից կարելի է եզրակացնել, որ նստվածքի միայն չափազանց հզոր պինդ և հսկայական զանգվածները կարող են դիմանալ ալեբախության չդադարող ներգործությանը իրենց առաջին բարձրացման ու ծովի մակարդակի հետագա փոփոխությունների ժամանակ, ինչպես նաև մթնոլորտի քայքայիչ աշխատանքին։ Նստվածքի այդպիսի հզոր ու ընդարձակ կուտակումներ կարող են առաջանալ երկու ձևով. կամ ծովի խոր անդունդներում և այս դեպքում ծովի հատակն այնպես հարուստ կերպով բնակված չի լինում կյանքի բազմազան ձևերով, ինչպես լինում է ավելի ծանծաղ ծովը, և նստվածքի զանգվածը բարձրանալուց հետո միայն ոչ լրիվ գաղափար կտա այն օրգանիզմների մասին, որոնք նրա կուտակման ժամանակաշրջանում ապրել են ծովի ամենամոտ շրջաններում. կամ որևէ հզորությամբ և տարածությամբ նստվածք կարող է կուտակվել ծովի դանդաղորեն իջնող հատակին։ Այս վերջին դեպքում, քանի դեռ հատակի ցածրացումը և նստվածքի կուտակումը գրեթե հավասարակշռում են միմյանց, ծովը պետք է մնա ծանծաղ և ներկայացնի բարենպաստ պայմաններ շատ ու բազմազան ձևերի համար և այսպիսով կարող է առաջանալ մի ֆորմացիա, որը հարուստ է բրածոներով և բավականաչափ հզոր է, որպեսզի դիմանա այն քայքայիչ ազդեցություններին, որոնց նա կենթարկվի, երբ կբարձրանա ծովի մակարդակից վեր։
Ես համոզված եմ, որ համարյա մեր բոլոր հնագույն ֆորմացիաները, որոնց մեջ գերակշռում են բրածոներով հարուստ շերտերը, այդպես են առաջացել հատակի ցածրացման դեպքում։ Այդ խնդրի մասին իմ տեսակետների հրապարակման ժամանակից՝ 1845 թ. սկսած ես անդադար հետևել եմ երկրաբանության հաջողություններին և զարմանքով նկատել եմ, թե ինչպես մի հեղինակ մյուսի հետևից խոսելով երբեմն մի, երբեմն մի այլ մեծ ֆորմացիայի մասին, եկել են այն եզրակացության, որ նա նստել է ծովի հատակի ցածրացման ժամանակ։ Ես դրան կարող եմ ավելացնել, որ միայն մեկ երրորդային հին ֆորմացիա, Հարավային Ամերիկայի արևմտյան ափին, այնքան մեծ շերտ է ներկայացնում, որ նա դիմացել է քայքայմանը, որին մինչև այժմ էլ ենթարկվել է, և այդ ֆորմացիան կուտակվել է ցածրացման դարաշրջանում և այդ պատճառով ունի զգալի հզորություն, սակայն նա էլ հազիվ թե պահպանվի մինչև ապագա մի քիչ հեռավոր երկրաբանական դարաշրջանը։
Բոլոր երկրաբանական փաստերը պարզ կերպով մեզ ցույց են տալիս, որ երկրագնդի յուրաքանչյուր մարզ մակերևույթի բազմաթիվ դանդաղ տատանումներ է կրել, և, ըստ երևույթին, այդ տատանումներն ընդգրկել են մեծ տարածություններ։ Հետևաբար, բրածոներով հարուստ և բավականաչափ հզոր ու ընդարձակ ֆորմացիաները, որպեսզի պահպանվեին հետագա քայքայումից, կարող էին ցածրացման դարաշրջանում գոյանալ ընդարձակ մարզերում, բայց միայն այնտեղ, որտեղ բերվող նստվածքը բավական էր, որպեսզի ծովի խորությունը շարունակեր մնալ չնչին և որպեսզի մնացորդները ծածկվեին ու պահպանվեին մինչև իրենց քայքայումը։ Մյուս կողմից, քանի դեռ ծովի հատակը մնում է անշարժ, նստվածքի հզոր շերտեր չեն կարող կուտակվել ծանծաղաջուր մարզերում, որոնք ամենից ավելի նպաստավոր են կյանքի համար։ Ավելի ևս քիչ հնարավոր է այդ բարձրացման միջակա դարաշրջաններում, կամ, ավելի ճիշտ ասած, այդ ժամանակ նստած շերտերը պետք է մեծ մասամբ ենթարկվեին քայքայման, իրենց բարձրացման ու ծովի ափակոծության ներգործության ոլորտի մեջ մտնելուն զուգընթաց։
Այդ դիտողությունները վերաբերում են գլխավորապես առափնյա և ենթաառափնյա նստվածքներին։ Ինչ վերաբերում է ընդարձակ, բայց ծանծաղ ծովին, ինչպիսին, օրինակ, գոյություն ունի Մալայան արշիպելագի մեծ մասի սահմաններում, որտեղ խորությունը փոփոխվում է 30 կամ 40-ից մինչև 60 ֆաթոմ[66], այնտեղ կարող է բարձրացման դարաշրջանում առաջանալ լայնածավալ տարածությամբ մի ֆորմացիա, և նա այնքան շատ չի տուժի իր դանդաղ բարձրացման ժամանակ, բայց այդպիսի ֆորմացիայի հզորությունը չի կարող մեծ լինել, որովհետև հատակի բարձրացող շարժման պատճառով նա ավելի փոքր կլինի, քան այն խորությունը, որի մեջ տեղի է ունեցել կուտակումը. նստվածքը նաև շատ պնդացած չի լինի և ծածկված չի լինի վերևից նոր ֆորմացիայով, այնպես որ շատ հավանականություններ կան, որ նա կողողվի ու կտարվի մթնոլորտի և ծովի ալեբախության քայքայիչ աշխատանքի հետևանքով՝ մակարդակի հետագա տատանումների ժամանակ։ Սակայն, մ-ր Հոպկինսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 198] այն կարծիքն է հայտնել, որ եթե մարզի մի որոշ մասը բարձրանում է և նախքան դենուդացիայի ենթարկվելը կսկսի նորից իջնել, այդ դեպքում բարձրացման դարաշրջանում առաջացած նստվածքների թեկուզև աննշան շերտը կարող է հետո ծածկվել նոր նստվածքներով և, այսպիսով, պահպանվել երկար ժամանակ։
Մ-ր Հոպկինսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 198] նաև այն կարծիքն է հայտնում, որ հորիզոնական զգալի տարածություն գրավող նստվածքային շերտերը միայն հազվագյուտ դեպքերում են վերջնականապես քայքայվել։ Բայց բոլոր երկրաբանները, բացառությամբ այն քչերի, որոնք ընդունում են, թե մեր այժմյան ձևափոխված թերթաքարերը և պլուտոնական ապարները մի ժամանակ կազմել են երկրի սկզբնական միջուկը, կհամաձայնվեն, որ այդ ապարների մակերևույթից կտրված են նրանց մի ժամանակ ծածկող հսկայական շերտեր․ որովհետև հազիվ թե հնարավոր է ընդունել, որ նման ապարները կարող էին կարծրանալ և բյուրեղանալ, մնալով բաց. սակայն եթե ձևափոխումը տեղի է ունեցել օվկիանոսային մեծ խորության մեջ, այդ ապարների նախկին պաշտպանողական ծածկույթը չպետք է հասներ զգալի հաստության։ Ենթադրելով, թե գնեյսը, փայլարի թերթաքարը, գրանիտը, դիորիտը և այլն առաջ անպայման ծածկված են եղել, ինչպե՞ս կարող ենք բացատրել, որ այդ ապարներն այժմ մերկացած են ընդարձակ տարածությունների վրա աշխարհի զանազան մասերում, բացի այն ենթադրությունից, որ նրանց ծածկող բոլոր շերտերն ամբողջովին ոչնչացել են։ Որ այդպիսի ընդարձակ մարզեր իրոք կան, դրանում ոչ մի կասկած չկա. Պարիմի գրանիտային մարզը, համաձայն Հումբոլդտի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 199] նկարագրության, տասնևինը անգամ մեծ է Շվեյցարիայից։ Բուեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 200] քարտեզի վրա Ամազոնից դեպի հարավ ցույց է տրված այդ տիպի ապարներից կազմված մի մարզ, որ հավասար է Իսպանիային, Ֆրանսիային, Իտալիային, Գերմանիային և Մեծ Բրիտանիայի մի մասին միասին վերցրած։ Այդ մարզը դեռ հանգամանորեն ուսումնասիրված չէ, բայց ճանապարհորդների միաբերան վկայությամբ գրանիտային ապարները այդտեղ չափազանց լայն տարածում ունեն։ Ֆոն-Էշվեգը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 201] տալիս է այդ ապարների մանրամասն կտրվածքը, որն սկսվում է Ռիո-Ժանեյրոյից և ձգվում է ուղիղ գծով 260 աշխարհագրական մղոն դեպի երկրի ներսը. ես ինքս ճանապարհորդել եմ մի այլ ուղղությամբ 150 մղոն տարածության վրա և ոչինչ չեմ տեսել, բացի գրանիտի ապարներից։ Ես հետազոտել եմ ապարների բազմաթիվ նմուշներ, որ հավաքված էին ամբողջ ափի երկարությամբ Ռիո-Ժանեյրոյից մինչև Լա-Պլատայի գետաբերանը 1100 աշխարհագրական մղոն տարածության վրա, և նրանք բոլորն էլ ապարների այդ դասին էին պատկանում։ Երկրի ներսում, Լա-Պլատայի հյուսիսային ամբողջ ափի երկարությամբ, ես դիտել եմ, բացի նոր երրորդային ապարներից, թույլ ձևափոխված ապարների միայն մի փոքր հողամաս, որը միայն կարող էր ներկայացնել գրանիտային սերիայի սկզբնական ծածկույթի մնացորդը։ Դառնալով լավ հայտնի մի մարզի, այսինքն Միացյալ Նահանգներին և Կանադային, և օգտվելով պրոֆեսոր Ռոջերսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 202] կազմած հիանալի քարտեզից, ես հաշվել եմ տարածությունները, կտրելով դրանք քարտեզից ու թուղթը կշռելով, և գտել եմ, որ ձևափոխված (բացի կիսաձևափոխվածներից) և գրանիտային ապարները 19:12,5 հարաբերությամբ գերակշռում են ավելի նոր պալեոզոյիկ ֆորմացիաներին՝ բոլորին միասին վերցրած։ Շատ երկրներում ձևափոխված և գրանիտային ապարները զգալիորեն ավելի շատ տարածված կլինեին, քան այժմ նրանք թվում են, եթե հեռացվեր նստվածքային շերտերի ամբողջ հաստությունը, որն անհամաչափորեն ընկած է նրանց վրա և որը չէր կարող կազմել այն նախկին ծածկույթի մասը, որի տակ նրանք բյուրեղացել են։ Հետևաբար, միանգամայն հավանական է, որ աշխարհի որոշ մասերում ամբողջ ֆորմացիաներ իսպառ քայքայվել են, և նրանցից ոչ մի հետք չի մնացել։
Այստեղ ուշադրության արժանի է ևս մի դիտողություն։ Բարձրացման ժամանակաշրջաններում ցամաքի մարզը և նրան հարակից ծանծաղաջրային մարզը մեծանում են, և որի հետ միասին կարող են առաջանալ բնակվելու նոր կայաններ. և այս բոլորը, ինչպես բացատրվել է ավելի առաջ, նպաստում է նոր տարատեսակների ու տեսակների առաջացմանը, բայց հենց այդ ժամանակաշրջաններն էլ սովորաբար բաց են մնում երկրաբանական տարեգրության մեջ։ Մյուս կողմից, ցածրացման դարաշրջանում բնակելի տարածությունը և բնակիչների թիվը պետք է կրճատվեին (բացառությամբ այն ձևերի, որոնք ապրում են արշիպելագի վերածվող մայր ցամաքի ափերի մոտ). հետևաբար, այդ դարաշրջաններում պետք է տեղի ունենա ուժեղ մահաջնջում, և միայն քիչ նոր տարատեսակներ ու տեսակներ կարող են առաջանալ, և հենց այդ իջեցման ժամանակաշրջաններում էլ կուտակվել են բրածոներով ամենահարուստ նետվածքները։
ՅՈՒՐԱՔԱՆՉՅՈՒՐ ՖՈՐՄԱՑԻԱՅՈՒՄ ԲԱԶՄԱԹԻՎ ՄԻՋԱԿԱ ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԲԱՑԱԿԱՅՈՒԹՅԱՆ ՄԱՍԻՆ
Այս բոլոր կշռադատությունների հիման վրա չի կարելի կասկածել, որ երկրաբանական տարեգրությունն ամբողջությամբ վերցված վերին աստիճանի թերի է. բայց եթե ուշադրություն դարձնենք յուրաքանչյուր ֆորմացիայի վրա առանձին, ապա ավելի ևս դժվար է ըմբռնել, թե ինչու նրա մեջ մենք չենք գտնում իսկական միջակա տարատեսակներ այն ազգակից տեսակների միջև, որոնք ապրել են ֆորմացիայի սկզբում և վերջում։ Հայտնի են շատ օրինակներ, երբ միևնույն տեսակը ներկայացնում է տարատեսակներ նույն ֆորմացիայի վերին և ստորին մասերում, այսպես, Տրաուտշոլդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 203] բերում է մեծ թվով օրինակներ ամմոնիտների մասին. Հիլգենդորֆը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 204] նույնպես նկարագրել է մի շատ հետաքրքրական դեպք, որտեղ Planorbis multiformis-ի տասը ձևեր Շվեյցարիայում մի քաղցրահամ ջրերի ֆորմացիայի հաջորդական շերտերում աստիճանաբար փոփոխվում են։ Թեև յուրաքանչյուր ֆորմացիա անվիճելիորեն պահանջում է իր կուտակման համար հսկայական թվով տարիներ, այնուամենայնիվ կարելի է բերել շատ պատճառներ, որոնք բացատրում են, թե ինչու նրանցից յուրաքանչյուրը սովորաբար իր մեջ չի պարփակում շաղկապող օղակների անընդհատ շարքեր այն տեսակների միջև, որոնք ապրել են ֆորմացիայի սկզբում և վերջում, բայց ես չեմ կարող հարկ եղած չափով գնահատական տալ հետևյալ կշռադատումների հարաբերական նշանակությանը։
Թեև յուրաքանչյուր ֆորմացիա ինքնին կարող է նշել տարիների շատ երկար շարք, բայց նրանցից ամեն մեկը հավանորեն կարճ է այն ժամանակաշրջանի համեմատությամբ, որ հարկավոր է մի տեսակի փոփոխվելու և այլ տեսակ դառնալու համար։ Ինձ հայտնի է, որ երկու հնէաբան, որոնց կարծիքները արժանի են լիակատար հարգանքի, այսինքն Բրոննը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 136] և Վուդվորդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 205] եկել են այն եզրակացության, որ յուրաքանչյուր ֆորմացիայի միջին տևողությունը երկու կամ երեք անգամ գերազանցում է տեսակային ձևերի գոյության միջին տևողությանը։ Բայց անհաղթահարելի դժվարություններ են խանգարում մեզ, ինչպես ինձ թվում է, այդ տեսակետից գալու մի քիչ վստահելի եզրակացության։ Որևէ տեսակի առաջին անգամ հանդիպելով տվյալ ֆորմացիայի նստվածքներում, վերին աստիճանի համարձակություն կլինի դրանից եզրակացնել, թե նա առաջ էլ գոյություն չի ունեցել որևէ մի տեղ. ինչպես նաև նկատելով, որ տեսակն անհետանում է ֆորմացիայի վերջին շերտերի կուտակումից ավելի վաղ, նույնքան անմտածված կլիներ ենթադրել, թե տեսակը մահաջնջվել է հենց այդ ժամանակ։ Մենք մոռանում ենք, թե ինչքան փոքր է Եվրոպայի մակերևույթը ամբողջ աշխարհի մակերևույթի համեմատությամբ, բայց Եվրոպայի սահաններում էլ միևնույն ֆորմացիայի տարբեր հարկերը դեռևս միանգամայն ստույգ կերպով բաղդատված չեն միմյանց հետ։
Ծովային կենդանիների մասին մենք համարձակ կերպով կարող ենք ենթադրել, որ այստեղ լայն չափով տեղի են ունեցել գաղթեր, որոնց դրդապատճառ եղել են կլիմայական և այլ փոփոխությունները, և երբ մենք տեսնում ենք, որ մի տեսակ առաջին անգամ է երևում որևէ ֆորմացիայի մեջ, մենք կարող ենք հավանական համարել, որ նա այդ ժամանակ միայն առաջին անգամ վերաբնակվել է այդ մարզում։ Օրինակ, քաջ հայտնի է, որ շատ տեսակներ մի քիչ ավելի վաղ են երևացել Հյուսիսային Ամերիկայի պալեոզոյան շերտերում, քան Եվրոպայի միևնույն շերտերում․ ըստ երևույթին ամերիկական ծովերից եվրոպական ծովերը նրանց գաղթելու համար որոշ ժամանակ է պահանջվել։ Աշխարհի զանազան մասերում հետագա նստվածքներն ուսումնասիրելիս ամենուրեք նկատվել է, որ որոշ մինչև այժմ ապրող քիչ տեսակներ սովորական են տվյալ նստվածքում, բայց հարևան ծովում անհետացել են, կամ ընդհակառակը, որ որոշ տեսակներ հարևան ծովում շատ առատ են, բայց հազվագյուտ են կամ բոլորովին բացակայում են առանձին վերցրած այդ նստվածքի մեջ։ Վերին աստիճանի ուսանելի է խորհել Եվրոպայի բնակիչների վերաբնակման միանգամայն ապացուցված շատ դեպքերի մասին, որոնք տեղի են ունեցել մի ամբողջ երկրաբանական ժամանակաշրջանի միայն մի մասը ներկայացնող սառցադաշտային դարաշրջանի ընթացքում, ինչպես նաև խորհել ծովի մակարդակի փոփոխությունների մասին, կլիմայի ծայրահեղ փոփոխությունների և ժամանակի հսկայական տևողության մասին դարձյալ նույն սառցադաշտային ժամանակաշրջանի սահմաններում։ Սակայն, կարելի է կասկածել այն բանում, որ աշխարհի որևէ մասում անընդհատ կուտակվել են իրենց մեջ բրածո մնացորդներ պարունակող նստվածքային շերտեր ամբողջ այդ ժամանակաշրջանի ընթացքում հենց միևնույն մարզում։ Անհավատալի է, օրինակ, որ ամբողջ սառցադաշտային ժամանակաշրջանի ընթացքում նստվածքները կուտակվել են Միսիսիպիի գետաբերանի մոտ այն խորության սահմաններում, որի մեջ ծովային կենդանիները կարող են ամենից ավելի լավ զարգանալ, որովհետև մենք գիտենք, որ Ամերիկայի այլ մասերում հենց այդ ժամանակամիջոցում կատարվել են աշխարհագրական մեծ փոփոխություններ։ Երբ այդ շերտերը, որոնք Միսիսիպիի գետաբերանի մոտ ծանծաղ ծովում նստել են սառցադաշտային ժամանակաշրջանի մի որոշ մասի ընթացքում, կենթարկվեն բարձրացման, ապա օրգանական մնացորդները հավանորեն առաջին անգամ երևան կգան նստվածքների մեջ և կանհետանան զանազան մակարդակների վրա՝ տեսակների տեղափոխության և աշխարհագրական փոփոխությունների հետևանքով։ Եվ հեռավոր ապագայում երկրաբանը, ուսումնասիրելով այդ շերտերը, կհակվի այն եզրակացության գալու, որ այդ շերտերում պարփակված օրգանիզմների գոյության միջին տևողությունն ավելի քիչ է, քան սառցադաշտային ժամանակաշրջանի տևողությունը, մինչդեռ իրականում նա զգալիորեն ավելի մեծ է, որովհետև այդ օրգանիզմները սկսել են գոյություն ունենալ սառցադաշտային ժամանակաշրջանից առաջ և գոյություն ունեն մինչև այժմ։
Որպեսզի կարելի լինի երկու ձևերի միջև եղած անցումների լրիվ սերիան ստանալ միևնույն ֆորմացիայի վերին ու ստորին մասերից, նստվածքը պետք է կուտակվի անընդհատ կերպով երկար ժամանակաշրջանի ընթացքում, որը բավականանար փոփոխելիության դանդաղ պրոցեսի համար. հետևապես, նստվածքը պետք է ունենա շատ մեծ հզորություն, և փոփոխվող տեսակները պետք է ապրեն ամբողջ այդ ժամանակ հենց նույն մարզում։ Բայց մենք տեսանք, որ իր ամբողջ շերտերում բրածոներ պարունակող հզոր ֆորմացիան կարող է կուտակվել միայն հատակի իջեցման ժամանակաշրջանում, և որպեսզի խորությունը մոտավորապես միանման մնա, որը անհրաժեշտ պայման է հանդիսանում ծովային օրգանիզմների համար հենց նույն մարզում, նստվածքի կուտակման արագությունը պետք է մոտավորապես հավասարակշռի իջեցման արագությանը։ Բայց իջեցման հենց այդ պրոցեսի հետևանքը լինում է այն մարզի ընկղմումը, որտեղից բերվում է նստվածքի նյութը, և այսպիսով բերվող նյութի քանակը պակասում է, մինչդեռ իջեցումը դեռ շարունակվում է։ Իրականության մեջ բերվող նյութի քանակության և իջնելու արագության այդ գրեթե ճիշտ համապատասխանությունը հավանորեն հազվագյուտ պատահականություն է ներկայացնում, որովհետև շատ հնէաբաններ նկատել են, որ շատ հզոր նստվածքները սովորաբար չեն պարունակում բրածո օրգանիզմներ, որոնք հանդիպում են նստվածքների այդ ստվարության միայն վերին կամ միայն ստորին սահմանի մոտ։
Ըստ երևույթին նաև յուրաքանչյուր ֆորմացիան, որևէ երկրի ֆորմացիաների ամբողջ սերիայի նման, սովորաբար ներկայացնում է ընդմիջվող շերտակուտակում։ Երբ մենք դիտում ենք, ինչպես այդ հաճախ պատահում է, զանազան հանքաբանական կազմություն ունեցող շերտերից բաղկացած մի ֆորմացիա, մենք իրավունք ունենք ենթադրելու, որ նրա կուտակման պրոցեսում եղել են ավելի մեծ կամ փոքր ընդմիջումներ։ Եվ որևէ ֆորմացիայի ամենաուշադիր հետազոտությունը մեզ ոչ մի պատկերացում չի տալիս այն ժամանակի տևողության մասին, որը հարկավոր է եղել նրա կուտակման համար։ Կարելի է շատ օրինակներ բերել, որ ընդամենը մի քանի ոտնաչափ հաստությամբ շերտերը իրենցից ներկայացնում են ֆորմացիաներ, որոնք այլ վայրերում ունեն հազարավոր ոտնաչափ հզորություն և իրենց կուտակման համար հսկայական ժամանակաշրջան են պահանջել, սակայն չգիտենալով այս փաստը, ոչ ոքի մտքով էլ չի անցնի, որ այդ ամենաքիչ հզորություն ունեցող ֆորմացիան այդպիսի հսկայական ժամանակամիջոց է արտահայտում։ Կարելի է նշել շատ դեպքեր, որտեղ որևէ ֆորմացիայի ստորին շերտերը բարձրացել են, ենթարկվել քայքայման, կրկին իջել են և ծածկվել նույն ֆորմացիայի վերին շերտերով, փաստեր, որոնք ցույց են տալիս, որ երկարատև, բայց ուշադրությունից հեշտությամբ վրիպող ընդմիջումներ տեղի են ունեցել այդպիսի շերտերի կուտակման ժամանակ։ Այլ դեպքերում բրածո մեծ ծառերը, որոնք դեռ պահպանել են ուղղաձիգ դիրքը, ինչ դիրքի մեջ նրանք աճել են, մեզ պարզ կերպով ցույց են տալիս ժամանակի այն շատ երկարատև ժամանակամիջոցները և երկրի մակարդակի փոփոխությունները կուտակման պրոցեսի ընթացքում՝ փոփոխություններ, որոնց մասին չէինք կարող կռահել, եթե չպահպանվեին այդ ծառերը. այսպես, սըր Չ. Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] և դ-ր Դոուսոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 196] Նոր Շոտլանդիայում գտել են ածուխ պարունակող շերտեր 1400 ոտնաչափ հաստությամբ, որի մեջ գտնվել են արմատներ պարունակող հին միջնաշերտեր, որոնք դասավորված են եղել մեկը մյուսի վրա 68-ից ոչ պակաս տարբեր մակարդակների վրա։ Այսպիսով, երբ միևնույն տեսակը հանդիպում է որևէ ֆորմացիայի հիմքում, միջին շերտերում և վերին հորիզոններում, խիստ հավանական է ենթադրել, որ նա միևնույն տեղում չի ապրել կուտակման ամբողջ ժամանակաշրջանի ընթացքում, այլ անհետացել ու վերստին երևացել է, և գուցե թե շատ անգամ, քանի դեռ շարունակվել է միևնույն երկրաբանական ժամանակաշրջանը։ Հետևաբար, եթե այդ տեսակը զգալի փոփոխություն է կրել երկրաբանական մեկ ֆորմացիայի կուտակման ժամանակ, նրա մեկ կտրվածքը իր մեջ չի պարփակի բոլոր այն նուրբ միջակա անցումները, որոնք, մեր տեսության համաձայն, պետք է գոյություն ունեցած լինեին, այլ կպարունակի այդ ձևի ցայտուն կերպով արտահայտված, թերևս նաև աննշան փոփոխությունները։
Չափազանց կարևոր է հիշել, որ բնախույզները չունեն ոսկյա կանոն, որի միջոցով նրանք կկարողանային տարբերել տեսակները տարատեսակներից։ Նրանք ընդունում են յուրաքանչյուր տեսակի սահմաններում որոշ թույլ փոփոխելիություն, բայց երբ նրանք հանդիպում են մի քիչ ավելի մեծ տարբերությունների երկու որևէ ձևերի միջև, նրանք այդ ձևերը համարում են առանձին տեսակներ, եթե միայն հնարավորություն չեն ունենում շաղկապել միմյանց հետ նրանց մերձավոր միջակա անցումներով, իսկ այդ բանի վրա, հենց նոր շարադրված պատճառներով, շատ հազվագյուտ դեպքում կարելի է հույս դնել, երբ գործ ունենք երկրաբանական կտրվածքի հետ։ Ենթադրենք, թե B և C ներկայացնում են երկու տեսակներ, իսկ երրորդը՝ A-ն գտնված է ավելի հին, ավելի ցած ընկած շերտի մեջ, եթե A-ն նույնիսկ խիստ միջակա ձև լիներ B-ի և C-ի միջև, նա պարզապես կհամարվեր մի երրորդ և առանձին տեսակ, եթե միայն միևնույն ժամանակ նրան չէր կարելի սերտ կերպով կապել միջակա տարատեսակների միջոցով ձևերից մեկի կամ երկուսի հետ։ Սակայն չպետք է մոռանալ, ինչպես այդ բացատրվել է ավելի առաջ, որ թեկուզ A-ն եղել է B-ի և C-ի իսկական նախածնողը, դրանից դեռևս անպայման չի հետևում, որ նա բոլոր տեսակետներից պիտի լինի նրանց միջև խիստ միջակա ձև։ Այսպիսով, մենք կարող ենք միևնույն ֆորմացիայի ստորին ու վերին շերտերից ստանալ ծնողական տեսակները և նրանց բազմազան կերպով փոփոխված հաջորդները, և եթե մենք չունենք բազմաթիվ անցումային ձևեր, մենք չենք էլ կարող կռահել նրանց միջև արյունակցական ազգակցության գոյության մասին և նրանց կհամարենք տարբեր տեսակներ։
Հայտնի է, թե ինչպիսի աննշան տարբերությունների հիման վրա շատ հնէաբաններ որոշել են իրենց տեսակները. նրանք այդ անում են ավելի ևս հաճույքով, եթե օրինակները ծագում են միևնույն ֆորմացիայի տարբեր ստորաբաժանումներից։ Մի քանի հմուտ հնէաբաններ դ’Օրբինիի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 206] և մյուսների կողմից ճանաչված տեսակներից շատերին այժմ իջեցնում են տարատեսակների աստիճանի, և դրանում, իհարկե, մենք գտնում ենք փոփոխելիության որոշ ապացույց, որը մեր տեսությանն համաձայն պետք է գտնվեր։ Այժմ մի անգամ էլ դառնանք երրորդային ուշ նստվածքներին, որոնք պարունակում են, բնախույզների մեծամասնության կարծիքով, այժմ ապրող տեսակներին նման բազմաթիվ խեցիներ։ Մի քանի հիանալի բնախույզներ, օրինակ Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] և Պիկտեն,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 207] պնդում են, որ այդ բոլոր երրորդային ձևերը ներկայացնում են այլ տեսակներ, թեև ընդունում են, որ նրանք այժմյան ձևերից շատ քիչ են տարբերվում. այսպիսով, եթե չենթադրենք, որ այդ ականավոր բնախույզներն իրենց երևակայության շնորհիվ ընկել են մոլորության մեջ և որ այդ վերին երրորդային տեսակները էապես ոչնչով չեն տարբերվում իրենց այժմյան ներկայացուցիչներից, կամ եթե չընդունենք, հակառակ բնախույզների մեծամասնության կարծիքի, որ այդ երրորդային տեսակները բոլորն էլ իսկապես տարբերվում են այժմյան ձևերից, այդ դեպքում մենք ցուցում ենք գտնում, որ հաճախ հանդիպում են այնպիսի թույլ փոփոխություններ, որպիսիք ենթադրում է իմ տեսությունը։ Եթե մենք դառնանք մի քիչ ավելի մեծ ժամանակամիջոցներին, այսինքն միևնույն ֆորմացիայի տարբեր, բայց հաջորդական հարկերին, մենք կտեսնենք, որ նրանց մեջ պարունակվող բրածոները թեև բոլոր կողմից ճանաչվում են որպես տեսակային տեսակետից տարբեր ձևեր, բայց նրանք ավելի սերտ կերպով կապված են միմյանց հետ, քան իրարից հեռու ընկած ֆորմացիաներում գտնվող տեսակները. այնպես որ այստեղ ևս մենք հանդիպում ենք աներկբայելի մի վկայության հօգուտ տեսության պահանջած ուղղությամբ կատարվող փոփոխությունների։ Այս վերջին հարցին ես դեռ կվերադառնամ հետևյալ գլխում։
Այն կենդանիների և բույսերի վերաբերմամբ, որոնք արագ բազմանում են և աչքի չեն ընկնում շարժունությամբ, հիմք կա ենթադրելու, ինչպես այղ տեսանք ավելի առաջ, որ նրանց տարատեսակները սկզբում սովորաբար հարմարված են լինում մի վայրի և որ այդպիսի տեղական տարատեսակները լայնորեն տարածվում են ու դուրս են մղում իրենց ազգակից ձևերին միայն այն բանից հետո, երբ նրանք բավական զգալի չափով փոփոխվել են ու կատարելագործվել։ Այդպիսի պայմաններում քիչ հավանականություն կա հայտնաբերելու որևէ երկրի մի ֆորմացիայում բոլոր նախկին անցումային աստիճանները որևէ երկու ձևերի միջև, որովհետև կարելի է ենթադրել, որ հաջորդական փոփոխությունները տեղական բնույթ են ունեցել կամ կատարվել են մի սահմանափակ մարզի սահմաններում։ Ծովային կենդանիները մեծ մասամբ ունեն լայն տարածում, և մենք տեսանք, որ բույսերի մեջ տարատեսակներով ամենից ավելի հարուստ են նրանք, որոնք ամենից ավելի լայն են տարածված։ Խեցիների և ծովային այլ կենդանիների նկատմամբ միանգամայն հավանական է, որ նրանցից ամենից ավելի հաճախ նրանք են սկիզբ տվել սկզբում տեղական տարատեսակների, իսկ հետո նաև նոր տեսակների, որոնք ամենից ավելի լայն կերպով են տարածված, և նրանց տարածումը անցնում է Եվրոպայում հայտնի երկրաբանական ֆորմացիաների սահմաններից հեռու, իսկ դա ևս դարձյալ զգալիորեն իջեցնում է որևէ երկրաբանական ֆորմացիայում անցումային աստիճաններ որոնելու, գտնելու հնարավորության հավանականության աստիճանը։
Կարելի է նշել նաև Ֆոկոների (Falconer)[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 73] վերջերս արտահայտված մի ավելի կարևոր կշռադատությունը, որը բերում է այն եզրակացության, որ այն ժամանակաշրջանը, որի ընթացքում յուրաքանչյուր տեսակ փոփոխություն է կրել, թեև շատ երկարատև է, եթե այն չափենք տարիներով, հավանորեն կարճ է եղել ա,յն ժամանակի համեմատությամբ, որի ընթացքում տեսակը ոչ մի փոփոխության չի ենթարկվել։
Չպետք է մոռանալ, որ ներկայումս հետազոտության համար ունենալով նույնիսկ լրիվ օրինակները, մենք միայն հազվագյուտ դեպքերում կարող ենք երկու ձև միջակա տարատեսակներով շաղկապել միմյանց հետ և այսպիսով ապացուցել նրանց պատկանելիությունը միևնույն տեսակին, եթե չունենք զանազան վայրերից հավաքած մեծ թվով օրինակներ, իսկ բրածո տեսակների վերաբերմամբ դա հազվադեպ է լինում հնարավոր։ Մենք թերևս ամենից ավելի լավ կպատկերացնենք այն ենթադրության ամբողջ անհավանականությունը, որ մեզ կհաջողվի բազմաթիվ բրածո անցումային օղակներով միմյանց հետ շաղկապել տեսակները, եթե մեզ հարց տանք, օրինակ՝ թե ապագա մի որոշ ժամանակաշրջանի երկրաբան ի վիճակի՞ է արդյոք ապացուցելու, որ մեր անասունների՝ ոչխարների, ձիերի, շների բազմազան ցեղերը առաջացել են մեկ արմատից կամ մի քանի նախասկզբնական արմատներից․ կամ ինչպես նաև այն ծովային խեցիները, որոնք ապրում են Հյուսիսային Ամերիկայի ափերի մոտ և որոնց կոնխիլիոլոգներից ոմանք համարում են նրանց եվրոպական ներկայացուցիչների տարբեր տեսակներ, իսկ մյուսները համարում են տարատեսակներ, արդյոք իսկապե՞ս տարատեսակներ են միայն, թե նրանք տարբերվում են միմյանցից որպես տեսակներ։ Դա ապագայի երկրաբանի համար հնարավոր կլինի այն դեպքում, եթե նա կհայտնաբերի բրածո վիճակում բազմաթիվ անցումային ձևեր։ Բայց նման հաջողությունը վերին աստիճանի անհավանական է։
Տեսակների անփոփոխելիության հավատացող հեղինակները չեն դադարել պնդելուց, թե երկրաբանությունը բոլորովին չի հայտնաբերում անցումային ձևեր։ Այդ պնդումը, ինչպես մենք կտեսնենք ամենամոտ գլխում, միանգամայն սխալ է։ Սըր Ջոն Լեբբոկի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 60] դիտողության համաձայն՝ «յուրաքանչյուր տեսակ ներկայացնում է շաղկապող օղակ մյուս ազգակից ձևերի միջև»։ Եթե մենք վերցնենք մի սեռ, որը բաղկացած է ապրող և մահաջնջված երկու տասնյակ տեսակներից և ոչնչացնենք նրանցից չորս հինգերորդը, ապա մնացած տեսակները, առանց որևէ կասկածի, կսկսեն զգալիորեն ավելի շատ տարբերվել մեկը մյուսից։ Եթե լինի, որ պատահմամբ այսպիսով ոչնչացեք են հատկապես ծայրային ձևերը, ապա ամբողջ սեռն այն ժամանակ ավելի առանձնացած կլինի մյուս մերձավոր սեռերից։ Այն, ինչ դեռ ևս չեն հայտնաբերել երկրաբանական հետազոտությունները, դա անսահմանորեն բազմաթիվ անցումների գոյությունն է, որոնք այնքան մոտ են միմյանց, որքան մոտ են այժմյան տարատեսակները, անցումներ, որոնք միմյանց հետ շաղկապում են բոլոր գոյություն ունեցող և մահաջնջված տեսակներին։ Բայց դրա վրա հույս դնել չի կարելի, թեև այդպիսի անցումների բացակայությունը բազմիցս նշվել է որպես ամենալուրջ առարկություն իմ տեսակետների դեմ։
Գուցե տեղին կլիներ երկրաբանական տարեգրության անլրիվության պատճառների մասին եղած նախորդ դատողությունները եզրափակենք հետևյալ երևակայական լուսաբանությամբ։ Մալայան արշիպելագն իր մեծությամբ համարյա հավասար է Եվրոպային՝ սկսած Նորդ-Կապից մինչև Միջերկրական ծովը և Մեծ Բրիտանիայից մինչև Ռուսաստան, և հետևաբար տարածությամբ հավասար է այն բոլոր երկրաբանական ֆորմացիաներին, որոնք որքան և իցե հանգամանորեն ուսումնասիրված են, չհաշված Ամերիկայի Միացյալ Նահանգներում զարգացած ֆորմացիաները։ Ես միանգամայն համաձայն եմ մ-ր Գոդվին-Աուստենի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 208] հետ, որ Մալայան արշիպելագի այժմյան վիճակն իր բազմաթիվ մեծ կղզիներով, որ մեկը մյուսից բաժանված են լայն, բայց ծանծաղ ծովերով, հավանորեն մեզ ներկայացնում են այն դրությունը, որի մեջ առաջներում գտնվել է Եվրոպան, այն ժամանակ, երբ տեղի է ունեցել մեր ֆորմացիաների մեծ մասի կուտակումը։ Մալայան արշիպելագը այն մարզերից մեկն է, որոնք ամենից ավելի հարուստ են օրգանական էակներով. սակայն եթե հավաքենք այն բոլոր տեսակները, որոնք երբևիցե ապրել են այստեղ, ինչքա՜ն թերի կներկայացնեին նրանք աշխարհի բնապատմությանը։
Բայց մենք ունենք բոլոր հիմքերը մտածելու, որ արշիպելագի մայր ցամաքային բնակչությունը կարող էր պահպանվել միայն չափազանց ոչ լրիվ ձևով այն ֆորմացիաներում, որոնց կուտակումը մենք այնտեղ ենթադրում ենք։ Այնտեղ կարող էին թաղվել միայն իսկական առափնյա կամ ստորջրյա մերկ ժայռերի վրա ապրող կենդանիներից քչերը և դրանցից էլ նրանք, որոնք թաղված են խճաքարի կամ ավազի մեջ, չէին կարող պահպանվել մինչև հեռավոր դարաշրջանը։ Ամենուրեք, որտեղ նստվածքներ չեն կուտակվում ծովի հատակին կամ որտեղ նրանք նստում են ոչ բավարար արագությամբ, որպեսզի քայքայումից պահպանեն օրգանական մարմինները, բրածո մնացորդներ բոլորովին չէին պահպանվի։
Բազմազան բրածոներով հարուստ և բավականաչափ հզոր ֆորմացիաները որպեսզի պահպանվեին մինչև ապագա մի դարաշրջան, որը մեզանից այնքան հեռու է, որքան անցյալում հեռու են երկրորդային ֆորմացիաները, կարող էին արշիպելագում առաջանալ միայն իջնելու ժամանակաշրջաններում։ Իջնելու այդ ժամանակաշրջանները պետք է մեկը մյուսից բաժանված լինեն հսկայական ժամանակամիջոցներով, որոնց ընթացքում տվյալ մարզը կամ մնացել է անշարժ, կամ բարձրացել է. բայց բարձրանալու ժամանակ, եթե ափերը զառիվայր են, բրածո պարունակող ֆորմացիաները պիտի քայքայվեին ծովի ափակոծության ներգործությունից նույնքան արագ կերպով, որքան արագ որ կուտակվել էին, որի օրինակը մենք այժմ տեսնում ենք Հարավային Ամերիկայի ափերի մոտ։ Նույնիսկ արշիպելագի ընդարձակ և ծանծաղ ծովերում բարձրացման ժամանակաշրջաններում նստվածքային շերտերը հազիվ թե կարողանային մեծ զանգվածներով կուտակվել կամ ծածկվել հետագա նստվածքների պաշտպանական ծածկույթով և այսպիսով ապահովել մինչև հեռավոր ապագա ժամանակները իրենց պահպանման հավանականությունը։ Իջեցման ժամանակաշրջաններում այստեղ մեծ չափերով մահաջնջում պիտի տեղի ունենար, բարձրացման ժամանակաշրջաններում այստեղ հավանորեն ձևերի ուժեղ փոփոխելիություն է ընթացել, բայց այդ վերջին ժամանակաշրջանների ընթացքում երկրաբանական տարեգրությունը չէր կարող առանձնապես լրիվ լինել։
Կարելի է կասկածել, որ արշիպելագի ամբողջ տարածության վրա կամ նրա մի մասում իջեցման նույնիսկ ամենաերկար ժամանակաշրջանի և դրա հետ միաժամանակ նստվածքների կուտակման տևողությունը գերազանցել է միևնույն տեսակային ձևերի գոյության միջին տևողությանը, իսկ այդ դեպքերը անհրաժեշտ պայման են երկու կամ մի քանի տեսակների միջև բոլոր միջակա ձևերի պահպանման համար։ Եթե այղ միջակա ձևերը ոչ բոլորն են պահպանվել, ապա անցումային բոլոր տարատեսակները պետք է թվան նոր, թեև մերձավոր ազգակից տեսակներ։ Հավանական է նույնպես, որ յուրաքանչյուր իջեցման մեծ ժամանակաշրջան պետք է ընդհատվեր մակարդակի տատանումներով և որ այդ երկարատև ժամանակաշրջաններում պետք է տեղի ունենային նաև որոշ կլիմայական փոփոխություններ, իսկ այդ պայմաններում արշիպելագի բնակիչները պետք է գաղթեին և չէին կարող որևէ ֆորմացիայում պահպանվել նրանց փոփոխությունների խիստ հաջորդական հուշարձանները։
Արշիպելագի ծովային բնակիչներից շատ շատերը այժմ տարածված են նրա սահմաններից հազարավոր մղոններով հեռու, և ըստ անալոգիայի մենք լիակատար իրավունք ունենք կարծելու, որ գլխավորապես հենց այդ լայնորեն տարածված տեսակները, կամ գոնե նրանցից մի քանիսը պետք է ամենից ավելի հաճախ տան նոր տարատեսակներ, իսկ տարատեսակները սկզբում եղել են տեղական կամ տարածվել են փոքր տարածության վրա, բայց եթե նրանք ունեցել են վճռական առավելություններ կամ եթե ավելի շատ են փոփոխվել ու կատարելագործվել, ապա նրանք պետք է դանդաղ կերպով տարածվեին և դուրս մղեին իրենց ազգակից ձևերին։ Իրենց նախկին հայրենիքը վերադարձած այդպիսի տարատեսակները շնորհիվ այն հանգամանքի, որ նրանք գրեթե հավասարաչափ կերպով, թեև շատ թույլ շեղվել են իրենց նախկին ձևից և որ նրանք պարփակված են տվյալ ֆորմացիայի ոչ միևնույն հորիզոնում, համաձայն այն կանոնների, որոնցով ղեկավարվում են շատ հնէաբաններ, պետք է համարվեն նոր ինքնուրույն տեսակներ։
Այսպես ուրեմն՝ եթե այս դիտողություններում ճշմարտության նշույլ կա, ապա մենք իրավունք չունենք հուսալու, որ մեր երկրաբանական ֆորմացիաներում հնարավոր է գտնել անսահման թվով այն անցումային ձևերը, որոնց միջոցով, մեր տեսության համաձայն, միևնույն խմբին պատկանող բոլոր նախկին և ժամանակակից ձևերը շաղկապված են միմյանց հետ՝ կազմելով կյանքի երկար ու ճյուղավորված մի շղթա։ Մենք կարող ենք հուսալ, որ կգտնենք միայն քիչ օղակներ, և դրանք մենք իրոք գտնում ենք, մի քանիսը ավելի հեռավոր, մյուսներն ավելի սերտորեն կապված իրար հետ. և այդպիսի օղակները, ինչքան էլ մոտ լինեն միմյանց, բայց եթե միայն նրանք գտնվում են միևնույն ֆորմացիայի տարբեր հարկերում, շատ հնէաբանների կողմից համարվում են առանձին տեսակներ։ Չեմ թաքցնում, որ ես երբեք էլ չէի կասկածի, թե ինչքան աղքատիկ են երկրաբանական հուշարձանները ամենալավ պահպանված երկրաբանական կտրվածքներում, եթե յուրաքանչյուր ֆորմացիայի սկզբում և վերջում ապրող տեսակների միջև կապող անհամար օղակների բացակայությունը այնքան ծանրակշիռ փաստարկ չլիներ իմ տեսության դեմ։
ԱԶԳԱԿԻՑ ՏԵՍԱԿՆԵՐԻ ԱՄԲՈՂՋ ԽՄԲԵՐԻ ՀԱՆԿԱՐԾԱԿԻ ԵՐԵՎԱՆ ԳԱԼՈՒ ՄԱՍԻՆ
Շատ հնէաբաններ, օրինակ Ագասսիսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] Պիկտեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 207] և Սեջվիկը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 209] համառորեն նշել են տեսակների ամբողջ խմբերի երևան գալը զանազան ֆորմացիաներում որպես ճակատագրական առարկություն, որը հերքում է տեսակների փոփոխելիության մասին եղած կարծիքը։ Եթե միևնույն սեռերին կամ ընտանիքներին պատկանող բազմաթիվ տեսակները իսկապես մի անգամից են սկսել իրենց գոյությունը, ապա այդ փաստը ճակատագրական կլիներ բնական ընտրության միջոցով կատարվող էվոլյուցիայի տեսության համար, որովհետև մի որոշ ընդհանուր նախածնողից առաջացած բոլոր ձևերի խմբի այդ ճանապարհով զարգացումը պետք է ներկայացներ չափազանց դանդաղ մի պրոցես, և նախածնողներն էլ պետք է ապրեին իրենց փոփոխված հաջորդներից շատ առաջ։ Բայց մենք միշտ չափազանցնում ենք երկրաբանական տարեգրության լրիվությունը, և այն փաստից, որ որոշ սեռեր կամ ընտանիքներ չեն գտնվել որոշ հարկից ներքև, սխալ կերպով եզրակացնում ենք, որ նրանք գոյություն էլ չեն ունեցել այդ հարկից առաջ։ Համենայն դեպս հնէաբանության դրական ցուցմունքներին կարելի է լիովին վստահել, մինչդեռ բացասական տվյալները նշանակություն չունեն, ինչպես այդ հաճախ հաստատվել է փորձով։ Մենք միշտ մոռանում ենք, թե որքան մեծ է աշխարհը այն մարզի համեմատությամբ, որի մեջ մեր երկրաբանական ֆորմացիաները մանրազնին կերպով հետազոտված են. մենք մոռանում ենք, որ տեսակների խմբերը նախքան երևան են եկել Եվրոպայի և Միացյալ նահանգների հին արշիպելագներում, կարող էին երկար ժամանակ գոյություն ունենալ և դանդաղ կերպով բազմանալ որևէ այլ տեղում։ Մենք բավարար չափով չենք գնահատում այն ժամանակամիջոցները, որոնք բաժանում են մեր հաջորդական ֆորմացիաները մեկը մյուսից և որոնք շատ դեպքերում գուցե ավելի երկարատև են, քան յուրաքանչյուր ֆորմացիայի կուտակման համար անհրաժեշտ ժամանակը։ Այդ ժամանակամիջոցները բավական են մեկ նախահայր ձևից ծագում առած տեսակների բազմացման համար, և հաջորդ ֆորմացիայում տեսակների այդպիսի խմբեր կարող են հանկարծ երևալ՝ կարծես թե հանկարծ ստեղծվածներ են։
Ես այստեղ ուզում եմ հիշեցնել մի դիտողություն, որ ես արել եմ առաջ, այդ այն է, որ կարող է դարերի երկար շարք պահանջվել, որպեսզի օրգանիզմը հարմարվի որոշ նոր և հատուկ կենսապայմանների, օրինակ օդում թռչելու, և որ, հետևաբար, անցումային ձևերը հաճախ պետք է երկար ժամանակ սահմանափակ տարածում ունենային որևէ մի մարզում, բայց երբ այդպիսի հարմարեցումը տեղի է ունեցել և քիչ տեսակներ այդպիսով ձեռք են բերել մեծ առավելություն այլ օրգանիզմների հանդեպ, բավական է արդեն համեմատաբար կարճ ժամանակ, որպեսզի առաջանան տարամիտվող շատ ձևեր, որոնք արագորեն ու լայնորեն տարածվում են ամենուրեք։ Պրոֆ. Պիկտեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 207] այս աշխատության մասին իր հիանալի վերլուծության մեջ խոսելով անցումային վաղ ձևերի մասին և որպես օրինակ վերցնելով թռչուններին, չի կարողանում պատկերացնել, թե ինչպես ենթադրյալ նախատիպի առջևի վերջավորությունների հաջորդական փոփոխությունները կարող էին որևէ տեսակետից առավելություն ներկայացնել։ Սակայն ուշադրություն դարձնենք Հարավային օվկիանոսի պինգվինների վրա։ Այդ թռչունների առջևի վերջավորություններն արդյոք չե՞ն գտնվում հենց ճիշտ այնպիսի անցումային վիճակում, որ նրանք «ոչ թաթեր են, ոչ էլ թևեր»։ Մինչդեռ այդ թռչունները հաղթականորեն պահպանում են իրենց տեղը գոյության կռվում, քանի որ նրանք ապրում են անհամար քանակությամբ և շատ ձևերով։ Ես չեմ ենթադրում, թե մենք այստեղ ունենք իսկական անցումային աստիճաններ, որոնցով անցել են թռչունների թևերն իրենց զարգացման ընթացքում, բայց արդյո՞ք մենք որևէ առանձին դժվարության կհանդիպենք, ընդունելով այն հնարավորությունը, որ պինգվինի որևէ փոփոխված հետնորդին օգտակար է եղել ձեռք բերել նախ տեղափոխվելու ընդունակություն՝ խփելով իր թևերը ջրի մակերեսին, ինչպես այդ անում է ֆալկլանդյան կարճաթև բադը, իսկ հետո նաև բարձրանալ ջրի մակերեսից ու փոխադրվել օդի մեջ։
Ես այժմ կբերեմ մի քանի օրինակներ, որոնք պարզաբանում են վերոհիշյալ դիտողությունները, և ցույց կտամ, թե ինչպես մենք սխալի մեջ ընկնելու վտանգի ենք ենթարկվում, ենթադրելով, որ տեսակների ամբողջ խմբեր հանկարծակի են առաջացել։ Նույնիսկ այնպիսի կարճատև ժամանակամիջոցում, որն անցել է Պիկտեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 207] հնէաբանական մեծ աշխատության առաջին և երկրորդ հրատարակությունների միջև, որ հրատարակվել են 1844—1846 և 1853—1857 թվականներին, մեր տեղեկությունները կենդանիների շատ խմբերի առաջին անգամ երևալու և անհետանալու մասին զգալիորեն փոխվել են, իսկ երրորդ հրատարակությունը հավանորեն կպահանջի է՛լ ավելի զգալի փոփոխություններ։ Ես կարող եմ հիշեցնել մի լավ հայտնի փաստ, որ ընդամենը մի քանի տարի առաջ հրատարակված երկրաբանական ձեռնարկներում ասվում էր, որ կաթնասունները հանկարծակի երևան են եկել երրորդային սերիայի սկզբում, իսկ այժմ բրածո կաթնասունների գտնվելու ամենահարուստ վայրերից մեկը հայտնի է երկրորդային սերիայի կեսին և բացի դրանից, իսկական կաթնասուններ են հայտնաբերվել նոր կարմիր ավազաքարի մեջ՝ այդ մեծ սերիայի գրեթե ամենասկզբում։ Կյուվիեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 133] բազմիցս հայտնել է այն համոզմունքը, որ երրորդային նստվածքներից ոչ մեկի մեջ բրածո կապիկներ չկան, իսկ այժմ բրածո տեսակներ հայտնաբերված են Հնդկաստանում, Հարավային Ամերիկայում և Եվրոպայում, նույնիսկ այնպիսի խոր շերտերում, ինչպես միոցենայիններն են։ Եթե չլինեին Միացյալ նահանգների նոր կարմիր ավազաքարի վրա ոտների հետքերի պահպանման հազվագյուտ դեպքերը, ո՞վ կարող էր ենթադրել, որ այդ ժամանակաշրջանում գոյություն են ունեցել առնվազն երեսուն զանազան տեսակի թռչնանման կենդանիներ և նրանցից մի քանիսները հասնում էին հսկայական մեծության։ Այդ շերտերում ոսկրի ոչ մի բեկոր չի գտնվել։ Դեռ ոչ այնքան վաղ հնէաբանները այն կարծիքին էին, թե թռչունների ամբողջ դասը հանկարծակի կերպով երևան է եկել էոցենի ժամանակաշրջաններում, իսկ այժմ մենք գիտենք ըստ պրոֆ. Օուենի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] վկայության, որ թռչունների մի ձև անկասկած գոյություն է ունեցել ներին կանաչ ավազաքարի նստվածքների դարաշրջանում, իսկ դեռ ավելի վերջերս Սոլենհոֆենի[67] յուրայի թերթաքարերում գտնված է Archeopteryx տարօրինակ թռչունը, որն ունեցել է մողեսի նման երկար պոչ, որի յուրաքանչյուր ողի վրա նստած են եղել երկու փետուրներ և թևեր, որոնց վրա եղել են երկուական ճանկեր։ Այդ հայտնագործությունը հազիվ թե ոչ ավելի քիչ, քան մի որևէ ուրիշը, ցույց տվեց, թե ինչքան մենք դեռ քիչ գիտենք երկրի հին բնակիչների մասին։
Ես կարող եմ բերել ևս մեկ օրինակ, որին ես ինքս վկա եմ եղել, և որը այղ պատճառով առանձնապես զարմացրել է ինձ։ Բրածո նստակյաց փնջոտնյա խեցգետինների (Cirripedia) մասին, իմ մեմուարում, հիմնվելով այժմ ապրող և երրորդային մահաջնջված տեսակների մեծ թվի վրա, անհատներով արտասովոր հարուստ շատ տեսակների վրա, որոնք տարածված են ամբողջ աշխարհում՝ սկսած արկտիկական մարզերից մինչև հասարակած և ապրում են խորության զանազան գոտիներում՝ սկսած մակընթացության վերին սահմանից մինչև 50 ֆաթոմը[66], հիմնվելով նույնիսկ ամենահին երրորդային շերտերում օրինակների հիանալի պահպանվածության վրա և այն հեշտության վրա, որով կարելի է ճանաչել այդ կենդանու փեղկի նույնիսկ բեկորը, հիմնվելով այս բոլոր հանգամանքների վրա, ես պնդել եմ, որ եթե բրածո Cirripedia-ները գոյություն ունենային երկրորդային ժամանակաշրջանում, նրանք անպայման պահպանված ու գտնված կլինեին. բայց քանի որ այն ժամանակ ոչ մի տեսակ չէր հայտնաբերվել այդ տարիքն ունեցող շերտերում, ուստի ես դրանից եզրակացրել եմ, որ այդ մեծ խումբը հանկարծակի կերպով զարգացել է երրորդային դարաշրջանի սկզբում։ Դա խիստ կերպով շփոթեցնում էր ինձ, ավելացնելով էլի մի ավելորդ օրինակ, ինչպես ես այն ժամանակ կարծում էի, որ տեսակների մի մեծ խումբ հանկարծակի կերպով է երևան եկել։ Բայց հենց որ լույս տեսավ իմ աշխատությունը, մի փորձառու հնէաբան՝ Բոսկեն,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 210] ինձ ուղարկեց նստակյաց Cirripedia-ի անկասկածելի մի ներկայացուցչի լրիվ օրինակի նկարը. այդ բրածոյին նա ինքն էր հանել Բելգիայի կավճային նստվածքներից, և կարծես թե դեպքը առանձնապես զարմանալի դարձնելու համար դուրս եկավ, որ դա Chthamalus էր, մի շատ սովորական, խոշոր և ամենուրեք տարածված սեռ, որի ոչ մի տեսակը մինչև այժմ գտնված չէ նույնիսկ ոչ մի երրորդային նստվածքում։ Ավելի ևս վերջերս մ-ր Վուդվորդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 205] վերին կավճի մեջ հայտնաբերել է մի Pyrgoma, որը նստակյաց Cirripedia-ի մի առանձին ենթաընտանիքի ներկայացուցիչ է. այնպես որ ներկայումս մենք բավականաչափ ապացույցներ ունենք, որ կենդանիների այդ խումբը գոյություն է ունեցել երկրորդային ժամանակաշրջանի ընթացքում։
Հնէաբանների առանձնապես հաճախ հիշատակած տեսակների ամբողջ խմբի հանկարծակի կերպով երևան գալու օրինակ հանդիսանում են ոսկրավոր ձկները, որոնք երևացել են, Ագասսիսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] վկայությամբ, կավճային ժամանակաշրջանին պատկանող շերտերում։ Այդ խումբն իր մեջ պարփակում է այժմ ապրող տեսակների հսկայական մեծամասնությունը։ Բայց յուրայի և տրիասի մի քանի ձևեր այժմ բոլորը համարվում են որպես ոսկրավորներին պատկանողներ. մի բարձր հեղինակություն այղ խմբի մեջ է զետեղել նույնիսկ պալեոզոյան որոշ ձևեր։ Եթե ոսկրավոր ձկները իրոք հանկարծակի կերպով են երևացել հյուսիսային կիսագնդում կավճային ֆորմացիայի սկզբում, այդ շատ ուշագրավ փաստ կլիներ, բայց դա անհաղթահարելի դժվարություն կներկայացներ միայն այն դեպքում, եթե կարելի լիներ ապացուցել, որ հենց նույն ժամանակաշրջանում տեսակները հանկարծակի և միաժամանակ զարգացել են աշխարհի մյուս մասերում։ Գրեթե ավելորդ է նշել, որ հազիվ թե գոնե մի բրածո ձուկ գտնված լինի հասարակածի մյուս կողմում, և աչքի անցկացնելով Պիկտեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 207] «Հնէաբանությունը», կարելի է համոզվել, որ Եվրոպայի մի քանի ֆորմացիաներում հայտնի են բրածո ձկների շատ քիչ թվով տեսակներ։ Ձկների մի քանի սակավաթիվ ընտանիքներ այժմ ունեն սահմանափակ տարածում. ոսկրավոր ձկները կարող էին առաջներում նույնպես ունենալ սահմանափակ տարածում, և որևէ ծովում զգալի զարգացման հասնելուց հետո նրանք կարող էին լայնորեն տարածվել ամենուրեք։ Մենք ոչ մի իրավունք չունենք ենթադրելու, որ երկրագնդի վրա ծովերը միշտ էլ այնպես ազատ կերպով բաց են եղել հյուսիսից մինչև հարավ, ինչպես այժմ մենք տեսնում ենք։ Նույնիսկ այժմ էլ, եթե Մալայան արշիպելագը վերածվեր ցամաքի, Հնդկական օվկիանոսի արևադարձային մասը կառաջացներ մի ընդարձակ ու բոլորովին փակ ավազան, որի մեջ ծովային կենդանիների ամեն մի խոշոր խումբ կարող էր բազմանալ, մնալով ինքնապարփակ, մինչև որ նրա մի քանի տեսակները կհարմարվեին ավելի զով կլիմայի և հնարավորություն կստանային շրջանցելու Աֆրիկայի կամ Ավստրալիայի հարավային հրվանդանները և այսպիսով հասնելու այլ հեռավոր ծովեր։
Հիմնվելով այս կշռադատությունների վրա և ուշադրության առնելով մեր անտեղյակությունը Եվրոպայի և Միացյալ նահանգների սահմաններից դուրս գտնվող այլ երկրների երկրաբանության մասին, ինչպես նաև մեր հնէաբանական գիտելիքներում կատարված հեղաշրջումները, որոնք առաջացել են վերջին տասներկու տարիների հայտնագործություններից, ես կարծում եմ, որ չափազանց համարձակություն կլիներ լիակատար հեղինակավորությամբ դատել ամբողջ աշխարհում ձևերի հաջորդականության մասին, դա նման կլիներ այն բանին, ինչպես եթե որևէ բնախույզ հինգ րոպեով իջներ Ավստրալիայի ամայի ափը և հետո սկսեր դատել այդ մայր ցամաքի բնական արտադրանքների քանակի և հատկությունների մասին։
ԲՐԱԾՈ ՄՆԱՑՈՐԴՆԵՐ ՊԱՐՈՒՆԱԿՈՂ ՄԵԶ ՀԱՅՏՆԻ ԱՄԵՆԱՍՏՈՐԻՆ ՇԵՐՏԵՐՈՒՄ ԱԶԳԱԿԻՑ ՏԵՍԱԿՆԵՐԻ ԽՄԲԵՐԻ ՀԱՆԿԱՐԾԱԿԻ ԵՐԵՎԱՆ ԳԱԼՈՒ ՄԱՍԻՆ
Կա մի այլ նման դժվարություն ևս, և այն էլ ավելի լուրջ։ Այդ այն հանգամանքն է, որ կենդանական թագավորության գլխավոր զանազան բաժինների պատկանող տեսակները հանկարծակի երևան են գալիս այն ամենաստորին շերտերում, որոնց մեջ օրգանական մնացորդները հայտնի են։ Այն փաստարկների մեծ մասը, որոնք բերել են ինձ այն համոզման, որ միևնույն խմբի բոլոր գոյություն ունեցող տեսակները ծագել են մեկ նախածնողից, միանման ուժով կիրառելի է նաև մեզ հայտնի հնագույն տեսակների նկատմամբ։ Օրինակ, չի կարելի կասկածել նրանում, որ կեմբրիի և սիլուրի բոլոր տրիլոբիտները առաջացել են մի որևէ խեցեմորթից, որը պետք է գոյություն ունեցած լինի կեմբրիի դարից շատ առաջ և որը հավանորեն խիստ տարբերվել է բոլոր մեզ հայտնի կենդանիներից։ Ամենահին կենդանիներից մի քանիսներից օրինակ Nautilus-ը, Lingula-ն և ուրիշներ, շատ խիստ չեն տարբերվում այժմյան տեսակներից, և, մեր տեսության համաձայն, չի կարելի ենթադրել, որ այդ հին տեսակներն են եղել նախածնողներ բոլոր այդ խմբերին պատկանող տեսակների համար, որոնք երևացել են հետագայում, որովհետև նրանք ոչ մի տեսակետից իրենց հատկանիշներով միջակա ձևեր չեն հանդիսանում նրանց միջև։
Հետևաբար, եթե այս տեսությունը ճիշտ է, ապա ոչ մի կասկած չկա, որ նախքան ամենաստորին կեմբրիական շերտի կուտակվելը անցել են երկարատև ժամանակաշրջաններ, նույնքան երկարատև կամ հավանորեն ավելի ևս երկարատև են եղել, քան կեմբրիական դարի և մեր օրերի միջև ընկած ամբողջ ժամանակամիջոցը, և որ այդ հսկայական ժամանակաշրջանների ընթացքում աշխարհը լիքն էր կենդանի էակներով։ Այստեղ մենք հանդիպում ենք մի ահեղ առարկության, որովհետև կասկածելի է թվում, որ երկիրը բավականաչափ երկարատև ժամանակ գոյություն է ունեցել այն վիճակում, որը բարենպաստ էր կենդանի էակների համար։ Սըր Վ. Թոմսոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 211] գալիս է այն եզրակացության, որ երկրի կեղևի պնդացումը հազիվ թե կարող էր տեղի ունենալ 20 միլիոնից ոչ պակաս կամ 400 միլիոնից ոչ ավելի տարիներ առաջ և հավանորեն տեղի է ունեցել 98 միլիոնից ոչ պակաս և 200 միլիոնից ոչ ավելի տարիների ընթացքում։ Այդ խիստ լայն սահմանները ցույց են տալիս, թե տվյալներն իրենք որքան կասկածելի են, և հնարավոր է, որ հետագայում այդ խնդրի մեջ պետք է մտցվեն նաև այլ տարրեր։ Մ-ր Կրոլլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 194] կարծում է, թե կեմբրիի ժամանակաշրջանից մոտավորապես 60 միլիոն տարի է անցել, սակայն դատելով նրանից, որ սառցադաշտային դարաշրջանի սկզբից օրգանական ձևերը շատ քիչ են փոփոխվել, այդ ժամանակը շատ կարճ է թվում կյանքի այն շատ ու զգալի փոփոխությունների համար, որոնք անպայման տեղի են ունեցել սկսած կեմբրիի ֆորմացիայի ժամանակից և դրան նախորդած 140 միլիոն տարիները հազիվ թե կարելի է բավական համարել կյանքի այն բազմազան ձևերի զարգացման համար, որոնք արդեն գոյություն են ունեցել կեմբրիի ժամանակաշրջանում։ Սակայն հավանական է, ինչպես համառորեն նշում է նաև սըր Վիլյամ Թոմսոնը, որ աշխարհը շատ վաղ ժամանակամիջոցում իր ֆիզիկական պայմաններում կրել է ավելի արագ և ավելի խիստ փոփոխություններ, քան ներկայումս կատարվող փոփոխությունները։ Իսկ այդ փոփոխությունները պետք է առաջացնեին համապատասխան չափով փոփոխություններ նաև այն ժամանակ գոյություն ունեցող օրգան ական բնակչության մեջ։
Այն հարցին, թե ինչո՞ւ մենք չենք գտնում բրածոներով հարուստ նստվածքներ, որոնք վերաբերում են կեմբրիի սիստեմին նախորդած այդ ենթադրյալ հնագույն ժամանակաշրջաններին, ես բավարար պատասխան չեմ կարող տալ։ Շատ ականավոր երկրաբաններ, սըր Ռ. Մուրչիսոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 197] գլխավորությամբ, մինչև վերջին ժամանակները համոզված էին, որ սիլուրի ամենաստորին շերտի օրգանական մնացորդների մեջ մենք տեսնում ենք կյանքի առաջին արշալույսը։ Այլ իրազեկ դատավորներ, ինչպես Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] և Է. Ֆորբսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] առարկում էին այդ կարծիքի դեմ։ Մենք չպետք է մոռանանք, որ աշխարհի միայն մի փոքր մասն է հանգամանորեն ուսումնասիրված։ Ոչ այնքան վաղ Բարրանդն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 212] ավելացրել է նոր և յուրօրինակ տեսակներով մի ավելի ցածր հարկ այն հարկերի վրա, ինչպիսիք հայտնի են սիլուրի սիստեմում, իսկ այժմ մ-ր Հիկսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 213] հարավային Ուելսում գտել է շերտեր, որոնք հարուստ են տրիլոբիտներով և պարունակում են կակղամորթների ու օղակավորների բազմազան ձևեր դեռ ավելի ցածրում՝ ստորին կեմբրիի ֆորմացիայում։ Ֆոսֆորիտային սերտ աճած կտորների և խեժային նյութի ներկայությունը ազոյան ամենաստորին շերտերում հավանորեն ցույց է տալիս, որ կյանք գոյություն է ունեցել այդ ժամանակաշրջաններում։ Eozoon-ի գոյությունը Կանադայում լավրենտյան ֆորմացիայի մեջ հանրաճանաչ է։ Կանադայում սիլուրի սիստեմի տակ գոյություն ունեն շերտերի երեք մեծ սերիաներ, և Eozoon-ը գտնված է նրանցից ամենաստորինի մեջ։ Սըր Վ. Լոգանը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 214] հաստատում է, որ նրանց «հզորությունը ամբողջովին վերցրած թերևս շատ ավելի կգերազանցի պալեոզոյան սերիայի սկզբից մինչև հիմա եղած հետագա բոլոր ապարների հզորությանը։ Այսպիսով մենք վերադառնում ենք այնքան հեռավոր ժամանակաշրջաններ, որ այսպես կոչված մերձմորդիալ կենդանիների (Բարրանդի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 212]) երևան գալը կարելի է համարել համեմատաբար նոր եղելություն»։ Eozoon-ը պատկանում է կենդանիների ամենից ավելի ցածր կազմվածք ունեցող դասին, բայց իր դասի համար նա բարձր կազմվածք ունի. նա գոյություն է ունեցել անհամար թվով և, ինչպես նկատել է դ-ր Դոուսոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 196] անկասկած սնվել է այլ մանր օրգանական էակներով, որոնք հավանորեն նույնպես գոյություն են ունեցել հսկայական քանակությամբ։ Այսպիսով այն արտահայտությունները, որոնցով ես 1859 թ. հայտնել եմ, որ կենդանի էակները գոյություն են ունեցել կեմբրիի ժամանակաշրջանից շատ առաջ, և որոնք համարյա նույնն են, ինչ որ հետագայում ասել է սըր Վ. Լոգանը, ճիշտ դուրս եկան։ Այնուամենայնիվ, կեմբրիի սիստեմից ավելի ցած բրածոներով հարուստ հզոր շերտերի բացակայության համար որևէ հարմար բացատրություն գտնելու դժվարությունը դեռևս շատ մեծ է։ Շատ հավանական չի թվում, որ ամենահին շերտերը բոլորովին քայքայվել են դենուդացիայի հետևանքով կամ նրանց բրածոները իսպառ ոչնչացել են մետամորֆիզացիայի պրոցեսի շնորհիվ, որովհետև եթե այդ այդպես լիներ, մենք կունենայինք ըստ հասակի նրանց հետևից անմիջապես հաջորդող ֆորմացիաների միայն աննշան մնացորդները և նրանք միշտ կմնային մասամբ մետամորֆոզի ենթարկված վիճակում։ Սակայն Ռուսաստանի և Հյուսիսային Ամերիկայի ընդարձակ տերիտորիաների վրա սիլուրի նստվածքների մասին մեր ունեցած նկարագրությունները չեն հաստատում այղ ենթադրությունը, թե որքան ավելի հին է ֆորմացիան, այնքան նա ավելի է ենթարկվել դենուդացիայի և մետամորֆոզի պրոցեսներին։
Այդ փաստը առայժմ պետք է համարել անբացատրելի և իրավամբ կարելի է նրա վրա մատնանշել որպես ուժեղ առարկություն այստեղ պաշտպանվող տեսակետների դեմ։ Ցույց տալու համար, որ նա կարող է հետագայում որևէ բացատրություն ստանալ, ես հայտնում եմ հետևյալ հիպոթեզը։ Եվրոպայի և Միացյալ նահանգների զանազան ֆորմացիաներում գտնվող և ըստ երևույթին ոչ մեծ խորություններում ապրած օրգանիզմների բնույթի հիման վրա, ինչպես նաև այն նստվածքների հզորության հիման վրա, որոնցից կազմված են մղոններով հաստություն ունեցող ֆորմացիաները, մենք կարող ենք եզրակացնել, որ Եվրոպայի և Հյուսիսային Ամերիկայի այժմյան մայր ցամաքների մոտերքը ամբողջ ժամանակ գոյություն են ունեցել մեծ կղզիներ կամ ցամաքի տարածություններ, որոնք նյութ են մատակարարել նստվածքների առաջացման համար։ Միանգամայն նույնպիսի ենթադրություն են արել վերջերս Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] և ուրիշները։ Բայց մենք չգիտենք, թե ինչպես է եղել իրերի դրությունը հաջորդական զանազան ֆորմացիաների միջև ընկած ժամանակամիջոցում. արդյոք Եվրոպան և Միացյալ նահանգները այդ ժամանակամիջոցներում եղե՞լ են ցամաք, թե՞ առափնյա ստորջրյա մակերևույթ, որի վրա նստվածքներ չեն կուտակվել, թե՞ բաց ու անտակ ծովի հատակ։
Դառնալով այժմյան օվկիանոսներին, որոնք ցամաքի համեմատությամբ երեք անգամ ավելի մեծ տարածություն են գրավում, մենք տեսնում ենք, որ նրանք ծածկված են բազմաթիվ կղզիներով, բայց հազիվ թե հայտնի է գեթ մեկ իսկական օվկիանոսային կղզի (բացառությամբ Նոր Զելանդիայի, եթե նա կարող է անվանվել իսկական օվկիանոսային կղզի), որի վրա գոյություն ունենային պալեոզոյան կամ երկրորդային որևէ ֆորմացիաների գոնե մնացորդները։ Մենք կարող ենք ըստ երևույթին դրանից եզրակացնել, որ պալեոզոյան և երկրորդային ժամանակաշրջանների ընթացքում ոչ ցամաքներ, ոչ էլ կղզիներ են եղել այնտեղ, որտեղ այժմ տարածվում են մեր օվկիանոսները, որովհետև եթե նրանք գոյություն ունենային, այդ դեպքում պալեոզոյան և երկրորդային ֆորմացիաները ամենայն հավանականությամբ կառաջանային այն նստվածքների հաշվին, որոնք կգոյանային նրանց քայքայման ժամանակ. և այդ ֆորմացիաները պետք է գեթ մասամբ բարձրանային ծովի մակարդակից վեր երկրի մակերևույթի այն տատանումների ժամանակ, որոնք պետք է տեղի ունենային այդ չափազանց երկարատև ժամանակաշրջանների ընթացքում։ Հետևաբար, եթե մենք այս փաստերից կարող ենք որևէ եզրակացություն հանել, պետք է ընդունենք, որ այնտեղ, որտեղ այժմ տարածվում են մեր օվկիանոսները, վերջիններս գոյություն են ունեցել այն ամենահեռավոր ժամանկաշրջանից սկսած, որի մասին մենք որևէ տեղեկություն ունենք. մյուս կողմից, այնտեղ, որտեղ այժմ գտնվում են մեր մայր ցամաքները, գոյություն են ունեցել ցամաքի ընդարձակ տարածություններ, որոնք անպայման ենթարկվել են մակերևույթի մեծ տատանումների՝ սկսած կեմբրիի ժամանակաշրջանից։ Կորալյան ծովախութերի մասին իմ աշխատության կցված գունավոր քարտեզը բերում է ինձ այն եզրակացության, որ մեծ օվկիանոսներն այժմ էլ ներկայացնում են իջեցման գլխավոր մարզերը, մեծ արշիպելագները ներկայացնում են մակերևույթի տատանումների մարզեր, իսկ մայր ցամաքները՝ բարձրացման մարզեր։ Բայց մենք ոչ մի հիմք չունենք կարծելու, որ իրերի դրությունը աշխարհի սկզբից այդպես էլ մնացել է։ Մեր մայր ցամաքները հավանորեն գոյացել են այն բանի հետևանքով, որ երկրի մակերևույթի բազմաթիվ տատանումների ժամանակ գերակշռել է բարձրացման ուժը, բայց մի՞թե չէին կարող գերակշռող բարձրացման մարզերը դարերի ընթացքում փոխել իրենց տեղերը։ Կեմբրիի դարաշրջանից շատ առաջ եղած ժամանակաշրջանում մայր ցամաքները կարող էին գոյություն ունենալ այնտեղ, որտեղ այժմ տարածվում են օվկիանոսները և օվկիանոսները կարող էին գոյություն ունենալ այնտեղ, որտեղ այժմ գտնվում են մեր մայրցամաքները։ Եվ մենք հիմք չունենք ենթադրելու, որ եթե, օրինակ, Խաղաղ օվկիանոսի հատակը այժմ դառնար մայր ցամաք, մենք կարող էինք այնտեղ ճանաչել ավելի հին ֆորմացիաներ, քան կեմբրիյան շերտերը, կարծելով, որ այդ ֆորմացիաներն առաջ այնտեղ են գոյացել, որովհետև շատ հեշտությամբ կարող էր պատահել, որ այն շերտերը, որոնք մի քանի մղոն իջել են դեպի երկրի կենտրոնը և ենթարկվել են վերևում գտնվող ջրի հսկայական ճնշմանը, կարող էին շատ ավելի մեծ չափով կերպարանափոխվել, քան շարունակ մակերևույթին մոտիկ մնացած շերտերը։ Ինձ միշտ թվացել է, թե այն մետամորֆիկ ապարների հսկայական տարածությունները, որոնք մերկացած են աշխարհի զանազան մասերում, օրինակ Հարավային Ամերիկայում, պետք է ենթարկված լինեն տաքացման մեծ ճնշման տակ, պահանջում են որոշ յուրահատուկ բացատրություն, և թվում է, թե կարելի է կարծել, որ այդ ընդարձակ տարածությունները ներկայացնում են կեմբրիյանից շատ ավելի հին ֆորմացիաներ բոլորովին կերպարանափոխված և ողողման պրոցեսների ազդեցությամբ մերկացած։
Այստեղ մեր քննության առած դժվարությունները, այսինքն այն, որ թեև մենք մեր երկրաբանական ֆորմացիաներում գտնում ենք միացուցիչ օղակներ այն տեսակների միջև, որոնք այժմ գոյություն ունեն և առաջ գոյություն են ունեցել, մենք չենք գտնում նրանց միասին սերտ կերպով շաղկապող անհամար նուրբ անցումային ձևեր, որ եվրոպական ֆորմացիաներում տեսակների շատ խմբերի առաջին անգամ երևան գալը տեղի է ունենում հանկարծակի, որ կեմբրիի շերտերից ցած, ինչքան այժմ հայտնի է, գրեթե բոլորովին բացակայում են բրածոներով հարուստ ֆորմացիաները, այս բոլոր դժվարություններն անկասկած չափազանց լուրջ են։ Այդ երևում է հենց այն փաստից, որ ականավոր շատ հնէաբաններ, ինչպես Կյուվիեն,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 133] Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97], Բարրանդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 212], Պիկտեն,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 207] Ֆոկոները,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 73] Է. Ֆորբսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] և ուրիշներ, և մեր խոշորագույն երկրաբանները, ինչպես Լյաելը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] Մուրչիսոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 197] Սեջվիկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 209] և ուրիշներ, միաձայն ու ջերմ կերպով պաշտպանել են տեսակների անփոփոխելությունը։ Բայց այժմ սըր Չարլզ Լյաելը իր բարձր հեղինակությամբ աջակցություն է ցույց տալիս հակառակ կողմին, և շատ երկրաբաններ ու հնէաբաններ խիստ տատանվում են իրենց նախկին կարծիքում։ Նրանք, ովքեր կարծում են, թե երկրաբանական տարեգրությունը որքան և իցե լրիվ է, անկասկած իսկույն կժխտեն այս տեսությունը։ Ինչ վերաբերում է ինձ, ապա ես հետևելով Լյաելի այլաբանությանը, նայում եմ երկրաբանական տարեգրության վրա, որպես աշխարհի պատմության վրա, որը լրիվ չի պահպանվել, գրված է փոփոխված բարբառով, մի պատմություն, որի միայն վերջին հատորն ունենք, որ վերաբերում է երկու կամ երեք երկրների, այդ հատորից միայն այստեղ ու այնտեղ պահպանվել է մի համառոտ գլուխ և յուրաքանչյուր էջից անվնաս է մնացել տեղ-տեղ միայն մի քանի տող։ Դանդաղ կերպով փոփոխվող, հաջորդական գլուխներում ավելի կամ պակաս տարբեր բարբառի յուրաքանչյուր բառը ներկայացնում է կյանքի ձևեր, որոնք թաղված են մեր հաջորդական ֆորմացիաներում և որոնց մենք սխալ կերպով համարում ենք հանկարծակի երևացածներ։ Այսպիսի տեսակետից մոտենալով, վերևում քննարկված դժվարությունները զգալիորեն նվազում են կամ նույնիսկ անհետանում։
ԳԼՈՒԽ XI։ ՕՐԳԱՆԻԶՄՆԵՐԻ ԵՐԿՐԱԲԱՆԱԿԱՆ ՀԱՋՈՐԴԱԿԱՆՈՒԹՅԱՆ ՄԱՍԻՆ
ՆՈՐ ՏԵՍԱԿՆԵՐԻ ԴԱՆԴԱՂ ՈՒ ԱՍՏԻՃԱՆԱԿԱՆ ԵՐԵՎԱՆ ԳԱԼՈՒ ՄԱՍԻՆ։ ՆՐԱՆՑ ՓՈՓՈԽԵԼԻՈՒԹՅԱՆ ՏԱՐԲԵՐ ԱՍՏԻՃԱՆԻ ՄԱՍԻՆ։— ՄԻ ԱՆԳԱՄ ԱՆՀԵՏԱՑԱԾ ՏԵՍԱԿՆԵՐԸ ՆՈՐԻՑ ԵՐԵՎԱՆ ՉԵՆ ԳԱԼԻՍ։— ՏԵՍԱԿՆԵՐԻ ԽՄԲԵՐՆ ԻՐԵՆՑ ՀԱՅՏՆՎԵԼՈՒ ԵՎ ԱՆՀԱՅՏԱՆԱԼՈՒ ՄԵՋ ՀԵՏԵՎՈՒՄ ԵՆ ՆՈՒՅՆ ԿԱՆՈՆՆԵՐԻՆ, ԻՆՉ ՈՐ ԱՌԱՆՁԻՆ ՏԵՍԱԿՆԵՐԸ։— ՄԱՀԱՋՆՋՄԱՆ ՄԱՍԻՆ։— ԱՄԲՈՂՋ ԱՇԽԱՐՀՈՒՄ ԿՅԱՆՔԻ ՁԵՎԵՐԻ ՄԻԱԺԱՄԱՆԱԿՅԱ ՓՈՓՈԽՈՒԹՅԱՆ ՄԱՍԻՆ։— ՄԱՀԱՋՆՋՎԱԾ ՏԵՍԱԿՆԵՐԻ ՄԻՄՅԱՆՑ ՀԵՏ ԵՎ ԱՅԺՄ ԱՊՐՈՂ ՏԵՍԱԿՆԵՐԻ ՀԵՏ ՈՒՆԵՑԱԾ ԱԶԳԱԿՑՈՒԹՅԱՆ ՄԱՍԻՆ։ ՀԻՆ ՁԵՎԵՐԻ ԶԱՐԳԱՑՄԱՆ ԱՍՏԻՃԱՆԻ ՄԱՍԻՆ։— ԲՆԱԿՈՒԹՅԱՆ ՄԻԵՎՆՈՒՅՆ ՄԱՐԶԵՐԻ ՍԱՀՄԱՆՆԵՐՈՒՄ ՄԻԱՆՄԱՆ ՏԻՊԵՐԻ ՀԱՋՈՐԴԱԿԱՆ ԵՐԵՎԱՆ ԳԱԼՈՒ ՄԱՍԻՆ։— ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ ՆԱԽՈՐԴ ԵՎ ՆԵՐԿԱ ԳԼՈՒԽՆԵՐԻՑ։
Այժմ տեսնենք, թե ինչին ավելի շատ համապատասխանում են օրգանական էակների երկրաբանական հաջորդականության զանազան փաստերն ու օրենքները, արդյոք տեսակների անփոփոխելիության մասին ընդհանուրի կողմից ընդունված կարծիքին, թե այն տեսակետին, որ նրանք դանդաղ ու աստիճանաբար փոփոխվում են տարատեսակների առաջացման և բնական ընտրության միջոցով։
Նոր տեսակները երևացել են շատ դանդաղ կերպով, մեկը մյուսի հետևից թե՛ ցամաքի վրա և թե՛ ջրերի մեջ։ Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] ցույց է տվել, որ երրորդային զանազան հարկերի վերաբերմամբ հազիվ թե կարելի է դրանում կասկածել, տարեցտարի լրացվում են այդ հարկերի միջև եղած բաց տեղերը և մահաջնջված ու դեռ գոյություն ունեցող ձևերի քանակությունների միջև տարբերությունը ավելի ու ավելի նվազում է։ Մի քանի ավելի նոր, թեև անկասկած շատ հին շերտերում, եթե ժամանակը որոշելու լինենք տարիներով, մահաջնջվում են ընդամենը մեկ կամ երկու տեսակներ և լինում են միայն մեկ կամ երկու նոր տեսակ, որ երևան են գալիս այստեղ առաջին անգամ մի տեղում կամ, որքան մեզ հայտնի է, երկրի վրա ամենուրեք։ Երկրորդային ֆորմացիաների միջև ընդմիջումներն ավելի ցայտուն կերպով են արտահայտված. բայց ինչպես նկատել է Բրոննը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 136] յուրաքանչյուր ֆորմացիայում թաղված տեսակների բազմության ոչ երևան գալը և ոչ էլ անհետանալը հանկարծակի չի եղել։
Զանազան սեռերի և դասերի պատկանող տեսակները փոփոխվել են ոչ միանման արագությամբ և ոչ էլ միանման աստիճանով։ Հին երրորդային շերտերում բազմաթիվ մահաջնջված ձևերի մեջ կարելի է գտնել այժմ ապրող խեցիներից քչերին։ Ֆոկոները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 73] ցույց է տվել նման երևույթի մի զարմանալի օրինակ՝ դա այժմ ապրող կոկորդիլոսների համատեղ գոյությունն է, բազմաթիվ մահաջնջված կաթնասունների և սողունների հետ Հիմալայների նախալեռների նստվածքներում։ Սիլուրյան Lingula-ն միայն քիչ բանով է տարբերվում այդ սեռի այժմ ապրող տեսակներից, մինչդեռ սիլուրյան այլ կակղամորթների մեծ մասը և բոլոր խեցեմորթները մեծ չափով են փոփոխվել։ Ցամաքի բնակիչներն ըստ երևույթին ավելի արագ են փոփոխվել, քան ծովային օրգանիզմները, և դրա մի զարմանալի օրինակը վերջերս դիտվել է Շվեյցարիայում։ Հիմք կա կարծելու, որ բարձրակարգ կազմվածքով ձևերն ավելի արագ են փոփոխվում, քան ստորակարգները, թեպետ կան նաև բացառություններ այդ կանոնից։ Օրգանական աշխարհում փոփոխությունների աստիճանը, ինչպես նկատել է Պիկտեն,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 207] միևնույնը չէ յուրաքանչյուր հաջորդական, այսպես կոչված, ֆորմացիայում։ Սակայն եթե մենք համեմատենք ոչ ամենամերձավորների թվից որևէ ֆորմացիաներ իրար հետ, մենք կգտնենք, որ բոլոր տեսակները որոշ փոփոխություն են կրել։ Եթե տեսակը մի անգամ անհետացել է երկրի երեսից, մենք հիմք չունենք կարծելու, որ նույն ձևը երբևիցե նորից երևան կգա։ Այդ վերջին կանոնից ամենից ավելի նկատելի թվացող բացառությունը Բարրանդի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 212], այսպես կոչված, «գաղութներն» են, որոնք որոշ ժամանակով ներխուժում են մի ավելի հին ֆորմացիայի մեջ և հետո իրենց տեղը նորից զիջում են առաջուց գոյություն ունեցած կենդանական աշխարհին։ Բայց այդ փաստին Լյաելի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] տված բացատրությունը, այսինքն, որ դա մի այլ աշխարհագրական գավառից ժամանակավոր ներգաղթի դեպք է, բավարար է թվում։
Այդ զանազան փաստերը լավ համապատասխանում են իմ տեսությանը, որը չի ենթադրում զարգացման որոշակի օրենքներ, որոնք պահանջում են, որպեսզի երրորդային մարզի բոլոր բնակիչները փոփոխվեին հանկարծակի կամ միաժամանակ, կամ միանման չափով։ Փոփոխության պրոցեսը պետք է ընթանար դանդաղ կերպով և տարածվեր միաժամանակ միայն քիչ տեսակների վրա, որովհետև յուրաքանչյուր տեսակի փոփոխելիությունը կախված չէ մյուս բոլոր տեսակների փոփոխելիությունից։ Բնական ընտրությունը մեծ կամ փոքր չափով կկուտակի՞ արդյոք այդ փոփոխությունները կամ առաջացող անհատական տարբերությունները, դրանով առաջ բերելով ավելի կամ պակաս կայուն փոփոխություններ, դա կախված կլինի բազմաթիվ և բազմազան պատահականություններից, այսինքն նրանից, թե արդյոք օգտակար է փոփոխությունը, խաչաձևման ազատությունից, երկրի ֆիզիկական պայմանների դանդաղ փոփոխությունից, տվյալ երկիրը նոր գաղթականների գալուստից և մյուս բնակիչների առանձնահատկություններից, որոնց հետ մրցման մեջ են մտնում փոփոխվող տեսակները։ Ուստի բոլորովին զարմանալի չէ, որ որևէ տեսակ պահպանվում է իր անփոփոխ ձևով ավելի երկար, քան մյուս տեսակները, կամ եթե նույնիսկ փոփոխվում է, ապա պակաս չափով։ Նման հարաբերություններ մենք նկատում ենք զանազան երկրների այժմ գոյություն ունեցող բնակիչների միջև․ օրինակ, Մադեյրայի ցամաքային խեցիները և կարծրաթև միջատները զգալիորեն շեղվել են Եվրոպայի մայր ցամաքի վրա ապրող իրենց մերձավոր ազգակիցներից, մինչդեռ ծովային խեցիները և թռչունները մնացել են անփոփոխ։ Ցամաքային և բարձր կազմակերպված ձևերի ավելի արագ փոփոխելիությունը՝ համեմատած ծովային ու կազմվածքով ավելի ցածր ձևերի հետ, մենք կարող ենք ըստ երևույթին բացատրել կյանքի օրգանական և անօրգանական պայմանների հետ բարձր ձևերի ունեցած ավելի բարդ փոխհարաբերություններով, ինչպես այդ պարզաբանված է նախորդ գլուխներից մեկում։ Երբ որևէ մարզի բնակիչներից շատերը փոփոխվել կամ կատարելագործվել են, մենք կարող ենք ըմբռնել, որ գոյության կռվում օրգանիզմների միջև, եղած մրցման ու անխուսափելի փոխհարաբերությունների սկզբունքի հետևանքով յուրաքանչյուր ձև, որը չի ենթարկվել որոշ փոփոխության և կատարելագործության, դատապարտված է մահաջնջման։ Դա բացատրում է մեզ, թե ինչու միևնույն մարզի բոլոր տեսակները, եթե մենք նրանց ուսումնասիրում ենք բավականաչափ երկար ժամանակամիջոցներից հետո, ի վերջո փոփոխված են լինում, որովհետև առանց փոփոխվելու նրանք պետք է անհետանային։
Միևնույն դասի ներկայացուցիչների մոտ փոփոխելիության միջին հանրագումարը երկարատև ու հավասար ժամանակաշրջանների ընթացքում ըստ երևույթին կարող է գրեթե միանման լինել, բայց քանի որ երկար ժամանակ պահպանվող և բրածոներով հարուստ ֆորմացիաների կուտակումը պայմանավորվում է նրանով, որ նստվածքների մեծ զանգվածները կուտակվում են իջվածքների մարզերում, ապա համարյա մեր բոլոր ֆորմացիաները պետք է կուտակվեին ժամանակի մեծ և անկանոն ընդմիջումներից հետո. հետևաբար և օրգանական կյանքի փոփոխությունների ընդհանուր հանրագումարը, որը դրսևորում են հաջորդական ֆորմացիաներում թաղված բրածոները, նույնը չէ։ Այդ տեսակետից յուրաքանչյուր ֆորմացիա ստեղծագործական նոր և լրիվ ակտ չի ներկայացնում, այլ միայն մի պատահական տեսարան, որը համարյա դիպվածաբար վերցված է դանդաղ ու անընդհատ փոփոխվող դրամայից։
Դժվար չէ հասկանալ, թե ինչու մի անգամ անհետացած տեսակը երբեք չի կարող նորից երևան գալ, եթե նույնիսկ նորից կրկնվեին կյանքի թե օրգանական և թե անօրգանական միանգամայն նույնանման պայմանները։ Եթե որևէ տեսակի հաջորդը թեկուզ կարողացել է (այդ բանը, անկասկած, հաճախ էլ տեղի է ունեցել) հարմարվել այնպես, որպեսզի ի վիճակի լիներ բնության տնտեսության մեջ գրավելու մի այլ տեսակի տեղը և այդպիսով դուրս մղել նրան, այնուամենայնիվ երկու ձևերն էլ թե՛ հինը և թե՛ նորը լիովին նույնանման չէին լինի, որովհետև նրանք երկուսն էլ գրեթե անկասկածելիորեն պիտի ժառանգեին իրենց զանազան նախածնողների զանազան հատկանիշներ, իսկ զանազան օրգանիզմներն էլ արդեն կփոփոխվեին տարբեր ձևով։ Օրինակ, հնարավոր է, որ եթե մեր բոլոր սիրամարգանման աղավնիները անհետանային, աղավնաբույծները կստեղծեին ներկայումս եղածից համարյա չտարբերվող մի նոր ցեղ. բայց եթե անհետանար նաև նախասկզբնական ձևը ժայռաղավնին (իսկ մենք բոլոր հիմքերն ունենք կարծելու, որ բնության մեջ նախասկզբնական ձևերը սովորաբար դուրս մղվում ու ոչնչացվում են իրենց կատարելագործված հետնորդների կողմից), այդ դեպքում անհավատալի է. որ հնարավոր լիներ որևէ այլ աղավնատեսակից կամ նույնիսկ ընտանի աղավնիների որևէ այլ որոշակի տեսակից առաջ բերել գոյություն ունեցողին նման սիրամարգանման աղավնի, որովհետև հաջորդական փոփոխությունները համարյա հաստատապես մի քիչ տարբեր կլինեին և նոր ստեղծված տարատեսակը հավանորեն կժառանգեր իր նախածնողից որոշ բնորոշ առանձնահատկություններ։
Տեսակների խմբերը, այսինքն սեռերն ու ընտանիքները իրենց երևան գալու և անհետանալու ընթացքում ղեկավարվում են նույն կանոններով, ինչ կանոններով որ ղեկավարվում են առանձին տեսակները, փոփոխվելով ավելի կամ պակաս արագությամբ և մեծ կամ փոքր չափով։ Մի անգամ անհետացած խումբը երբեք կրկին երևան չի գալիս. այլ խոսքով՝ նրա գոյությունը քանի դեռ շարունակվում է, այնքան ժամանակ անընդհատ է։ Ես գիտեմ, որ այս կանոնից կան որոշ առերևույթ բացառություններ, բայց նրանք չափազանց քիչ են, այնքան քիչ, որ Է. Ֆորբսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] Պիկտեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 207] և Վուդվորդն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 205] ընդունում են կանոնի ճշտությունը, չնայած որ նրանք բոլորն էլ խիստ հակառակ են իմ պաշտպանած հայացքներին․ իսկ այդ կանոնը խիստ կերպով համապատասխանում է իմ տեսությանը, որի իմաստի համաձայն միևնույն խմբի բոլոր տեսակները ինչքան էլ երկար ժամանակ խումբը գոյություն ունենա, առաջացել են մեկը մյուսից աստիճանական փոփոխությունների միջոցով, իսկ բոլորը միասին առաջացել են մի ընդհանուր նախածնողից։ Օրինակ, Lingula սեռի մեջ բոլոր տեսակները, որոնք հաջորդաբար երևացել են զանազան դարաշրջաններում, պետք է շաղկապված լինեն մեկը մյուսի հետ սերունդների հաջորդական շարքով, որը չի ընդհատվել սկսած սիլուրի ամենաստորին շերտի կուտակման ժամանակից մինչև ներկա ժամանակ։
Մենք նախորդ գլխում տեսանք, որ տեսակների ամբողջ խմբեր երբեմն թվում են հանկարծակի երևան եկած, և ես աշխատել եմ բացատրություն տալ այդ փաստին, որ եթե ճիշտ լիներ, նա ճակատագրական նշանակություն կունենար իմ տեսության համար։ Բայց նման դեպքերն անկասկած բացառություն են, իսկ ընդհանուր կանոն է տեսակների թվի աստիճանական ավելանալը, մինչև որ խումբը հասնում է իր գագաթնակետին, որից հետո վաղ թե ուշ հետևում է նրա աստիճանական պակասումը։ Եթե որևէ սեռի տեսակների թիվը կամ որևէ ընտանիքի սեռերի թիվը պատկերենք ոչ միանման հաստություն ունեցող ուղղաձիգ գծով, որը բարձրանում է երկրաբանական հաջորդական այն ֆորմացիաների միջով, որտեղ այդ տեսակները գտնվում են, այդ դեպքում կարող է երբեմն սխալ կերպով թվալ, թե այդ գիծը ներքևում սկսվում է ոչ թե սուր ծայրով, այլ միանգամից ընթանում է ցայտուն գծով. հետո նա դեպի վեր աստիճանաբար հաստանում է, երբեմն որոշ տարածության վրա պահպանելով միանման հաստություն, և վերջապես վերին շերտերում բարակում է, դրանով արտահայտելով տեսակների նվազումը և վերջնական անհայտացումը։ Խմբի մեջ տեսակների թվի այդպիսի աստիճանական աճումը ամբողջովին համապատասխանում է իմ տեսությանը, որովհետև միևնույն սեռի տեսակները և միևնույն ընտանիքի սեռերը կարող են բազմանալ միայն դանդաղորեն ու աստիճանաբար, քանի որ փոփոխության և ազգակից շատ ձևերի առաջացման պրոցեսն անկասկած պետք է լինի դանդաղ ու աստիճանական, այսինքն՝ մի տեսակ ամենից առաջ սկիզբ է տալիս երկու կամ երեք տարատեսակների, վերջինները դանդաղ կերպով դառնում են տեսակներ, որոնք իրենց հերթին նույն դանդաղ ճանապարհով առաջացնում են այլ տարատեսակներ ու տեսակներ, և այդպես շարունակ, նման մի մեծ ծառի, որը ճյուղավորվում է մի ընդհանուր բնից, մինչև որ վերջապես խումբը հասնում է լայնածավալ զարգացման։
ՄԱՀԱՋՆՋՄԱՆ ՄԱՍԻՆ
Մինչև այժմ մենք ի միջի այլոց ենք խոսել տեսակների և նրանց խմբերի մահաջնջման մասին։ Բնական ընտրության տեսության համաձայն հին ձևերի մահաջնջվելը և նոր ու կատարելագործված ձևերի երևան գալը սերտ կերպով կապված են միմյանց հետ։ Նախկին ուսմունքը, թե երկրի ամբողջ բնակչությունը ոչնչացել է պարբերաբար կրկնվող աղետներով, այժմ լքված է գրեթե բոլոր, նույնիսկ այնպիսի երկրաբանների կողմից, ինչպես Էլի դը-Բոմոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 215] Մուրչիսոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 197] Բարրանդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 212] և ուրիշներ, որոնց ընդհանուր հայացքները պետք է որ բնականորեն բերեին այդպիսի եզրակացության։ Ընդհակառակը, հիմնվելով երրորդային ֆորմացիաների ուսումնասիրության վրա, մենք իրավունք ունենք կարծելու, որ տեսակները և տեսակների խմբերը աստիճանաբար մեկը մյուսի հետևից անհետանում են՝ սկզբում մի տեղում, հետո մի ուրիշ և վերջապես ամենուրեք երկրի վրա։ Ասենք՝ որոշ հազվագյուտ դեպքերում, օրինակ մի պարանոցի պատռվելու դեպքում և դրա հետևանքով հարևան ծովի մեջ բազմաթիվ նոր բնակիչների ներխուժման ժամանակ կամ որևէ կղզու վերջնական խորասուզման ժամանակ մահաջնջման պրոցեսը կարող էր արագ կատարվել։ Ինչպես առանձին տեսակների, այնպես էլ տեսակների ամբողջ խմբերի գոյության տևողությունը շատ տարբեր է. շատ խմբեր, ինչպես մենք տեսանք, գոյություն ունեին կյանքի մեզ հայտնի առաջին նշույլների երևալու ժամանակից սկսած մինչև մեր օրերը, մյուսները անհետացել են դեռ մինչև պալեոզոյան էրայի վերջանալը։ Ըստ երևույթին ճշտորոշ օրենք չկա, որով սահմանվեր առանձին տեսակի կամ առանձին սեռի գոյության տևողությունը։ Հիմք կա կարծելու, որ տեսակների մի ամբողջ խմբի մահաջնջումը ավելի դանդաղ է ընթանում, քան նրանց առաջացումը, եթե նրանց հայտնվելը և անհետանալը պատկերացնենք, ինչպես այդ արել ենք առաջ, անհավասար հաստություն ունեցող ուղղաձիգ գծով, ապա այդ գիծը իր վերին ծայրում, որը պատկերում է մահաջնջման ընթացքը, կնեղանա ավելի աստիճանաբար, քան ստորին ծայրում, որը ներկայացնում է տեսակների առաջին անգամ հայտնվելը և նրանց թվի սկզբնական աճումը։ Սակայն որոշ դեպքերում տեսակների ամբողջ խմբերի, օրինակ երկրորդային ժամանակաշրջանի վերջին ամմոնիտների անհետացումը տեղի է ունեցել զարմանալիորեն արագ կերպով։
Տեսակների անհետացման պրոցեսը թվում էր անըմբռնելի գաղտնիքով սքողված։ Մի քանի հեղինակներ նույնիսկ ենթադրել են, որ ինչպես անհատի կյանքն ունի միայն որոշակի տևողություն, այնպես էլ տեսակները գոյություն ունեն միայն մի որոշակի ժամանակ։ Տեսակների մահաջնջումը ինձ ավելի զարմանալի էր թվում, քան որևէ մեկ ուրիշին։ Երբ ես Լա-Պլատայում գտա ձիու մի ատամ, որը գտնվում էր մաստոդոնտի, մեգաթերիայի, տոքսոդոնտի և այլ մահաջնջված հրեշների մնացորդների հետ միասին, որոնք բոլորն էլ վերջին երկրաբանական ժամանակաշրջաններից մեկում գոյություն են ունեցել այժմ ևս գոյություն ունեցող խեցիների հետ միաժամանակ, դա ինձ վերին աստիճանի զարմացրեց, որովհետև ես գիտեի, որ ձին այն ժամանակից սկսած, երբ իսպանացիները նրան բերել են Հարավային Ամերիկա, այնտեղ վայրենացել է և բազմացել ամենուրեք անսովոր չափով, և ես ինձ հարց էի տալիս, թե ինչ պատճառ կարող էր այդքան նոր ժամանակներում այդտեղ առաջացնել նախկին ձիու անհետացումբ, երբ կյանքի պայմանները ակնհայտորեն բարենպաստ են։ Բայց իմ զարմանքը անհիմն էր։ Պրոֆ. Օուենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] շուտով նկատեց, որ թեև այդ ատամը շատ նման է այժմյան ձիու ատամին, բայց պատկանում է մահաջնջված մի տեսակի։ Եթե այդ ձին մինչև այժմ գոյություն ունենար, բայց լիներ բավական հազվագյուտ, ոչ մի բնախույզի դա տարօրինակ չէր թվա, որովհետև բոլոր երկրներում, բոլոր դասերի տեսակների ահագին քանակություն պատկանում է հազվագյուտների թվին։ Երբ մենք հարց ենք տալիս, թե ինչո՞ւ այս կամ այն տեսակը հազվագյուտ է, մենք գտնում ենք պատասխանը, որ նրա կենսապայմաններում ինչ որ անբարենպաստ բան կա. բայց ի՞նչ է այդ ինչ-որ բանը, մենք համարյա երբեք չենք կարող ասել։ Եթե այդ բրածո ձին մինչև այժմ գոյություն ունենար որպես հազվագյուտ տեսակ, մենք կարող էինք հավատացած լինել, մյուս բոլոր կաթնասունների անալոգիայով, չբացառելով նաև դանդաղ բազմացեղ փղին, ինչպես նաև գիտենալով ընտանի ձիու նատուրալիզացիայի պատմությունը Հարավային Ամերիկայում, որ ավելի բարենպաստ պայմաններում նա շատ կարճ ժամանակում կլցներ ամբողջ մայր ցամաքը։ Բայց մենք չէինք կարողանա ասել, թե ինչպիսին են եղել այն անբարենպաստ պայմանները, որոնք արգելք են եղել նրա զարգացմանը, արդյոք այստեղ ներգործել է որևէ մե՞կ պատճառ, թե շատ պատճառներ, ձիու կյանքի ի՞նչ շրջանում և ի՞նչ չափով գործել են նրանք։ Եթե այդ պայմանները, թեև դանդաղ կերպով, շարունակ ավելի ու ավելի անբարենպաստ են դարձել, մենք հավանորեն չէինք նկատի այդ հանգամանքը, մինչդեռ բրածո ձին ավելի ու ավելի հազվադեպ կհանդիպեր ու վերջապես կանհետանար, իսկ նրա տեղը կգրավեր որևէ ավելի բախտավոր մրցակից։
Դժվար է միշտ մտքում պահել այն հանգամանքը, որ յուրաքանչյուր կենդանի էակի բազմացումը միշտ կասեցվում է աննկատելի թշնամական պատճառներով և որ այդ աննկատելի պատճառները միանգամայն բավական են, որպեսզի տեսակը դառնա սակավաթիվ և վերջապես իսպառ մահաջնջվի։ Այս հարցերն այնքան քիչ են պարզաբանված, որ հաճախ ես լսել եմ, թե ինչպես զարմանք են հայտնել այն առթիվ, որ մահաջնջվել են այնպիսի խոշոր հրեշներ, ինչպես մաստոդոնտը և ավելի ևս հին դինոզավրները, կարծես թե միայն մարմնի ամրությունը կարող է հաղթանակ ապահովել գոյության կռվում։ Ընդհակառակը, մարմնի մեծ չափերով որոշ դեպքերում, ինչպես այդ նկատել է Օուենը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] պայմանավորվում է ավելի արագ անհետացումը, որովհետև խոշորության դեպքում պահանջվում է մեծ քանակությամբ կերակուր։ Հնդկաստանում կամ Աֆրիկայում մարդու երևան գալուց առաջ երևի ինչ որ պատճառ է եղել, որը կասեցրել է այժմ ապրող փղի անընդհատ բազմացումը։ Այդ հարցում շատ իրազեկ դատավոր դոկտոր Ֆոկոները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 73] կարծում է, որ Հնդկաստանում փղի բազմացմանը խոչընդոտ են հանդիսանում գլխավորապես միջատները, որոնք անդադար ուժասպառ են անում և թուլացնում նրան, նույնպիսի եզրակացության է եկել Բրուսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 216] աֆրիկական փղի վերաբերմամբ Հաբեշստանում։ Անկասկած է, որ միջատները և վամպիրները վճռական ազդեցություն ունեն Հարավային Ամերիկայի զանազան մասերում նատուրալիզացիայի ենթարկված ավելի խոշոր չորքոտանիների գոյության վրա։
Ուսումնասիրելով երրորդային ավելի նոր նստվածքները, մենք շատ դեպքերում նկատում ենք, որ տեսակների սակավաթվությունը նախորդում է մահաջնջմանը, և մենք գիտենք, որ միևնույնն է նկատվել նաև այն կենդանիների վերաբերմամբ, որոնք տեղ-տեղ կամ իսպառ ոչնչացվել են մարդու գործունեությունից։ Ես այստեղ կարող եմ կրկնել այն, ինչ գրել եմ 1845 թվականին, այն է՝ ընդունել, որ տեսակն իր անհայտանալուց առաջ սովորաբար սակավաթիվ է դառնում, չզարմանալ տեսակների սակավաթվության վրա, իսկ հետո խիստ զարմանալ, որ տեսակը դադարում է գոյություն ունենալուց, դա միևնույնն է, թե ընդունել, որ հիվանդությունը նախորդում է անհատի մահվանը, և չզարմանալ, լսելով հիվանդության մասին, իսկ հետո, երբ հիվանդ մարդը մեռել է, զարմանալ դրա վրա և կասկածել, որ մահվան պատճառը եղել է բռնության գործողություն։
Բնական ընտրության տեսությունը հիմնվում է այն դրույթի վրա, որ յուրաքանչյուր նոր տարատեսակ և վերջ ի վերջո յուրաքանչյուր նոր տեսակ առաջանում է և մնում է շնորհիվ այն հանգամանքի, որ ունենում է որևէ առավելություն նրա հանդեպ, որի հետ նա ընդհարման մեջ է մտնում, դրան իր գրեթե անխուսափելիորեն հետևում է ավելի պակաս հաջողակորեն կազմակերպված ձևերի մահաջնջում։ Նույնը նկատվում է նաև մեր ընտանի ձևերի վերաբերմամբ։ Հենց որ ստացվում է նոր, փոքր ինչ ավելի լավ տարատեսակ, նա դուրս է մղում իր հարևան ավելի նվազ կատարելագործված տարատեսակներին. ավելի ևս կատարելագործված լինելով, նա տարածվում է ամենուրեք, մեր շորտհորն (կարճաեղջյուր) անասունների նման, և իրենով փոխարինում է նախկին ցեղերին նաև այլ երկրներում։ Հետևաբար նոր ձևերի հայտնվելը և նախկինների անհետանալը, թեկուզ ստացվել են այդ ձևերը բնական ճանապարհով, թե արհեստականորեն, սերտ կերպով կապված են միմյանց հետ։ Բարգավաճող վիճակում գտնվող խմբերում որոշ ժամանակի ընթացքում առաջացող նոր տեսակային ձևերի քանակությունը որոշ ժամանակաշրջաններում ըստ երևույթին եղել է ավելի մեծ, քան նախկին, անհետացած տեսակային ձևերի քանակությունը. բայց մենք գիտենք, որ տեսակների թիվը, գոնե նորագույն երկրաբանական դարաշրջաններում, անսահման կերպով չի ավելացել, այնպես որ նորագույն ժամանակների վերաբերմամբ մենք կարող ենք կարծել, որ նոր ձևերի առաջացում պատճառել է համարյա նույն քանակությամբ նախկին ձևերի մահաջնջում։
Ինչպես ավելի առաջ պարզաբանված և օրինակներով լուսաբանված է, որ մրցումն ավելի համառ է լինում այն ձևերի միջև, որոնք բոլոր տեսակետներից ավելի մոտ են միմյանց, այդ պատճառով որևէ տեսակի փոփոխված ու կատարելագործված հաջորդները սովորաբար ոչնչացնում են նախահայր տեսակին, իսկ եթե որևէ մի տեսակից զարգացել են մի քանի նոր ձևեր, այն ժամանակ նրան մերձավոր, այսինքն նրա հետ միասին մի սեռի պատկանող տեսակները ամենից ավելի մեծ չափով են ենթարկվում ոչնչացման։ Հատկապես այդ եղանակով, ինչպես ես կարծում եմ, մի տեսակից ծագում առած նոր տեսակների խումբը, այսինքն նոր սեռը, կարող է դուրս մղել միևնույն ընտանիքին պատկանող նախկին սեռը։ Բայց հաճախ պիտի պատահեր, որ մի որևէ խմբի պատկանող նոր տեսակ գրավել է մի այլ խմբի տեսակին պատկանող տեղը և դրանով էլ առաջ բերել նրա ոչնչացումը։ Եթե մերձավոր շատ ձևեր զարգանում են այդպիսի հաղթական մրցակցից, այն ժամանակ նաև այլ շատ ձևեր պետք է իրենց տեղը զիջեն նրանց և դրանք կլինեն սովորական ձևեր, առաջուց դուրս մղված ձևին ամենամոտ կանգնածներ, որոնք բոլորը միասին երևան կհանեն որևէ ժառանգված անկատարելություն իրենց կազմվածքում։ Բայց արդյոք այդ տեսակները, որ իրենց տեղերը զիջել են փոփոխված ու կատարելագործված այլ տեսակներին, կպատկանեն միևնույն դասին, թե մի այլ դասի։ Զիջողների թվից մի քանիսները երբեմն կարող են երկար ժամանակ պահպանվել շնորհիվ այն բանի, որ նրանք գրավում են հեռավոր ու մեկուսացած մի մարզ, որտեղ նրանք խուսափել են ուժեղ մրցակցությունից։ Օրինակ, երկրորդային ֆորմացիաներում լայնորեն տարածված խեցիների խոշոր սեռի՝ Trigonia-ի որոշ տեսակները պահպանվել են ավստրալիական ծովերում, գանոիդ ձկների մեծ և համարյա մահաջնջված խմբի քիչ ներկայացուցիչներ մինչև հիմա ապրում են մեր քաղցրահամ ջրերում։ Հետևաբար, մենք տեսնում ենք, որ որևէ խմբի վերջնական անհետացումը սովորաբար տեղի է ունենում ավելի դանդաղ ճանապարհով, քան նրա առաջացումը։
Ինչ վերաբերում է ամբողջ ընտանիքների կամ կարգերի, օրինակ պալեոզոյան ժամանակաշրջանի վերջին տրիլոբիտների, երկրորդային ժամանակաշրջանի վերջին ամմոնիտների թվացող հանկարծակի անհետացմանը, մենք պետք է այստեղ վերհիշենք այն, ինչ որ արդեն ասվել է հնարավոր մեծ ընդմիջումների մասին, որոնք բաժանում են մեր հաջորդական ֆորմացիաները միմյանցից. այդ ընդմիջումների ընթացքում կարող էին տեղի ունենալ դանդաղ մահաջնջման բազմաթիվ դեպքեր։ Բացի դրանից, եթե նոր գաղթականների հանկարծակի երևան գալու կամ արտասովոր արագությամբ բազմանալու հետևանքով որևէ նոր խմբի շատ տեսակներ գրավել են մի որոշ մարզ, այդ դեպքում նախկին տեսակների թվից շատերը պետք է համապատասխանորեն արագ կերպով անհետանան. իրենց տեղը այս կերպով զիջող ձևերը սովորաբար միմյանց մոտ են, ունենալով իրենց կազմվածքում որևէ ընդհանուր անկատարություն։
Ուրեմն՝ առանձին տեսակի և տեսակների ամբողջ խմբերի անհետացման եղանակը, ինչպես ինձ թվում է, լավ համաձայնվում է բնական ընտրության տեսության հետ։ Մահաջնջման փաստը մեզ ոչնչով չպետք է զարմացնի, եթե մի բան էլ կա զարմանալու, դա մեր ինքնավստահությունն է, որը մեզ թույլ է տալիս երևակայելու, թե մենք ըմբռնում ենք այն պատահականությունների ամբողջ միագումարությունը, որոնցից կախված է յուրաքանչյուր տեսակի գոյությունը։ Եթե մենք գեթ մի րոպե մոռանանք, որ յուրաքանչյուր տեսակ ձգտում է անսահմանորեն բազմանալ և որ միշտ որևէ խոչընդոտ դրան հակառակ է գործում, թեև մենք այդ խոչընդոտները հազվագյուտ դեպքում ենք նկատում, ապա բնության ամբողջ կարգ ու կանոնը մեզ անհասկանալի կդառնա։ Միայն այն ժամանակ, երբ մենք ի վիճակի կլինենք ճշգրտորեն ցույց տալու, թե ինչու այսինչ տեսակը ավելի հարուստ է անհատներով, քան մյուսը, թե ինչու այս, այլ ոչ թե մյուս որևէ տեսակ կարող է նատուրալիզացիայի ենթարկվել տվյալ երկրում, միայն այդ ժամանակ, բայց ոչ ղրանից առաջ, մենք իրավունք կունենանք զարմանալու, որ մենք չենք հասկանում որևէ առանձին տեսակի կամ տեսակների խմբերի անհետացման պատճառները։
ԳՐԵԹԵ ԱՄԲՈՂՋ ԵՐԿՐԱԳՆԴԻ ՎՐԱ ԿՅԱՆՔԻ ՁԵՎԵՐԻ ՄԻԱԺԱՄԱՆԱԿՅԱ ՓՈՓՈԽՈՒԹՅԱՆ ՄԱՍԻՆ
Հազիվ թե ամենազարմանալի հնէաբանական հայտնագործությունը պետք է համարել այն փաստը, որ կյանքի ձևերը երկրագնդի վրա գրեթե միաժամանակ են փոփոխվում։ Մեր եվրոպական կավճային ֆորմացիան կարելի է որոշել ըստ բրածոների չափազանց տարբեր կլիմա ունեցող միմյանցից խիստ շատ հեռու գտնվող մարզերում, նույնիսկ այնտեղ, որտեղ չի կարելի գտնել կավճի, որպես ապարի ոչ մի բեկոր, այն է՝ Հյուսիսային Ամերիկայում, հասարակածային Հարավային Ամերիկայում, Հրո երկրում, Բարեհուսո հրվանդանում և Արևելյան Հնդկաստանում։ Այդ հեռավոր կետերում որոշ շերտերի մեջ գտնված օրգանական մնացորդները ցուցաբերում են անկասկածելի նմանություն այն օրգանական մնացորդների հետ, որոնք բնորոշ են կավճի համար։ Չի կարելի ասել, որ այնտեղ մենք հանդիպում ենք միևնույն տեսակներին, որոշ դեպքերում ոչ մի միանգամայն նույնանման տեսակ չկա, բայց նրանք պատկանում են միևնույն սեռերին ու ենթասեռերին և հաճախ նմանություն են ցուցաբերում նույնիսկ այնպիսի քիչ կարևոր հատկանիշներում, ինչպիսին է խեցիների մակերեսի քանդակը։ Բացի դրանից, այլ ձևերը, որոնք չեն գտնվում եվրոպական կավճի մեջ, բայց հանդիպում են կամ վերևում կամ ներքևում գտնվող ֆորմացիայում, երկրագնդի այդ հեռավոր կետերում միևնույն կարգով են դասավորված։ Զանազան հեղինակներ Ռուսաստանում, Արևմտյան Եվրոպայում և Հյուսիսային Ամերիկայում պալեոզոյան խմբի զանազան հաջորդական ֆորմացիաներում նկատել են կյանքի օրգանական ձևերի նման զուգահեռություն, նույնն էլ, Լյաելի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] խոսքերով, նկատվում է Եվրոպայի և Հյուսիսային Ամերիկայի երրորդային նստվածքներում։ Եթե նույնիսկ ուշադրություն չդարձնենք այն սակավաթիվ բրածո տեսակների վրա, որոնք ընդհանուր են Հին և Նոր աշխարհներին, այնուամենայնիվ ակներև կլինի երկու երկրների պալեոզոյան և երրորդային հարկերում կյանքի հաջորդական ձևերի ընդհանուր զուգահեռությունը և զանազան ֆորմացիաները հեշտությամբ կարող են համեմատվել իրար հետ։
Սակայն այս դիտողությունները վերաբերում են երկրագնդի ծովային բնակիչներին, մենք բավականաչափ տվյալներ չունենք, որպեսզի դատենք այն մասին, թե արդյոք ցամաքի և քաղցրահամ ջրերի բնակիչները այդքան մեկը մյուսից հեռավոր վայրերում ներկայացնում են նույնպիսի զուգահեռություն իրենց փոփոխություններում։ Հիմք կա կասկածելու դրանում, որ նրանք փոփոխվում են այդպիսի ձևով, եթե Megatherium-ը, Mylodon-ը, Macrauchenia-ն և Toxodon-ը Լա Պլատայից Եվրոպա բերված լինեին առանց նշելու նրանց երկրաբանական դրությունը, ոչ ոք չէր կասկածի, որ նրանք ապրել են այն խեցիների հետ միաժամանակ, որոնք ապրում են այժմ էլ. բայց քանի որ այդ արտասովոր հրեշներն ապրել են մաստոդոնտի և ձիու հետ միաժամանակ, ուստի և կարելի է առնվազն ենթադրել, որ նրանք ապրել են երրորդային ուշ դարաշրջաններից մեկում։
Երբ ասում են, թե կյանքի ծովային ձևերը փոփոխվել են ամբողջ աշխարհում միաժամանակ, չպետք է կարծել, որ այդ արտահայտությունը վերաբերում է մեկ տարվա կամ մեկ դարի, կամ թե ընդհանրապես այն ունի միանգամայն ճշգրիտ երկրաբանական իմաստ։ Եթե Եվրոպայում այժմ ապրող բոլոր ծովային կենդանիներին և բոլոր նրանց, որոնք ապրել են Եվրոպայում պլեյստոցենյան ժամանակաշրջանում (շատ հեռավոր մի ժամանակաշրջան, որը եթե տարիներով չափենք, իր մեջ առնում է ամբողջ սառցադաշտային դարաշրջանը), համեմատենք Հարավային Ամերիկայում կամ Ավստրալիայում այժմ ապրողների հետ, ապա ամենափորձված բնախույզն անգամ հազիվ թե ի վիճակի կլինի ասելու, թե Եվրոպայի այժմյան բնակիչներն ավելի մեծ նմանություն ունեն հարավային կիսագնդում ապրողների հետ, թե պլեյստոցենյան բնակիչները։ Նմանապես և շատ իրազեկ դիտողներից շատերը պնդում են, որ Միացյալ նահանգներում այժմ ապրող ձևերից մի քանիսը ցուցաբերում են ավելի սերտ նմանություն այն ձևերի հետ, որոնք ապրել են Եվրոպայում ուշ երրորդային դարաշրջաններից մեկում, քան Եվրոպայի այժմյան բնակիչների հետ. իսկ եթե այդ այդպես է, ապա ակներև է, որ բրածոներ ունեցող այն շերտերը, որ այժմ նստում են Հյուսիսային Ամերիկայի ափերի մոտ, հետագայում կհամեմատվեն եվրոպական մի քիչ ավելի ուշ ժամանակի շերտերի հետ։ Այնուամենայնիվ, հազիվ թե կարելի է կասկածել այն բանում, որ հեռավոր ապագայում բոլոր նորագույն ծովային ֆորմացիաները, այն է՝ վերին պլիոցենը, պլեյստոցենը և Եվրոպայի, Հյուսիսային ու Հարավային Ամերիկայի և Ավստրալիայի ամենանոր շերտերը, պետք է երկրաբանական իմաստով դիտվեն որպես միաժամանակյա, որովհետև նրանք իրենց մեջ պարունակում են որոշ չափով իրար նմանվող բրածո մնացորդներ և նրանց մեջ արդեն չեն հանդիպում ձևեր, որոնք հատուկ են ավելի հին՝ ներքևում գտնվող նստվածքներին։
Աշխարհի հեռավոր մասերում կյանքի ձևերի միաժամանակյա փոփոխության փաստը վերոհիշյալ լայն իմաստով խիստ զարմացրել է այնպիսի հիանալի դիտողների, ինչպես Վերնեյլն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 217] ու դ’Արշիակն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 218] են։ Նրանք ցույց տալով կյանքի պալեոզոյան ձևերի զուգահեռությունը Եվրոպայի զանազան մասերում, ավելացնում են. «Եթե այդ տարօրինակ հաջորդականությամբ ապշած մենք մեր ուշադրությունը դարձնենք Հյուսիսային Ամերիկայի վրա և այնտեղ գտնենք մի շարք անալոգիական երևույթներ, ապա մեզ անկասկածելի կթվա, որ տեսակների այդ բոլոր փոփոխությունները, նրանց անհետանալը և նորերի երևան գալը չի կարելի բացատրել միայն ծովային հոսանքների փոփոխություններով կամ շատ թե քիչ տեղական ու ժամանակավոր այլ պատճառներով, այլ կախված են ընդհանուր օրենքներից, որոնք ղեկավարում են ամբողջ կենդանական թագավորությունը»։ Բարրանդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 212] հենց նույն իմաստով անում է ծանրակշիռ դիտողություններ։ Եվ իրոք, բոլորովին անօգուտ է դիմել հոսանքների, կլիմայի կամ այլ ֆիզիկական պայմանների փոփոխություններին, նրանց մեջ որոնելով կյանքի ձևերի այդ մեծ փոփոխությունների պատճառները. փոփոխություններ, որոնք կատարվում են երկրի վրա ամենուրեք՝ ամենաբազմազան կլիմաներում։ Մենք պետք է, ինչպես նկատել է Բարրանդը, այստեղ որոնենք ինչ-որ մի առանձին օրենք։ Դա ավելի պարզ կլինի մեզ, երբ մենք ուշադրություն դարձնենք օրգանական ձևերի այժմյան տեղաբաշխման վրա և կտեսնենք, թե որքան թույլ է զանազան երկրների ֆիզիկական պայմանների և նրանց բնակչության բնույթի միջև եղած կախումը։
Ամբողջ աշխարհում կյանքի ձևերի զուգահեռ հաջորդականության այդ մեծ փաստը հեշտ կերպով բացատրվում է բնական ընտրության տեսությամբ։ Նոր տեսակներն առաջանում են շնորհիվ այն բանի, որ ունենում են որոշ առավելություններ այլ ձևերի հանդեպ, իսկ այն ձևերը, որոնք արդեն գերիշխող են կամ ունեն որևէ առավելություն նույն երկրում ապրող այլ ձևերի հանդեպ, սկիզբ են տալիս ամենամեծ թվով տարատեսակների կամ սկզբնավորվող տեսակների։ Մենք դրա պարզ ապացույցն ունենք նրանում, որ այն բույսերը, որոնք գերիշխում են, այսինքն ամենից ավելի սովորական են ու լայնորեն տարածված, առաջացնում են ամենաշատ թվով նոր տարատեսակներ։ Բնական է նաև այն, որ գերիշխող փոփոխական ու լայն կերպով տարածված տեսակները, որոնք արդեն որոշ չափով տիրապետել են այլ տեսակների տերիտորիան, ամենից ավելի մեծ շանսեր կունենան տարածվելու ավելի ևս հեռու և տալու նոր տարատեսակներ ու տեսակներ նոր երկրներում։
Տարածման պրոցեսը հաճախ շատ դանդաղ է կատարվում՝ կախված կլիմայական ու աշխարհագրական փոփոխություններից, զարմանալի պատահականություններից և նոր տեսակների մշտական հարմարումից զանազան պայմանների նկատմամբ, որոնց մեջ նրանք ստիպված են ընկնել, բայց ժամանակի ընթացքում գերիշխող ձևերը սովորաբար հաջողությամբ տարածվում են և վերջ ի վերջո դառնում են տիրապետող։ Առանձին մայր ցամաքների վրա ապրող ցամաքային ձևերը տարածվում են հավանորեն ավելի դանդաղ, քան անընդմեջ տարածվող ծովի բնակիչները։ Ուստի մենք իրավունք ունենք սպասելու, որ կհանդիպենք, և մենք իրոք հանդիպում ենք ցամաքի վրա ապրող ձևերի հաջորդականության մեջ ավելի պակաս խստիվ զուգահեռություն, քան ծովերում ապրող ձևերի համար։
Ուրեմն, ամբողջ աշխարհում կյանքի միևնույն ձևերի զուգահեռ և բառիս լայն իմաստով միաժամանակյա հաջորդականությունն, ըստ երևույթին, լավ համապատասխանում է այն դրույթին, որ նոր տեսակները ծագել են լայն կերպով տարածված ու փոփոխված տիրապետող տեսակներից։ Այս ձևով առաջացած նոր տեսակներն իրենք էլ կլինեն գերիշխող, որովհետև նրանք ձեռք են բերել որոշ առավելություններ իրենց արդեն գերիշխող եղած նախնիների, ինչպես նաև այլ տեսակների հանդեպ, նրանք կտարածվեն ավելի հեռու, կփոփոխվեն ու կառաջացնեն նոր ձևեր։ Նախկին ձևերը, որոնք դուրս են մղվում և իրենց տեղը զիջում են նոր և հաղթող ձևերին, սովորաբար կկազմես ազգակից խմբեր, որովհետև նրանք ժառանգել են մի որևէ ընդհանուր թերություն, և այդ պատճառով էլ որքան նոր և կատարելագործված խմբերը տարածվում են ամբողջ աշխարհում, այնքան հին խմբերն ամենուրեք անհետանում են. և ամենուրեք երևան կգա ձևերի հաջորդականության մեջ համապատասխանություն ինչպես նրանց հայտնվելու, այնպես էլ նրանց վերջնական անհետանալու տեսակետից։
Այդ հարցի վերաբերմամբ ուշադրության արժանի է մի դիտողություն ևս։ Ես նշել եմ պատճառները, որոնք բերում են ինձ այն եզրակացության, որ մեր բրածո պարունակող խոշոր ֆորմացիաների մեծ մասը կուտակվել է իջնելու ժամանակաշրջանների ընթացքում և հսկայական տևողությամբ ընդմիջումները, որ նկատվում են ըստ բրածոների, համընկնում են այն ժամանակաշրջանների հետ, երբ ծովի հատակը կամ մնացել է անշարժ, կամ բարձրացել է, կամ էլ երբ նստվածքները կուտակվել են ոչ բավարար չափով արագ, որպեսզի օրգանական մնացորդները կարողանային նրանց տակ թաղվել ու պահպանվել։ Ես ենթադրում եմ, որ այդ միջակա երկար ժամանակաշրջանների ընթացքում յուրաքանչյուր մարզի բնակիչներ ենթարկվել են զգալի փոփոխությունների ու մահաջնջման և տեղի են ունեցել մեծ չափերով ներգաղթումներ աշխարհի այլ մասերից։ Քանի որ մենք հիմք ունենք կարծելու, որ մեծ մարզեր են ենթարկվել միանման բնույթի շարժման, ուստի չափազանց հավանական է, որ աշխարհի միևնույն մասի շատ ընդարձակ տարածությունների վրա հաճախ նստել են խիստ միաժամանակյա ֆորմացիաներ, սակայն մենք ոչ մի իրավունք չունենք կարծելու, որ այդ միշտ տեղի է ունեցել այդպես և որ մեծ մարզերը միշտ ունեցել են միանման շարժումներ։
Եթե երկու ֆորմացիաներ կուտակվել են երկու մարզերում գրեթե միևնույն, բայց ոչ բոլորովին ճիշտ նույն ժամանակաշրջանում, ապա մենք պետք է վերոհիշյալ պատճառներով նրանց մեջ հանդիպենք կյանքի ձևերի միևնույն ընդհանուր հաջորդականությանը, բայց տեսակների խիստ համապատասխանություն չի լինի, որովհետև փոփոխությունը, մահաջնջումը և նոր ձևերի հայտնվելը, կարող էին տեղի ունենալ մի մարզում ավելի երկար ժամանակի ընթացքում, քան մի այլ մարզում։
Ես կարծում եմ, որ նման դեպքեր հանդիպում են Եվրոպայում։ Միստեր Պրեստվիչին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 219] հաջողվել է Անգլիայի և Ֆրանսիայի էոցենյան նստվածքների մասին իր գրած հիանալի մեմուարներում հաստատել երկու երկրներում հաջորդական հարկերի ընդհանուր սերտ զուգահեռություն. սակայն անգլիական որոշ հարկեր ֆրանսիականների հետ համեմատելիս, թեև երկու երկրներում էլ ստացվում է միևնույն սեռերին պատկանող տեսակների թվի զարմանալի համապատասխանություն, բայց տեսակներն իրենք տարբերվում են միմյանցից, և ի նկատի առնելով երկու մարզերի մոտիկությունը, դժվար է բացատրել այդ տարբերությունները, եթե չդիմենք այն ենթադրության, որ տարբեր բնակիչներ, բայց միաժամանակյա կենդանական աշխարհներ ունեցող երկու ծովերը մի պարանոցով բաժանված են եղել իրարից։ Նման դիտողություններ արել է նաև Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] մի քանի երրորդային ամենաուշ ֆորմացիաների վերաբերմամբ։ Բարրանդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 212] նույնպես նշում է, որ խիստ ընդհանուր զուգահեռություն գոյություն ունի Բոհեմիայի և Սկանդինավիայի սիլուրյան հաջորդական նստվածքների միջև. բայց միաժամանակ նա զարմանալի տարբերություններ է գտնում տեսակների մեջ։ Եթե այդ երկրներում տարբեր ֆորմացիաները միանգամայն նույն ժամանակաշրջաններում էլ չեն կուտակվել, այնպես որ մի երկրում մի ֆորմացիան հաճախ համապատասխանել է մյուս երկրում բաց մնացած տեղին, և եթե երկու երկրներում տեսակները շարունակել են դանդաղ կերպով փոփոխվել զանազան ֆորմացիաների կուտակման ժամանակ և նրանց միջև ընկած երկարատև ժամանակամիջոցների ընթացքում, այդ դեպքում երկու երկրների զանազան ֆորմացիաները կարող էին դասավորվել միանման կարգով՝ կյանքի ձևերի ընդհանուր հաջորդականության համապատասխանաբար, այդ կարգը կարող էր սխալմամբ թվալ խիստ զուգահեռ, և այնուամենայնիվ տեսակները երկու երկրների համապատասխան թվացող հարկերում լիովին միանման չէին լինի։
ՄԱՀԱՋՆՋՎԱԾ ՏԵՍԱԿՆԵՐԻ ՄԻՄՅԱՆՑ ՀԵՏ ԵՎ ԱՅԺՄ ԱՊՐՈՂ ՏԵՍԱԿՆԵՐԻ ՀԵՏ ՈՒՆԵՑԱԾ ԱԶԳԱԿՑՈՒԹՅԱՆ ՄԱՍԻՆ
Այժմ մեր ուշադրությունը սևեռենք մահաջնջված և այժմ ապրող տեսակների փոխհարաբերությունների վրա։ Նրանք բոլորը խմբավորվում են փոքրաթիվ ընդարձակ դասերի մեջ, և այդ փաստը անմիջապես բացատրվում է զարգացման տեսությամբ։ Որքան հին է որևէ ձև, այնքան սովորաբար նա ավելի շատ է տարբերվում այժմ ապրողներից։ Բայց մահաջնջված տեսակները, ինչպես արդեն վաղուց նկատել է Բոկլենդը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 220] բոլորն էլ կարող են տեղավորվել կամ այժմ գոյություն ունեցող խմբերում կամ նրանց միջև եղած արանքներում։ Որ կյանքի մահաջնջված ձևերն օգնում են լցնելու այժմյան սեռերի, ընտանիքների և կարգերի միջև եղած արանքները, դա միանգամայն ճիշտ է. բայց քանի որ այդ դրույթը հաճախ անտեսվում է կամ նույնիսկ բացասվում է, ավելորդ չի լինի մի քանի դիտողություններ անել այդ առթիվ և բերել մի քանի օրինակներ։ Եթե մենք սահմանափակվենք միևնույն դասի կամ ապրող, կամ մահաջնջված տեսակներն առանձին-առանձին քննարկելով, ապա մենք ամենևին այնպես լրիվ շարքեր չենք ունենա, ինչպես նրանք կստացվեն, եթե մեկը և մյուսը դասենք մի ընդհանուր սիստեմի մեջ։ Պրոֆեսոր Օուենի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] աշխատություններում մենք շարունակ հանդիպում ենք մահաջնջվածների վերաբերմամբ գործածվող «ընդհանրացած ձևեր» արտահայտությանը, իսկ Ագասսիսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] աշխատություններում «մարգարեական կամ սինթետիկ տիպեր» արտահայտությունը, և այդ տերմինները ցույց են տալիս, որ այդպիսի ձևերը իրոք հանդիսանում են միջակա կամ շաղկապող օղակներ։ Մի այլ հեղինակավոր հնէաբան՝ Գոդրին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 221] ամենապարզ կերպով ցույց է տվել, որ Ատտիկայում[68] իր հայտնաբերած բրածո կաթնասուններից շատերը լրացնում են գոյություն ունեցող սեռերի միջև եղած տարածությունները։ Կյուվիեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 133] որոճողներին և հաստակաշիներին համարել է կաթնասունների երկու բոլորովին տարբեր կարգեր, բայց մինչև այժմ այնքան բրածո միջակա օղակներ են հայտնաբերվել, որ Օուենը ստիպված է եղել փոխել ամբողջ դասակարգությունը և մի քանի հաստակաշիների զետեղել որոճողների հետ մեկ ենթակարգի մեջ. օրինակ, խոզին ուղտից բաժանող մեծ տարածությունը լցվել է միջակա ձևերով։ Ungulata կամ սմբակավոր չորքոտանիները այժմ ստորաբաժանվում են զուգամատավորների և անզուգամատավորների, բայց հարավամերիկյան Macrauchenia-ն որոշ լափով այդ երկու մեծ ստորաբաժանումները կապում է միմյանց հետ։ Ոչ ոք չի ժխտի, որ հիպպարիոնը հանդիսանում է անցումային ձև այժմյան ձիու և մի քանի ավելի հին սմբակավորների միջև։ Ինչ զարմանալի միացուցիչ օղակ է ներկայացնում կաթնասունների շղթայի մեջ հարավամերիկյան տիպոթերիան, որը և արտահայտում է պրոֆ. Ժերվեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 222] կողմից նրան տրված անունը, մի ձև, որը չի կարելի տեղավորել գոյություն ունեցող կարգերից և ոչ մեկի մեջ։ Կաթնասունների բոլորովին մի առանձին խումբ են կազմում ծովակովերը, և ներկայումս ապրող դյուգոնի և լամանտինի ամենաուշագրավ առանձնահատկություններից մեկն այն է, որ նրանք չունեն հետևի վերջավորություններ, որոնցից նույնիսկ սաղմեր չեն մնացել, բայց բրածո Halitherium-ը, ըստ պրոֆեսոր Ֆլոուերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 161] խոսքերի, ունի ոսկրացած ազդոսկրը, «որը հոդավորվում է կոնքի պարզ կերպով արտահայտված ազդրային հոդափոսի հետ», և, այսպիսով, այս կենդանին փոքր ինչ մոտենում է սմբակավոր չորքոտանիներին, որոնց հետ ծովակովերը նմանություն ունեն ուրիշ տեսակետներից։ Cetacea-ը կամ կետերը խիստ կերպով տարբերվում են մյուս բոլոր կաթնասուններից, բայց երրորդային Zeulodon-ը և Squalodon-ը, որոնց մի քանի բնագետներ զետեղել են մի առանձին ինքնուրույն կարգի մեջ, ներկայացնում են, պրոֆ. Հյոքսլիի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 27] կարծիքով, իսկական Cetacea «և շաղկապող օղակ են հանդիսանում ջրում ապրող գիշատիչների հետ»։
Նույնիսկ թռչունների և սողունների միջև եղած հսկայական տարածությունը, համաձայն նույն պրոֆ. Հյոքսլիի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 27] հետազոտությունների, մասամբ լրացվում է, և ամենաանսպասելի կերպով, մի կողմից ջայլամով ու բրածո արխեոպտերիքսով, իսկ մյուս կողմից բրածո Compsognathus-ով, որը դինոզավրների խմբի ներկայացուցիչներից մեկն է, իսկ այդ խումբն իր մեջ պարփակում է բոլոր ցամաքային սողունների ամենահսկաներին։ Անողնաշարավորների վերաբերմամբ Բարրանդը՝[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 212] ամենաբարձր հեղինակությունը, որին միայն մենք կարող ենք վկայակոչել, հավատացնում է, որ նա ամեն օր ավելի ու ավելի է համոզվում այն կարծիքին, որ թեև պալեոզոյան կենդանիները կարող են, իհարկե, զետեղվել գոյություն ունեցող խմբերում, սակայն այդ հին ժամանակաշրջանում խմբերն իրենք այնպես պարզորոշ կերպով սահմանազատված չէին մեկը մյուսից, ինչպես այժմ նրանք սահմանազատված են։
Մի քանի հեղինակներ ժխտում են այն, որ հնարավոր է որևէ մահաջնջված տեսակ կամ տեսակների խումբ դիտել որպես միջակա ձև այժմ ապրող երկու տեսակների կամ տեսակների խմբերի միջև։ Եթե դրա տակ հասկանանք թե մահաջնջված ձևը իր բոլոր հատկանիշներով ուղղակի միջակա դիրք է գրավում այժմ ապրող երկու ձևերի կամ խմբերի միջև, այդ դեպքում այդպիսի ժխտումը հավանորեն հիմնավոր է։ Բայց բնական դասակարգության մեջ բրածո շատ տեսակներ անկասկածելիորեն կանգնած են այժմյան տեսակների միջև, և մահաջնջված սեռերը կանգնած են այժմ ապրող սեռերի միջև, նույնիսկ այն սեռերի միջև, որոնք պատկանում են զանազան ընտանիքների։ Ամենասովորական դեպքը, մանավանդ շատ հեռավոր խմբերի վերաբերմամբ, ինչպիսիք են, օրինակ, ձկներն ու սողունները, կլինի ըստ երևույթին հետևյալը, եթե երկու խմբերը ներկայումս միմյանցից տարբերվում են երկու տասնյակ հատկանիշներով, ապա նրանց հին ներկայացուցիչները միմյանցից տարբերվել են ավելի փոքրաթիվ հատկանիշներով, այնպես որ երկու խմբերն առաջ մեկը մյուսին մի քիչ ավելի մոտ են կանգնած եղել, քան այժմ։
Սովորաբար կարծում են, թե որքան հին է որևէ մի ձև, այնքան նրա մեջ ավելի շատ կարելի է գտնել հատկանիշներ, որոնք միմյանց հետ շաղկապում են մեկը մյուսից հեռու կանգնած խմբերը։ Այդ դիտողությունը, իհարկե, պետք է վերաբերի միայն այն խմբերին, որոնք խիստ փոփոխության են ենթարկվել, երկրաբանական ժամանակների ընթացքում. դժվար կլիներ ապացուցել այդ կարծիքի արդարացիությունը, որովհետև այժմ էլ ժամանակ առ ժամանակ գտնում են մինչև հիմա գոյություն ունեցող կենդանի, ինչպես, օրինակ, Lepidosiren-ը, որը ցուցաբերում է ազգակցության հատկանիշներ շատ հեռու կանգնած խմբերի հետ։ Սակայն, եթե մենք համեմատենք հին սողուններին ու երկկենցաղներին, հին ձկներին, հին գլխոտանիներին և էոցենի կաթնասուններին նույն դասերի ավելի նոր ներկայացուցիչների հետ, ապա մենք պետք է ընդունենք, որ ճշմարտության բաժին կա այդ դիտողության մեջ։
Այժմ տեսնենք, թե այդ բազմազան փաստերն ու եզրակացություններն ինչպես են համաձայնվում փոփոխությունների միջոցով տեղի ունեցած զարգացման տեսության հետ։ Քանի որ այդ հարցը մի քիչ բարդ է, ուստի կխնդրեմ ընթերցողին դիմել չորրորդ գլխում զետեղված դիագրամին (էջ 205)։ Ենթադրենք, թե շեղագիր տպված և թվանշաններ ունեցող տառերը ներկայացնում են սեռեր, իսկ նրանցից ճյուղավորվող կետագծերը՝ յուրաքանչյուր սեռի տեսակներ։ Հորիզոնական գծերը թող ցույց տան հաջորդական երկրաբանական ֆորմացիաները, և դիցուք, բոլոր այն ձևերը, որոնք զետեղված են ամենավերին գծից ներքև, պատկերեն մահաջնջված ձևեր։ Այժմ գոյություն ունեցող երեք՝ 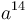 ,
, 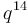 ,
,  սեռերը կազմում են մի փոքրիկ ընտանիք,
սեռերը կազմում են մի փոքրիկ ընտանիք,  և
և  ` մոտ ազգակից մի ընտանիք կամ ենթաընտանիք, իսկ
` մոտ ազգակից մի ընտանիք կամ ենթաընտանիք, իսկ  ,
, 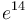 ,
, 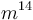 ` երրորդ ընտանիքը։ Այս երեք ընտանիքները, մահաջնջված շատ սեռերի հետ միասին, որոնք կանգնած են նախահայր (A) ձևից ճյուղավորվող տոհմաբանական զանազան գծերի վրա, կազմում են մի կարգ, որովհետև նրանք բոլորն էլ իրենց հին նախածնողից պիտի ժառանգած լինեն ինչ-որ ընդհանուր բան։ Հատկանիշների տարամիտվելու մշտական ձգտման սկզբունքով, որի լուսաբանության համար էլ բերված է այս դիագրամը, որքան նոր է մի որևէ ձև, այնքան նա ավելի կտարբերվի իր հին նախածնողից։ Սրանից հասկանալի է այն կանոնը, որ ամենահին ձևերը ամենից շատ են տարբերվում այժմ ապրողներից։ Սակայն մենք չպետք է կարծենք, որ հատկանիշների տարամիտումը մի ինչ-որ անխուսափելի բան է. նա կախված է միայն նրանից, որ որևէ տեսակի հաջորդներին հաջողվում է բնության տնտեսության մեջ գրավել շատ ու բազմազան վայրեր։ Ուրեմն, միանգամայն հնարավոր է, ինչպես մենք այդ տեսանք սիլուրյան մի քանի ձևերի օրինակի վրա, որ տեսակը կարող է իր գոյությունը շարունակել, թույլ կերպով փոփոխել իր կենսապայմանների թույլ փոփոխության պատճառով և, այնուամենայնիվ, հսկայական ժամանակաշրջանի ընթացքում պահպանել միևնույն ընդհանուր հատկանիշները։ Դիագրամի վրա այդ պատկերված է
` երրորդ ընտանիքը։ Այս երեք ընտանիքները, մահաջնջված շատ սեռերի հետ միասին, որոնք կանգնած են նախահայր (A) ձևից ճյուղավորվող տոհմաբանական զանազան գծերի վրա, կազմում են մի կարգ, որովհետև նրանք բոլորն էլ իրենց հին նախածնողից պիտի ժառանգած լինեն ինչ-որ ընդհանուր բան։ Հատկանիշների տարամիտվելու մշտական ձգտման սկզբունքով, որի լուսաբանության համար էլ բերված է այս դիագրամը, որքան նոր է մի որևէ ձև, այնքան նա ավելի կտարբերվի իր հին նախածնողից։ Սրանից հասկանալի է այն կանոնը, որ ամենահին ձևերը ամենից շատ են տարբերվում այժմ ապրողներից։ Սակայն մենք չպետք է կարծենք, որ հատկանիշների տարամիտումը մի ինչ-որ անխուսափելի բան է. նա կախված է միայն նրանից, որ որևէ տեսակի հաջորդներին հաջողվում է բնության տնտեսության մեջ գրավել շատ ու բազմազան վայրեր։ Ուրեմն, միանգամայն հնարավոր է, ինչպես մենք այդ տեսանք սիլուրյան մի քանի ձևերի օրինակի վրա, որ տեսակը կարող է իր գոյությունը շարունակել, թույլ կերպով փոփոխել իր կենսապայմանների թույլ փոփոխության պատճառով և, այնուամենայնիվ, հսկայական ժամանակաշրջանի ընթացքում պահպանել միևնույն ընդհանուր հատկանիշները։ Դիագրամի վրա այդ պատկերված է 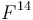 տառով։
տառով։
A-ից առաջացած բոլոր մահաջնջված և այժմյան ձևերը կազմում են, ինչպես արդեն նշվել է, մի կարգ, իսկ այդ կարգը մահաջնջման ու հատկանիշների տարամիտման շարունակ գործող ազդեցությունների հետևանքում բաժանվել է զանազան ենթաընտանիքների և ընտանիքների, որոնցից մի քանիսը, հավանորեն, պետք է ոչնչանային զանազան ժամանակաշրջաններում, իսկ մյուսները շարունակում են ապրել մինչև ներկա ժամանակները։
Զննելով այդ դիագրամը, կարելի է տեսնել, որ եթե մահաջնջված ձևերից շատերը, որոնք պետք է գոյություն ունենային հաջորդական ֆորմացիաներում, հայտնաբերվեին սերիայի ստորին մասի զանազան տեղերում, ապա ամենավերին գծի վրայի երեք այժմյան րնտանիքները միմյանցից ավելի քիչ կզանազանվեին։ Օրինակ, եթե հայտնաբերվեին 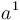 ,
, 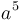 ,
, 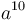 ,
, 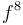 ,
, 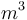 ,
,  ,
,  սեռերը, ապա այդ ընտանիքներն այնպես սերտորեն միացած կլինեին մի մեծ ընտանիքի մեջ, ինչպես որ այդ տեղի է ունեցել որոճողների ու մի քանի հաստակաշիների հետ։ Սակայն նա, ով չի համաձայնի երեք ընտանիքների այժմ ապրող սեռերն այս ձևով իրար հետ շաղկապող մահաջնջված սեռերի վրա նայել որպես միջակա սեռերի, կարող է մասամբ իրավացի լինել, որովհետև նրանք միջակա լինում են ոչ թե անմիջականորեն, այլ երկար ու զարտուղի ճանապարհով՝ անցնելով բազմաթիվ խիստ տարբեր ձևերով։ Եթե մահաջնջված ձևերից շատերը հայտնաբերվեին երկրաբանական ֆորմացիաները ցույց տվող միջին հորիզոնական գծերից մեկի վերևում, օրինակ VI գծից բարձր և ոչ մեկը չհայտնաբերվեր այդ գծից ցած, այդ դեպքում ընտանիքներից միայն երկուսը (այսինքն ձախ կողմում գտնվող և այլն և և այլն) կմիանային իրար հետ և այն ժամանակ կմնային երկու ընտանիքներ, որոնք ավելի քիչ կտարբերվեին միմյանցից, քան տարբերվում էին մինչև բրածոների հայտնաբերումը։ Այնուհետև, եթե ենթադրենք, որ այն երեք ընտանիքները, որոնք կանգնած են վերին գծի վրա և իրենց մեջ պարփակում են ութ սեռեր (սկսած -ից մինչև ), տարբերվում են մեկը մյուսից կես դյուժին կարևոր հատկանիշներով, այդ դեպքում այն ընտանիքները, որոնք գոյություն են ունեցել VI թվանշանով նշանակված ժամանակաշրջանում, հավանորեն կտարբերվեին մեկը մյուսից ավելի քիչ թվով հատկանիշներով, որովհետև իրենց զարգացման այդ վաղ շրջանում նրանք պետք է ավելի քիչ չափով տարբերվեին իրենց ընդհանուր նախածնողից։ Այսպիսով, հին և մահաջնջված սեռերը հաճախ իրենց հատկանիշներով շինում են ավելի կամ պակաս չափով միջականեր իրենց փոփոխված հաջորդների միջև կամ իրենց կողմնակի ազգականների միջև։
սեռերը, ապա այդ ընտանիքներն այնպես սերտորեն միացած կլինեին մի մեծ ընտանիքի մեջ, ինչպես որ այդ տեղի է ունեցել որոճողների ու մի քանի հաստակաշիների հետ։ Սակայն նա, ով չի համաձայնի երեք ընտանիքների այժմ ապրող սեռերն այս ձևով իրար հետ շաղկապող մահաջնջված սեռերի վրա նայել որպես միջակա սեռերի, կարող է մասամբ իրավացի լինել, որովհետև նրանք միջակա լինում են ոչ թե անմիջականորեն, այլ երկար ու զարտուղի ճանապարհով՝ անցնելով բազմաթիվ խիստ տարբեր ձևերով։ Եթե մահաջնջված ձևերից շատերը հայտնաբերվեին երկրաբանական ֆորմացիաները ցույց տվող միջին հորիզոնական գծերից մեկի վերևում, օրինակ VI գծից բարձր և ոչ մեկը չհայտնաբերվեր այդ գծից ցած, այդ դեպքում ընտանիքներից միայն երկուսը (այսինքն ձախ կողմում գտնվող և այլն և և այլն) կմիանային իրար հետ և այն ժամանակ կմնային երկու ընտանիքներ, որոնք ավելի քիչ կտարբերվեին միմյանցից, քան տարբերվում էին մինչև բրածոների հայտնաբերումը։ Այնուհետև, եթե ենթադրենք, որ այն երեք ընտանիքները, որոնք կանգնած են վերին գծի վրա և իրենց մեջ պարփակում են ութ սեռեր (սկսած -ից մինչև ), տարբերվում են մեկը մյուսից կես դյուժին կարևոր հատկանիշներով, այդ դեպքում այն ընտանիքները, որոնք գոյություն են ունեցել VI թվանշանով նշանակված ժամանակաշրջանում, հավանորեն կտարբերվեին մեկը մյուսից ավելի քիչ թվով հատկանիշներով, որովհետև իրենց զարգացման այդ վաղ շրջանում նրանք պետք է ավելի քիչ չափով տարբերվեին իրենց ընդհանուր նախածնողից։ Այսպիսով, հին և մահաջնջված սեռերը հաճախ իրենց հատկանիշներով շինում են ավելի կամ պակաս չափով միջականեր իրենց փոփոխված հաջորդների միջև կամ իրենց կողմնակի ազգականների միջև։
Բնության մեջ այդ հարաբերությունները պետք է անհամեմատ ավելի բարդ լինեն, քան այդ պատկերված է դիագրամով, որովհետև խմբերը եղել են ավելի բազմաթիվ, նրանք պետք է գոյություն ունենային ամենևին ոչ միաժամանակ և փոփոխվեին շատ տարբեր չափով։ Քանի որ մենք ունենք երկրաբանական տարեգրության միայն վերջին հատորը, և այն էլ խիստ կցկտուր ձևով, ուստի մենք ոչ մի իրավունք չունենք ակնկալելու, բացառությամբ միայն մի քանի հազվագյուտ դեպքերի, որ հնարավորություն կունենանք լրացնելու մեծ բացերը բնական սիստեմի մեջ, և այդպիսով միացնելու առանձին ընտանիքները կարգերի մեջ։ Ամենաշատը, որ մենք իրավունք ունենք սպասելու, այդ այն է, որպեսզի որոշ երկրաբանական ժամանակաշրջանում մեծ փոփոխությունների ենթարկված խմբերը ավելի հին ֆորմացիաներում մեկը մյուսից փոքր ինչ ավելի մոտ կանգնած լինեն, այնպես որ խմբերի հին ներկայացուցիչները իրենց որոշ հատկանիշներով ավելի քիչ տարբերվեին միմյանցից, քան տարբերվում են այդ խմբերի այժմյան ներկայացուցիչները միմյանցից, իսկ այդ, համաձայն մեր լավագույն հնէաբանների միահամուռ վկայության, պատահում է շատ հաճախ։
Ուրեմն, փոփոխությունների միջոցով կատարվող զարգացման տեսությունը բավարար կերպով է բացատրում այն գլխավոր փաստերը, որոնք վերաբերում են կյանքի մահաջնջված տեսակների միմյանց հետ և այժմ ապրող ձևերի հետ ունեցած փոփոխություններին։ Որևէ այլ հայեցակետով նրանք բոլորովին անբացատրելի են։
Այդ տեսության տեսակետից ակնհայտ է, որ երկրի պատմության որևէ մեծ ժամանակաշրջանի կենդանական աշխարհն իր ընդհանուր բնույթով պետք է միջակա տեղ գրավի նախորդ ու հաջորդ կենդանական աշխարհների միջև. օրինակ, այն տեսակները, որոնք ապրել են զարգացման վեցերորդ մեծ դարաշրջանում, որը նշված է դիագրամի վրա, հանդիսանում են հինգերորդ դարաշրջանում ապրածների փոփոխված հաջորդները և յոթերորդ դարաշրջանում ավելի ևս փոփոխված ձևերի նախահայրերը, հետևաբար, նրանք հազիվ թե իրենց հատկանիշներով չեն կարող համարյա միջակա ձևեր լինել կյանքի ավելի վաղ և ուշ ձևերի միջև։ Սակայն մենք պետք է ուշադրության առնենք, որ նախորդ որոշ ձևերն իսպառ մահաջնջվել են, տվյալ մարզում երևացել են այլ մարզերից եկած նոր ձևեր և մեծ չափով փոփոխություններ են կատարվել հաջորդական ֆորմացիաների միջև ընկած երկարատև ընդմիջումների ժամանակ։ Այդ սահմանափակումներով յուրաքանչյուր երկրաբանական ժամանակաշրջանի կենդանական աշխարհն անկասկած պետք է իր հատկանիշներով միջակա լինի նախորդ և հաջորդ կենդանական աշխարհների միջև։ Ես կբերեմ միայն մեկ օրինակ, այսինքն այն, որ երբ դեվոնյան սիստեմը առաջին անգամ հայտնաբերվել է, հնէաբանները նրա մեջ գտնվող բրածոներին, նրանց հատկանիշների համաձայն, համարել են միջակա այն բրածոների միջև, որոնք պարունակվել են վերևում գտնվող քարածխային և ցածում գտնվող սիլուրյան սիստեմների մեջ։ Բայց յուրաքանչյուր կենդանական աշխարհ չպետք է անպայման լինի միջակա, բառիս նեղ իմաստով, որովհետև հաջորդական ֆորմացիաների միջև անցած ժամանակամիջոցները կարող էին նաև անհավասար լինել։
Այն հանգամանքը, որ որոշ տեսակներ այս կանոնից բացառություն են ներկայացնում, չի հակասում այն դրույթի ճշմարտությանը, որ յուրաքանչյուր ժամանակաշրջանի կենդանական աշխարհն ամբողջությամբ վերցրած իր հատկանիշներով միջակա է լինում նախորդ և հաջորդ կենդանական աշխարհների միջև։ Օրինակ, երբ դոկտոր Ֆոկոները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 73] մաստոդոնտների և փղերի տեսակները դասավորել է երկու շարքում՝ մեկն ըստ նրանց փոխադարձ ազգակցության և մյուսն ըստ նրանց գոյության ժամանակաշրջանների, ապա ստացվել է այն, որ երկու շարքը չեն համապատասխանում իրար ձևերի դասավորությամբ, այսինքն՝ ծայրային հատկանիշներ ունեցող տեսակները լինում են ոչ ամենահները և ոչ էլ ամենանորերը, իսկ միջակա հատկանիշներ ունեցող տեսակները տարիքի համեմատ միջակա տեղ չեն գրավում։ Բայց, մի րոպե ենթադրելով, որ այդ և ուրիշ նման դեպքերում մենք ստույգ տվյալներ ունենք տեսակների առաջին անգամ երևան գալու և անհետանալու ժամանակի մասին, որը իրականում բնավ չկա, մենք հիմք չունենք կարծելու, որ հաջորդականորեն առաջացած ձևերի կյանքը պետք է տևի համապատասխան ժամանակամիջոցների ընթացքում։ Որոշ դեպքերում ավելի հին ձևը կարող էր գոյություն ունենալ ավելի երկար, քան որևէ տեղում ավելի ուշ առաջացած մի ձև, և դա առանձնապես բնական է ցամաքային ձևերի վերաբերմամբ, որոնք բնակվել են բաժան-բաժան մարզերում։ Խոշոր ձևերից հետո, օրինակի համար վերցնենք փոքրերին։ Եթե մենք ընտանի աղավնու այժմ ապրող և մահաջնջված գլխավոր տեսակները շարքերով դասավորենք ըստ նրանց ազգակցության, ապա այդպիսի դասավորությունը ոչ լիովին կհամապատասխանի այն կարգին, ինչ կարգով ծագել են այդ ցեղերը, և ավելի ևս քիչ նրանց անհետացման կարգին, որովհետև նախահայր ժայռաղավնին ապրում է մինչև հիմա, իսկ մի քանի տարատեսակներ, որ միջակա են նրա և փոստատար աղավնու միջև, մահաջնջվել են. փոստատար աղավնիները, որոնք էական հատկանիշով՝ կտուցի երկարությամբ ծայրային տեղ են գրավում, ավելի վաղ են առաջացել, քան կարճակտուց թուրմանները, որոնք այդ տեսակետից տվյալ շարքում հակադիր տեղ են գրավում։
Եզրակացությունն այն մասին, որ որևէ միջակա ֆորմացիայի օրգանական մնացորդներն ունեն որոշ աստիճանի միջակա հատկանիշներ, սերտ առնչության մեջ է գտնվում բոլոր հնէաբանների կողմից պաշտպանվող այն փաստերի հետ, որ երկու հաջորդական ֆորմացիաների բրածոներն ավելի սերտ կերպով են կապված միմյանց հետ, քան երկու հեռավոր ֆորմացիաների բրածոները։ Պիկտեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 207] բերում է օրգանական մնացորդների ընդհանուր նմանության մի լավ օրինակ կավճային ֆորմացիայի զանազան հարկերում, թեև յուրաքանչյուր հարկում տեսակները տարբեր են։ Մենակ այդ փաստն իր ընդհանրությամբ խախտեց պրոֆ. Պիկտեի հավատը դեպի տեսակների անփոփոխելիությունը։ Ով ծանոթ է երկրի մակերեսին այժմ ապրող տեսակների բաշխմանը, նա չի աշխատի մեկը մյուսին հաջորդող երկու մերձավոր ֆորմացիաներում զանազան տեսակների այդ մոտ նմանության բացատրությունները որոնել այն բանում, որ հին մարզերի ֆիզիկական պայմանները գրեթե միանման են մնացել։ Պետք է վերհիշել, որ օրգանիզմները, գոնե նրանք, որոնք ապրել են ծովերում, փոփոխվել են ամբողջ աշխարհում գրեթե միաժամանակ և, հետևապես, միանգամայն տարբեր կլիմաներում և պայմաններում։ Վերհիշենք այն զարմանալի կլիմայական փոփոխությունները, որոնք տեղի են ունեցել պլեյստոցենյան ժամանակաշրջանում, որն իր մեջ պարփակում է նաև ամբողջ սառցադաշտային դարաշրջանը, և նկատենք, թե նրանք ինչքան քիչ են անդրադարձել ծովի բնակիչների տեսակային ձևերի վրա։
Զարգացման տեսության տեսակետից միանգամայն հասկանալի է, որ մեկը մյուսին մոտիկ հաջորդող ֆորմացիաների օրգանական մնացորդները սերտ կերպով կապված են միմյանց հետ, թեև նրանք պատկանում են զանազան տեսակների։ Քանի որ յուրաքանչյուր ֆորմացիայի կուտակումը հաճախակի ընդհատվել է և քանի որ հաջորդական ֆորմացիաների միջև գոյություն ունեն խոշոր բաց տեղեր, ուստի, ինչպես ես փորձել եմ ցույց տալ վերջին գլխում, մենք չենք կարող սպասել, որ որևէ մեկ կամ երկու ֆորմացիաներում կգտնենք բոլոր միջակա տարատեսակները այն տեսակների միջև, որոնք երևան են եկել այդ ժամանակաշրջանների սկզբում և վերջում. բայց այդպիսի ընդմիջումներից հետո, որոնք շատ երկար են, եթե չափում ենք տարիներով, և այնքան էլ երկարատև չեն, եթե նրանց չափում ենք երկրաբանորեն, մենք կարող ենք գտնել իրար հետ սերտ կերպով կապված ձևեր կամ, ինչպես մի քանի հեղինակներ անվանում են դրանց, ներկայացուցիչ-տեսակներ կամ փոխարինող տեսակներ և նրանց մենք իսկապես գտնում ենք։ Մի խոսքով, մենք գտնում ենք տեսակային ձևերի դանդաղ և հազիվ նշմարելի փոփոխությունների հենց այնպիսի ապացույցներ, ինչպիսիք մենք իրավունք ունենք սպասելու։
ՆԵՐԿԱՅՈՒՄՍ ԱՊՐՈՂ ՁԵՎԵՐԻ ՀԱՄԵՄԱՏՈՒԹՅԱՄԲ ՀԻՆ ՁԵՎԵՐԻ ԶԱՐԳԱՑՄԱՆ ԱՍՏԻՃԱՆԻ ՄԱՍԻՆ
Չորրորդ գլխում մենք արդեն տեսանք, որ հասունության հասած օրգանիզմների մասերի դիֆերենցման ու մասնագիտացման աստիճանը նրանց կազմվածքի հարաբերական կատարելության բարձրության ամենալավ ցուցանիշն է։ Նմանապես և տեսանք, որ մասերի մասնագիտացումը քանի որ առավելություն է տալիս օրգանիզմին, ապա բնական ընտրությունը ձգտում է, որպեսզի յուրաքանչյուր էակի կազմվածքը դարձնի ավելի մասնագիտացված և կատարելագործված և այդ իմաստով ավելի բարձր. դրա հետ միասին բնական ընտրությունը կարող է պահպանել պարզ ու անկատարյալ կառուցվածք ունեցող շատ ձևեր, որոնք հարմարված են պարզ կենսապայմանների, և որոշ դեպքերում նույնիսկ կարող է իջեցնել ու պարզեցնել կազմվածքը, որը այդպիսի պարզացած ձևերին դարձնում է ավելի լավ հարմարված իրենց նոր պայմաններին։ Այլև ավելի սովորական դեպքերում նոր տեսակները դառնում են կազմվածքով ավելի բարձր, քան իրենց նախորդները, որովհետև գոյության կռվում նրանք հաղթում են նախկին ձևերին, որոնց հետ նրանք մրցության մեջ են մտնում։ Սրանից մենք կարող ենք եզրակացնել, որ եթե գրեթե միանման կլիմայական պայմաններում երկրագնդի էոցենյան բնակիչները մրցության մեջ մտնեին այժմյան բնակիչների հետ, վերջիններս կհաղթահարեին ու կոչնչացնեին առաջիններին, ինչպես որ էոցենյանները երկրորդայիններին և երկրորդայինները պալեոզոյաններին։ Այնպես որ թե գոյության կռվում հաղթանակի այդ հիմնական չափանիշի շնորհիվ և թե ի նկատի առնելով օրգանների մասնագիտացման աստիճանը, նոր ձևերը, բնական ընտրության տեսության համաձայն, պետք է ավելի բարձր դասվեն, քան հները։ Արդյոք, իրո՞ք այդ այդպես է։ Հնէաբանների հսկայական մեծամասնությունը դրան դրական պատասխան կտար, և թվում է, թե այդ պատասխանը հարկավոր է ճշմարտություն համարել, թեպետև դժվար է այն ապացուցել։
Այդ եզրակացության դեմ չի կարելի լուրջ առարկություն համարել այն հանգամանքը, որ մի քանի բազկոտանիներ ամենահեռավոր երկրաբանական դարաշրջաններից սկսած, մնացել են քիչ փոփոխված և որոշ ցամաքային ու քաղցրահամ ջրերի կակղամորթներ մնացել են համարյա նույնը այն ժամանակվանից սկսած, ինչքան մենք գիտենք, երբ նրանք առաջին անգամ երևացել են։ Անհաղթահարելի դժվարություն չի ներկայացնում նաև այն, որ արմատոտանիները, դոկտոր Կարպենտերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 223] հավաստիացմամբ, իրենց կազմվածքով չեն առաջադիմել սկսած նույնիսկ լավրենտյան դարաշրջանից, որովհետև որոշ օրգանիզմներ պետք է պարզ կենսապայմաններին հարմարված մնային, իսկ ի՞նչը կարող էր ավելի լավ հարմարվել այդ պայմաններին, քան այդ ցածր կազմվածքով Protozoa-ն։ Վերոհիշյալներին նման առարկությունները իմ տեսության համար ճակատագրական կլինեին, եթե նա կազմվածքի առաջադիմությունը որպես անհրաժեշտ պայման ընդուներ, նմանապես նրանք ճակատագրական կլինեին, եթե կարելի լիներ ապացուցել, որ, օրինակ, վերոհիշյալ արմատոտանիներն առաջին անգամ երևացել են լավրենտյան դարաշրջանում կամ վերոհիշյալ բազկոտանիները կեմբրիյան դարաշրջանում, որովհետև այդ դեպքում բավականաչափ ժամանակ չէր լինի, որպեսզի այդ օրգանիզմները հասնեին զարգացման այն աստիճանին, որին նրանք հասել են այդ դարաշրջաններում. բայց քանի որ նրանք հասել են զարգացման տվյալ աստիճանին, ապա բնական ընտրության տեսակետից բոլորովին անհրաժեշտություն չկա, որ նրանք շարունակեին է՛լ ավելի կատարելագործվել, թեև նրանք պետք է որ յուրաքանչյուր հաջորդող դարաշրջանում թեթևակի փոփոխվեին, որպեսզի պահեին իրենց տեղերը այն թույլ փոփոխությունների ժամանակ, ինչպիսիք տեղի են ունենում նրանց գոյության պայմաններում։ Վերոհիշյալ առարկությունները սերտ կերպով կապված են այն հարցի հետ, թե մենք արդյոք գիտե՞նք, թե իսկապես որքան հին է մեր աշխարհը և կյանքի զանազան ձևերը առաջին անգամ ո՞ր ժամանակաշրջաններում են երևացել. այդ մասին կարելի է դեռ շատ վիճել։
Այն հարցը, թե արդյոք կազմվածքն ամբողջությամբ վերցրած առաջադիմել է, շատ տեսակետներից չափազանց խճողված է։ Երկրաբանական տարեգրությունը, որ բոլոր ժամանակներում թերի է, չի տարածվում մինչև բավականաչափ հեռավոր ժամանակները, որպեսզի աներկբայելի հստակությամբ դրսևորեր, որ աշխարհի մեզ ծանոթ պատմության ընթացքում կազմվածքը մեծ չափով առաջ է շարժվել։ Նույնիսկ ներկայումս, քննարկելով միևնույն դասի ներկայացուցիչներին, բնախույզները չեն համաձայնվում միմյանց հետ, թե որ ձևերը պետք է համարել բարձրակարգ։ Այսպես, ոմանք սելախիաներին կամ շնաձկներին համարում են ձկների բարձրակարգ ներկայացուցիչներ այն հիման վրա, որ նրանք իրենց կազմվածքի մի քանի կարևոր գծերով մոտենում են սողուններին, մյուսները բարձրակարգ են համարում ոսկրավոր ձկներին։ Գանոիդ ձկներին դասում են սելախիաների և ոսկրավորների միջև. վերջիններս այժմ ներկայացնում են մի խումբ,, որը ձևերի թվով զգալի չափով գերակշռում է, բայց առաջներում գոյություն են ունեցել միայն սելախիաները և գանոիդները, և տվյալ դեպքում մենք կարող ենք որոշել, թե ձկներն իրենց կազմվածքով արդյոք առաջադիմել են կամ ցածրացել միայն համապատասխան այն բանի, թե կազմվածքի բարձրության ինչպիսի չափանիշ կցանկանանք ընտրել։ Կազմվածքի բարձրության տեսակետից զանազան տիպերի ներկայացուցիչներին համեմատելու փորձերը միանգամայն անհույս են։ Ո՞վ կորոշի, թե որն է ավելի բարձր, թանաքաձո՞ւկը, թե մեղուն, այդ միջատը, որին մեծ Բերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 25] համարել է «ավելի բարձր կազմակերպված, քան ձուկը, թեև այլ պլանով»։ Կյանքի համար տեղի ունեցող բարդ մրցության մեջ միանգամայն հնարավոր է, որ խեցեմորթները, որոնք իրենց դասի ներկայացուցիչների մեջ այնքան էլ բարձր կազմվածք չունեն, հաղթում են գլխոտանիներին՝ կակղամորթների բարձրակարգ ներկայացուցիչներին և այդպիսի խեցեմորթները, որոնք թեև բարձր զարգացում չունեն, պետք է դասվեն շատ բարձր անողնաշարավոր կենդանիների շարքում, եթե նրանց գնահատելիս ղեկավարվենք բոլոր ստուգումներից ամենավճռականով՝ գոյության կռվի օրենքով։ Բացի դրանից, որ մենք հանդիպում ենք այդպիսի դժվարությունների այն հարցը որոշելիս, թե որ ձևերն են ամենից ավելի առաջ գնացել իրենց կազմվածքով. մենք չպետք է համեմատենք որևէ դասի միմիայն բարձրակարգ ներկայացուցիչներին, որոնք գոյություն են ունեցել թե մեկ և թե մյուս դարաշրջաններում, թեև կասկած չկա, որ դա կարևոր, գուցե և ամենակարևոր տարրն է հետևություններ անելու ժամանակ, բայց մենք պետք է համեմատենք երկու ժամանակաշրջաններում եղած ստորակարգ և բարձրակարգ բոլոր ներկայացուցիչներին միմյանց հետ։ Մի որոշ հին դարաշրջանում թվով գերազանցել են բարձրակարգ և ստորակարգ կակղամորթները, այսինքն, գլխոտանիները և բազկոտանիները. ներկայումս այդ երկու խմբերն էլ խիստ կրճատվել են, մինչդեռ մյուսները, կազմվածքով միջակները, զգալիորեն բազմացել են. դրա հետևանքով մի քանի բնախույզներ պաշտպանում են այն կարծիքը, որ կակղամորթները առաջներում հասել են ավելի բարձր զարգացման, քան այժմ. բայց հակառակ կարծիքի օգտին կարելի է բերել ավելի ծանրակշիռ փաստարկ, այն է՝ ցույց տալ բազկոտանիների խիստ կրճատումը և այն փաստը, որ մեր այժմյան գլխոտանիները թեև սակավաթիվ են, բայց ավելի բարձր կազմվածք ունեն, քան այդ դասի հին ներկայացուցիչները։ Մենք պետք է համեմատենք նաև երկու ժամանակաշրջանների ընթացքում եղած բարձրակարգ և ստորակարգ դասերի հարաբերական քանակությունը երկրի ամբողջ մակերևույթի վրա։ Եթե, օրինակ, գոյություն ունեն ողնաշարավոր կենդանիների հիսուն հազար զանազան ձևեր և եթե մենք գիտենայինք, որ մի ինչ-որ նախկին ժամանակաշրջանում դրանցից գոյություն են ունեցել միայն տասը հազար ձևեր, մենք կկարողանայինք բարձրակարգ դասում ձևերի թվի այդ ավելացումը, որը ենթադրում է ստորակարգ ձևերի մեծ կրճատում, դիտել որպես կազմվածքի վճռական առաջադիմություն աշխարհի մեջ։ Այստեղից մենք տեսնում ենք, թե որքան անհուսալիորեն դժվար է, այդպիսի չափազանց բարդ պայմաններում, լիակատար ճշտությամբ համեմատել մեկը մյուսին հաջորդած երկրաբանական ժամանակաշրջանների այնքան անկատար կերպով հայտնի կենդանական աշխարհների կազմվածքի հարաբերական բարձրությունը։
Մենք էլ ավելի պարզ կերպով կգնահատենք այդ դժվարությունը, ուսումնասիրելով գոյություն ունեցող մի քանի կենդանիներ ու բույսեր։ Այն անսովոր արագությունը, որով եվրոպական օրգանիզմները վերջերս տարածվել են Նոր Զելանդիայում և գրավել տեղական ձևերի առաջվա բռնած տեղերը, մեզ իրավունք է տալիս կարծելու, որ եթե Մեծ Բրիտանիայի բոլոր կենդանիներն ու բույսերը մտցվեին Նոր Զելանդիա, բրիտանական շատ ու շատ ձևեր այնտեղ ժամանակի ընթացքում լրիվ կենթարկվեին նատուրալիզացիայի և դուրս կմղեին տեղական ձևերին։ Մյուս կողմից հիմնվելով այն փաստի վրա, որ հազիվ թե հարավային կիսագնդի գոնե մի բնակիչ վայրենանա Եվրոպայում որևէ մի տեղում, մենք լիակատար հիմք ունենք կասկածելու, որ նոր զելանդական բոլոր ձևերը Մեծ Բրիտանիա տեղափոխելու դեպքում կարող էին մի քիչ զգալի թվով գրավել այն տեղերը, որոնք այժմ գրավված են մեր հայրենի բույսերի և կենդանիների կողմից։ Այդ տեսակետից Մեծ Բրիտանիայում բնակվող օրգանիզմները ձևերի շարքում ավելի բարձր տեղ են գրավում նոր զելանդականների համեմատությամբ։ Չնայած դրան, ամենաիրազեկ բնախույզն էլ, մեկ և մյուս երկրների տեսակներն ուսումնասիրելու հիման վրա, չէր կարողանա կանխատեսել այդպիսի հետևանքը։
Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] և շատ այլ գիտակ դատավորներ պնդում են, որ հին կենդանիները որոշ չափով նման են նույն դասի նոր կենդանիների սաղմերին և մահաջնջված ձևերի երկրաբանական հաջորդականությունը գրեթե զուգահեռ է՝ այժմյան ձևերի սաղմնաբանական զարգացման հետ։ Այդպիսի հայացքը զարմանալիորեն լավ համաձայնվում է մեր տեսության հետ։ Հաջորդ գլուխներից մեկում ես կաշխատեմ ցույց տալ, որ չափահաս ձևը տարբերվում է իր սաղմից այն փոփոխությունների հետևանքով, որոնք սկսվում են ոչ ամենավաղ հասակում և երևան են գալիս զարգացման համապատասխան ժամանակաշրջաններում որպես ծնողներից ժառանգվածներ։ Այդ պրոցեսը, թեև նա սաղմը թողնում է համարյա առանց փոփոխությունների, հաջորդական սերունդներում շարունակ կուտակում է չափահաս ձևի ավելի ու ավելի մեծ փոփոխություններ։ Այսպիսով, սաղմը կարծես ներկայացնում է տեսակի նախկին և ավելի քիչ փոփոխված վիճակի բնության կողմից պահպանած պատկերը։ Հնարավոր է, որ այդ դրույթը ճիշտ է, բայց հազիվ թե երբևիցե կարելի կլինի այն ապացուցել։ Եթե, օրինակ, մենք տեսնում ենք, որ մեզ հայտնի ամենահին կաթնասունները, սողունները և ձկները խիստ կերպով վերցրած պատկանում են միևնույն դասերին, թեև այդ հին ձևերից մի քանիսները քիչ ավելի պակաս են տարբերվում միմյանցից, քան նույն խմբերի այժմյան տիպիկ ներկայացուցիչները, ապա զուր կլիներ որոնել ողնաշարավորների ընդհանուր սաղմնաբանական գծերն ունեցող կենդանիներ, քանի դեռ հայտնաբերված չեն բրածոներով հարուստ շերտեր կեմբրիյան ամենաստորին շերտերից ավելի ցած. սակայն այդ հայտնադործման վրա քիչ հույս կա։[69]
ԵՐՐՈՐԴԱՅԻՆ ԺԱՄԱՆԱԿԱՇՐՋԱՆԻ ԱՄԵՆԱՎԵՐՋԻՆ ԴԱՐԵՐԻ ԸՆԹԱՑՔՈՒՄ ԲՆԱԿՈՒԹՅԱՆ ՆՈՒՅՆ ՄԱՐԶԵՐԻ ՍԱՀՄԱՆՆԵՐՈՒՄ ՄԻԱՆՄԱՆ ՏԻՊԵՐԻ ՀԱՋՈՐԴԱԿԱՆՈՐԵՆ ԵՐԵՎԱՆ ԳԱԼՈՒ ՄԱՍԻՆ
Դեռ շատ տարիներ առաջ մ-ր Կլիֆտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 224] ցույց է տվել, որ ավստրալիական քարանձավներում գտնված բրածո կաթնասունները մերձավոր ազգակից են այդ մայր ցամաքի վրա այժմ ապրող պարկավորների հետ։ Հարավային Ամերիկայում նույնիսկ անպատրաստ դիտողը նկատում է նման հարաբերություններ՝ տեսնելով Լա-Պլատայի զանազան վայրերում գտնված զրահակրի զրահին նմանվող հսկա զրահի կտորները, և պրոֆեսոր Օուենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] զարմանալի կերպով ապացուցել է, որ այդպիսի առատությամբ այնտեղ թաղված բրածո կաթնասունների մեծ մասը պատկանում է հարավամերիկյան տիպին։ Այդ ազգակցությունն ավելի ևս պարզ կերպով հաստատվում է բրածո ոսկորների զարմանայի կոլեկցիայով, որ հավաքել են Լունդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 225] և Կլաուզենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 226] Բրազիլիայի քարանձավներում։ Այդ փաստերն ինձ վրա այնպիսի ուժեղ տպավորություն են գործել, որ 1839 և 1845 թվականներին ես հանդես եմ եկել որպես «տիպերի հաջորդականության օրենքի» միևնույն մայր ցամաքի վրա ապրածի և ապրողի այդ զարմանալի հարաբերակցության ջերմ պաշտպան։ Հետագայում պրոֆ. Օուենը նույն ընդհանրացումը տարածել է նաև Հին Աշխարհի կաթնասունների վրա։ Այդ հեղինակը նույն օրենքը երևան է հանում նաև Նոր Զելանդիայի մահաջնջված հսկա թռչունների վերաբերյալ իր տված ռեստավրացիաներում. մենք այդ նույնը տեսնում ենք նաև բրազիլիական քարանձավներում գտնված թռչունների վրա։ Միստեր Վուդվորդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 205] ցույց է տվել, որ այդ օրենքը կիրառելի է նաև ծովային խեցիների վերաբերմամբ, բայց կակղամորթների մեծամասնության լայն տարածման շնորհիվ այդ օրենքն այնքան էլ պարզորոշ կերպով չի արտահայտվում նրանց վրա։ Կարելի է նշել այլ օրինակներ էլ, օրինակ Մադերայի բրածո և այժմ ապրող քաղցրահամ ջրերի կակղամորթների միջև կամ Արալա-Կասպիական[70] ծովի բրածո և կիսաքաղցրահամ ջրերում այժմ ապրող խեցիների միջև եղած հարաբերակցությունը։
Բայց հատկապես ի՞նչ է արտահայտում միևնույն մարզերում և միևնույն տիպերի հաջորդականության այս ուշագրավ օրենքը։ Պետք է շատ համարձակ լինել, որպեսզի համեմատելով Ավստրալիայի և Հարավային Ամերիկայի միևնույն լայնության տակ գտնվող մասերի կլիման, վճռեինք բացատրել, մի կողմից, այդ երկու մայր ցամաքների բնակիչների իրար նման չլինելը ֆիզիկական պայմանների տարբերությամբ կամ, մյուս կողմից, պայմանների նմանությամբ բացատրել միևնույն տիպերի գոյությունը յուրաքանչյուր մայր ցամաքի վրա երրորդային ժամանակաշրջանի ամենավերջին դարաշրջաններում։ Չի կարելի վկայակոչել նաև այն, որ գոյություն ունի մի անխախտելի օրենք, որի համաձայն պարկավորները պետք է գլխավորապես կամ բացառապես երևան գային Ավստրալիայում, կամ թե թերատամները և այլ ամերիկական տիպեր պետք է ծագում առնեին միայն Ամերիկայում։ Մենք գիտենք, որ նախկին ժամանակներում Եվրոպան բնակված է եղել բազմաթիվ պարկավորներով, և վերևում հիշատակված աշխատության մեջ ես ցույց եմ տվել, որ Ամերիկայում ցամաքային կաթնասունների բաշխման օրենքը առաջներում այլ է եղել, քան այժմ։ Հյուսիսային Ամերիկան առաջներում ցուցաբերել է շատ ընդհանուր հատկանիշներ մայր ցամաքի այժմյան հարավային կեսի նկատմամբ, և մայր ցամաքի հարավային կեսը առաջներն ավելի սերտ հարաբերակցության մեջ է եղել հյուսիսային կեսի հետ, քան այժմ։ Նմանապես Ֆոկոների[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 73] և Կոտլիի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 227] հայտնագործություններից մենք գիտենք, որ հյուսիսային Հնդկաստանը իր կաթնասուն կենդանիներով առաջներն ավելի սերտ հարաբերակցության մեջ է եղել Աֆրիկայի հետ, քան այժմ։ Կարելի է նույնանման փաստեր բերել նաև ծովային կենդանիների տարածման մասին։
Փոփոխությունների միջոցով զարգացման տեսությունը ճիշտ կերպով բացատրում է միևնույն մարզերում միևնույն տիպերի երկարատև, բայց ոչ անպայման հաջորդաւկանության մեծ օրենքը, որովհետև աշխարհի յուրաքանչյուր մասի բնակիչները, իհարկե, պետք է ձգտեին թողնել նույն մարզում մոտ ազգակից, բայց որոշ չափով փոփոխված հաջորդներ նաև հետագա ամենամոտ ժամանակաշրջանի համար։ Եթե մի մայր ցամաքի բնակիչներն առաջները խիստ տարբերվել են մյուսի բնակիչներից, ապա նրանց փոփոխված հաջորդներն էլ շարունակում են տարբերվել գրեթե նույն ձևով և նույն չափով։ Սակայն խիստ երկար ժամանակամիջոցներ անցնելուց և աշխարհագրական այնպիսի մեծ փոփոխություններից հետո, որոնք հնարավոր են դարձնում բնակավայրերի փոփոխություն կատարել, ավելի թույլ ձևեր դուրս կմղվեն գերիշխող ձևերի կողմից, այնպես որ էակների տարածման գործում ոչ մի անփոփոխ բան չի լինի։
Գուցե որևէ մեկը կատակի համար կհարցնի՝ մի՞թե ես կարծում եմ, որ առաջները Հարավային Ամերիկայում ապրած մեգաթերիումը և նրան ազգակից այլ հսկայական հրեշները իրենցից հետո թողել են համրուկին, զրահակրին և մրջնակերին, որպես իրենց այլասերված հաջորդների։ Այդ ոչ մի րոպե չի կարելի ընդունել։ Այդ հսկա հրեշները իսպառ մահաջնջվել են և չեն թողել հաջորդներ։ Սակայն Բրազիլիայի քարայրներում գտնվել են շատ տեսակներ, որոնք մեծությամբ և այլ հատկանիշներով մոտ նմանություն ունեն Հարավային Ամերիկայում այժմ ապրող տեսակների հետ և այդ բրածոներից մի քանիսը կարող էին լինել այժմ ապրող տեսակների նախնիներ։ Չպետք է մոռանալ, որ մեր տեսության համաձայն միևնույն սեռին պատկանող բոլոր տեսակները որևէ մի տեսակի հաջորդներ են. այնպես որ եթե որևէ երկրաբանական ֆորմացիայում գտնվել են վեց սեռեր, յուրաքանչյուրը ութական տեսակներով, և նրան հաջորդող ֆորմացիայում՝ վեց այլ մերձավոր կամ իրար փոխարինող սեռեր նույնքան տեսակներով յուրաքանչյուրի մեջ, այդ դեպքում մենք կարող ենք սրանից եզրակացնել, որ ընդհանրապես ավելի հին սեռերից յուրաքանչյուրի միայն մեկ տեսակն է թողել փոփոխված հաջորդներ, որոնք էլ կազմում են զանազան տեսակներ պարփակող նոր սեռեր, յուրաքանչյուր հին սեռի մյուս յոթ տեսակները մահաջնջվել են և հաջորդներ չեն թողել. կամ, և այս ամենասովորական դեպքն է, վեց հին սեռերից միայն երկուսի կամ երեքի երկու կամ երեք տեսակները դառնում են նոր սեռերի նախահայրեր, իսկ այդ սեռերի մյուս տեսակները և այլ ամբողջական սեռեր իսպառ անհետանում են։ Դեպի անկում հակվող կարգերում, որոնց մեջ նկատվում է սեռերի և տեսակների թվի կրճատում, ինչպես հարավամերիկական թերատամնավորների մոտ, է՛լ ավելի քիչ թվով սեռեր ու տեսակներ թողնում են փոփոխված հաջորդներ։
ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ ՆԱԽՈՐԴ ԵՎ ՆԵՐԿԱ ԳԼՈՒԽՆԵՐԻՑ
Ես աշխատել եմ ցույց տալ, որ երկրաբանական տարեգրությունը վերին աստիճանի թերի է, որ երկրաբանները խնամքով հետազոտել են միայն երկրի մակերեսի մի փոքր մասը, որ օրգանիզմների մի քանի դասերը միայն առատությամբ են պահպանվել բրածո վիճակում, որ մեր թանգարաններում պահպանվող ինչպես նմուշների, այնպես էլ տեսակների թիվը միանգամայն աննշան է այն սերունդների թվի համեմատությամբ, որոնք հաջորդել են մեկը մյուսին նույնիսկ մեկ ֆորմացիայի ընթացքում։ Ես աշխատել եմ ցույց տալ, որ քանի որ ծովի հատակի իջնելը համարյա անհրաժեշտ պայման է նստվածքների կուտակման համար, որոնք պիտի լինեն բրածո բազմազան տեսակներով հարուստ և բավականաչափ հզոր, որպեսզի կարողանան դիմանալ հետագա քայքայմանը, ուստի մեր հաջորդական ֆորմացիաների միջև պետք է մեծ ժամանակամիջոցներ անցնեին, որ մահաջնջումը իջեցման ժամանակաշրջաններում պետք է որ գերակշռեր, իսկ փոփոխելիությունը՝ բարձրացման ժամանակաշրջաններում և որ այդ երկրորդ ժամանակաշրջաններից հուշարձաններ պահպանվել են ավելի թերի ձևով. որ յուրաքանչյուր առանձին ֆորմացիա անընդհատ կերպով չի կուտակվել․ որ յուրաքանչյուր ֆորմացիայի տևողությունը հավանորեն կարճ է տեսակային ձևերի միջին տևողության համեմատությամբ։ Այնուհետև, ես աշխատել եմ ցույց տալ, որ գաղթերը կարևոր դեր են կատարել որևէ մարզում և ֆորմացիայում նոր ձևերի առաջին անգամ երևան գալու գործում, որ հենց այդ լայն տարածված տեսակներն են ամենից շատ փոփոխվել և ամենից ավելի հաճախ սկիզբ տվել նոր տեսակների և որ տարատեսակները սկզբում եղել են տեղական։ Վերջապես, ես ձգտել եմ նաև ցույց տալ, որ թեև յուրաքանչյուր տեսակ պետք է անցներ բազմաթիվ անցումային ստադիաներ, այնուամենայնիվ, այն ժամանակաշրջանները, որոնց ընթացքում յուրաքանչյուր տեսակ ենթարկվել է փոփոխությունների, բազմաթիվ են ու երկար, եթե չափենք նրանց տարիներով, հավանորեն կարճատև են եղել այն ժամանակաշրջանների համեմատությամբ, որոնց ընթացքում յուրաքանչյուր տեսակ մնացել է առանց փոփոխությունների։ Այս պատճառները բոլորը միասին վերցրած մեծ չափով բացատրում են, թե ինչու մենք թեև գտնում ենք շատ անցումային օղակներ, բայց և այնպես չենք գտնում այն անհամար տարատեսակները, որոնք կկապակցեին բոլոր մահաջնջված և գոյություն ունեցող ձևերը միմյանց հետ նուրբ աստիճանական անցումներով։ Պետք է միշտ ինկատի ունենալ նաև այն, որ երկու ձևերի միջև գտնվող մի տարատեսակ մեզ հանդիպելիս կարող է համարվել որպես մի նոր ուրույն տեսակ, եթե միայն չի հաջողվել վերականգնել անցումների ամբողջ շղթան, որովհետև չի կարելի պնդել, թե կարծես մենք ունենք ճիշտ չափանիշ, որով կարելի է զանազանել տեսակները տարատեսակներից։
Ով որ չի համաձայնի ընդունել երկրաբանական տարեգրության անլրիվությունը, նա իրավացիորեն պետք է ժխտի նաև իմ տեսությունը, որովհետև նա իզուր կհարցնի՝ հապա ո՞ւր են այն անհամար անցումային օղակները, որոնք պետք է կապակցեին միևնույն ֆորմացիայի հաջորդական հարկերում գտնվող մոտ նմանություն ունեցող կամ փոխարինող տեսակները միմյանց հետ։ Նա կարող է չհավատալ, որ հսկայական ժամանակամիջոցներ պետք է անցած լինեն մեր հաջորդական ֆորմացիաների միջև, նա կարող է ուշադրություն չդարձնել այն կարևոր դերի վրա, որ կատարել են գաղթերը որևէ մեծ մարզի, օրինակ Եվրոպայի ֆորմացիաների վերաբերմամբ․ նա կարող է համառորեն մատնանշել տեսակների ամբողջ խմբերի տեսանելի, թեև հաճախ միայն սխալ կերպով տեսանելի, հանկարծակի երևան գալը։ Նա կարող է հարցնել, ո՞ւր են այն անհամար օրգանիզմների մնացորդները, որոնք պետք է գոյություն ունեցած լինեին կեմբրիյան սիստեմի նստվածքներից շատ առաջ։ Մենք այժմ գիտենք, որ գոնե մի կենդանի հաստատապես գոյություն է ունեցել այն ժամանակ․ բայց վերջին հարցին ես կարող եմ պատասխանել այն ենթադրությամբ, որ այնտեղ, որտեղ այժմ տարածվում են մեր օվկիանոսները, նրանք գոյություն են ունեցել հսկայական ժամանակաշրջանի ընթացքում և այնտեղ, որտեղ այժմ գտնվում են մեր տատանվող մայր ցամաքները, նրանք գտնվել են կեմբրիյան սիստեմի սկզբից, բայց այդ դարաշրջանից շատ առաջ աշխարհը ներկայացրել է բոլորովին այլ պատկեր և որ հին մայր ցամաքները, որ կազմված են ավելի հին ֆորմացիաներից, քան մեզ հայտնի բոլոր ֆորմացիաները, այժմ պահպանվել են միայն կերպարանափոխված վիճակում գտնվող մնացորդների ձևով կամ մինչև այժմ թաղված են օվկիանոսի տակ։
Մի կողմ թողնելով այդ դժվարությունները, մենք տեսնում ենք, որ հնէաբանությունը զարմանալի կերպով համաձայնվում է տեսակների ծագման և փոփոխականության ու բնական ընտրության միջոցով նրանց ձևափոխության վերաբերյալ տեսության հետ։ Մենք այսպիսով կարող ենք հասկանալ, թե ինչու նոր տեսակներն առաջանում են դանդաղորեն ու հաջորդականորեն. թե ինչու տարբեր դասերի տեսակները չեն փոփոխվում անպայման միասին, կամ միանման արագությամբ, կամ միանման չափով, բայց և այնպես ժամանակի ընթացքում բոլորն էլ որոշ չափով փոփոխվում են։ Հին ձևերի մահաջնջումը նորերի առաջացման գրեթե անխուսափելի հետևանք է։ Մենք կարող ենք հասկանալ, թե ինչու մի անգամ անհետացած տեսակն այլևս երբեք նորից երևան չի գալիս։ Տեսակների խմբերը թվով դանդաղ կերպով ավելանում են և գոյություն են ունենում անհավասար ժամանակամիջոցների ընթացքում, որովհետև փոփոխությունների պրոցեսը պետք է անպայման դանդաղ կերպով ընթանա և կախված է բազմաթիվ բարդ պատահականություններից։ Ընդարձակ և տիրապետող խմբերին պատկանող տիրապետող տեսակները ձգտում են թողնել բազմաթիվ փոփոխված հաջորդներ, որոնք առաջ են բերում նոր ենթախմբեր և խմբեր։ Նրանք առաջացման հետ զուգընթաց ավելի պակաս ուժեղ խմբերի տեսակները՝ որպես ընղհանուր նախահորից ժառանգած լինելով կազմվածքի որոշ անկատարելություն, ցուցաբերում են բոլորը միասին մահաշնջվելու տենդենց և չեն թողնում երկրի երեսին փոփոխված հաջորդներ։ Բայց տեսակների ամբողջ խմբի իսպառ մահաջնջումը երբեմն կարող է տեղի ունենալ շատ դանդաղ կերպով շնորհիվ այն հանգամանքի, որ սակավաթիվ հաջորդներ գերապրում են, երկար մնալով պաշտպանված և մեկուսացած վայրերում։ Եթե որևէ խումբ բոլորովին անհետացել է, նա կրկին երևան չի գա, որովհետև սերունդների շղթան ընդհատված է։
Մեզ համար հասկանալի է դառնում, որ տիրապետող ձևերը, որոնք լայն կերպով տարածված են և տալիս են ամենամեծ թվով տարատեսակներ, ձգտում են աշխարհը լցնել իրենց մերձավոր, բայց փոփոխված հաջորդներով, և այս վերջիններս սովորաբար հաջողությամբ դուրս են մղում գոյության կռվում իրենց առջև տեղի տվող խմբերին։ Դրա հետևանքով էլ երկար ժամանակամիջոցներից հետո երկրագնդի օրգանական բնակչությունը թվում է միաժամանակ փոփոխված։
Մեզ հասկանալի է դառնում, թե ինչու կյանքի բոլոր հին ու նոր ձևերը բոլորը միասին դասավորվում են սակավաթիվ մեծ դասերի մեջ։ Մեզ հասկանալի է դառնում, որ որքան որևէ ձև ավելի հին է, այնքան ավելի՝ հատկանիշների տարամիտման մշտական ձգտում ունենալու շնորհիվ սովորաբար տարբերվում է այժմ ապրողից․ ինչու հին և մահաջնջված ձևերը հաճախ լցնում են գոյություն ունեցող ձևերի միջև եղած տարածությունները, երբեմն մի խմբի մեջ կապակցում են երկու խմբեր, որոնք առաջ համարվել են առանձին խմբեր, բայց ավելի հաճախ միայն փոքր ինչ ավելի մոտեցնում են իրանց միմյանց։ Որքան որևէ ձև հին է, այնքան ավելի հաճախ նա գրավում է այնպիսի դրություն, որը որոշ չափով միջակա է այժմ առանձին կանգնած խմբերի միջև, որովհետև որքան հին է ձևը, այնքան նա ավելի սերտորեն կապված է, հետևապես և ավելի նման է այն խմբերի նախահորը, որոնք վաղուց արդեն իրարից հեռացել են։ Մահաջնջմած ձևերը հազվագյուտ դեպքերում են գրավում այժմ գոյություն ունեցող ձևերի միջև ուղիղ միջին տեղը, բայց նրանք միջակա են, որոնք երկար ու զարտուղի ճանապարհով անցնում են զանազան այլ մահաջնջված ու իրենցից տարբերվող ձևերի միջով։ Մեզ համար պարզ է դառնում, թե ինչու մեկը մյուսին անմիջականորեն հաջորդող ֆորմացիաների օրգանական մնացորդները մոտ նմանություն ունեն միմյանց հետ, որովհետև նրանց միջև գոյություն ունի մոտ ազգակցական կապ։ Մեզ համար միանգամայն հասկանալի է, թե ինչու որևէ միջակա ֆորմացիայից վերցված օրգանական մնացորդները իրենց հատկանիշներով էլ գրավում են միջակա տեղ։
Աշխարհի բնակիչները նրա պատմության յուրաքանչյուր հաջորդական ժամանակաշրջանում գոյության կռվի մեջ հաղթել են իրենց նախորդներին, այդ իմաստով նրանք ավելի բարձր են իրենց նախորդներից և կազմվածքով սովորաբար ավելի մասնագիտացած են. գուցե դրանով է բացատրվում այն ընդհանուր կարծիքը, որին համաձայն են հնէաբաններից շատերը, որ կազմվածքը ամբողջությամբ վերցրած զարգացել է առաջադիմորեն։ Մահաջնջված և հին կենդանիները որոշ չափով նման են նույն դասերին պատկանող ավելի նոր կենդանիների սաղմերին, և այդ զարմանալի փաստը մեր տեսության տեսակետից պարզ բացատրություն է ստանում։ Միևնույն մարզերում կազմվածքի միևնույն տիպերի հաջորդականությունը հետագա երկրաբանական ժամանակաշրջաններում դադարում է հանելուկային լինելուց և հասկանալի է դառնում, ժառանգականության սկզբունքի տեսակետից։
Ուրեմն, եթե երկրաբանական տարեգրությունը այդքան թերի է, ինչպես շատերը կարծում են,— բայց կարելի է առնվազն պնդել, որ չի կարելի պաշտպանել նրա լրիվությունը,— ապա բնական ընտրության տեսության դեմ եղած գլխավոր առարկությունները զգալի չափով թուլանում են կամ վերանում։ Մյուս կողմից, հնէաբանության բոլոր գլխավոր օրենքները պարզ կերպով, ինչպես ինձ թվում է, վկայում են այն մասին, որ տեսակները ծագել են բնական ծննդի միջոցով, ըստ որում հին ձևերը դուրս են մղվել կյանքի նոր և կատարելագործված ձևերի կողմից, որոնք ծագել են փոփոխականության և ամենալավ հարմարվածների գերապրելու շնորհիվ։
ԳԼՈՒԽ XII։ ԱՇԽԱՐՀԱԳՐԱԿԱՆ ՏԱՐԱԾՈՒՄ
ԱՐԴԻ ՏԱՐԱԾՈՒՄԸ ՉԻ ԿԱՐՈՂ ԲԱՑԱՏՐՎԵԼ ՖԻԶԻԿԱԿԱՆ ՊԱՅՄԱՆՆԵՐՈՒՄ ԵՂԱԾ ՏԱՐԲԵՐՈՒԹՅՈՒՆՆԵՐՈՎ։— ԱՐԳԵԼՔՆԵՐԻ ՆՇԱՆԱԿՈՒԹՅՈՒՆԸ։— ՄԻԵՎՆՈՒՅՆ ՄԱՅՐ ՑԱՄԱՔԻ ՏՆԱԿԱՆ ԱՐՏԱԴՐԱՆՔՆԵՐԻ ՆՄԱՆՈՒԹՅՈՒՆԸ։— ՍՏԵՂԾԱԳՈՐԾՈՒԹՅԱՆ ԿԵՆՏՐՈՆՆԵՐԸ։— ՏԱՐԱԲՆԱԿԵՑՈՒՄԸ ՈՐՊԵՍ ԿԼԻՄԱՅԻ, ՑԱՄԱՔԻ ՄԱԿԵՐԵՎՈՒՅԹԻ ԵՎ ՊԱՏԱՀԱԿԱՆ ՊԱՏՃԱՌՆԵՐԻ ՀԵՏԵՎԱՆՔ։— ՏԱՐԱԲՆԱԿԵՑՈՒՄԸ ՍԱՌՑԱԴԱՇՏԱՅԻՆ ԺԱՄԱՆԱԿԱՇՐՋԱՆԻ ԸՆԹԱՑՔՈՒՄ։— ՍԱՌՑԱԴԱՇՏԱՅԻՆ ԺԱՄԱՆԱԿԱՇՐՋԱՆՆԵՐԻ ՀԵՐԹԱՓՈԽՈՒՄԸ ՀՅՈՒՍԻՍՈՒՄ ԵՎ ՀԱՐԱՎՈՒՄ։
Երկրագնդի մակերևույթի վրա կազմակերպված էակների տարածումը ուսումնասիրելիս մեր ուշադրությունն ամենից առաջ իր վրա է գրավում այն ուշագրավ փաստը, որ զանազան մարզերի բնակիչների ոչ նման լինելը և ոչ էլ նման լինելը չի կարելի լիովին բացատրել կլիմայական և այլ ֆիզիկական պայմաններով։
Վերջերս այդպիսի եզրակացության է եկել այդ հարցով զբաղվողներից համարյա թե ամեն մեկը։ Եվ նրա ճշտությունը ապացուցելու համար բավական է բերել թեկուզ Ամերիկայի ներկայացրած օրինակը. եթե մենք հանենք նրա բևեռային և հյուսիսային բարեխառն մասերը, ապա բոլոր հեղինակները համաձայն են այն բանում, որ օրգանիզմների աշխարհագրական տարածման մեջ ամենահիմնական բաժանումներից մեկը Նոր և Հին աշխարհների միջև եղած տարբերությունն է։ Սակայն ճանապարհորդելով Ամերիկայի ընդարձակ մայր ցամաքի վրա՝ սկսած Միացյալ Նահանգների կենտրոնական մասերից և վերջացրած նրա ամենահարավային կետով, մենք հանդիպում ենք ամենաբազմազան ֆիզիկական պայմանների, ինչպիսիք միայն կարող են ունենալ խոնավ մարզերը, ջրազուրկ անապատները, բարձր լեռները, խոտավետ հարթավայրերը, անտառները, ճահիճները, լճերը և մեծ գետերը գրեթե ամեն մի ջերմաստիճաններով։ Հին աշխարհում հազիվ թե գտնվեին այնպիսի կյիմայական կամ որևէ այլ ֆիզիկական պայմաններ, որոնց համապատասխանը չգտնվեր Նոր աշխարհում, գոնե այնքան, որքան այդ անհրաժեշտ է միևնույն տեսակի համար։ Իհարկե, Հին աշխարհում կարելի է նշել ավելի տաք երկրամասեր, քան որևէ մեկը նոր աշխարհում, սակայն նրանց բնակչությունը չի տարբերվում շրջապատող մարզերի բնակչությունից, որովհետև հազվադեպ է, որ օրգանիզմների մի խումբ հարմարված է լինում մի փոքրիկ երկրամասի, որը տարբերվում է միայն փոքր ինչ յուրահատուկ պայմաններով։ Բայց, չնայած Նոր և Հին աշխարհների ֆիզիկական պայմաններում եղած այդ ընդհանուր զուգահեռությանը, ինչքա՜ն մեծ է նրանց բնակիչների միջև եղած տարբերությունը։
Եթե դառնանք հարավային կիսագնդին և համեմատության համար վերցնենք ցամաքի լայնածավալ տարածությունները, որ ներկայացնում են Ավստրալիան, Հարավային Աֆրիկան և Հարավային Ամերիկայի արևմտյան մասերը՝ լայնության 25° և 35° միջև, ապա կարելի է գտնել մարզեր, որոնք ծայր աստիճան նման են իրենց ֆիզիկական պայմաններով, բայց անհնարին է ցույց տալ երեք ֆաունա կամ ֆլորա, որոնք հիմնական կերպով այդչափ տարբեր լինեն միմյանցից։ Այնուհետև համեմատելով Հարավային Ամերիկայի 35° լայնությունից հարավ ընկած երկրների բնական արտադրանքները լայնության 25°-ից հյուսիս գտնվող երկրների համապատասխան արտադրանքների հետ, հետևաբար լայնության տասը աստիճան տարածությամբ միմյանցից բաժանված և խիստ տարբեր պայմաններում գտնվող էակները, մենք կգտնենք, որ նրանք անհամեմատ ավելի մոտ են միմյանց, քան Ավստրալիայի և Աֆրիկայի գրեթե միևնույն կլիմայական պայմաններում գտնվող բնական արտադրանքները։ Նույնանման փաստեր կարելի է բերել նաև ծովերի բնակչության վերաբերմամբ։
Մյուս կարևոր փաստը, որը մեզ զարմացնում է մեր ընդհանուր ակնարկի ժամանակ, այդ այն է, որ ազատ տեղափոխության համար հնարավոր արգելքները կամ խոչընդոտները ամենասերտ առնչության մեջ են գտնվում զանազան մարզերի բնական արտադրանքների հետ։ Մենք այդ տեսնում ենք Նոր և Հին աշխարհների գրեթե բոլոր ցամաքային ձևերի տարբերության մեջ, բացառելով նրանց հյուսիսային մասերը, որտեղ երկուսի ցամաքները համարյա միանում են իրար հետ և որտեղ փոքր ինչ տարբեր կլիմայի դեպքում կարող էր տեղի ունենալ բարեխառն գոտու հյուսիսային մասերի ձևերի ազատ տեղափոխություն, այնպես, ինչպես այժմ այդ տեղի է ունենում միայն իսկական բևեռային ձևերի համար։ Մենք նույնը տեսնում ենք Ավստրալիայի, Աֆրիկայի և Հարավային Ամերիկայի միևնույն լայնությունից վերցրած բնակիչների միջև եղած մեծ տարբերության մեջ, որովհետև այդ երկրները մեկուսացած են մեկը մյուսից այնքան, որքան որ դա հնարավոր է։ Վերջապես, նույն փաստին մենք հանդիպում ենք յուրաքանչյուր մայր ցամաքի վրա. բարձր և երկայնաձիգ լեռնաշղթաների, լայնածավալ անապատների, նույնիսկ մեծ գետերի հակադիր կողմերում մենք գտնում ենք զանազան ձևեր, բայց քանի որ լեռնաշղթաները, անապատները և այլն այնքան անանցանելի չեն, ինչքան մայր ցամաքները բաժանող օվկիանոսները, և ամենայն հավանականությամբ այնքան երկար ժամանակ գոյություն չունեն, դրա համար էլ այդ դեպքերում տարբերությունը շատ ավելի փոքր է, քան զանազան մայր ցամաքների միջև եղած տարբերությունը։
Դառնալով օվկիանոսին, մենք հանդիպում ենք նույն օրենքին։ Հարավային Ամերիկայի արևելյան ու արևմտյան ափերի ծովային բնակիչները շատ տարբեր են, բացառելով երկու ափերի համար ընդհանուր սակավաթիվ կակղամորթները, խեցեմորթները և փշամորթները. բայց դ-ր Գյունտերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 154] վերջերս ցույց է տվել, որ Պանամայի պարանոցի հակադիր ափերում ապրող ձկների մոտավորապես 30% նույնն է, որը հարկադրել է բնախույզներին ենթադրելու, որ պարանոցը նախկին ժամանակներում կտրված է եղել։ Հարավային Ամերիկայի ափերից դեպի արևմուտք գտնվում է բաց օվկիանոսի ընդարձակ տարածությունը, որտեղ ոչ մի կղզի չկա, որը կարող էր կայան լինել գաղթողների համար, այս դեպքում մենք հանդիպում ենք մի այլ ձևի արգելքի, և հենց որ մենք անցնում ենք այդ, Խաղաղ օվկիանոսի արևելյան կղզիների վրա հանդիպում ենք մի այլ, և այն էլ միանգամայն տարբեր, կենդանական աշխարհի։ Այսպիսով, երեք ծովային կենդանական աշխարհներ ձգվում են դեպի հյուսիս և հարավ զուգահեռ կերպով և իրարից ոչ հեռու, համապատասխանող կլիմայական պայմաններում, բայց մեկը մյուսից անջատված լինելով անհաղթահարելի արգելքներով, լինի դա ցամաք կամ բաց ծով, նրանք գրեթե բոլորովին տարբեր են։ Մյուս կողմից, գնալով Խաղաղ օվկիանոսի արևադարձային մասերի արևելյան կղզիներից ավելի դեպի արևմուտք, մենք անհաղթահարելի արգելքների չենք հանդիպում և գտնում ենք կամ անթիվ կղզիներ որպես կայաններ, կամ չընդհատվող ափային գիծ, մինչև որ վերջապես կտրում ենք կիսագունդը և մոտենում Աֆրիկայի ափերին։ Եվ այդ ամբողջ լայնածավալ տարածության վրա մենք չենք հանդիպում սուր կերպով պարագծված առանձին ծովային կենդանական աշխարհների։ Թեև շատ քիչ ծովային կենդանիներ ընդհանուր են Ամերիկայի արևելյան և արևմտյան ափերի և Խաղաղ օվկիանոսի կղզիների վերոհիշյալ երեք հարևան ֆաունաների համար, սակայն մի քանի ձկներ Խաղաղ օվկիանոսից մտնում են Հնդկականը և շատ կակղամորթներ ընդհանուր են Խաղաղ օվկիանոսի արևելյան կղզիների և Աֆրիկայի արևելյան ափերի համար, որոնք գտնվում են գրեթե հակադիր միջօրեականների տակ։
Երրորդ կարևոր փաստը, որը մասամբ արդեն ընդգրկվել է նախորդ տվյալներում, միևնույն մայր ցամաքի կամ ծովի բնական արտադրանքների նմանության մեջ է, թեև տեսակներն իրենք զանազան վայրերում և կայաններում կարող են տարբեր լինել։ Դա մի չափազանց ընդհանուր օրենք է, որն ապացուցվում է անթիվ օրինակներով յուրաքանչյուր մայր ցամաքի վրա։ Այնուամենայնիվ, բնախույզը ճանապարհորդելով, օրինակ, հյուսիսից դեպի հարավ, միշտ զարմանում է, թե ինչպես մերձավոր, բայց տեսակային տեսակետից տարբեր հաջորդական խմբերը փոխարինում են մեկը մյուսին։ Նա լսում է, որ թռչունների մերձավոր, բայց և այնպես տարբեր տեսակներն ունենում են գրեթե միևնույն ձայնը և տեսնում է, որ նրանց բները շատ նման են, բայց իրենց կառուցվածքով ոչ բոլորովին նույնն են, և նրանց ձվերի գույնն էլ համարյա միանման է։ Մագելլանի նեղուցի մոտ փռվող հարթավայրերում բնակվում է ամերիկական ջայլամի մի տեսակը, իսկ Լա-Պլատայի ավելի հյուսիս ընկած հարթավայրերում՝ միևնույն սեռի մի այլ տեսակ, բայց ոչ իսկական ջայլամ և ոչ էլ էմու, որոնք նման են Աֆրիկայում և Ավստրալիայում նույն լայնության տակ ապրողներին։ Լա-Պլատայի նույն հարթավայրերում մենք տեսնում ենք ագուտիին և վիսկաշային, որոնք իրենց բարքերով շատ նման են մեր նապաստակներին ու ճագարներին և պատկանում են կրծողների նույն կարգին, բայց ունեն կազմվածքի ցայտուն կերպով արտահայտված ամերիկական տիպ։ Բարձրանալով Կորդիլերների բարձր գագաթները, մենք գտնում ենք վիսկաշայի ալպիական տեսակը, ջրերում՝ կուղբի և մշկամկան փոխարեն՝ հարավ ամերիկական տիպի կրծողներ կոիպուին և կապիբարային։ Կարելի է բերել անթիվ քանակությամբ նաև այլ օրինակներ։ Այսպես, ամերիկական ափերի մոտ գտնվող կղզիների բնակիչները, ինչքան էլ այդ կղզիները երկրաբանական տեսակետից տարբեր լինեն, միշտ ունենում են խիստ արտահայտված ամերիկական տիպ, թեև կարող են ներկայացված լինել տարբեր տեսակներով, որոնք հատուկ են առանձին կղզիների։ Դառնալով հեռավոր անցյալին, ինչպես այդ ցույց է տրված վերջին գլխում, մենք գտնում ենք, որ ամերիկական տիպը տիրապետող է ինչպես ամերիկական մայր ցամաքի վրա, այնպես էլ ամերիկական ծովերում։ Այդ փաստերում մենք տեսնում ենք այն խոր օրգանական կապը, որ գոյություն ունի ժամանակի և տարածության մեջ ցամաքի կամ ջրի մի որոշ տարածության վրա՝ անկախ ֆիզիկական պայմաններից։ Եվ քննախույզ չէ այն բնագետը, որն իր առաջ հարց չի դնում, թե ինչ է նշանակում այդ կապը։
Այդ կապը պարզապես ժառանգականության արտահայտությունն է, որով միայն, ինչպես մենք այղ հաստատապես գիտենք, պայմանավորված է կամ իրար միանգամայն նման, կամ, ինչպես մենք այդ տեսնում ենք տարատեսակների դեպքում, առավել կամ պակաս նմանություն ունեցող օրգանիզմների երևան գալը։ Զանազան մարզերի բնակիչների միջև եղած տարբերությունը կարող է վերագրվել նրանց փոփոխությանը, որն առաջանում է փոփոխականության և բնական ազդեցության տակ, և հավանորեն ավելի նվազ չափով զանազան ֆիզիկական պայմանների որոշակի ազդեցությանը։ Չնմանվելու աստիճանը պայմանավորվում է նրանով, թե ինչքան լրիվ չափով է կանխվել տիրապետող ձևերի գաղթը մի մարզից մյուսը շատ թե քիչ հեռավոր դարաշրջանում, առաջին գաղթականների առանձնահատկություններով ու քանակությամբ և բնակիչների մի մասի ազդեցությամբ մյուսների վրա՝ զանազան փոփոխությունների պահպանման իմաստով, որովհետև գոյության կռվում օրգանիզմների փոխհարաբերությունները, ինչպես ես այդ արդեն բազմիցս նշել եմ, կազմում են կենսապայմաններից ամենակարևորը։ Սրանից հետևում է գաղթը կասեցնող արգելքների կարևոր նշանակությունը, այնպես, ինչպես ժամանակը կարևոր է բնական ընտրության ազդեցության տակ փոփոխության դանդաղ պրոցեսի համար։ Լայնորեն տարածված, անհատներով հարուստ տեսակները, որոնք իրենց հիմնական ընդարձակ մարզերում արդեն հաղթել են բազմաթիվ մրցակիցներին, ունենում են նաև լավագույն շանսեր նոր տեղեր գրավելու համար, երբ նրանք ընկնում են նոր երկրներ։ Իրենց նոր բնակավայրերում նրանք ենթարկվում են իրենց համար նոր կենսապայմաններին և այդ պատճառով հաճախ տալիս են նոր փոփոխություններ և հարմարումներ, դրա հետևանքով նրանք է՛լ ավելի հաղթելու ընդունակ են դառնում և առաջացնում են ձևափոխված հաջորդների խմբեր։ Հիմնվելով փոփոխելիության հետ միասին ժառանգականության այդ սկզբունքի վրա, կարելի է հասկանալ այնքան սովորական ու լավ հայտնի փաստը, որ սեռի ստորաբաժանումները, ամբողջ սեռերը և նույնիսկ ընտանիքները իրենց տարածմամբ սահմանափակված են միևնույն մարզերով։
Ինչպես նկատվել է վերջին գլխում, մենք ոչ մի ցուցում չունենք, որ գոյություն ունի անհրաժեշտ զարգացման օրենք։ Քանի որ փոփոխականության հակումը յուրաքանչյուր տեսակի անկախ հատկությունն է և բնական ընտրության կողմից կօգտագործվի միայն այնքան ժամանակ, քանի դեռ նա նպաստում է ամեն մի անհատի նրա գոյության բարդ կռվի մեջ, ուստի զանազան տեսակների փոփոխականության սահմանները չեն կարող միանման լինել։ Եթե որոշ թվով տեսակներ, որոնք երկար ժամանակ ապրել են միասին և իրենց բուն մարզում մրցել իրար հետ, մասսայորեն գաղթել են մի նոր և հետագայում առանձնացած մարզ, այդ դեպքում նրանց փոփոխությունը չի կարող մեծ լինել, որովհետև ոչ գաղթը, ոչ էլ մեկուսանալը ինքնին չեն կարող որևէ բան առաջ բերել այդ ուղղությամբ։ Այդ գործոնները նշանակություն ունեն միայն այն դեպքում, երբ օրգանիզմները դրվում են նոր պայմանների մեջ միմյանց նկատմամբ և արդեն ավելի փոքր չափով էլ շրջապատող ֆիզիկական պայմանների նկատմամբ։ Ինչպես որ որոշ ձևեր պահպանել են իրենց առանձնահատկությունները համարյա անփոփոխ վիճակում՝ սկսած չափազանց հեռավոր երկրաբանական ժամանակաշրջանից, ինչպես մենք տեսանք վերջին գլխում, այնպես էլ որոշ տեսակներ տարաբնակվել են ընդարձակ տարածությունների վրա, մնալով գրեթե կամ բոլորովին անփոփոխ։
Այս տեսակետների համաձայն ակներև է, որ միևնույն սեռի տարբեր տեսակները, թեկուզ նրանք գրավեին երկրագնդի ամենահեռավոր տեղամասերը, պետք է ծագում առնեին մի տեղից, որովհետև առաջացել են մի ընդհանուր նախածնողից։ Այն տեսակների վերաբերմամբ, որոնք երկրաբանական ամբողջ շրջանների ընթացքում միայն աննշան փոփոխություններ են կրել, դժվար չէ ապացուցել, որ նրանք մի մարզից տարաբնակվել են այն պատճառով, որ ամենահին ժամանակներից սկսած տեղի ունեցած աշխարհագրական ու կլիմայական զգալի փոփոխությունների ընթացքում գաղթեր կարող էին տեղի ունենալ համարյա անսահմանափակ չափերով։ Բայց այլ շատ դեպքերում, երբ մենք հիմք ունենք կարծելու, որ մի սեռի տեսակները առաջացել են համեմատաբար նոր ժամանակներում, այդ մեծ դժվարություններ է ներկայացնում։ Ճիշտ այդպես էլ ակներև է, որ միևնույն տեսակի անհատները, որոնք թեև ներկայումս ապրում են հեռավոր ու մեկուսացած մարզերում, պետք է առաջացած լինեն մի տեղից, որտեղ սկզբում ապրել են նրանց ծնողները, որովհետև, ինչպես այդ բացատրվել է, անհավանական է, որ միմյանց միանգամայն նման անհատները կարող էին առաջանալ զանազան տեսակների պատկանող ծնողներից։
ԵՆԹԱԴՐՅԱԼ ՍՏԵՂԾԱԳՈՐԾՈՒԹՅԱՆ ԿԵՆՏՐՈՆԻ ՄԻԱՍՆԱԿԱՆՈՒԹՅՈՒՆԸ
Մենք մոտեցել ենք այն հարցին, որը բազմիցս քննարկվել է բնախույզների կողմից, այն է՝ թե արդյոք յուրաքանչյուր տեսակ ստեղծվել է երկրագնդի երեսին մե՞կ, թե մի քանի տեղերում։ Իհարկե, կան դեպքեր, երբ շատ դժվար է ըմբռնել, թե ինչ կերպով տեսակը որևէ տեղից կարող էր տարածվել մինչև հեռավոր ու մեկուսացած վայրերը, որտեղ նա այժմ գտնվում է։ Այնուամենայնիվ, այդ տեսակետի պարզությունը, ըստ որի յուրաքանչյուր տեսակ սկզբում երևացել է միայն մեկ մարզում, գերում է մեր միտքը։ Նա, ով մի կողմ է նետում այդ տեսակետը, հրաժարվում է ըմբռնել, թե ինչումն է տեսակի սովորական ծննդի և նրան հաջորդող տարաբնակման vera causa-ն[71], օգնության է կանչում հրաշագործությանը։ Առհասարակ ընդունում են, որ մեծ մասամբ յուրաքանչյուր տեսակի մարզը լինում է համատարած, և եթե կենդանին կամ բույսը հանդիպում են երկու կետերում, որոնք այնքան հեռացած են մեկը մյուսից կամ բաժանված են այնպիսի տարածությամբ, որ դա տեղափոխության համար խոչընդոտ է հանդիսանում, ապա այդպիսի դեպքը ցուցադրվում է որպես ինչ որ ուշագրավ ու բացառիկ բան։ Կաթնասուն կենդանիների համար ծովի ընդարձակ տարածության վրայով տեղափոխվելու անընդունակությունը գուցե ավելի պարզորոշ է, քան որևէ մի այլ էակի համար, բայց և այնպես մենք ոչ մի անբացատրելի բան չենք տեսնում, որ միևնույն կաթնասունները կան երկրագնդի զանազան կետերում։ Ոչ մի երկրաբան դժվարություն չի տեսնում այն բանում, որ Բրիտանիայի կղզիները Եվրոպայի հետ ունեն ընդհանուր կաթնասուններ, որովհետև կասկած չկա, որ նրանք իրար հետ մի ժամանակ միացած են եղել։ Բայց եթե միևնույն տեսակը կարող է երևան գալ երկու տարբեր կետերում, հապա ինչո՞ւ մենք ոչ մի կաթնասուն չենք գտնում, որը ընդհանուր լիներ Եվրոպայի և Ավստրալիայի կամ Հարավային Ամերիկայի համար։ Գոյության պայմանները այնտեղ ու այստեղ այնքան միանման են, որ եվրոպական բազմաթիվ կենդանիներ ու բույսեր նատուրալիզացիայի են ենթարկվել Ամերիկայում և Ավստրալիայում, և հյուսիսային ու հարավային կիսագնդերի այդ հեռավոր կետերից վերցրած մի քանի բույսեր բոլորովին նույնական են։ Դրա պատասխանը, ես կարծում եմ, այն է, որ կաթնասուններն անընդունակ են տեղափոխվելու ընդարձակ ընդհատվող տարածությունների վրայով, մինչդեռ բույսերը կատարել են այդ տեղափոխությունը տարածման համար իրենց ունեցած զանազան միջոցների շնորհիվ։ Բազմազան արգելքների հսկայական և կարևոր նշանակությունը հասկանալի է միայն այն դեպքում, եթե մենք կանգնած ենք այն տեսակետի վրա, որ տեսակների մեծամասնությունը երևացել է արգելքի մի կողմում և չի կարողացել տեղափոխվել մյուս կողմը։ Փոքրաթիվ ընտանիքներ, շատ ենթաընտանիքներ, խիստ շատ սեռեր և էլ ավելի մեծ թվով սեռային ստորաբաժանումներ հարմարված են մի միասնական մարզի, բացի դրանից, շատ բնախույզներ նկատել են, որ ամենաբնական սեռերը, այսինքն այնպիսիները, որոնց տեսակները ամենից ավելի մոտ են իրար, սովորաբար կապված են մի մարզի հետ, կամ եթե ունեն լայն տարածում, ապա գրավում են անընդմիջվող մարզեր։ Շատ տարօրինակ կլիներ, եթե դրան միանգամայն հակառակը նկատվեր այն դեպքում, երբ մենք նույն շարքում իջնում ենք ավելի ևս ցած, հասնում ենք միևնույն տեսակի անհատներին և հանկարծ նրանք, թեկուզ սկզբում, սահմանափակված չեն լինում մի մարզով։
Այդ պատճառով էլ ինձ, ինչպես նաև շատ ուրիշ բնախույզների, թվում է, որ ամենից ավելի հավանական է այն տեսակետը, ըստ որի յուրաքանչյուր տեսակ սկզբում երևան է եկել միայն մի մարզում և հետո այստեղից տարածվել է այնքան հեռու, որքան այդ նրան թույլ են տվել նրա ճանապարհորդելու միջոցները և անցյալ ու ներկա ժամանակների պայմանները։ Անկասկած է, որ կան շատ դեպքեր, երբ մենք չենք կարող բացատրել, թե ինչպես միևնույն տեսակը կարող էր մի կետից ընկնել մյուսը։ Բայց աշխարհագրական և կլիմայական փոփոխությունները, որոնք հավանորեն տեղի են ունեցել ամենավերջին երկրաբանական ժամանակշրջանների ընթացքում, կարող էին շատ տեսակների համար ընդմիջվող դարձնել սկզբում համատարած մարզերը։ Այսպիսով, մենք պետք է քննարկենք, թե արդյոք որքան բազմաթիվ են և արդյոք իրենց նշանակությամբ այդքան կարևոր են բացառություններն այն կանոնից, որ յուրաքանչյուր տեսակ ունի տարածման համատարած մարզ, որպեսզի կարողանան հարկադրել մեզ հրաժարվել վերոհիշյալ ընդհանուր կշռադատումների հիման վրա ճշմարտացի տեսակետից, որ տեսակն առաջացել է մի մարզում և այստեղից տարածվել է այնքան հեռու, որքան այդ հնարավոր է եղել։ Չափազանց տաղտկալի կլիներ քննարկել բոլոր այն առանձին դեպքերը, երբ այժմ ապրող տեսակները գրավում են տարբեր մեկուսացած մարզեր, և ես ոչ մի րոպե չեմ հավակնում շատ դեպքերում գեթ որևէ բացատրություն տալու հնարավորության վրա։ Բայց որոշ նախնական դիտողություններից հետո ես կվերլուծեմ փաստերի մի քանի ամենաուշագրավ խմբեր, այսինքն՝ նախ միևնույն տեսակի գոյությունը իրարից հեռու գտնվող լեռնաշղթաների գագաթների վրա և արկտիկայի ու անտարկտիկայի մարզերի զանազան մասերում, երկրորդ (հաջորդ գլխում), քաղցրահամ ջրերի էակների լայն տարածումը և երրորդ, ցամաքային միևնույն տեսակների գոյությունը կղզիների և ամենամոտ մայր ցամաքների վրա, որոնք երբեմն միմյանցից բաժանված են բաց ծովի հարյուրավոր մղոն տարածությամբ։ Եթե միևնույն տեսակի գոյությունը երկրագնդի հեռավոր ու մեկուսացած կետերում կարող է շատ դեպքերում բացատրվել այն տեսակետի հիման վրա, որ յուրաքանչյուր տեսակ տարածվել է մի հիմնական տեղից, ապա, նաև ուշադրության առնելով անցած կլիմայական ու աշխարհագրական փոփոխությունների մասին մեր անտեղյակ լինելը և տեղափոխության պատահական եղանակները, այն գաղափարը, որ յուրաքանչյուր տեսակի ծագման կենտրոնի միասնականությունը օրենք է, ինձ թվում է, համեմատաբար ամենից ավելի վստահելի։
Քննարկելով այդ հարցը, մենք միևնույն ժամանակ պետք է կանգ առնենք մեզ համար մի շատ կարևոր կետի վրա, այն է՝ կարո՞ղ են արդյոք մի սեռի պատկանող զանազան տեսակները, որոնք մեր տեսության համաձայն առաջացել են մի ընդհանուր նախածնողից, տարաբնակվել մի մարզից, տարաբնակման ժամանակ ենթարկվելով փոփոխությունների։ Եթե այն դեպքում, երբ մի մարզի տեսակների մեծ մասը տարբերվում է մյուս մարզի թեկուզև մերձավոր տեսակներից, հաջողվի ապացուցել, որ գաղթը մի մարզից մյուսը տեղի է ունեցել հավանորեն ավելի վաղ ժամանակաշրջանում, ապա մեր ընդհանուր եզրակացությունը զգալի չափով կհաստատվի, որովհետև այդպիսի բացատրությունը համաձայնվում է փոփոխությունների միջոցով ծագում առնելու սկզբունքի հետ։ Օրինակ, մի հրաբխային կղզի, որ բարձրացել և գոյացել է մայր ցամաքից մի քանի հարյուր մղոն հեռավորության վրա, ժամանակի ընթացքում, հավանորեն, նրանից կարող էր ստանալ փոքրաթիվ գաղթողներ, և վերջինների սերունդները, թեև փոփոխված, բայց և այնպես ժառանգականության շնորհիվ կպահպանեին իրենց ազգակցական կապը մայր ցամաքի բնակիչների հետ։ Նման դեպքերը սովորական են և, ինչպես մենք կտեսնենք հետագայում, անկախ ստեղծման տեսակետից, անբացատրելի են։ Մի մարզի տեսակների մի այլ մարզի տեսակների հետ ունեցած առնչության վերաբերյալ այդպիսի տեսակետը էականորեն չի տարբերվում մ-ր Ուոլլեսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] կարծիքից, որը գալիս է այն եզրակացության, որ «յուրաքանչյուր տեսակ իր գոյությունն սկսում է թե՛ ժամանակի և թե՛ տարածության տեսակետից սերտ կապի մեջ գտնվելով իրեն նախորդած մերձավոր տեսակի հետ»։ Ներկայումս լավ հայտնի է, որ նա այդ կապը բացատրում է ընդհանուր ծագմամբ, որին ուղեկցում է փոփոխությունը։
Ստեղծագործական մեկ թե մի քանի կենտրոնների գոյության հարցը տարբերվում է մի այլ, թեև մոտ հարցից, այսինքն՝ արդյո՞ք մի տեսակին պատկանող բոլոր անհատներն առաջացել են մեկ զույգից, մեկ հերմաֆրոդիտ անհատից, թե՞, ինչպես ենթադրում են մի քանի հեղինակներ, միաժամանակ ստեղծված շատ անհատներից։ Այնպիսի օրգանիզմների մոտ, որոնք բեղմնավորության կարիք չեն զգում, եթե միայն այդպիսիները գոյություն ունեն, յուրաքանչյուր տեսակ պետք է առաջանար տարատեսակների հաջորդական փոփոխության միջոցով, որոնք միմյանց դուրս են մղել, բայց երբեք չեն խառնվել նույն տեսակի անհատների կամ տարատեսակների հետ. այնպես որ փոփոխության յուրաքանչյուր հաջորդական ստադիայի վրա, այդ ձևի բոլոր անհատները կառաջանային մեկ ծնողից։ Բայց մեծ մասամբ, հատկապես այն, դեպքերում, երբ բեղմնավորումը նախորդում է յուրաքանչյուր ծննդին կամ երբ օրգանիզմները պատահաբար խաչաձևվում են միմյանց հետ, մի մարզում ապրող մի տեսակի անհատները պահպանում են իրենց միակերպությունը խաչաձևման շնորհիվ, այնպես որ նրանցից շատերը պետք է ենթարկվեն միաժամանակյա փոփոխության, և փոփոխությունների ամբողջ գումարը յուրաքանչյուր ստադիայի վրա չպետք է որոշվի միայն մի ծնողից առաջացման միջոցով։ Պարզաբանեմ իմ միտքը օրինակով. մեր անգլիական արշավաձին տարբերվում է ձիերի մյուս բոլոր ցեղերից, բայց նա իր բնորոշ առանձնահատկություններով և գերազանցությամբ պարտական է ոչ թե մի զույգից ծագում առնելուն, այլ ամեն մի սերնդի բազմաթիվ անհատների մշտական ընտրությանը և մարզումին։
Նախքան այն երեք կարգի փաստերի քննարկումը, որոնք ես ընտրել եմ որպես «ստեղծագործության միասնական կենտրոններ» տեսության համար ամենամեծ դժվարություն ներկայացնողներ, ես պետք է մի քանի խոսք ասեմ տարաբնակման եղանակների մասին։
ՏԱՐԱԲՆԱԿՄԱՆ ԵՂԱՆԱԿՆԵՐԸ
Սըր Չ. Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] և ուրիշ հեղինակներ արդեն հմտորեն պարզաբանել են այդ հարցը։ Ես իմ կողմից կարող եմ այստեղ ներկայացնել միայն շատ համառոտ քաղվածք ամենակարևոր փաստերից։ Կլիմայական փոփոխումները պետք է հզոր ազդեցություն ունենային տեղափոխության վրա։ Այն մարզը, որն իր կլիմայի պատճառով ներկայումս անմատչելի է որոշ օրգանիզմների համար, կարող էր լայն ճանապարհ ծառայել տեղափոխությունների համար, երբ նրա կլիման այլ է եղել։ Սակայն շուտով ես առիթ կունենամ այդ հարցի վրա ավելի մանրամասնորեն կանգ առնելու։ Ցամաքի մակարդակի փոփոխությունները նույնպես կարող էին մեծ ազդեցություն ունենալ։ Օրինակ մի նեղ պարանոց ներկայումս բաժանում է երկու ծովային կենդանական աշխարհներ. բայց եթե նա խորասուզվի կամ առաջներում խորասուզվել է ծովի մակարդակի տակ, այդ դեպքում երկու կենդանական աշխարհները կարող են միանալ, դառնալ մեկ կամ կարող էին առաջներում միանալ։ Այնտեղ, որտեղ ներկայումս տարածվում է ծովը, նախկին ժամանակներում ցամաքը կարող էր միացնել կղզիները և նույնիսկ մայր ցամաքները, որը կարող էր առաջ բերել ցամաքային բնակիչների գաղթ մեկից մյուսը։ Երկրաբաններից ոչ ոք չի առարկում, որ ցամաքի մակարդակի մեծ փոփոխություններն են տեղի ունեցել այն ժամանակի ընթացքում, ինչ գոյություն ունեն օրգանիզմները։ Էդվարդ Ֆորբսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] պնդել է, որ ատլանտյան բոլոր կղզիները ոչ շատ առաջ միացած են եղել Եվրոպայի կամ Աֆրիկայի հետ, իսկ Եվրոպան իր հերթին՝ Ամերիկայի հետ։ Այլ հեղինակներ այդ հիպոթեզը փոխադրել են համարյա բոլոր օվկիանոսների վրա և համարյա յուրաքանչյուր կղզի միացրել են այս կամ այն մայր ցամաքի հետ։ Եվ իրոք, եթե Ֆորբսի բերած ապացույցները ճշմարտացի են, ապա մենք պետք է ընդունենք, որ հազիվ թե կա գոնե մի կղզի, որը ոչ այնքան վաղուց միացած չի եղել որևէ մայր ցամաքի հետ։ Այդ տեսակետը կտրում է մեկը մյուսից խիստ հեռավոր մարզերում, միևնույն տեսակի տարածման գորդյան հանգույցը և վերացնում է շատ դժվարություններ, բայց որքան ես կարող եմ դատել, մենք իրավունք չունենք ենթադրելու, որ այդպիսի հսկայական աշխարհագրական փոփոխություններ են կատարվել այժմյան տեսակների գոյության ժամանակի ընթացքում։ Ինձ թվում է, թե մենք բազմաթիվ ապացույցներ ունենք, որ ցամաքի և ծովի մակարդակը մեծ տատանումներ է կրել, բայց մենք որոշակի ցուցմունքներ չունենք մայր ցամաքների դիրքի և չափերի այնպիսի զգալի փոփոխությունների մասին, որպեսզի միացնենք նրանց նորագույն ժամանակաշրջանի ընթացքում ինչպես մեկը մյուսի հետ, այնպես էլ զանազան նրանց միջև ընկած օվկիանոսային կղզիների հետ։ Ես հաճությամբ ընդունում եմ, որ առաջները եղել են կղզիներ, որոնք այժմ սուզվել են ծովի մեջ, որոնք կարող էին մեկնակետեր ծառայել բույսերի և շատ կենդանիների հետագա տարաբնակման համար։ Կորալյան կղզիներ ունեցող ծովերում այդպիսի սուզված կղզիները ներկայումս նկատվում են ծովի մակերևույթից վեր բարձրացող կորալյան կառուցումների օղակներով, կամ ատոլլներով։ Երբ լիակատար կերպով կընդունվի, իսկ այդ երբևիցե կլինի, որ յուրաքանչյուր տեսակ առաջացել է մի հիմնական մարզից և երբ ժամանակի ընթացքում մենք կիմանանք որևէ որոշակի բան տարածման եղանակների մասին, մենք կկարողանանք մեծ համոզվածությամբ կշռադատել ցամաքի նախկին տարածման մասին։ Բայց ես չեմ կարծում, որ երբևիցե կապացուցեն, որ այժմ միմյանցից բոլորովին անջատված մայր ցամաքները մեծ մասամբ անընդմեջ կամ համարյա անընդմեջ կերպով միացած են եղել մեկը մյուսի և այժմյան օվկիանոսային շատ կղզիների հետ։ Օրգանիզմների տարածման վերաբերյալ մի քանի փաստերը, այն է՝ համարյա յուրաքանչյուր մայր ցամաքի հակադիր կողմերում ծովային կենդանական աշխարհների կազմում եղած մեծ տարբերությունը, զանազան երկրների և նույնիսկ ծովերի երրորդային բնակիչների սերտ կապը նրանց ժամանակակից բնակիչների հետ, կղզիների և նրանց ամենամերձավոր մայր ցամաքի կաթնասունների միջև ազգակցությունը, որը մասամբ որոշվում է նրանց միջև ընկած օվկիանոսի խորությամբ (դրա հետ մենք կծանոթանանք հետագայում), այդ և այլ նման փաստերը խոսում են ընդդեմ նորագույն շրջանի ընթացքում այդպիսի հսկայական աշխարհագրական հեղաշրջումներ ընդունելուն, որոնք անհրաժեշտ են Ֆորբսի տեսակետից և ընդունվում են նրա հետևորդների կողմից։ Օվկիանոսային կղզիների բնակիչների առանձնահատկությունները և հարաբերական քանակը նույնպես վկայում են, որ կղզիները մայր ցամաքների հետ առաջներում անընդհատ միացած չեն եղել։ Այդ կղզիների համարյա բացառապես հրաբխային կազմությունը նույնպես չի վկայում այն ենթադրության օգտին, որ նրանք անհետացած մայր ցամաքների մնացորդներ են, որովհետև եթե նրանք լինեին սկզբնապես չընդհատվող լեռնաշարքեր, ապա առնվազն նրանցից մի քանիսը այլ լեռնագագաթների նման կազմված կլինեին գրանիտից, մետամորֆիկ ապարներից, բրածոներ պարունակող հին շերտերից և այլ լեռնային ապարներից, այլ ոչ թե կներկայացնեին բացառապես հրաբխային ծագում ունեցող պարզ կուտակումներ։
Այժմ ես պետք է մի քանի խոսք ասեմ այն մասին, որ անվանում են տարաբնակման պատահական եղանակներ, բայց որն ավելի ճիշտ կլիներ անվանել տարաբնակում բարենպաստ դեպքում, ըստ որում ես կսահմանափակվեմ բույսերով։ Բուսաբանական աշխատություններում հաճախ պնդում են, որ այս կամ այն բույսը վատ է հարմարված իր սերմերը մեծ տարածությունների վրա տարածելու համար, բայց ծովի վրայով բույսերի տեղափոխվելու առավել կամ պակաս ընդունակությունը, կարելի է ասել, գրեթե բոլորովին անհայտ է։ Քանի դեռ ես մ-ր Բերկլեի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 228] օգնությամբ չէի կատարել մի քանի փորձեր, անհայտ էր մնացել նույնիսկ այն, թե սերմերն ինչքան երկար ժամանակ կարող են դիմանալ ծովի ջրի վնասակար ներգործությանը։ Ի զարմանս իմ, ես գտա, որ 87 սերմից 64-ը կարողացան ծլել 28 օր ծովի ջրում նրանց պահելուց հետո, իսկ մի քանիսը դիմացան նույնիսկ 137 օր տևող թրջոցին։ Պետք է նշել, որ մի քանի կարգեր տուժեցին ավելի, քան մյուսները. փորձի համար վերցրած ինը թիթեռնածաղկավոր բույսերից, բացառությամբ մեկի, վատ դիմացան ծովի ջրի ներգործությանը, Hydrophyllaceae և Polemoinaceae մերձավոր ընտանիքներից վերցրած յոթ տեսակները փչացան մի ամիս թրջոցից հետո։ Հարմարության համար ես փորձի ենթարկում էի գլխավորապես մանր սերմեր առանց տուփիկների կամ պտուղների, և որովհետև մի քանի օր հետո նրանք բոլորն էլ սուզվեցին, ուրեմն նրանց չէին կարողանա լողալով անցնել ծովի մեծ տարածություն, անկախ նրանց վրա աղի ջրի գործած վնասակար ազդեցությունից։ Դրանից հետո ես վերցրի մի քանի խոշոր պտուղներ, տուփիկներ և այլն, և նրանցից մի քանիսը երկար ժամանակ մնում էին ջրի երեսին։ Լավ հայտնի է, թե ինչքան մեծ է թարմ և չոր փայտի լողունության տարբերությունը, և իմ մեջ միտք ծագեց, որ ջրերը հաճախ կարող են ծովը տանել չոր բույսեր կամ նրանց ճյուղերը՝ նրանց վրա մնացած սերմերով ու պտուղներով միասին։ Դրա համար ես վճռեցի չորացնել 94 բույսերից վերցրած ցողուններ ու ճյուղեր հասունացած պտուղներով միասին և դրանք դրեցի ծովի ջրի մեջ։ Նրանց մեծամասնությունը շուտով սուզվեց, բայց մի քանիսը, որ կանաչ վիճակում լողում էին միայն կարճատև ժամանակ, չոր վիճակում լողում էին անհամեմատ ավելի երկար ժամանակ։ Այսպես, օրինակ, հասած անտառային կաղինն իսկույն սուզվեց, իսկ չորացրածը լողում էր 90 օր շարունակ և դրանից հետո հողի մեջ դրվելով՝ ծլեց. հասունացած հատապտուղներ ունեցող ծնեբեկի բույսը լողաց 23 օր, չորացրածը՝ 85 օր, որից հետո նրա սերմերն, այնուամենայնիվ, ծլեցին. հովանացավոր Helosciadium-ի հասունացած սերմերը սուզվեցին երկու օրից հետո, իսկ չորացրածները լողացին 90 օրից ավելի և, դրանից հետո ծլեցին։ Ընդհանրապես 94 չորացած բույսերից 18-ը լողացին 28 օրից ավելի, և այդ 18-ից մի քանիսը նույնիսկ շատ ավելի երկար։ Քանի որ վերցրած սերմերի  -ը ծլեցին 20 օր ջրի մեջ մնալուց հետո և հասունացած սերմեր ունեցող զանազան տեսակի բույսերի
-ը ծլեցին 20 օր ջրի մեջ մնալուց հետո և հասունացած սերմեր ունեցող զանազան տեսակի բույսերի  մասը (բայց ոչ բոլոր նույն տեսակներից, ինչպես նախորդ փորձում) լողացին նախապես չորացնելուց հետո 28 օրից ավելի, ուստի կարող ենք եզրակացնել (ընդհանրապես որքան կարելի է եզրակացություններ անել այդ աղքատիկ տվյալների հիման վրա), որ որևէ երկրի բույսերի տեսակների
մասը (բայց ոչ բոլոր նույն տեսակներից, ինչպես նախորդ փորձում) լողացին նախապես չորացնելուց հետո 28 օրից ավելի, ուստի կարող ենք եզրակացնել (ընդհանրապես որքան կարելի է եզրակացություններ անել այդ աղքատիկ տվյալների հիման վրա), որ որևէ երկրի բույսերի տեսակների 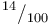 -ի սերմերը կարող են ծովային հոսանքներով տարվել 28 օր շարունակ, պահպանելով իրենց ծլունակությունը։ Ջոնստոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 229] ֆիզիկական ատլասում Ատլանտյան օվկիանոսի զանազան հոսանքների միջին արագությունը ցույց է տրված օրական 33 մղոն (որոշ հոսանքները օրական անում են միջին հաշվով 60 մղոն)․ ելնելով սրանից, մենք ունենք, որ մի երկրի պատկանող բույսերի մասի սերմերը կարող են ծովով լողալ 924 մղոն մինչև մի այլ երկիր և եթե հասնելով ափին, քամու միջոցով կքշվեն մի որևէ բարենպաստ տեղ, այդ դեպքում կծլեն։
-ի սերմերը կարող են ծովային հոսանքներով տարվել 28 օր շարունակ, պահպանելով իրենց ծլունակությունը։ Ջոնստոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 229] ֆիզիկական ատլասում Ատլանտյան օվկիանոսի զանազան հոսանքների միջին արագությունը ցույց է տրված օրական 33 մղոն (որոշ հոսանքները օրական անում են միջին հաշվով 60 մղոն)․ ելնելով սրանից, մենք ունենք, որ մի երկրի պատկանող բույսերի մասի սերմերը կարող են ծովով լողալ 924 մղոն մինչև մի այլ երկիր և եթե հասնելով ափին, քամու միջոցով կքշվեն մի որևէ բարենպաստ տեղ, այդ դեպքում կծլեն։
Ինձանից հետո նման փորձեր կատարել է Մ. Մարտենսոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 230] բայց այդ փորձերը շատ ավելի լավ են դրվել, որովհետև նա սերմերն արկղի մեջ իջեցրել է ուղղակի ծովը, որից նրանք հետզհետե թրջվել են ու ենթարկվել օդի ներգործության, ինչպես իսկական ծովում լողացող բույսեր։ Փորձի համար նա վերցրել է 98 տեսակի սերմեր, մեծ մասամբ իմ վերցրածներից տարբեր, բայց դրա հետ միասին վերցրել է մի քանի խոշոր պտուղներ, ինչպես նաև ծովի շուրջը ապրող բույսերի սերմեր, որը նպաստել է ինչպես նրանց լողունության տևողությանը, այնպես էլ նրանց դիմադրությանը աղի ջրի վնասակար ներգործության դեմ։ Սակայն դրա փոխարեն նա նախապես չի չորացրել ոչ բույսերը, ոչ պտուղներով ճյուղերը, որը, ինչպես մենք տեսանք, նրանցից մի քանիսի համար կարող է զգալիորեն երկարացնել լողունությունը։ Դրա հետևանքով փորձի համար վերցրած զանազան տեսակների սերմերի 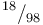 մասը մնացել է ջրի մեջ 42 օր և դրանից հետո պահպանել է ծլունակությունը։ Բայց ես չեմ կասկածում, որ այդ բույսերը, ենթարկվելով ալիքների ազդեցությանը, ավելի քիչ կլողային, քան ուժեղ շարժումներից պաշտպանված լինելով, ինչպես եղել է մեր փորձերում։ Այդ պատճառով գուցե ավելի վստահելի լինի ընդունել, որ որևէ բուսական տեսակների սերմերի
մասը մնացել է ջրի մեջ 42 օր և դրանից հետո պահպանել է ծլունակությունը։ Բայց ես չեմ կասկածում, որ այդ բույսերը, ենթարկվելով ալիքների ազդեցությանը, ավելի քիչ կլողային, քան ուժեղ շարժումներից պաշտպանված լինելով, ինչպես եղել է մեր փորձերում։ Այդ պատճառով գուցե ավելի վստահելի լինի ընդունել, որ որևէ բուսական տեսակների սերմերի 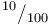 մասը չորացնելուց հետո կարող էր լողալով անցնել 900 մղոն լայնություն ունեցող ծովը և հետո ծլել։ Հետաքրքրական է այն հանգամանքը, որ խոշոր պտուղները հաճախ մանրերից ավելի երկար են լողում, որովհետև խոշոր պտուղներ կամ սերմեր ունեցող բույսերը, որոնք, ինչպես այդ ցույց է տվել Ալֆոնս դը-Կանդոլը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] ունեն տարածման նեղ մարզեր, հազիվ թե կարող են մի տեղից մի այլ տեղ տեղափոխվել որևէ այլ եղանակով։
մասը չորացնելուց հետո կարող էր լողալով անցնել 900 մղոն լայնություն ունեցող ծովը և հետո ծլել։ Հետաքրքրական է այն հանգամանքը, որ խոշոր պտուղները հաճախ մանրերից ավելի երկար են լողում, որովհետև խոշոր պտուղներ կամ սերմեր ունեցող բույսերը, որոնք, ինչպես այդ ցույց է տվել Ալֆոնս դը-Կանդոլը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] ունեն տարածման նեղ մարզեր, հազիվ թե կարող են մի տեղից մի այլ տեղ տեղափոխվել որևէ այլ եղանակով։
Սերմերը կարող են տարածվել նաև մի այլ եղանակով։ Լողացող ծառերը տարվում են կղզիների մեծամասնության վրա, և նույնիսկ նրանցից այնպիսիները, որոնք գտնվում են բաց օվկիանոսի մեջտեղում։ Խաղաղ օվկիանոսի կորալյան կղզիների բնակիչները իրենց գործիքների համար քարեր են գտնում բացառապես լողացող ծառերի արմատների մեջ. այդ քարերը պատկանում են թագավորներին և տալիս են նրանց մեծ եկամուտ։ Ես գտել եմ, որ եթե անկանոն գծագիր ունեցող քարը խրվել է ծառի արմատների միջև, ապա հողի մասնիկները հաճախ այնպես պինդ մտած են լինում նրա անհարթությունների մեջ, որ ջուրը չի կարողանում լվանալ նրանց այնտեղից նույնիսկ երկար ճանապարհի ընթացքում։ Եվ ես միանգամայն համոզված եմ մի դիտողության ճշտությանը, որ հողի մի փոքրիկ քանակությունից, որը բոլորովին թաքնված է եղել մոտավորապես հիսուն տարեկան կաղնու արմատներում, ծլել են երեք հատ երկշաքիլ բույսեր։ Բացի դրանից, ես կարող եմ հաղորդել, որ թռչունների դիակներն ընկնելով ծովը, երբեմն անմիջապես ոչնչացման չեն ենթարկվում, իսկ այդպիսի լողացող դիակների կտնառքներում զանազան սերմեր երկար ժամանակ պահպանում են իրենց կենսունակությունը. ոլոռը և վիկան, օրինակ, մնալով ծովի ջրի մեջ ընդամենը մի քանի օր, փչանում են, բայց 30 օր շարունակ ծովի ջրում լողալու թողնված աղավնու դիակի կտնառքից հանվելուց հետո, ի զարմանս իմ, նրանք գրեթե բոլորը ծլեցին։
Կենդանի թռչունները նույնպես սերմերի տարածման համար գործունյա գործոններ են։ Ես կարող էի բերել բազմաթիվ օրինակներ, ապացուցելու համար, թե ինչպես հաճախ թռչունների զանազան տեսակներ քամու միջոցով քշվում են օվկիանոսի ընդարձակ տարածությունների վրայով։ Ընդ որում մենք կարող ենք առանց տատանվելու ընդունել, որ նրանց թռիչքի միջին արագությունը այդպիսի պայմաններում հաճախ լինում է մեկ ժամում 35 մղոն [մոտավորապես 56 կմ], իսկ մի քանի հեղինակներ տալիս են է՛լ ավելի բարձր թվեր։ Ես ոչ մի անգամ չեմ տեսել, որ սննդարար սերմերն ամբողջական անցնեն թռչունի աղիքների միջով, բայց պտուղների պինդ սերմերն անվնաս անցնում են նույնիսկ հնդուհավի մարսողության օրգաններով։ Ես իմ այգում երկու ամսվա ընթացքում մանր թռչնակների կղկղանքներից հանել եմ 12 տեսակների սերմեր, որոնք անվնաս էին թվում, և նրանցից մի քանիսը փորձարկելիս ծլեցին։ Բայց հետևյալ փաստը առանձին նշանակություն ունի, թռչունների կտնառքը ստամոքսահյութ չի արտադրում և, ինչպես ես փորձերից գիտեմ, ոչ մի վնասակար ազդեցություն չի թողնում բույսերի ծլելու ընդունակության վրա. եթե թռչունը գտել է ու կուլ տվել մեծ քանակությամբ կեր, ապա, ինչպես հաստատապես հայտնի է, բոլոր հատիկները տասներկու և նույնիսկ տասնևութ ժամից շուտ չեն ընկնի ստամոքսը։ Այդ ժամանակամիջոցում թռչունը կարող է հեշտությամբ տարվել 500 մղոն հեռու․ ինչպես մենք գիտենք, գիշատիչները բռնում են հոգնած թռչուններ, և վերջիններիս պատառոտված կտնառքի պարունակությունը այդ ժամանակ հեշտությամբ կարող է դուրս թափվել։ Մի քանի ցերեկային գիշատիչ թռչուններ և բվերը իրենց որսն ուտում են ամբողջությամբ, իսկ տասներկուսից մինչև քսան ժամ հետո թափում են աղտեղությունները, որոնք, ինչպես ես գիտեմ կենդանական այգիներում կատարած փորձով, պարունակում են ծլելու ընդունակ սերմեր։ Որոշ դեպքերում վարսակը, ցորենը, կորեկը, կանեփը, դեղձանիկի կուտը, երեքնուկը և ճակնդեղը ծլել են, մնալով զանազան գիշատիչ թռչունների ստամոքսում տասներկուսից մինչև քսան և մեկ ժամ, ճակնդեղի երկու սերմիկներ ծիլ տվել են, թռչունի ստամոքսում երկու օր ու տասնչորս ժամ մնալուց հետո։ Քաղցրահամ ջրերի ձկները, իմ դիտումների համաձայն, ուտում են ցամաքային և ջրային բույսերի սերմեր և դրա շնորհիվ նույնպես սերմերը կարող են տեղից տեղ փոխադրվել։ Ես զանազան սերմեր խոթել եմ մեռած ձկների ստամոքսները և դրանցից հետո նրանց գցել եմ ձկնկեր գիշատիչներին, արագիլնեըին և պելիկաններին, մի քանի ժամից հետո այդ թռչունները սերմերը դուրս են շպրտել կամ բերանից աղտեղությունների հետ, կամ կղկղանքների հետ, ընդ որում սերմերից մի քանիսը պահպանել են ծլելու ընդունակությունը։ Սակայն այլ սերմեր այդ դեպքում միշտ մեռած էին լինում։
Մորեխը քամու միջոցով երբեմն տարվում է երկրից մեծ հեռավորություններ, ես ինքս բռնել եմ մեկը Աֆրիկայի ափից 370 մղոն հեռու և լսել եմ է՛լ ավելի մեծ հեռավորության վրա բռնվածների մասին։ Հարգարժան Ռ. Թ. Լոուն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 231] հաղորդել է սըր Չ. Լյաելին,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] որ 1844 թ. նոյեմբերին մորեխի անթիվ զանգվածներ այցելել են Մադեյրա կղզին, նրանք այնպիսի խտությամբ են թռել, ինչպես ձյունի փաթիլները ամենաուժեղ բուքի ժամանակ, և դեպի վերև նրանք տեսանելի են եղել այնքան բարձրությամբ, որ նրանց կարելի էր տեսնել հեռադիտակով։ Երկու կամ երեք օր շարունակ նրանք դանդաղ կերպով պտտվել են հսկայական ձվածիրով, առնվազն 5—6 մղոն տրամագծով, իսկ գիշերը իջել են բարձր ծառերի վրա, բոլորովին ծածկելով նրանց։ Մորեխը դեպի ծովը թռչելով անհետացել է նույնքան անսպասելի կերպով, ինչպես որ երևացել է, և այդ ժամանակից չի այցելել կղզին։ Նատալ երկրի որոշ մասերում ֆերմերները կարծում են, թեև առանց բավականաչափ հիմք ունենալու, որ վնասակար սերմերն իրենց մարգագետինների վրա բերվում են այն կղկղանքների հետ, որոնք մնում են այդ երկիրը շատ հաճախ այցելող մորեխների մեծ երամներից հետո։ Այդ ենթադրության հիման վրա մ-ր Ուիլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 232] նամակի մեջ ինձ ուղարկեց մորեխների արտաթորությունների չոր կտորների մի փոքրիկ ծրար, ես այդտեղից խոշորացույցի օգնությամբ հանեցի մի քանի սերմեր և աճեցրի նրանցից յոթ խոտաբույսեր, որոնք պատկանում էին երկու սեռերի երկու տեսակների։ Հետևաբար, մորեխի երամները, Մադեյրան այցելողների նման, կարող են իրոք որպես միջոց ծառայել բույսերի զանազան տեսակներ ցամաքից հեռու գտնվող կղզիների վրա տարածելու համար։
Թեև թռչունների կտուցներն ու ոտները սովորաբար մաքուր են լինում, բայց երբեմն հողը կպչում է նրանց, կաքավի ոտներից ես մի դեպքում պոկել եմ 61, մի ուրիշ անգամ՝ 22 գրան չոր կավային հող, և այդ հողի մեջ կային վիկի սերմի մեծությամբ քարեր։ Կամ էլ ավելի ուսանելի մի դեպք՝ իմ ընկերներից մեկն ուղարկել էր ինձ մի անտառային կտցար, որի սրունքին կպած էր հողի մի չոր կտոր ինը գրան կշռով, բայց նրա մեջ կար ճլիլի (Iunus bufonius) մի սերմիկ, որը ծլեց և ծաղիկներ տվեց։ Բրայտոնցի մ-ր Սվեյսլանդը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 233] որը վերջին քառասուն տարվա ընթացքում ուշադիր կերպով ուսումնասիրել է մեր չվող թռչուններին, հաղորդում է ինձ, որ ինքը հաճախ սպանել է խաղատտիկներ (Motacillae), մարգագետնային և քարային թռչուններ (Saxicolae) նրանց մեր ծովափ գալուց անմիջապես հետո, բայց նախքան գետնի վրա իջնելը, և այդ ժամանակ նա մի քանի անգամ նկատել է նրանց ոտներին կպած հողի կոշտիկներ։ Այդ հաստատելու համար, որ հողը սովորաբար սերմեր է պարունակում, կարելի է շատ օրինակներ բերել։ Այսպես, պրոֆ. Նյուտոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 234] ուղարկել է ինձ մի քարակաքավի (Caccabis rufa) ոտ, որը վիրավորված լինելով, չէր կարողացել թռչել, այդ ոտի վրա կար չոր հողակոշտ 6½ ունցիա կշռով։ Այդ հողը պահվել է երեք տարի շարունակ, բայց երբ հետո մանրացվել, թրջվել է ու զետեղվել ապակյա զանգի տակ, նրանից դուրս են եկել 82-ից ոչ պակաս բույսեր. դրանցից 12-ը միաշաքիլավորներ էին, այդ թվում մի սովորական վարսակ և առնվազն էլի հացազգի մի տեսակ, և 70 հատ երկշաքիլավորներ, որոնք պատկանում էին, որքան ես կարող էի դատել ըստ նրանց ծիլերի, ամենաքիչը երեք տարբեր տեսակների։ Մեր առջև ունենալով այսպիսի փաստեր, մենք արդյոք կարո՞ղ ենք կասկածել, որ շատ թռչուններ, որոնք ամեն տարի քամու միջոցով քշվում են օվկիանոսի մեծ տարածությունների վրայով և ամեն տարի չվերթ են կատարում, ինչպես օրինակ, միլիոնավոր լորերը, որոնք չվում են Միջերկրական ծովի վրայով, կարող են պատահաբար տեղափոխել իրենց ոտներին կամ կտուցին կպած չոր հողի հետ նաև մի քանի սերմ։ Բայց ես կրկին կվերադառնամ այդ խնդրին։
Մեզ հայտնի է նաև այն, որ լողացող սառցակույտերն իրենց հետ բերում են հող և քարեր, իսկ որոշ դեպքերում էլ ջախ, ոսկորներ և ցամաքային թռչունների բներ, այնպես որ հազիվ թե կարելի է կասկածել, որ նրանք կարող են պատահական կերպով, ինչպես այդ ենթադրել է Չ. Լյաելը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] սերմերը տեղափոխել արկտիկական կամ անտարկտիկական մարզի մի մասից մյուսը, իսկ սառցադաշտային ժամանակաշրջանում նրանք կարող էին սերմերը տարածել այժմյան բարեխառն գոտու սահմաններում։ Այսպես, ելնելով այն հանգամանքից, որ Ազորյան կղզիների և Եվրոպայի բուսական շատ տեսակներ, որոնք նման են իրար, համեմատած Ատլանտյան օվկիանոսի այն կղզիների տեսակների հետ, որոնք ավելի մոտ են գտնվում մայր ցամաքին, և ինկատի առնելով (ինչպես այդ նկատել է մ-ր Հ. Կ. Ուոտսոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 62]) այդ բույսերի փոքր ինչ հյուսիսային բնույթը աշխարհագրական լայնության համեմատությամբ, ես կասկածում եմ, որ Ազորյան կղզիները սերմեր ստացել են սառցադաշտային ժամանակաշրջանում լողացող սառցակույտերի մասնակցությամբ։ Իմ խնդրանքով սըր Չ. Լյաելը գրել է Մ. Հարտունգին,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 235] հարցնելով, թե արդյոք նա չի՞ գտել այդ կղզիների վրա գլաքարային գոյացումներ, և պատասխան է ստացել, որ նա գտել է գրանիտի և այլ այնպիսի ապարների մեծ կտորներ, որոնք արշիպելագի վրա չեն հանդիպում։ Այդ պատճառով մենք կարող ենք համոզված կերպով ենթադրել, որ լողացող սառույցները ժայռերի բեկորներից բաղկացած իրենց բեռը մի ժամանակ թափել են օվկիանոսային այդ կղզիների ափերի վրա և համենայն դեպս հնարավոր է, որ նրանք կարող էին այստեղ բերել հյուսիսային բույսերի որոշ սերմեր։
Ուշադրության առնելով այն, որ սերմերի տարածման վերոհիշյալ եղանակները, ինչպես նաև այլ եղանակներ, որոնք անկասկած դեռ մնում են չհայտնագործված, տեղի են ունեցել տարեցտարի՝ տասնյակ հազարամյակների ընթացքում, ես կարծում եմ, տարօրինակ կլիներ, եթե շատ բույսեր չկարողանային լայն կերպով տարածվել։ Այդ եղանակները երբեմն անվանվում են պատահական, բայց այդպիսի արտահայտությունը այնքան էլ ճիշտ չէ, որովհետև ծովային հոսանքները պատահական չեն, ինչպես որ պատահական չէ նաև իշխող քամիների ուղղությունը։ Պետք է նշել, որ հազիվ թե լինի մեծ տարածության վրա սերմեր տեղափոխելու որևէ եղանակ, որը կարող է նրանց պահել լավ վիճակում, որովհետև մի կողմից սերմերը չեն կարող պահպանել իրենց ծլունակությունը ծովի ջրի երկարատև ազդեցության տակ, մյուս կողմից նրանք չեն կարող պահպանվել թռչունների կտնառքի և աղիքների մեջ։ Բայց և այնպես այդ եղանակները բավարար են, որպեսզի սերմերը պատահականորեն տեղափոխվեն մի քանի հարյուր մղոն լայնությամբ ծովի վրայով, ինչպես նաև տեղափոխվեն մի կղզուց մյուս կղզու վրա կամ մի մայր ցամաքից մոտիկ գտնվող կղզիների վրա, թեև անբավարար են մի մայր ցամաքից հեռու գտնվող մի այլ մայր ցամաքի վրա տեղափոխելու համար։ Միմյանցից հեռու գտնվող մայր ցամաքների բուսական աշխարհները չեն կարող խառնվել այդպիսի պատճառների ազդեցության տակ և մնում են այնքան տարբեր, որքան մենք այդ տեսնում ենք ներկայում։ Ծովային հոսանքներն իրենց ուղղության շնորհիվ երբեք չեն բերում սերմեր Հյուսիսային Ամերիկայից Բրիտանական կղզիները, բայց նրանք կարող են սերմեր բերել և իրոք բերում են Արևելյան Հնդկաստանից դեպի մեր արևմտյան ծովափերը, որտեղ նրանք, եթե չեն էլ փչանում ծովի ջրում չափազանց երկարատև մնալուց, այնուամենպյնիվ փչանալու են, որովհետև չեն դիմանում մեր կլիմային։ Գրեթե ամեն տարի մեկ կամ երկու ցամաքային թռչուն Հյուսիսային Ամերիկայից ամբողջ Ատլանտյան օվկիանոսի վրայով անցնում են Իռլանդիայի կամ Անգլիայի արևմտյան ափերը. բայց սերմերն այդ հազվագյուտ թափառաշրջիկների միջոցով միայն մի դեպքում կարող են տեղափոխվել, այն է՝ պահպանվելով նրանց ոտներին և կտուցին կպած հողի մեջ, որն ինքնըստինքյան քիչ է պատահում։ Սակայն նույնիսկ այդ դեպքում էլ շատ քիչ հավանականություն կա, որ սերմը կընկնի բարենպաստ հողի մեջ և նրանից դուրս եկած բույսը կհասնի լրիվ զարգացման։ Այնուամենայնիվ մեծ սխալ կլիներ կարծել, որ եթե Մեծ Բրիտանիայի նման հարուստ բնությամբ մի կղզի, որքան հայտնի է (թեև այդ շատ դժվար կլիներ ապացուցել), վերջին մի քանի դարերի ընթացքում թափառաշրջիկների տեղափոխության պատահական այդ եղանակներով սերմեր չի ստացել Եվրոպայից կամ մի այլ մայր ցամաքից, ապա իր բնությամբ աղքատ և մայր ցամաքից ավելի հեռավոր մի այլ կղզի չէր կարող գաղթականներ ստանալ այդ նույն եղանակներով։ Բրիտանիայից նույնիսկ անհամեմատ ավելի քիչ հարուստ մի կղզու վրա ընկած սերմերի կամ կենդանիների հարյուրավոր տեսակներից թերևս մեկը միայն կլիներ այնքան լավ հարմարված իր նոր բնակավայրին, որպեսզի ենթարկվեր նատուրալիզացիայի։ Բայց դա ամենևին չի կարող լուրջ առարկություն լինել տեղափոխության պատահական եղանակների նշանակության նկատմամբ, որոնք գործել են երկրաբանական երկարատև ժամանակաշրջանների ընթացքում, քանի դեռ կղզին բարձրանում էր և մինչև որ նա լիովին լցվել է բնակիչներով։ Գրեթե մերկ հողի վրա, որը բնակված է միայն փոքրաթիվ միջատներով և թռչուններով, կամ նույնիսկ բոլորովին զուրկ է դրանցից, համարյա յուրաքանչյուր սերմ հնարավորություն կունենար ծլելու և ապրելու, եթե միայն հարմարվել է կլիմային։
ՏԱՐԱԲՆԱԿՈՒՄԸ ՍԱՌՑԱԴԱՇՏԱՅԻՆ ԺԱՄԱՆԱԿԱՇՐՋԱՆԻ ԸՆԹԱՑՔՈՒՄ
Մի վայրից մյուսը տեղափոխության անհնարին թվացող պայմաններում առանձնացած տեղամասերում տեսակի տարածման ամենազարմանալի դեպքերից մեկն է այն նմանությունը, որ ունեն այնպիսի լեռնագագաթների վրա ապրող շատ բույսեր ու կենդանիներ, որոնք միմյանցից անջատված են հարյուրավոր մղոն տարածություն գրավող ցածրավայրերով, որտեղ ալպիական տեսակները չեն կարող գոյություն ունենալ։ Իրոք, ուշագրավ է այն փաստը, որ միևնույն տեսակին պատկանող շատ բույսեր ապրում են Ալպերի և Պիրենյան լեռների ձյունի շրջանում և Եվրոպայի ծայր հյուսիսում, բայց առավել ևս ուշագրավ է այն, որ Ամերիկայի Միացյալ նահանգներում Սպիտակ լեռների բույսերը նույնն են, ինչ որ Լաբրադորում, և համարյա նույն բույսերն են, ինչպես այդ գիտենք Ազա-Գրեյից,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 68] որոնք աճում են Եվրոպայի ամենաբարձր լեռների վրա։ Նման փաստերը դեռ 1747 թվականին Գմելինին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 236] բերել են այն եզրակացության, որ միևնույն տեսակը կարող է ստեղծված լինել անկախ կերպով զանազան կետերում և մենք կմնայինք այդ տեսակետի վրա, եթե Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] և ուրիշներ լուրջ ուշադրություն չդարձնեին սառցադաշտային ժամանակաշրջանի վրա, որը, ինչպես մենք այժմ կտեսնենք, պարզ բացատրություն է տալիս այդ փաստերին։ Մենք ունենք վկայություններ, որոնք ընձեռում են թե՛ օրգանական և թե՛ անօրգանական ամենաբազմազան առարկաները, որ կենտրոնական Եվրոպան և Հյուսիսային Ամերիկան նորագույն երկրաբանական ժամանակաշրջանի ընթացքում կրել են բևեռային կլիմա։ Հրդեհից ոչնչացած տան փլատակները ավելի լրիվ չեն կարող հաղորդել պատահածի մասին, քան Շոտլանդիայի և Ուելսի լեռները պատմում են մեզ իրենց խզբզված լանջերով, հղկված մակերեսներով և կուտակված գլաքարերով սառցի հեղեղների մասին, որոնք ոչ այնքան վաղուց լցրել են լեռնային հովիտները։ Եվրոպայի կլիման այնքան շատ է փոխվել, որ հյուսիսային Իտալիայում նախկին սառցադաշտերի թողած հսկայական քարակարկառները ծածկված են խաղողի այգիներով և եգիպտացորենի դաշտերով։ Էռատիկական գլաքարերը և խզբզված ժայռերը համոզիչ կերպով ապացուցում են ավելի վաղ ցուրտ ժամանակաշրջանի գոյությունը Միացյալ Նահանգների զգալի մասի տարածության վրա։
Սառցադաշտային կլիմայի անցյալ ներգործությունը Եվրոպայի բնակիչների տարածման վրա, Էդվարդ Ֆորբսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] բացատրության համաձայն, ըստ էության հետևյալն է։ Նրա հարուցած փոփոխություններն ավելի լավ հասկանալու համար մենք ենթադրենք, թե մի նոր սառցադաշտային ժամանակաշրջան նորից դանդաղ կերպով գալիս է և անցնում, ինչպես այդ եղել է նաև առաջներում։ Քանի ցուրտն ուժեղացել է, այնքան մեկը մյուսի հետևում գտնվող հարավային գոտիները պիտանի են դարձել հյուսիսի բնակիչների համար և վերջինները գրավել են բարեխառն երկրների նախկին բնակիչների տեղերը։ Միաժամանակ այդ նախկին ձևերը նահանջել են շարունակ ավելի ու ավելի հարավ, մինչև որ նրանց կանգնեցրել են արգելքները,— այդպիսի դեպքում նրանք պետք է ոչնչանային։ Լեռները ծածկվել են ձյունով ու սառույցներով, և նախկին ալպիական բնակիչները պետք է իջնեին հովիտները։ Իսկ այդ ժամանակ, երբ ցուրտը հասել է իր maximum-ին, արկտիկական կենդանական և բուսական աշխարհները գրավել են Եվրոպայի կենտրոնական մասերը, Ալպերից և Պիրենյան լեռներից դեպի հարավ, և նույնիսկ մտել են Իսպանիա։ Միացյալ Նահանգների այժմյան բարեխառն մարզերում նույն ձևով պետք է բնակություն հաստատեին բևեռային կենդանիներն ու բույսերը, որոնք համարյա չեն տարբերվում եվրոպականներից, քանի որ այժմյան մերձբևեռային ձևերը, որոնց վերաբերմամբ մենք ենթադրում ենք, որ նրանք ամեն տեղից գաղթել են դեպի հարավ, զարմանալիորեն միակերպ են ամբողջ բևեռային մարզում։
Տաքության վերադարձի հետ զուգընթաց արկտիկական ձևերը պետք է նահանջեին դեպի հյուսիս, որոնց նահանջի ընթացքում կրնկակոխ հետևում են ավելի բարեխառն երկրների օրգանիզմները։ Մյուս կողմից, քանի որ ձյունը հալվում էր սկսած լեռների ստորոտներից, ուստի արկտիկական ձևերը պետք է գրավեն բացված ու հալչող վայրերը, հետզհետե բարձրանալով ավելի վեր ու վեր՝ ինչ չափով որ ավելի տաքություն էր լինում և ձյունը ավելի շատ էր հալվում, մինչդեռ նրանց եղբայրակիցները շարժվում էին դեպի իրենց հյուսիսային բնակավայրը։ Այդ պատճառով էլ, երբ բոլորովին տաքացել է, մինչև այդ Եվրոպայի և Հյուսիսային Ամերիկայի ցածրավայրերում բնակվող տեսակները պետք է կրկին ընկնեին Հին և Նոր աշխարհների արկտիկական մարզերը և մեկը մյուսից հեռու գտնվող լեռնագագաթների վրա։
Այստեղից մենք կարող ենք հասկանալ, թե ինչու իրար նման են այն բույսերը, որոնք գտնվում են միմյանցից այնքան հեռավոր կետերում, ինչպես Միացյալ նահանգների և Եվրոպայի լեռներն են։ Այստեղից մենք կարող ենք նաև հասկանալ, թե ինչու յուրաքանչյուր լեռնաշղթայի ալպիական բույսերն առանձնապես մոտ են այն արկտիկական ձևերին, որոնք ապրում են նրանցից ուղիղ դեպի հյուսիս կամ գրեթե ուղիղ դեպի հյուսիս, որովհետև ցրտերի վրա հասնելու ժամանակ առաջին գաղթը և տաքերն ընկնելիս նրանց վերադարձը տեղի են ունեցել ընդհանրապես կամ ուղիղ դեպի հարավ կաս ուղիղ դեպի հյուսիս։ Օրինակ, Շոտլանդիայի ալպիական բույսերը, ըստ մ-ր Հ. Կ. Ուոտսոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 62] դիտողության, և Պիրենյան բույսերը, ըստ Ռամոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 237] դիտողության, առանձնապես մոտիկ են հյուսիսային Սկանդինավիայի բույսերին. Միացյալ նահանգների բույսերը՝ լաբրադորական բույսերին. Սիբիրի լեռների բույսերը՝ այդ երկրի արկտիկական մարզի բույսերին։ Այդպիսի բացատրությունը, որը հիմնվում է նախկին սառցադաշտային ժամանակաշրջանի միանգամայն հաստատված փաստի վրա, իմ կարծիքով այնքան բավարար կերպով է բացատրում Եվրոպայի և Ամերիկայի ալպիական և արկտիկական արտադրանքների ժամանակակից տարածումը, որ երբ մենք միևնույն տեսակները գտնում ենք այլ երկրներում իրարից հեռու ընկած լեռնագագաթներ իր վրա, մենք ավելի մեծ կամ փոքր իրավունքով կարող ենք, առանց հետագա ապացույցների, գալ այն եզրակացության, որ մի ավելի ցուրտ կլիմա ղրանից առաջ հնարավորություն է տվել այդ բույսերին տեղափոխվելու լեռների միջև ընկած ցածրավայրերով, որոնք այժմ նրանց համար չափազանց տաք են։
Քանի որ արկտիկական ձևերը, կլիմայի փոփոխությունների կապակցությամբ, սկզբում շարժվել են դեպի հարավ, իսկ հետո ետ՝ դեպի հյուսիս, ուստի նրանք իրենց այդ երկարատև թափառումների ընթացքում ջերմության առանձին տարբերություն չեն կրել, իսկ քանի որ նրանք գաղթել են մասսայորեն, ապա նրանց փոխհարաբերություններն էլ զգալի չափով չեն խանգարվել։ Այդ հիման վրա, այս գրքում անցկացվող սկզբունքների համաձայն, այդ ձևերը մեծ փոփոխությունների չեն ենթարկվել։ Բայց ինչ վերաբերում է ալպիական ձևերին, որոնք տաքության վերադարձի ժամանակից սկսած մնացել են մեկուսացած սկզբում ստորոտներում, իսկ հետագայում լեռնագագաթների վրա, ապա նրանց դրությունը մի քիչ տարբեր էր, որովհետև անհավատալի է, որ բոլոր արկտիկական ձևերը, որոնք մնացել են միմյանցից հեռու գտնվող լեռնագագաթների վրա, այն ժամանակվանից պահպանվեին այստեղ, ամենայն հավանականությամբ նրանք խառնվել են նաև հին ալպիական տեսակների հետ, որոնք պետք է գոյություն ունենային լեռների վրա մինչև սառցադաշտային դարաշրջանի սկիզբը, բայց ամենացուրտ ժամանակաշրջանի ընթացքում դուրս մղվեին դեպի հարթավայրերը. բացի դրանից, նրանք որոշ չափով ենթարկվել են նաև կլիմայական ազդեցությունների։ Այդ պատճառով նրանց փոխհարաբերությունները կարող էին ավելի կամ պակաս չափով խանգարվել, իսկ դրա հետևանքով նրանք կարող էին փոփոխվել և փոփոխվել են․ այսպես, եթե մենք համեմատենք Եվրոպայի զանազան մեծ լեռնաշղթաների այժմյան ալպիական կենդանիներին ու բույսերին միմյանց հետ, ապա կգտնենք, որ թեև շատ տեսակներ մնացել են բոլորովին անփոփոխ, բայց մի քանիսն առաջացրել են տարատեսակներ, մյուսները գոյություն ունեն որպես կասկածելի ձևեր, կամ ենթատեսակներ, և վերջապես մի քանիսն էլ դարձել են տարբեր, թեպետ և մերձավոր տեսակներ, որոնք զանազան մարզերում փոխարինում են միմյանց։
Նախորդ շարադրանքում ես ընդունեցի, որ մեր երևակայական սառցադաշտային ժամանակաշրջանի սկզբում արկտիկական արտադրանքները բևեռային մարզերում նույնքան միակերպ են եղել, ինչքան որ ներկայումս են. բայց անհրաժեշտ է ընդունել նաև այն, որ մերձարկտիկական ձևերից շատերը և բարեխառն կլիմայի ձևերից քչերը նույնպես ամենուրեք եղել են նույնը, որովհետև Հյուսիսային Ամերիկայի և Եվրոպայի լեռների ցածր լանջերի և հարթավայրերի վրա այժմ ապրող տեսակներից մի քանիսը նույնն են։ Այդ պատճառով կարելի է հարցնել, ինչպե՞ս եմ ես բացատրում ամբողջ աշխարհի մերձարկտիկական և բարեխառն գոտու ձևերի միակերպության այդ աստիճանը իսկական սառցադաշտային ժամանակաշրջանի սկզբում։ Ներկայումս Հին և Նոր աշխարհների մերձարկտիկական և հյուսիսային բարեխառն գոտու արտադրանքները միմյանցից բաժանված են ամբողջ Ատլանտյան օվկիանոսով և Խաղաղ օվկիանոսի հյուսիսային մասով։ Սառցադաշտային ժամանակաշրջանի ընթացքում, երբ Հին և Նոր աշխարհների այդ բնակիչները ապրել են դեռ է՛լ ավելի հարավ, քան այժմ, նրանք պետք է ավելի ևս կատարելապես բաժանված լինեին օվկիանոսի ընդարձակ տարածությամբ, ուստի կարելի է հարցնել, թե ինչպե՞ս կարող էր միևնույն տեսակն այդ ժամանակ կամ ավելի վաղ մուտք գործել երկու մայր ցամաքները։ Ես կարծում եմ, որ դրա բացատրությունը գտնվում է մինչսառցադաշտային ժամանակաշրջանի կլիմայի բնույթի մեջ։ Այդ նորագույն պլիոցենյան ժամանակաշրջանում երկրագնդի բնակիչների մեծամասնությունը եղել է նույնը, ինչ որ ներկայումս է։ Ելնելով սրանից, մենք կարող ենք ենթադրել, որ 60° լայնության տակ այժմ ապրող օրգանիզմները պլիոցենյան ժամանակաշրջանի ընթացքում ապրել են ավելի հյուսիսում, բևեռային շրջանագծի մոտերը, այսինքն՝ լայնության 66—67°-ի տակ, իսկ այժմյան արկտիկական ձևերն այն ժամանակ ապրել են բևեռին ավելի մոտ՝ կտրատված ցամաքի վրա։ Բայց դիտելով երկրագունդը, մենք տեսնում ենք, որ բևեռային շրջանագծի մոտերը ցամաքը ձգվում է գրեթե չընդհատված շերտով արևմտյան Եվրոպայից Սիբիրի վրայով մինչև արևելյան Ամերիկա։ Իսկ մերձբևեռային ցամաքի այդ անընդհատությունը և դրա հետևանքով տեղափոխությունների ազատությունը, ավելի բարենպաստ կլիմայական պայմանների հետ միասին, բավական են, որպեսզի բացատրենք Հին և Նոր աշխարհների մերձարկտիկական և բարեխառն ձևերի թույլատրելի միակերպությունը։
Վերոհիշյալի հիման վրա ընդունելով, որ մեր մայր ցամաքները երկար ժամանակ պահպանել են մոտավորապես միևնույն հարաբերական դիրքը, թեև նրանց մակերևույթը մեծ փոփոխությունների է ենթարկվել, ես որոշակիորեն հակվում եմ ընդարձակելու իմ ավելի առաջ արտահայտած տեսակետը և ընդունելու, որ մի է՛լ ավելի վաղ և էլ ավելի տաք ժամանակաշրջանի ընթացքում, ինչպիսին, օրինակ, եղել է վաղ պլիոցենը, միևնույն բույսերի և կենդանիների մեծ քանակություն է բնակվել գրեթե չընդհատված շուրջբևեռային ցամաքի վրա և որ այդ բույսերն ու կենդանիները թե Հին և թե Նոր աշխարհում սկսել են աստիճանաբար շարժվել դեպի հարավի կլիմայի ցրտելու հետ զուգընթաց, դեռ սառցադաշտային ժամանակաշրջանից շատ առաջ։ Ես կարծում եմ, որ ներկայումս մենք տեսնում ենք նրանց հետնորդներին մեծ մասամբ փոփոխված վիճակում. Եվրոպայի և Միացյալ Նահանգների կենտրոնական մասերում։ Այս տեսակետից մենք կարող ենք հասկանալ Հյուսիսային Ամերիկայի և Եվրոպայի օրգանիզմների ազգակցությունը շատ նվազ նմանության դեպքում, մի ազգակցություն, որ վերին աստիճանի ուշագրավ է, եթե ուշադրության առնենք այդ երկու մարզերի հեռավորությունը միմյանցից և բաժանվածությունը ամբողջ Ատլանտյան օվկիանոսով։ Այնուհետև մենք կարող ենք հասկանալ մի քանի դիտողների նկատած ուշագրավ փաստը, որ Եվրոպայի և Ամերիկայի օրգանիզմները երրորդային դարաշրջանի ավելի ուշ շրջանների ընթացքում անհամեմատ ավելի մոտ են եղել միմյանց հետ, քան այժմ․ այդ բացատրվում է նրանով, որ Հին ու Նոր աշխարհների հյուսիսային մասերի ավելի տաք ժամանակաշրջանների ընթացքում գրեթե առանց ընդհատվելու միացած են եղել իրար հետ որպես կամուրջ ծառայող ցամաքով, որը ցրտերի հետևանքով դարձել է անօգուտ թե մեկ և թե մյուս ուղղությամբ տեղափոխվելու համար։
Պլիոցենյան ժամանակաշրջանում, ջերմության դանդաղ կերպով իջնելու ընթացքում, Հին և Նոր աշխարհների տեսակները բևեռային գոտուց գաղթելով դեպի հարավի դրանով իսկ դարձել են մեկը մյուսից կտրված։ Այդ անջատումը, որքանով որ դա վերաբերում է բարեխառն կլիմայի արտադրանքներին, կարող էր տեղի ունենալ շատ դարեր առաջ։ Քանի որ բույսերն ու կենդանիները բոլորը շարժվել են դեպի հարավ, ապա մի լայնածավալ մարզում նրանք պետք է խառնվեին և մրցման մեջ մտնեին բուն ամերիկական արտադրանքների հետ, մյուս մարզում՝ Հին աշխարհի արտադրանքների հետ։ Հետևաբար, մենք այստեղ ունենք բոլոր բարենպաստ պայմանները մեծ փոփոխությունների համար, շատ ավելի մեծ, քան, օրինակ, ալպիական բնակիչների դեպքում, որոնք մեկուսացած մնացել են անհամեմատ ավելի նոր ժամանակաշրջանում զանազան լեռնաշղթաների վրա և Եվրոպայի ու Հյուսիսային Ամերիկայի արկտիկական երկրներում։ Այստեղից էլ հետևում է այն, որ Նոր և Հին աշխարհների բարեխառն մարզում այժմ ապրող ձևերը համեմատելիս մենք գտնում ենք շատ քիչ նույնանման տեսակներ (թեև Ազա-Գրեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 68] վերջերս ցույց է տվել, որ նույնանման բույսերն անհամեմատ ավելի շատ են, քան ենթադրվել է առաջ), բայց յուրաքանչյուր մեծ դասի մեջ մենք գտնում ենք շատ ձևեր, որոնք մի քանի բնախույզների կողմից համարվում են աշխարհագրական ցեղեր, մյուսների կողմից՝ որպես առանձին տեսակներ, և չափազանց շատ մերձավոր կամ միմյանց փոխարինող ձևեր, որոնք բոլոր բնախույզների կողմից համարվում են որպես առանձին տեսակներ։
Պլիոցենյան կամ նույնիսկ փոքր ինչ ավելի վաղ ժամանակաշրջանի ընթացքում բևեռային շրջանի չընդհատվող եզրային գծի երկարությամբ եղած ծովային ֆաունայի դանդաղ տարածումը ինչպես ցամաքի վրա, այնպես էլ ծովի ջրերում, կարող էր առաջ բերել, փոփոխության տեսության համաձայն, խիստ շատ մերձավոր ձևեր, որոնք այժմ ապրում են բոլորովին անջատված ծովային մարզերում։ Այսպիսով, ինչպես ես կարծում եմ, կարելի է բացատրել մի քանի շատ մերձավոր ինչպես դեռևս ապրող, այնպես էլ մահաջնջված երրորդային ձևերի գոյությունը Հյուսիսային Ամերիկայի բարեխառն մարզի արևելյան և արևմտյան ափերին, ինչպես նաև բացատրել շատ մերձավոր խեցեմորթների (ինչպես այդ նկարագրված է Դանայի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 96] հիանալի աշխատության մեջ), մի քանի ձկների և այլ ծովային կենդանիների գոյության զարմանալի փաստը, մի կողմից, Միջերկրական ծովում, մյուս կողմից՝ Ճապոնական ծովերում, թեև այդ մարզերն այժմ բոլորովին բաժանված են իրարից ամբողջ մայր ցամաքով և օվկիանոսի լայնածավալ տարածություններով։
Հյուսիսային Ամերիկայի արևելյան ու արևմտյան ափերի, Միջերկրական և Ճապոնական ծովերի, ինչպես նաև Հյուսիսային Ամերիկայի և Եվրոպայի բարեխառն երկրների այժմյան կամ առաջները բնակված տեսակների մոտ ազգակցության այդ դեպքերը չի կարելի բացատրել տեսակների առանձին-առանձին ստեղծագործության տեսությամբ։ Մենք չենք կարող պնդել, որ այդ տեսակներն ստեղծվել են մարզերի ունեցած գրեթե միանման պայմանների համաձայն, որովհետև համեմատելով, օրինակ Հարավային Ամերիկայի, հարավային Աֆրիկայի և Ավստրալիայի որոշ մասերը, մենք տեսնում ենք երկրներ, որոնք իրենց ֆիզիկական պայմաններով շատ մոտ են միմյանց, բայց բնակիչներով շատ տարբեր են։
ՍԱՌՑԱԴԱՇՏԱՅԻՆ ԺԱՄԱՆԱԿԱՇՐՋԱՆՆԵՐԻ ՀԵՐԹԱՓՈԽՈՒՄԸ ՀՅՈՒՍԻՍՈՒՄ ԵՎ ՀԱՐԱՎՈՒՄ
Սակայն մենք պետք է վերադառնանք այն հարցին, որն անմիջականորեն է վերաբերում մեզ։ Ես համոզված եմ, որ Ֆորբսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] տեսակետները պետք է ընդունվեն ավելի լայն չափերով։ Եվրոպայում մենք հանդիպում ենք չափազանց լրիվ ապացույցների սառցադաշտային ժամանակաշրջանի գոյության մասին՝ սկսած Բրիտանիայի արևմտյան ափերից մինչև Ուրալյան լեռնաշղթան և դեպի հարավ Պիրենյան լեռները։ Սիբիրի լեռնային բուսականության բնույթի և սառած կաթնասունների ուսումնասիրությունը մեզ բերում է այն եզրակացության, որ այդ երկիրն էլ գտնվել է նման պայմաններում։ Ըստ Հուկերի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] Լիբանանում հավերժական ձյունը առաջները ծածկել է կենտրոնական լեռնաշղթան և սկիզբ է տվել սառցադաշտերի, որոնք իջել են 4000 ոտնաչափ [1219 մետր] հովիտների մեջ։ Հենց նույն դիտողը վերջերս հայտնաբերել է խոշոր քարակարկառներ Հյուսիսային Աֆրիկայում Ատլասի լեռնաշղթայի ցածրադիր մակարդակի վրա։ Հիմալայների երկարությամբ սառցադաշտերն իրենց նախկին զարգացման հետքերը թողել են շատ ցածրում, այնպիսի տեղերում, որոնք գտնվում են 900 մղոն հեռու, իսկ Սիկկիմում դոկտ. Հուկերը տեսել է հսկայական հին քարակարկառների վրա աճող եգիպտացորեն։ Ասիական մայր ցամաքից դեպի հարավ, հասարակածի մյուս կողմում, ինչպես մենք գիտենք դոկտ. Յու. Հաաստի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 238] և դոկտ. Հեկտորի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 239] հիանալի հետազոտություններից, նոր Զելանդիայում հսկայական սառցադաշտերն առաջներում իջել են մինչև ցածրադիր մակարդակները, իսկ միևնույն բույսերը, որոնց գտել է Հուկերը այդ կղզու լայն տարածություններով միմյանցից բաժանված լեռների վրա, մեզ պատմում են ծանոթ պատմություն նախկին ցուրտ ժամանակաշրջանի մասին։ Մեծարգո Վ. Բ. Կլարկի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 240] ինձ հաղորդած փաստերից դատելով, ըստ երևույթին, սառցադաշտերի նախկին ներգործության հետքեր են հայտնաբերվել Ավստրալիայի հարավ-արևելյան անկյունի լեռների վրա։
Դառնանք Ամերիկային։ Նրա հյուսիսային կեսում գտնված են գլաքարեր մայր ցամաքի արևելյան մասում՝ մինչև 36—37° դեպի հարավ, իսկ Խաղաղ օվկիանոսի ափերին, որտեղ կլիման ներկայումս այնքան տարբեր է, մինչև 46° դեպի հարավ։ Էռատիկական գլաքարեր նկատված են նաև ժայռոտ լեռների վրա։ Հարավային Ամերիկայի Կորդիլերներում, համարյա հենց հասարակածի մոտերքը, սառցադաշտերը մի ժամանակ իրենց այժմյան մակարդակից շատ ավելի ցած են իջել։ Կենտրոնական Չիլիում ես ուսումնասիրել եմ Պորտիլոյի հովիտը կտրվող խոշոր գլաքարերի մնացորդներից կազմված մի ընդարձակ թումբ, որի վերաբերմամբ հազիվ թե կասկած կարող է լինել, որ այստեղ մի ժամանակ եղել է մի հսկայական քարակարկառ. իսկ մ-ր Դ. Ֆորբսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] հաղորդում է ինձ, որ նա Կորդիլերների զանազան մասերում՝ հարավային լայնության 13° և 30° միջև մոտավորապես 12000 ոտնաչափ [3658 մետր] բարձրության վրա գտել է խոր ակոսված ժայռեր, որոնք նման են իրեն լավ հայտնի նորվեգիական ժայռերին, ինչպես նաև գծված գետաքարեր ունեցող բեկորների զանգվածներ։ Կորդիլերների նշված ամբողջ տարածության վրա այժմ չկան իսկական սառցադաշտեր նույնիսկ շատ ավելի մեծ բարձրությունների վրա։ Ավելի դեպի հարավ, մայր ցամաքի երկու կողմերում, սկսած 41°-ից մինչև նրա հարավային ծայրը, մենք գտնում ենք նախկին սառցադաշտերի ներգործության ամենաակներև ապացույցներն այն բազմաթիվ գլաքարերի մեջ, որոնք իրենց գտնվելու վայրերից հեռու են տարվել։
Այս բազմազան փաստերից, այսինքն նրանից, որ սառցադաշտային գործունեությունը լայն զարգացում է ունեցել հյուսիսային և հարավային կիսագնդերում, նրանից, որ այդ ժամանակաշրջանը երկու կիսագնդերում էլ երկրաբանական իմաստով վերջերս է եղել, թե այնտեղ և թե այստեղ շատ երկար է տևել, ինչպես այդ մասին կարելի է դատել ըստ նրանց թողած հետևանքների, և վերջապես նրանից, որ սառցադաշտերը ոչ այնքան վաղուց շատ ցած են իջել ամբողջ Կորդիլերների երկարությամբ, այս բոլորից ինձ մի ժամանակ թվում էր, թե մենք չենք կարող հրաժարվել այն եզրակացությունից, որ սառցային ժամանակաշրջանի ընթացքում ջերմությունը միաժամանակ է իջել ամբողջ երկրագնդի վրա։ Բայց դրանից հետո մ-ր Կրոլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 194] մի շարք ուշագրավ մեմուարներում փորձել է ապացուցել, որ կլիմայի սառցային պայմանները զանազան ֆիզիկական այնպիսի պատճառների հետևանք են, որոնք պայմանավորված են երկրի ուղեծրի կենտրոնազանցության ավելացմամբ։ Այդ բոլոր պատճառները հասցնում են մի հետևանքի, սակայն իմ աչքում նրանցից ամենակարևորն է ուղեծրի կենտրոնազանցության անուղղակի ազդեցությունը օվկիանոսային հոսանքների վրա։ Ըստ մ-ր Կրոլի՝ ցուրտ ժամանակաշրջանները կրկնվում են յուրաքանչյուր 10 կամ 15 հազար տարին մի անգամ, և այդ ժամանակաշրջանները մեծ ժամանակամիջոցներից հետո լինում են չափազանց դաժան՝ կախված զանազան պատահական հանգամանքներից, որոնցից, ինչպես ցույց է տվել Չ. Լյաելը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] ամենակարևորն է ցամաքի և ջրի հարաբերական դրությունը։ Մ-ր Կրոլը կարծում է, որ վերջին մեծ սառցադաշտային ժամանակաշրջանը եղել է մոտ 240000 տարի առաջ և կլիմայի փոքրիկ տատանումներով տևել է շուրջ 160000 տարի։ Ինչ վերաբերում է ավելի հին սառցադաշտային ժամանակաշրջաններին, ապա մի քանի երկրաբաններ ուղղակի ապացույցների հիման վրա համոզված են, որ նրանք տեղի են ունեցել միոցենի և էոցենի ընթացքում, չհիշատակելով էլ ավելի հին ֆորմացիաները։ Սակայն մ-ր Կրոլի հասած հետևություններից մեզ համար ամենակարևոր է այն, որ երբ հյուսիսային կիսագունդը ցուրտ ժամանակաշրջան է ապրում, հարավային կիսագնդի ջերմությունը բարձրանում է, և նրա ձմեռները անհամեմատ ավելի մեղմանում են, գլխավորապես օվկիանոսային հոսանքների ուղղության փոփոխության շնորհիվ։ Եվ ընդհակառակը, նույնը կլինի հյուսիսային կիսագնդի հետ, երբ հարավային կիսագունդը սառցադաշտային ժամանակաշրջան է անցնում։ Այդ եզրակացություններն այնքան առատ լույս են սփռում օրգանիզմների աշխարհագրական տարածման վրա, որ ես լիովին հակված եմ դրանք ճիշտ համարելու, բայց սկզբում ես կհաղորդեմ փաստեր, որոնք բացատրության կարիք ունեն։
Դոկտոր Հուկերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] Հարավային Ամերիկայի մասին ցույց է տվել, որ բացի բազմաթիվ մերձավոր տեսակներից, Հրո երկրի քառասունից մինչև հիսուն ծաղկավոր բույսեր, որոնք կազմում են նրա աղքատիկ բուսական աշխարհի բավական զգալի մասը, ընդհանուր են Հյուսիսային Ամերիկայի և Եվրոպայի բույսերի հետ, չնայած այն հսկայական հեռավորությանը, որր բաժանում է տարբեր կիսագնդերում գտնվող այդ երկու մայր ցամաքները։ Հասարակածային Ամերիկայի բարձր լեռների վրա հանդիպում են բազմաթիվ յուրահատուկ տեսակներ, որոնք պատկանում են եվրոպական սեռերին։ Բրազիլիայի Օրգանի լեռներում Գարդները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 241] գտել է բարեխառն Եվրոպայի մի քանի, անտարկտիկական և անդական մի քանի սեռերի ներկայացուցիչներ, որոնք չեն հանդիպում հարակից տաք ցածրավայրերում։ Սիլյա դը Կարակաս լեռնաշղթայի վրա հռչակավոր Հումբոլդտը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 199] դեռ վաղուց գտել է տեսակներ, որոնք պատկանում են Կորդիլերների բնորոշ սեռերին։
Եվրոպայի համար զանազան բնորոշող ձևերը և Բարեհուսո հրվանդանի բուսական աշխարհի փոքրաթիվ ներկայացուցիչները հանդիպում են Աֆրիկայում Հաբեշստանի լեռների վրա։ Բարեհուսո հրվանդանի վրա գտնված են շատ քիչ եվրոպական տեսակներ, որոնք, ինչպես կարծում են, մարդու միջոցով չեն բերվել այնտեղ, իսկ լեռների վրա գտնվել են եվրոպականներին փոխարինող մի քանի ձևեր, որոնք չեն հայտնաբերված Աֆրիկայի մերձարևադարձային մասերում։ Դոկտ. Հուկերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] վերջերս ցույց է տվել նաև մի քանի բույսեր, որ ապրում են Ֆերնանդո-Պո բարձր կղզու բարձրադիր մասերում և հարևան Կամերունի լեռների վրա, Գվինեի ծոցում, շատ մոտ են այն ձևերին, որոնք ապրում են Հաբեշստանի լեռների վրա, ինչպես նաև նրանց, որոնք պատկանում են բարեխառն Եվրոպային։ Ներկայումս, ինչպես ես լսել եմ դոկտ. Հուկերից, բարեխառն գոտու նույն բույսերից մի քանիսին հայտնաբերել է մեծարգո Ռ. Թ. Լոուն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 231] Կանաչ հրվանդանի կղզիների լեռների վրա։ Բարեխառն գոտու միևնույն ձևերի այդպիսի տարածվածությունը գրեթե հասարակածի մոտերը, Աֆրիկայի ամբողջ մայր ցամաքի լայնությամբ և մինչև Կանաչ հրվանդանի արշիպելագի լեռները, հանդիսանում է բույսերի տարածման ամենազարմանալի փաստերից մեկը։
Հիմալայան լեռների վրա, Հնդկաստանի առանձնացած լեռնաշղթաների, Ցեյլոնի բարձունքների և Ճավայի հրաբխային գագաթների վրա կան շատ բույսեր, որոնք կամ նման են, կամ փոխարինում են միմյանց և դրա հետ միասին փոխարինում են Եվրոպայի բույսերին, բայց գտնված լեն նրանց միջև գտնվող տաք ցածրավայրերում։ Ճավայի ավելի բարձր գագաթների վրա հավաքված բույսերի սեռերի ցուցակը հիշեցնում են այն կոլեկցիայի ցուցակագրությունը, որ հավաքված է Եվրոպայի բլուրների վրա։ Ավելի ևս զարմանալի է այն փաստը, որ ավստրալիական յուրահատուկ ձևերը ունեն որոշ ներկայացուցիչներ, որոնք աճում են Բորնեո կղզու լեռնագագաթների վրա։ Ավստրալիական ձևերից մի քանիսը, ինչպես ես այդ իմացել եմ դոկտ. Հուկերից,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] տարածված են Մալակկա թերակղզու լեռներում և երբեմն էլ ցրված են մի կողմից մինչև Հնդկաստան, մյուս կողմից՝ դեպի հյուսիսից մինչև Ճապոնիա։
Ավստրալիայի հարավային լեռների վրա դոկտ. Ֆ. Մյուլլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] հայտնաբերել է եվրոպական մի քանի տեսակներ․ մարդու միջոցով չբերված այլ տեսակներ հանդիպում են ցածրավայրերում և, ինչպես հաղորդում է ինձ Հուկերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] կարելի է տալ եվրոպական սեռերի մի երկար ցուցակ, որոնք գտնված են Ավստրալիայում, բայց գտնված չեն միջանկյալ տաք մարզերում։ Դոկտոր Հուկերն իր «Ներածություն Նոր Զելանդիայի ֆլորայի ուսումնասիրության» սքանչելի աշխատության մեջ բերում է նույնանման զարմանալի փաստեր այդ մեծ կղզու բույսերի վերաբերյալ։ Այստեղից մենք տեսնում ենք, որ որոշ բույսեր, որոնք աճում են աշխարհիս բոլոր մասերի արևադարձային գոտու ավելի բարձր լեռների վրա և հյուսիսային ու հարավային կիսագնդերի բարեխառն գոտիների հարթավայրերում, կամ միևնույն տեսակներն են, կամ թե միևնույն տեսակների տարատեսակները։ Սակայն անհրաժեշտ է նկատել, որ այդ բույսերը բառիս ճիշտ իմաստով արկտիկական չեն, որովհետև, ըստ մ-ր Հ. Կ. Ուոտսոնի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 62] դիտողության, «բևեռներից դեպի հասարակածային լայնությունները նահանջելով ալպիական կամ լեռնային բուսական աշխարհը ավելի ու ավելի քիչ արկտիկական են դառնում»։ Բացի այս նույնական և չափազանց մերձավոր ձևերից, շատ տեսակներ, որոնք բնակվում են միևնույն, միմյանցից հեռու գտնվող մարզերում, պատկանում են արևադարձային միջանկյալ ցածրավայրերում այժմ չհանդիպող սեռերին։
Այս համառոտ դիտողությունները վերաբերում են միայն բույսերին, բայց կան մի քանի փաստեր նաև ցամաքային կենդանիների վերաբերմամբ։ Ծովային արտադրանքների համար նմանապես կարելի է բերել նման տվյալներ, այսպես, օրինակ, ես կարող եմ հիմնվել այնպիսի մի բարձր հեղինակության վկայությունների վրա, որպիսին է պրոֆ. Դանան,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 96] որ «իհարկե, զարմանալի է, որ Նոր Զելանդիան իր խեցեմորթներով ավելի մոտ է կանգնած Մեծ Բրիտանիային, իր հակոտնյային, քան աշխարհի որևէ այլ մի մասի»։ Սըր Ջ. Ռիչարդսոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 112] նույնպես ասում է, որ Նոր Զելանդիայի, Թասմանիայի և այլ ափերի մոտ երևան են գալիս ձկների հյուսիսային ձևեր։ Դ-ր Հուկերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] հաղորդում է ինձ, որ քսանհինգ տեսակ ջրիմուռներ ընդհանուր են Նոր Զելանդիայի և Եվրոպայի համար, բայց գտնված չեն միջանկյալ արևադարձային ծովերում։
Բերված փաստերի հիման վրա, այսինքն, որ բարեխառն գոտիների ձևերը գտնվում են ամբողջ հասարակածային Աֆրիկայի և Հնդկաստանի լեռների վրա՝ մինչև Ցեյլոն և Մալայան արշիպելագը և ավելի նվազ չափով արևադարձային Հարավային Ամերիկայի ընդարձակ տարածությունների վրա, կարելի է համարյա հաստատապես ընդունել, որ ավելի հին ժամանակաշրջաններից մեկում, անպայման սառցադաշտային ժամանակաշրջանի ամենախիստ ժամանակ, այդ ընդարձակ մայր ցամաքների ցածրավայրերը հասարակածի մոտերքը ամենուրեք բնակված են եղել մեծ քանակությամբ բարեխառն գոտու ձևերով։ Այդ ժամանակաշրջանում հասարակածային կլիման ծովի մակարդակի վրա հավանորեն եղել է մոտավորապես այնպես, ինչպես այժմ է այդ լայնության վեց հազար ոտնաչափ բարձրության վրա, գուցե թե նույնիսկ մի քիչ ավելի ցուրտ։ Այդ ամենացուրտ ժամանակաշրջանի ընթացքում հասարակածի մոտերքի ցածրավայրերը պետք է ծածկվեին հասարակածային և բարեխառն երկրների խառը բուսականությամբ, նման Հուկերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] նկարագրած այն փարթամ բուսականությանը, որը պատում է Հիմալայան լեռների ստորին լանջերը չորսից հինգ հազար ոտնաչափ բարձրության վրա, բայց գուցե և բարեխառն գոտու ձևերն ավելի գերակշռություն են ունեցել։ Ճիշտ այդպես էլ մ-ր Մենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 242] հայտնաբերել է, որ Գվինեի ծոցում Ֆերնանդո-Պո լեռնոտ կղզու եվրոպական բարեխառն ձևերն սկսում են երևան գալ մոտավորապես հինգ հազար ոտնաչափ բարձրության վրա։ Պանամայի լեռների վրա, ընդամենը երկու հազար ոտնաչափ բարձրության վրա, դոկտ. Զեեմանը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 243] գտել է մեքսիկականին նման բուսականություն, որն «ունի տաք գոտու ձևեր, հավասարաչափորեն խառնված բարեխառն ձևերի հետ»։
Այժմ տեսնենք, թե մ-ր Կրոլի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 194] եզրակացությունը, որի համաձայն այն ժամանակ, երբ հյուսիսային կիսագունդը կրում էր մեծ սառցադաշտային ժամանակաշրջանի ծայրահեղ ցուրտը, հարավայինը եղել է զգալիորեն ավելի տաք, արդյոք ինչքան է լուսաբանում տարբեր օրգանիզմների անբացատրելի թվացող այժմյան տարածումը երկու կիսագնդերի բարեխառն մասերում և արևադարձային գոտու լեռներում։ Սառցադաշտային ժամանակաշրջանը տարիներով հաշված պետք է շատ երկար ժամանակ տևած լինի, և եթե մենք հիշենք, թե նատուրալիզացիայի ենթարկված մի քանի բույսեր և կենդանիներ փոքրաթիվ դարերի ընթացքում ինչպիսի հսկայական տարածությունների վրա կարողանում են տարաբնակվել, ապա այդ ժամանակաշրջանը բավական կլինի ամեն չափի գաղթերի համար։ Մենք գիտենք, որ քանի ցուրտն ավելի ու ավելի է ուժեղացել, այնքան արկտիկական ձևերն ավելի տարածվել են բարեխառն մարզերում, իսկ հենց նոր մեր հաղորդած տվյալների հիման վրա հազիվ թե մեզ մոտ որևէ կասկած կարող է մնալ, որ ամենաուժեղ տիրապետող և լայն տարածված բարեխառն ձևերից մի քանիսն իրենց հերթին գրավել են հասարակածային ցածրավայրերը։ Այդ տաք ցածրավայրերի բնակիչները միաժամանակ պետք է գաղթեին դեպի հարավային կիսագնդի արևադարձային և մերձարևադարձային մասերը, որովհետև նա այդ ժամանակաշրջանի ընթացքում ավելի տաք է եղել։ Սառցադաշտային ժամանակաշրջանի վերջերին, երբ երկու կիսագնդերը հետզհետե ձեռք են բերել իրենց նախկին ջերմությունը, հյուսիսային բարեխառն ձևերը, որ մինչև այդ ապրել են հասարակածի մոտերքի ցածրավայրերում, կամ դուրս են մղվել դեպի իրենց նախկին մարզերը, կամ էլ ոչնչացել են ու փոխարինվել են հարավից վերադարձած հասարակածային ձևերով։ Սակայն շատ հավանական է, որ հյուսիսային բարեխառն ձևերից մի քանիսը բարձրացել են մոտակա բարձրությունները, որտեղ բավականաչափ բարձրության դեպքում նրանք կարող էին պահպանվել երկար ժամանակ Եվրոպայի լեռների վրա պահպանված արկտիկական ձևերի նման։ Նրանք կարող էին այստեղ պահպանվել, եթե նույնիսկ կլիման այնքան էլ պիտանի չլիներ նրանց համար, որովհետև ջերմության փոփոխությունբ պետք է տեղի ունենար չափազանց դանդաղորեն, իսկ բույսերը, անկասկած, որոշ չափով ընդունակություն ունեն կլիմայավարժվելու. այդ երևում է նրանից, որ նրանք իրենց հաջորդներին են փոխանցում զանազան կառուցվածքային առանձնահատկություններ, որոնք թույլ են տալիս նրանց դիմադրել տաքին ու ցրտին։
Երևույթների կանոնավոր հաջորդականության դեպքում հարավային կիսագունդն իր հերթին ենթարկվել է դաժան սառցադաշտային ժամանակաշրջանին, մինչդեռ հյուսիսային կիսագնդում ավելի տաք էր, որի հետևանքով հարավային բարեխառն ձևերը կարող էին գրավել հասարակածային ցածրավայրերը։ Հյուսիսային ձևերը, որոնք մինչև այդ մնացել էին լեռների վրա, այդ ժամանակ կարող էին իջնել ցածրավայրերը և խառնվել հարավային ձևերի հետ։ Վերջիններս, տաքության վերադարձի հետ միասին, պետք է վերադառնային իրենց նախկին մարզերը, թողնելով որոշ տեսակներ լեռների վրա և տանելով իրենց հետ մի քանի հյուսիսային բարեխառն ձևեր, որոնք իջել են իրենց լեռնային ապաստարաններից։ Դրա հետևանքով մենք կկարողանայինք գտնել ինչպես հյուսիսային և հարավային գոտիներում, այնպես էլ միջանկյալ արևադարձային մարզերի լեռների վրա սակավաթիվ տեսակներ, որոնք միանգամայն միևնույնն են։ Սակայն այն տեսակները, որոնք երկար ժամանակ մնացել են այդ լեռների վրա կամ զանազան կիսագնդերում, պետք է մրցման մեջ մտնեին որոշ նոր ձևերի հետ և ընկնեին մի քիչ տարբեր ֆիզիկական պայմանների մեջ. դրա շնորհիվ նրանք կարող էին ազատ կերպով ենթարկվել փոփոխության և ընդհանրապես դառնալ այժմ կամ տարատեսակներ, կամ փոխարինող տեսակներ, և իրոք այդ այդպես էլ է։ Մենք չպետք է մոռանանք նաև այն, որ երկու կիսագնդերում գոյություն են ունեցել սառցադաշտային շրջաններ ավելի վաղ դարաշրջաններում․ նրանք, համաձայն նույն սկզբունքների, մեզ բացատրում են, թե ինչու բոլորովին տարբեր տեսակներ բնակվում են միմյանցից հեռու ընկած մարզերում և պատկանում են այնպիսի սեռերի, որոնք այժմ չեն հանդիպում միջանկյալ տաք գոտիներում։
Վերին աստիճանի ուշագրավ է, որ նման կամ թեթևակի փոփոխված շատ տեսակներ գաղթել են հյուսիսից դեպի հարավ, այլ ոչ թե հակառակ ուղղությամբ. այդ խիստ կերպով մատնանշել է Հուկերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] Ամերիկայի և Ալֆոնս դը-Կանդոլը՝[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] Ավստրալիայի վերաբերմամբ։ Միայն քիչ թվով հարավային ձևեր մենք տեսնում ենք Բորնեոյի և Հաբեշստանի լեռների վրա։ Ես կասկածում եմ, որ այդ գերակշռող գաղթը հյուսիսից դեպի հարավ բացատրվում է ցամաքի հյուսիսում ունեցած ավելի մեծ տարածությամբ և նրանով, որ հյուսիսային ձևերն իրենց բուն մարզում գոյություն են ունեցել ավելի մեծ քանակությամբ. դրա հետևանքով բնական ընտրությունը և մրցությունը նրանց հասցրել են ավելի բարձր աստիճանի կատարելության կամ գերիշխելու ընդունակության, քան հարավային ձևերին. այդ պատճառով էլ, երբ երկու խմբերի անհատները խառնվել են միմյանց հետ հասարակածային մարզերում սառցադաշտային ժամանակաշրջանների փոխառփոխ կատարվող զարգացման ընթացքում, հյուսիսային ձևերն ավելի ուժեղ են դուրս եկել և ընդունակ են եղել սկզբում պահպանել իրենց գրաված վայրերը լեռների վրա, իսկ հետո նաև տարաբնակվել դեպի հարավի հարավային ձևերի հետ միասին. բայց այդպես չի եղել վերջիններիս հետ հյուսիսայինների նկատմամբ։ Ճիշտ այդպես էլ մենք տեսնում ենք ներկայումս, թե եվրոպական շատ ձևեր ծածկում են Լա-Պլատայի, Նոր-Զելանդիայի և ավելի քիչ չափով Ավստրալիայի հողը, դուրս մղելով տեղական բնիկներին, մինչդեռ չափազանց քիչ հարավային ձևեր որոշ տեղերում նատուրալիզացիայի են ենթարկել հյուսիսային կիսագնդում, թեև մորթիներ, բուրդ և այլ առարկաներ, որոնց մեջ սերմերը կարող էին պահպանվել ու տեղափոխվել, լայն չավերով Եվրոպա են ներմուծվում Լա-Պլատայից երկու կամ երեք դար շարունակ և Ավստրալիայից՝ վերջին քառասունից-հիսուն տարիների ընթացքում։ Սակայն Նիլգերի լեռները Հնդկաստանում բացառություն են ներկայացնում, որովհետև այստեղ, ինչպես ես այդ գիտեմ դ-ր Հուկերի ասածից, ավստրալիական ձևերն արագ կերպով և՛ բազմանում են, և՛ նատուրալիզացիայի ենթարկվում։ Կասկած չկա, որ վերջին սառցադաշտային ժամանակաշրջանից առաջ մերձարևադարձային լեռները ծածկված են եղել էնդեմիկ ալպիական ձևերով, բայց վերջիններս գրեթե ամենուրեք տեղի են տվել ավելի տարածված ձևերի առաջ, որոնք զարգացել են հյուսիսի ավելի լայնածավալ մարզերում և ավելի գործոն արտադրողական կենտրոններում։ Շատ կղզիների վրա բնական արտադրանքները թվով կամ հավասարվել են նատուրալիզացիայի ենթարկվածներին կամ նույնիսկ զիջում են նրանց, որը մահաջնջվելու առաջին քայլն է։ Լեռները դրանք կղզիներ են ցամաքի վրա, և նրանց բնակիչներն իրենց տեղը զիջել են այն ձևերին, որոնք զարգացել են հյուսիսի ընդարձակ մարզերում, ճիշտ այնպես, ինչպես որ իսկական կղզիների բնակիչները զիջել են ու շարունակում են զիջել իրենց տեղերը մայր ցամաքի այն ձևերին, որոնք այստեղ նատուրալիզացիայի են ենթարկվում մարդու միջոցով։
Այդ նույն սկզբունքներն էլ կիրառվում են բացատրելու համար ինչպես հյուսիսային ու հարավային բարեխառն գոտիների, այնպես էլ մերձարևադարձային լեռների ցամաքային և ծովային կենդանիների տարածումը։ Երբ սառցադաշտային ժամանակաշրջանի առավելագույն զարգացման ընթացքում օվկիանոսային հոսանքները խիստ տարբերվել են այժմյան հոսանքներից, բարեխառն ծովերի բնակիչներից մի քանիսը կարող էին հասնել հասարակածին. թերևս նրանցից քչերին է հաջողվել իսկույն տարածվել դեպի հարավ, օգտվելով ավելի սառ հոսանքներով, մինչդեռ մյուսները կարող էին մնալ և ապրել ավելի խոր ու սառ շերտերում, մինչև որ հարավային կիսագունդն էլ իր հերթին ձեռք կբերեր սառցադաշտային կլիմա, որը հնարավոր է դարձրել նրանց հետագա շարժումը ճիշտ այնպես, ինչպես, ըստ Ֆորբսի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] ներկայումս գոյություն ունեն արկտիկական օրգանիզմներ ունեցող մեկուսացած տեղամասեր հյուսիսային բարեխառն ծովերի ավելի խոր մասերում։
Ես հեռու եմ այն մտքից, որ ենթադրեմ, թե բոլոր դժվարությունները, որոնք կապված են հյուսիսում ու հարավում և որոշ դեպքերում միջանկյալ լեռնաշղթաների վրա մեկը մյուսից հսկայական հեռավորության վրա այժմ ապրող միևնույն մերձավոր տեսակների տարածման և հարաբերակցության հետ, վերանում են վերոհիշյալ կշռադատություններով։ Տարաբնակման ճշգրիտ սահմանները տրվել չեն կարող։ Մենք չենք կարող ասել, թե ինչու տարաբնակվել են այս տեսակները և ոչ մյուսները, թե ինչու որոշ տեսակներ փոփոխվել են ու նոր ձևերի սկիզբ տվել, մինչդեռ, մյուսները մնացել են անփոփոխ։ Մենք չենք կարող հույս ունենալ, որ կգտնենք այդ փաստերի բացատրությունը, քանի դեռ ի վիճակի չենք ասելու, թե ինչու մարդու միջոցով մեկը օտար երկրում նատուրալիզացիայի ենթարկվում է, իսկ մյուսը ոչ, թե ինչու մի տեսակ երկու և երեք անգամ ավելի հեռու է տարածվում և երկու երեք անգամ ավելի սովորական է, քան մյուսը իր գրաված մարզում։
Բացի դրանից, դեռ մնում են լուծելու մի շարք բոլորովին առանձնահատուկ դժվարություններ, ինչպես, օրինակ, միևնույն բույսերի ներկայությունը, որ ապացուցել է Հուկերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] միմյանցից այնքան հեռու գտնվող կետերում, ինչպիսիք են՝ Կերգուելենը, Նոր Զելանդիան և Հրո երկիրը. սակայն Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] կարծում է, որ այդ դեպքում տարաբնակում կարող է տեղի ունենալ լողացող սառույցների մասնակցությամբ։ Վերին աստիճանի ուշագրավ է նաև այն, որ հարավային կիսագնդի ինչպես նշված, այնպես էլ միմյանցից հեռու ցրված կետերում գոյություն ունեն տեսակներ, որոնք թեև տարբեր են, բայց պատկանում են բացառապես հարավին հատուկ սեռերին։ Այդ տեսակներից մի քանիսը այնքան տարբեր են, որ մենք չենք կարող ենթադրել, թե նրանք վերջին սառցադաշտային ժամանակաշրջանի սկզբից սկսած բավականաչափ ժամանակ են ունեցել տարաբնակվելու և դրա հետ կապված անհրաժեշտ աստիճան փոփոխություններ կրելու համար։ Կարծես փաստերը վկայում են այն մասին, որ մի սեռի առանձին տեսակները տարածվել են մի ընդհանուր կենտրոնից շառավիղների ուղղությամբ, և ես հակված եմ համարելու, որ ինչպես հարավային, այնպես էլ հյուսիսային կիսագնդում վերջին սառցադաշտային ժամանակաշրջանի սկսվելուց առաջ եղել է ավելի տաք ժամանակաշրջան, որի ընթացքում անտարկտիկական երկրները, որ այժմ ծածկված են սառույցով, ունեցել են վերին աստիճանի յուրահատուկ և առանձնացած բուսական աշխարհ։ Կարելի է կարծել, որ նախքան սառցադաշտային ժամանակաշրջանում այդ բուսականության մահաջնջվելը, նրա փոքրաթիվ ներկայացուցիչներն արդեն կարողացել էին լայնորեն տարածվել հարավային կիսագնդի զանազան մասերում տարաբնակման պատահական եղանակների մասնակցությամբ և կանգ առնելով այժմ սուզված կղզիների վրա։ Այս կերպով Ամերիկայի, Ավստրալիայի և Նոր Զելանդիայի հարավային ափերը կարողացել են ձեռք բերել կյանքի միևնույն յուրահատուկ ձևերը։
Սըր Չ. Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] իր գրքի մի ուշագրավ տեղում համարյա ինձ նման է արտահայտվել օրգանիզմների աշխարհագրական տարածման վրա երկրագնդի կլիմաների հերթափոխության ազդեցության մասին։ Այժմ մենք տեսանք, որ մ-ր Կրոլի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 194] եզրակացությունները մի կիսագնդի հաջորդական սառցադաշտային ժամանակաշրջանների մյուս կիսագնդի ավելի տաք ժամանակաշրջաններին զուգադիպելու մասին, ընդունելով դրա հետ միասին տեսակների դանդաղ փոփոխությունը, բացատրություն են տալիս այն բազմաթիվ փաստերին, որոնք վերաբերում են օրգանիզմների ինչպես միևնույն, այնպես էլ մերձավոր ձևերի տարածմանը աշխարհի բոլոր մասերում։ Կենդանի հեղեղը տարածվել է մի ժամանակաշրջանի ընթացքում հյուսիսից և մյուսի ընթացքում հարավից, ըստ որում թե մեկ և թե մյուս դեպքում հասել է հասարակածին. բայց նա ավելի մեծ ուժով ընթացել է հյուսիսից, քան հակառակ ուղղությամբ, և այդ պատճառով էլ հարավն ավելի լիովին է հեղեղվել։ Ինչպես որ մակընթացությունը թողնում է իր բերվածքները հորիզոնական շերտերով, ավելի վեր բարձրանալով եզրային շերտում, որտեղ ալիքները հասնում են առավելագույն բարձրության, այնպես էլ կենդանի հեղեղն իր կենդանի նստվածքները թողել է մեր լեռնագագաթների վրա, այն գծի ուղղությամբ, որն սկսվելով արկտիկական ցածրավայրերից, ամենամեծ բարձրության է հասնում հասարակածի մոտերքը։ Այդ ժամանակ ափ դուրս նետված զանազան էակներին կարելի է համեմատել մարդկային վայրենի ցեղերի հետ, որոնք շպրտված են անմատչելի լեռնային կիրճերը գրեթե յուրաքանչյուր երկրում, որտեղ նրանք պահպանվել են, ծառայելով մեզ հետաքրքրությամբ լի մի հիշեցում շրջապատող ցածրավայրերի նախկին բնակիչների մասին։
ԳԼՈՒԽ XIII։ ԱՇԽԱՐՀԱԳՐԱԿԱՆ ՏԱՐԱԾՈՒՄ (ՇԱՐՈՒՆԱԿՈՒԹՅՈՒՆ)
ՔԱՂՑՐԱՀԱՄ ՋՐԵՐԻ ՕՐԳԱՆԻԶՄՆԵՐԻ ՏԱՐԱԾՈՒՄԸ։— ՕՎԿԻԱՆՈՍԱՅԻՆ ԿՂԶԻՆԵՐԻ ԲՆԱԿԻՉՆԵՐԸ։— ԵՐԿԿԵՆՑԱՂՆԵՐԻ ԵՎ ՑԱՄԱՔԱՅԻՆ ԿԱԹՆԱՍՈՒՆՆԵՐԻ ԲԱՑԱԿԱՅՈՒԹՅՈՒՆԸ։— ԿՂԶԻՆԵՐԻ ԲՆԱԿԻՉՆԵՐԻ ԱՌՆՉՈՒԹՅՈՒՆԸ ՄԵՐՁԱՎՈՐ ՄԱՅՐ ՑԱՄԱՔԻ ԲՆԱԿԻՉՆԵՐԻՆ։— ՕՐԳԱՆԻԶՄՆԵՐԻ ԳԱՂԹԸ ԱՄԵՆԱՄՈՏ ՍԿԶԲՆԱՏԵՂԻՑ ԵՎ ՀԵՏԱԳԱ ՓՈՓՈԽՈՒԹՅՈՒՆԸ։— ՆԱԽՈՐԴ ԵՎ ՆԵՐԿԱ ԳԼՈՒԽՆԵՐԻ ՀԱՄԱՌՈՏ ԱԿՆԱՐԿ։
ՔԱՂՑՐԱՀԱՄ ՋՐԵՐԻ ՕՐԳԱՆԻԶՄՆԵՐԸ
Քանի որ լճերն ու գետային սիստեմները բաժանված են միմյանցից ցամաքի տեղամասերով, ապա կարելի է կարծել, որ քաղցրահամ ջրերի օրգանիզմները լայն կերպով տարածված չեն լինում միևնույն երկրի սահմաններում, իսկ քանի որ ծովը ավելի խոշոր արգելք է ներկայացնում, ապա կարելի է գալ այն եզրակացության, որ նրանք երբեք չեն տարածվում դեպի հեռավոր երկրները։ Իրականության մեջ ճիշտ հակառակն է նկատվում։ Հսկայական տարածում ունեն ոչ միայն զանազան դասերի պատկանող քաղցրահամ ջրային շատ տեսակներ, այլև նրանց մերձավոր տեսակները նկատելի կերպով գերիշխում են ամբողջ երկրագնդի վրա։ Ես լավ հիշում եմ, որ առաջին անգամ Բրազիլիայի քաղցրահամ ջրերում կոլեկցիա հավաքելիս ես վերին աստիճանի զարմացել էի այդտեղ քաղցրահամ ջրերի միջատների կակղամորթների և այլոց նմանությամբ ու դրանց կողքին ապրող ցամաքային էակների ոչ նմանությամբ Բրիտանիայում ապրող համապատասխան ձևերի հետ։
Ես կարծում եմ, որ քաղցրահամ ջրերի օրգանիզմների լայն կերպով տարածվելու ընդունակությունը կարող է մեծ մասամբ բացատրվել նրանով, որ նրանք ամենաօգտակար կերպով հարմարվել են իրենց հաճախակի տեղափոխությունների համար կարճ տարածությունների վրա՝ լճակից լճակ, կամ հոսանքից հոսանք իրենց բուն երկրների սահմաններում, լայնորեն տարաբնակվելու հակումը բխում է այդ ընդունակությունից գրեթե որպես անհրաժեշտ հետևանք։ Մենք այստեղ կարող ենք կանգ առնել միայն մի քանի դեպքերի վրա, և նրանց մեջ բացատրելու համար ամենադժվարներից մեկը վերաբերում է ձկներին։ Առաջները կարծում էին, թե քաղցրահամ ջրում ապրող միաբույն տեսակը երբեք չի հանդիպում երկու միմյանցից հեռու գտնվող մայր ցամաքների վրա։ Սակայն դոկտոր Գյունտերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 154] վերջերս ապացուցել է, որ Galaxios attenuatus-ը բնակվում է Թասմանիայիում, Նոր Զելանդիայում, Ֆալկլանդյան կղզիների և Հարավային Ամերիկայի մայր ցամաքի վրա։ Դա ներկայացնում է մի ուշագրավ դեպք և հավանորեն ցույց է տալիս, որ նրա տարածումը տեղի է ունեցել ավելի վաղ տաք ժամանակաշրջանում անտարկտիկական կենտրոնից։ Սակայն այդ դեպքը որոշ չափով ավելի պակաս զարմանալի է դառնում այն պատճառով, որ այդ սեռին պատկանող տեսակները օժտված են մեզ համար անհայտ ձևով բաց օվկիանոսի մեծ տարածություններ անցնելու ընդունակությամբ, այսպես, այդ տեսակներից մեկը Նոր Զելանդիայի և Աուկլանդյան կղզիների համար ընդհանուր է, թեև նրանք բաժանված են իրարից մոտավորապես 230 մղոն տարածությամբ։ Միևնույն մայր ցամաքի սահմաններում քաղցրահամ ջրերի ձկները հաճախ տարածված են լինում թե լայն կերպով և թե կարծես առանց պատճառի. այսպես, օրինակ, իրար կողքի գտնվող երկու գետային սիստեմներում որոշ տեսակներ կարող են ընդհանուր լինել, իսկ մյուսները բոլորովին տարբեր։
Հավանորեն նրանք երբեմն տարածվել են, այսպես կոչված, պատահական պատճառների օգնությամբ։ Այսպես, ձկները դեռ կենդանի վիճակում, ոչ այնքան հազվագյուտ դեպքերում, մրրիկներով տարվում են հեռավոր վայրեր, և հայտնի է, որ ձկնկիթը պահպանում է իր կենսունակությունը բավական երկար ժամանակ ջրից դուրս հանվելուց հետո։ Սակայն քաղցրահամ ջրերի ձկների տարաբնակումը կարելի է գլխավորապես վերագրել վերջին երկրաբանական ժամանակաշրջանի ընթացքում ցամաքի մակերևույթի փոփոխություններին, որի հետևանքով առաջացել է զանազան գետերի միախառնում։ Բացի դրանից, այդպիսի դեպքեր լինում են հեղեղների ժամանակ նաև առանց մակերևույթի որևէ փոփոխության։ Այն մեծ տարբերությունը, որ կա ձկների միջև չընդհատվող լեռնաշղթաների հակառակ կողմերում, որոնց գոյությունը, հետևաբար, կարող էր վաղուց ի վեր արգելակել այս կամ այն կողմի զանազան սիստեմների միացումը, բերում է նույն եզրակացությանը։ Քաղցրահամ ջրերի մի քանի ձկներ պատկանում են շատ հին ձկներին, և այդպիսի դեպքերում բավականաչափ ժամանակ է եղել ինչպես աշխարհագրական ընդարձակ փոփոխությունների համար, այնպես էլ ժամանակ և եղանակներ են գտնվել մեծ տեղափոխությունների համար։ Բացի դրանից, դոկտ. Գյունտերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 154] մի շարք նկատառումների հիման վրա, ձկների վերաբերմամբ եկել է այն եզրակացության, որ միևնույն ձևերն ունեն շատ երկարատև գոյություն։ Որոշ ջանք գործադրելով ծովային ձկանը կարելի է ընտելացնել քաղցրահամ ջրային կյանքին, իսկ ըստ Վալյանսենի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 244] հազիվ թե կա ձկների գեթ մի խումբ, որի բոլոր անդամները քաղցրահամ ջրի հարմարված լինեին, դրա հիման վրա ծովային տեսակը, որը պատկանում է քաղցրահամ ջրերի խմբին, կարող է ափի երկարությամբ մեծ տարածությունների վրա թափառել և հավանորեն կարող է առանց մեծ դժվարության հարմարվել հեռավոր երկրների քաղցրահամ ջրերին։
Քաղցրահամ ջրերի կակղամորթների որոշ տեսակներ ունեն տարածման շատ ընդարձակ մարզեր, և երկրագնդի վրա գերակշռող են հանդիսանում մերձավոր տեսակները, որոնք մեր տեսության համաձայն ծագել են մի ընդհանուր նախածնողից և պիտի տարածվեին՝ դուրս գալով մի հիմնական մարզից։ Այդ կակղամորթների տարածումը սկզբում ինձ շատ էր շփոթեցնում, որովհետև դժվար թե հավանական լինի, որ նրանց ձվերը կարող էին փոխադրվել թռչունների միջոցով, մանավանդ որ թե ձվերը և թե կենդանիներն իրենք անմիջապես փչանում են ծովի ջրի մեջ։ Ես չէի կարողանում նույնիսկ հասկանալ, թե նատուրալիզացիայի ենթարկված մի քանի տեսակներ ինչպես կարողացել են արագ կերպով տարածվել երկրի վրա։ Սակայն իմ կատարած երկու դիտողությունները (իսկ շատ այլ դիտողություններ անկասկած դեռ պետք է կատարել) որոշ լույս են սփռում այդ հարցի վրա։ Երբ բադերը արագ կերպով դուրս են գալիս լճակից, որը ծածկված է ջրոսպով, այդ բույսը, ինչպես ես նկատել եմ երկու անգամ, կպչում է նրանց մեջքին և պատահել է, որ ջրոսպը մի ակվարիումից մյուսը տեղափոխելով, առանց որևէ դիտավորության բազմացրել եմ նրանցից մեկում առաջինից վերցրած քաղցրահամ ջրերի խխունջներ։ Սակայն մյուս միջոցը թերևս ավելի իրական է. ես մտցրի բադի թաթիկները ակվարիումի մեջ, որտեղ կային քաղցրահամ ջրերի կակղամորթների շատ ձվեր և տեսա, որ որոշ քանակությամբ չափազանց մանր ձվեր և դեռ նոր դուրս եկած խխունջներ կպել են թաթիկներին և ամրացել են այնքան պինդ կերպով, որ նրանց դուրս հանելով ջրից, խխունջներին չէր կարելի թոթափել նրանց վրայից, թեև մի քիչ ավելի մեծ հասակի հասածները թափվում էին ազատ կերպով։ Այդպիսի դեռ նոր դուրս եկած խխունջները իրենց բնույթով ջրային կենդանիներ լինելով հանդերձ, ապրում էին բադի թաթիկների վրա՝ խոնավ մթնոլորտի մեջ, տասներկուսից մինչև քսան ժամ, այդ ժամանակի ընթացքում բադը կամ ձկնկուլը կարող է թռչել առնվազն վեց հարյուր կամ յոթ հարյուր մղոն և տարվելով քամու միջոցով ծովի վրայով դեպի մի օվկիանոսային կղզի կամ որևէ այլ հեռավոր կետ, անպայման կիջնի մի լճակի կամ գետակի վրա։ Սըր Չ. Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] հաղորդել է ինձ, որ բռնել է մի լողաբզեզ (Dytiscus), որին ամուր կերպով կպած է եղել մի Ancylus (ափսեի նմանվող քաղցրահամ ջրային խեցի), իսկ մի այլ ջրաբզեզ նույն ընտանիքից (Colymbetes) մի անգամ թռավ «Բիգլի» տախտակամածի վրա ամենամոտ երկրից քառասուն և հինգ մղոն հեռավորությամբ և ոչ ոք չի կարող ասել, թե բարենպաստ քամու դեպքում ինչքան հեռու կարող էր նա սլանալ։
Բույսերի վերաբերմամբ վաղուց արդեն հայտնի է, թե ինչպիսի հսկայական մարզեր են գրավում քաղցրահամ ջրերի և նույնիսկ ճահճային շատ տեսակներ ինչպես մայր ցամաքների, այնպես էլ ամենահեռավոր օվկիանոսային կղզիների վրա։ Ըստ Ա. դը-Կանդոլի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] այդ առանձնապես ցայտունորեն է արտահայտվում ցամաքային բույսերի այն մեծ խմբերում, որոնք պարփակում են ջրային շատ քիչ ներկայացուցիչներ, որովհետև վերջիններս կարծես հենց այդ պատճառով, անհապաղ գրավում են ընդարձակ մարզ։ Ես կարծում եմ, որ դա բացատրվում է տարաբնակման բարենպաստ եղանակներով։ Առաջները ես արդեն հիշատակել եմ, որ փոքր քանակությամբ հող պատահաբար կպչում է թռչունների ոտներին ու կտուցներին։ Լճակների տղմոտ եզերքները հաճախ այցելող բարձրասրունք թռչունները, երբ նրանց խրտնեցնում են, ամենայն հավանականությամբ ոտների վրա տիղմ են տանում։ Այդ խմբի թռչունները մյուսներից ավելի շատ են թափառում և հանդիպում են երբեմն բաց օվկիանոսի մեջ չափազանց հեռավոր և անպտուղ կղզիների վրա, հազիվ թե նրանք իջնում են ծովի մակերևույթի վրա և այսպիսով, նրանց ոտներին կպած տիղմը չի լվացվի, իսկ հասնելով ցամաքին նրանք անկասկած կգնան իրենց բնական քաղցրահամ ջրերի ապաստարանը։ Ես չեմ կարծում, որ բուսաբանները պարզ պատկերացում ունեն այն մասին, թե լճակների տիղմը ինչքան հարուստ է սերմերով։ Ես մի քանի փոքրիկ փորձեր եմ կատարել այդ ուղղությամբ, բայց այստեղ կհաղորդեմ ամենահետաքրքիր դեպքի մասին. փետրվարին ես վերցրի կերակրի երեք գդալ տիղմ ջրի տակից, փոքրիկ լճակի եզրի երեք զանազան տեղերից. չորացնելուց հետո այդ ցեխը կշռում էր 6¾ ունցիա[72]. վեց ամիս շարունակ ես այդ ցեխը պահում էի աշխատասենյակում խուփի տակ, պոկելով ու հաշվելով յուրաքանչյուր ծլող բույսը։ Բույսերը պատկանում էին զանազան տեսակների, և նրանց ընդհանուր թիվը հասնում էր 537-ի. և այդ մի թեյի գավաթում տեղավորված ջրիկ տիղմի մեջ։ Ուշադրության առնելով այդպիսի փաստերը, ես կարծում եմ, որ բոլորովին անբացատրելի կլիներ, եթե ջրային թռչունները քաղցրահամ ջրերում ապրող բույսերի սերմերը չտարածեին շատ հեռավոր կետերում գտնվող լճակներում ու հոսանքներում։ Նույնն է կատարվում նաև քաղցրահամ ջրերում ապրող մի քանի մանր կենդանիների ձվերի հետ։
Հավանորեն նաև այլ մեզ անհայտ պայմաններ են մասնակցում այդպիսի դեպքերում։ Ես արդեն մատնանշել եմ, որ քաղցրահամ ջրերի ձկներն ուտում են որոշ բույսերի սերմեր, բայց շատերին դուրս են նետում կուլ տալուց հետո, նույնիսկ մանր ձկները կուլ են տալիս միջին մեծությամբ սերմեր, օրինակ դեղին սափորիկի, կամ Potamogeton։ Ձկնկուլները և այլ թռչուններ դարեդար ամեն օր ուտում են ձկներ։ Հետո նրանք օդն են բարձրանում և չվում են դեպի այլ ջրեր կամ քամու միջոցով տարվում են ծովի վրայով, իսկ մենք տեսանք, որ սերմերն իրենց ծլելու ընդունակությունը պահպանում են, երբ դուրս են շպրտվում փղձկոցի միջոցով կամ կղկղանքի մեջ նույնիսկ շատ ժամեր անցնելուց հետո։ Երբ ես ծանոթացա փարթամ Nelumbium ջրաշուշանի սերմերի խոշոր չափերի հետ և հիշեցի Ա. դը-Կանդոլի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] դիտողությունը այդ բույսի տարածման մասին, ես մտածեցի, որ նրա տարածման եղանակները պետք է անբացատրելի մնան, բայց Օդյուբոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 117] պնդում է, որ գտել է հարավային մեծ ջրաշուշանի (դոկտոր Հուկերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] կարծիքով, հավանորեն, Nelumbium luteum-ի) սերմերը ձկնկուլի ստամոքսում։ Քանի որ այդ թռչունը պետք է թռչի լիքը լցրած ստամոքսով դեպի հեռավոր ջրավազանները և հետո սկսի ուտել կշտացնող ձկնային ուտելիքը, ապա ես, ըստ անալոգիայի կարծում եմ, որ նա կարող է փղձկոցի հետ դուրս նետել նաև ծլելու ընդունակ սերմեր։
Տարածման զանազան եղանակները ուսումնասիրելով, պետք է հիշել, որ երբ լճակը կամ հոսանքը դեռ նոր է գոյանում, օրինակ, մի բարձրացող կղզյակի վրա, նրանք դեռ ոչնչով չեն գրավված, և սերմը կամ ձուն հաջողությամբ զարգանալու շատ շանսեր ունեն։ Թեև գոյության կռիվը միշտ է տեղի ունենում լճակի բնակիչների միջև, ինչքան էլ նրանք քիչ լինեն, սակայն քանի որ տեսակների թիվը նույնիսկ լավ բնակված լճակում մեծ չէ այն տեսակների թվի համեմատությամբ, որոնք ապրում են ցամաքի համապատասխան տարածության վրա, ուստի մրցումը նրանց միջև հավանորեն ավելի թույլ է, քան ցամաքային տեսակների միջև՝ դրա հիման վրա օտար երկրի ջրերից գաղթողը նոր տեղ ձեռք բերելու ավելի մեծ շանսեր ունի, քան ցամաքային գաղթականները։ Մենք պետք է նաև հիշենք, որ քաղցրահամ ջրերի շատ օրգանիզմներ բնության սանդուխքի վրա ցածր տեղ են գրավում, և մենք հիմք ունենք կարծելու, որ այդպիսի էակները բարձրակարգներից ավելի դանդաղ են փոփոխվում, որը ժամանակ է տալիս ջրային տեսակների տարաբնակման համար։ Բացի դրանից, մենք չպետք է մոռանանք և այն հավանականությունը, որ քաղցրահամ ջրերի շատ տեսակներ առաջները անընդհատ կերպով տարածված են եղել հսկայական տարածության վրա, բայց հետո մահաջնջվել են միջակա կետերում։ Սակայն քաղցրահամ ջրերի բույսերը և ստորակարգ կենդանիները կպահպանե՞ն արդյոք իրենց նույնական ձևը կամ թե որոշ չափով փոփոխված կլինե՞ն, այդ միևնույն է, նրանց լայն տարածումը գլխավորապես կախված է սերմերի և ձվերի լայն տարածումից, որը կատարվում է կենդանիների, առանձնապես քաղցրահամ ջրերում ապրող թռչունների միջոցով, որոնք օժտված են թռչելու ընդունակությամբ, բնականաբար թափառում են մի ջրավազանից դեպի մյուսը։
ՕՎԿԻԱՆՈՍԱՅԻՆ ԿՂԶԻՆԵՐԻ ԲՆԱԿԻՉՆԵՐԸ
Այժմ մենք մոտենում ենք այն երեք կարգի փաստերից վերջինին, որոնք ես ընտրել եմ որպես օրգանիզմների տարածումը բացատրելու նկատմամբ ամենամեծ դժվարությունները ներկայացնողներ, երբ ընդունում ենք, որ ոչ միայն մի տեսակի բոլոր անհատներն են տարածվել մի մարզից, այլև մերձավոր, թեկուզ այժմ իրարից շատ հեռու կետերում ապրող տեսակները ծագել են մի մարզից՝ իրենց հեռավոր նախնիների հայրենիքից։ Ես արդեն բերել եմ իմ ապացույցները հերքելու համար այն կարծիքը, որ իբրև թե մայր ցամաքներն այժմյան տեսակների գոյության ժամանակաշրջանի ընթացքում հասել են այնպիսի հսկայական չափերի, որ բոլոր օվկիանոսային կղզիներն ստացել են իրենց այժմյան ցամաքային բնակիչները, կազմելով այդ մայր ցամաքների մասերը։ Այդ տեսակետը վերացնում է շատ դժվարություններ, բայց չի համապատասխանում այն բոլոր փաստերին, որոնք վերաբերում են կղզիների վրա ապրող օրգանիզմներին։ Հաջորդ դիտողություններում ես կանգ չեմ առնի բացառապես տարածման հարցի վրա, այլև կքննարկեմ մի քանի այլ դեպքեր, որոնք վերաբերում են երկու տեսությունների, այն է՝ անկախ ստեղծագործության և փոփոխության միջոցով առաջանալու տեսությունների ճշտության պարզաբանությանը։
Օվկիանոսային կղզիների վրա բնակվող զանազան էակների տեսակները սակավաթիվ են այն տեսակների համեմատությամբ, որոնք գրավում են ցամաքի հավասար տարածություն. Ա. դը-Կանդոլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] այդ ընդունում է բույսերի համար, Ուոլլաստոնը՝[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 65] միջատների համար։ Նոր Զելանդիան, օրինակ, իր բարձր լեռներով և բազմազան բնակավայրերով, հասնելով 780 մղոնից ավելի լայնության, ունի Աուկլենդյան, Կեմպբել և Չատամ կղզիների հետ միասին ընդամենը ծաղկավոր բույսերի 960 տեսակ. եթե մենք այս համեստ թիվը համեմատենք այն տեսակների թվի հետ, որոնք աճում են նույնքան տարածության վրա հարավ-արևմտյան Ավստրալիայում կամ Բարեհուսո հրվանդանի վրա, ապա մենք պետք է ընդունենք, որ թվի մեջ եղած մեծ տարբերությունը բացատրվում է ինչ որ պատճառով՝ անկախ ֆիզիկական պայմաններում եղած տարբերությունից։ Նույնիսկ Քեմբրիջի միակերպ կոմսությունն ունի 847 բույս, իսկ Էնգլեսի փոքրիկ կղզին՝ 764. բայց այդ թվերի մեջ մտնում են մի քանի պտերներ և նատուրալիզացիայի ենթարկված բույսեր, և համեմատությունը այնքան էլ ճիշտ չէ մի քանի այլ տեսակետներից։ Մենք ապացույց ունենք, որ Համբարձման անբնակ կղզին սկզբնապես ունեցել է կես դյուժինից պակաս ծաղկավոր բույսեր․ ներկայումս բուսական շատ տեսակներ նատուրալիզացիայի են ենթարկվել նրա վրա, ինչպես որ Նոր Զելանդիայի և օվկիանոսային կղզիներից ամեն մեկի վրա։ Հիմք կա կարծելու, որ Սուրբ Հեղինեի կղզին բերված բույսերն ու կենդանիները գրեթե կամ լիովին բնաջնջել են բնական շատ արտադրանքներ։ Նա, ով ընդունում է յուրաքանչյուր տեսակի առանձին ստեղծման ուսմունքը, պետք է նաև ընդունի, որ օվկիանոսային կղզիների համար մեծ թվով լավագույն կերպով հարմարված բույսեր ու կենդանիներ չեն ստեղծվել, որովհետև մարդը չնախամտածված կերպով դրանք բնակեցրել է շատ ավելի լրիվ և ավելի լավ, քան այդ արել է բնությունը։
Թեև օվկիանոսային կղզիների վրա տեսակների ընդհանուր թիվը քիչ է, բայց էնդեմիկ, այսինքն երկրագնդի վրա ուրիշ ոչ մի տեղ չհանդիպող, ձևերի քանակությունը հաճախ շատ մեծ է լինում։ Դրա ճշտությանը կհամոզվենք, եթե օրինակի համար համեմատենք Մադեյրա կղզու էնդեմիկ ցամաքային կակղամորթների կամ Գալապագոսի արշիպելագի էնդեմիկ թռչունների քանակը որևէ մայր ցամաքի նույն կենդանիների քանակության հետ, իհարկե եթե կղզիների տարածությունը հավասար է մայր ցամաքի տարածությանը։ Այդ կարելի է սպասել նաև տեսական կշռադատումների հիման վրա, որովհետև, ինչպես արդեն բացատրվել է, այն տեսակները, որոնք մեծ ժամանակամիջոցներից հետո պատահմամբ ընկնում են նոր և առանձնացած մարզերի մեջ և դրա հետ միասին մտնում են մրցության նոր պայմանների մեջ, անկասկած կենթարկվեն փոփոխության և շատ դեպքերում կտան փոփոխված հաջորդների խմբեր։ Բայց դրանից բնավ չի հետևում, որ եթե որևէ կղզու վրա մի դասի համարյա բոլոր տեսակները բնորոշ են այդ կղզու համար, ապա մյուս դասի տեսակները կամ առաջինի ստորաբաժանումներն էլ բնորոշ կլինեն. այդ տարբերությունը մասամբ կախված է նրանից, որ տեսակները չեն փոփոխվում, եթե մեծ թվով են ընկնում կղզին, որովհետև այդ դեպքում նրանց փոխհարաբերությունները որքան և իցե մեծ չափով չեն խախտվում, մասամբ էլ նրանից, որ չփոփոխված գաղթականները հաճախ կարող են կղզին ընկնել հիմնական մարզից, և հետո խաչաձևվել կղզու ձևերի հետ։ Կարելի է կարծել, որ այդպիսի խաչաձևումներից առաջացած հաջորդները զգալի չափով շահում են ուժի տեսակետից, և այսպիսով նույնիսկ պատահական խաչաձևումը ավելի մեծ նշանակություն կունենա, քան այդ կարելի էր ենթադրել։ Ասածիս օգտին ես կբերեմ մի քանի օրինակներ։ Գալապագոսյան կղզիների վրա կան ընդամենը 26 ցամաքային թռչուններ, որոնցից քսանմեկը կամ գուցե նույնիսկ քսաներեքը հատուկ են միայն այդ կղզիներին, մինչդեռ տասնմեկ ծովային թռչուններից էնդեմիկ են միայն երկուսը․ ակնհայտ է, որ ծովային թռչունները կարող են այդ կղզիներն ընկնել թե՛ ավելի հեշտ կերպով և թե՛ ավելի հաճախ, քան ցամաքայինները։ Մյուս կողմից, Բերմուդյան կղզիները, որոնք նույնպիսի հեռավորության վրա են գտնվում Հյուսիսային Ամերիկայից, ինչպիսի հեռավորության վրա են Գալապագոսյան կղզիները Հարավային Ամերիկայից, և ունեն չափազանց յուրահատուկ հող, չունեն և ոչ մի էնդեմիկ ցամաքային թռչուն, բայց մենք գիտենք մ-ր Ջ. Մ. Ջոնսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 245] Բերմուդյան կղզիների մասին գրած հիանալի նկարագրությունից, որ հյուսիս-ամերիկական շատ ու շատ թռչուններ ժամանակ առ ժամանակ և նույնիսկ համեմատաբար հաճախակի այցելում են այդ կղզիները։ Գրեթե յուրաքանչյուր տարի, ինչպես հաղորդում է ինձ մ-ր Է. Վ. Հարկուրտը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 235] շատ եվրոպական և աֆրիկական թռչուններ քամու միջոցով տարվում են Մադեյրա կղզին. այդ կղզու վրա բնակվում են իննսունինը տեսակներ, որոնցից միայն մեկն է հատուկ նրան, այն էլ շատ մերձավոր եվրոպական ձևին, և երեք կամ չորս այլ տեսակներ պատկանում են այդ կղզուն և Կանարյան կղզիներին։ Սրանից հետևում է, որ Բերմուդների և Մադեյրայի վրա բնակություն են հաստատել հարևան մայր ցամաքներից եկած թռչունները, որոնք երկար ժամանակ իրենց գոյության համար պայքարել են իրար հետ և հարմարվել մեկը մյուսին։ Դրա շնորհիվ յուրաքանչյուր տեսակ ընկնելով մի նոր մարզ, մյուսների կողմից կասեցվում է իր տեղում և իր սովորություններով և այդ պատճառով միայն քիչ չափով է հակված լինում փոփոխվելու։ Բացի դրանից, փոփոխվելու ամեն հակում ճնշվում է չփոփոխված գաղթականների հետ խաչաձևվելով, որոնք հաճախ գալիս են հիմնական մարզից։ Մադեյրայի վրա բնակվում են նաև զարմանալի մեծ թվով բնորոշ ցամաքային խեցիներ, մինչդեռ նրա ափերին չկա ոչ մի բնորոշ ծովային խեցի։ Թեև մենք չգիտենք, թե ինչպես են տարածվում ծովային խեցիները, սակայն կարող ենք հասկանալ, որ նրանց ձվերն ու թրթուրները կպչելով գուցե ջրիմուռներին կամ լողացող ծառերին կամ բարձրասրունք թռչունների ոտներին, կարող են ավելի հեշտությամբ տեղափոխվել բաց ծովի վրայով երեք կամ չորս հարյուր մղոն հեռավորության վրա, քան ցամաքային խեցիները։ Մադեյրայի վրա ապրող միջատների զանազան խմբերը ներկայացնում են գրեթե զուգահեռ դեպքեր։
Օվկիանոսային կղզիները երբեմն զուրկ են լինում որոշ դասերի կենդանիներից, ապա այդպիսի դեպքում նրանց տեղը գրավում են այլ դասերի ներկայացուցիչները. այսպես, Գալապագոսյան կղզիների վրա սողունները, Նոր Զելանդիայի վրա հսկա անթև թռչունները գրավում են կամ ոչ այնքան վաղուց գրավում էին կաթնասունների տեղը։ Թեև մենք այստեղ խոսում ենք Նոր Զելանդիայի մասին որպես օվկիանոսային մի կղզու մասին, բայց որոշ չափով կասկածելի է համարել նրան այդպիսին, որովհետև, մի կողմից, նա չափազանց մեծ է և Ավստրալիայից բաժանված չէ շատ խոր ծովով, մյուս կողմից, նրա լեռնաշղթաների երկրաբանական բնույթի և ուղղության հիման վրա հարգարժան Վ. Բ. Կլարկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 240] վերջերս հայտնել է այն կարծիքը, որ այդ կղզին, ինչպես նաև Նոր Կալեդոնիան, կարելի է համարել Ավստրալիային պատկանող։ Դառնալով բույսերին, մենք տեսնում ենք, որ ըստ Հուկերի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] Գալապագոսյան կղզիների վրա զանազան կարգերի հարաբերական թիվը բոլորովին տարբեր է նրանից, ինչ նկատվում է մի որևէ այլ տեղ։ Թվի բոլոր նման տարբերությունները և կենդանիների ու բույսերի որոշ խմբերի լիակատար բացակայությունը սովորաբար բացատրվում են ենթադրելի տարբերությամբ կղզիների ֆիզիկական պայմանների մեջ. բայց այդպիսի բացատրությունը շատ կասկածելի է։ Այն դյուրությունը, որով տեղի է ունենում նրանց վրա բնակություն հաստատելը, ըստ երևույթին, նույնքան մեծ նշանակություն ունի, ինչքան որ այդ կղզիների ներկայացրած պայմանների բնույթը։
Օվկիանոսային կղզիների բնակիչների վերաբերմամբ կարելի է հաղորդել բազմաթիվ ուշագրավ մանր փաստեր։ Օրինակ, մի քանի կղզիների վրա, որտեղ ոչ մի կաթնասուն չկա, էնդեմիկ բույսերից մի քանիսն ունեն հիանալի զարգացած կեռիկներով սերմեր, սակայն հազիվ թե որևէ փոխհարաբերություն այնքան ակնառու է, ինչպես այն, որ կեռիկները ծառայում են կաթնասունների մազերին կպչելու միջոցով սերմերը տարածելու համար։ Բայց այդպիսի սերմերը կարող էին ընկնել կղզու վրա նաև այլ եղանակով, իսկ բույսը փոփոխվելով ու դառնալով էնդեմիկ տեսակ, շարունակում է պահպանել կեռիկները, որոնք այժմ անօգուտ հավելվածների նշանակություն ունեն, նման կղզու շատ բզեզների վերնաթևերի տակ կծկված ետևի թևերին։ Այնուհետև կղզիների վրա հաճախ հանդիպում են այնպիսի ընտանիքների պատկանող ծառեր և թփուտներ, որոնք այլ տեղերում կազմված են միայն խոտանման տեսակներից. բայց ծառերը, ինչպես այդ ցույց է տվել Ալֆոնս դը-Կանդոլը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] ընդհանրապես ունեն, ինչ էլ ուզում է լինի դրա պատճառը, սահմանափակ տարածում, և դրա համար էլ քիչ հավանական է, որ ծառերը կարող էին իրարից հեռու գտնվող օվկիանոսային կղզիները հասնել։ Բայց այն խոտանման բույսը, որը շանսեր չունի հաջողությամբ մրցելու մայր ցամաքի վրա բուսնող լավ զարգացած շատ ծառերի հետ, հաստատվելով կղզու վրա կարող է ձեռք բերել առավելություններ այլ խոտանման բույսերի հանդեպ, շարունակ աճելով բարձր ու բարձր և ծավալելով իր գագաթը նրանց վրա։ Այս դեպքում բնական ընտրությունը ձգտում է բարձրացնել բույսի հասակը, ինչ ընտանիքի էլ նա պատկանի, և սկզբում դարձնում է նրան թփուտ, իսկ հետո ծառ։
ԵՐԿԿԵՆՑԱՂՆԵՐԻ ԵՎ ՑԱՄԱՔԱՅԻՆ ԿԱԹՆԱՍՈՒՆՆԵՐԻ ԲԱՑԱԿԱՅՈՒԹՅՈՒՆԸ ՕՎԿԻԱՆՈՍԱՅԻՆ ԿՂԶԻՆԵՐԻ ՎՐԱ
Օվկիանոսային կղզիների վրա կենդանիների ամբողջ կարգերի բացակայության մասին Բորի Սեն-Վենսենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 246] դեռ վաղուց նշել է, որ երկկենցաղները (գորտերը, դոդոշները, տրիտոնները) երբեք չեն գտնվել այն կղզիներից ոչ մեկի վրա, որոնք ցրված են օվկիանոսի լայնածավալ տարածության մեջ։ Ես աշխատեցի ստուգել այդ ցուցմունքը և գտա, որ նա միանգամայն ճիշտ է, բացառելով Նոր Զելանդիան, Նոր Կալեդոնիան, Անդամանյան և գուցե Սողոմոնյան ու Սեյշելյան կղզիները։ Բայց ես արդեն նշել եմ, որ կասկածելի է, թե արդյոք կարելի է Նոր Զելանդիան և Նոր Կալեդոնիան տեղավորել օվկիանոսային կղզիների թվում, և ավելի ևս կասկածելի նույնը Անդամանյան ու Սողոմոնյան խմբերի և Սեյշելյան կղզիների վերաբերմամբ։ Գորտերի, դոդոշների և տրիտոնների այդ ընդհանուր բացակայությունը անկասկածելիորեն օվկիանոսային շատ կղզիների վրա չի կարելի բացատրել վերջիններիս ֆիզիկական պայմաններով․ ընդհակառակր, թվում է, թե այդ կղզիներն առանձնապես հարմար են հիշված կենդանիների համար, որովհետև գորտերը բերվել են Մադեյրա, Ազորյան և Մավրիկիա կղզիները և այնտեղ այնքան են բազմացել, որ դարձել են վնասատուներ։ Քանի որ այդ կենդանիներն ու նրանց ձվերն անմիջապես փչանում են ծովի ջրում (որքան գիտենք՝ բացառությամբ մի հնդկական տեսակի) ուստի նրանց համար չափազանց դժվար է անցնել ծովի վրայով, և դա բացատրում է մեզ, թե ինչու նրանք բացակայում են իսկական օվկիանոսային կղզիների վրա։ Բայց թե ինչու նրանք, եթե հետևենք ստեղծագործության տեսությանը, այստեղ չեն ստեղծվել, դա բացատրել շատ դժվար է։
Կաթնասուններն իրենց կողմից ներկայացնում են նման դեպքեր։ Ես ուշադիր կերպով կարդացել եմ հին ճանապարհորդություններ և չեմ գտել ոչ մի անկասկածելի ցուցմունք այն մասին, որ մի ցամաքային կաթնասուն (բացառությամբ բնիկների բուծած ընտանի կենդանիների) ողջ վիճակում գտնված լինի այնպիսի կղզու վրա, որը 300 մղոնից ավելի հեռու է մայր ցամաքից կամ մայր ցամաքային մի մեծ կղզուց․ իսկ նույնիսկ անհամեմատ ավելի մոտ գտնվող շատ կղզիներ միանգամ այն անբնակ են։ Ֆալկլանդյան կղզիները, որոնց վրա ապրում է գայլի նմանությամբ մի աղվես, ավելի շուտ բացառության են համապատասխանում․ բայց կղզիների այդ խումբը չի կարելի դիտել որպես օվկիանոսային, որովհետև նա գտնվում է ծանծաղուտի վրա, որը միացնում է նրան մոտավորապես 280 մղոն հեռավորության վրա ընկած մայր ցամաքի հետ, բացի դրանից, լողացող սառույցները նախկին ժամանակները գլաքարեր են բերել այդ կղզիների արևմտյան ափերը և կարող էին տանել նաև աղվեսին, ինչպես այդ այժմ հաճախ լինում է արկտիկական երկրներում։ Չի կարելի ասել նաև այն, թե փոքր կղզին չի կարող կերակրել գոնե փոքրիկ կաթնասուններին, որովհետև այդպիսիները հանդիպում են աշխարհի շատ մասերում շատ փոքրիկ կղզիների վրա, եթե այդպիսի կղզիները հեռու չեն մայր ցամաքից, և հազիվ թե կարելի է անվանել մի կղզի, որի վրա մեր մանր չորքոտանիները չենթարկվեին նատուրալիզացիայի և չբազմանային։ Անկախ ստեղծագործություն ընդունող սովորական տեսակետի համաձայն չի կարելի ասել, որ այդ դեպքերում ժամանակ չի եղել կաթնասուններ ստեղծելու համար․ հրաբխային շատ կղզիներ բավականաչափ հին են, ինչպես այդ կարելի է տեսնել նրանց կրած հսկայական քայքայումից և նրանց երրորդային շերտերից․ այստեղ նաև ժամանակ եղել է այլ դասերի էնդեմիկ տեսակներ ստեղծելու համար, իսկ մայր ցամաքների մասին հայտնի է, որ կաթնասունների նոր տեսակներն ավելի արագ են երևան գալիս ու անհետանում, քան մյուսները, ցածրակարգ կենդանիները։ Թեև ցամաքային կաթնասունները չեն հանդիպում օվկիանոսային կղզիների վրա, բայց օդայինները հանդիպում են համարյա յուրաքանչյուր կղզու վրա։ Նոր Զելանդիայի վրա կան երկու տեսակ չղջիկներ, որոնք երկրագնդի վրա ուրիշ ոչ մի տեղ չկան. Նորֆոլկ կղզին, Վիտի արշիպելագը, Բոնին կղզիները, Կարոլինյան և Մարիանյան արշիպելագները և Մավրիկիա կղզին՝ բոլորն էլ ունեն իրենց բնորոշ չղջիկները։ Իսկ ինչո՞ւ, կարելի է հարցնել, ենթադրյալ ստեղծագործական գործունեությունը առաջացրել է հեռավոր կղզիների վրա չղջիկներ, այլ ոչ թե ուրիշ կաթնասուններ։ Իմ կարծիքով այդ հարցին հեշտ է պատասխանել, որովհետև ոչ մի ցամաքային կենդանի չի կարող անցնել ծովի ընդարձակ տարածությունը, իսկ չղջիկները կարող են նրա վրայով թռչել։ Այդ կենդանիներին տեսել են ցերեկը Ատլանտյան օվկիանոսի վրայով հեռու թռչելիս, հյուսիս-ամերիկական երկու տեսակներ կանոնավոր կերպով կամ ժամանակ առ ժամանակ այցելում են Բերմուդները, որոնք գտնվում են մայր ցամաքից 600 մղոն հեռու։ Ես իմանում եմ մ-ր Տոմսից,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 247] որը հատկապես ուսումնասիրել է այդ ընտանիքը, որ շատ տեսակներ ունեն տարածման հսկայական շրջան և գտնված են թե՛ մայր ցամաքների և թե՛ հեռավոր կղզիների վրա։ Այստեղից մեզ մնում է միայն ենթադրել, որ այդպիսի թափառող տեսակը փոփոխվել է իր բնակավայրի նոր հանգամանքներին հարմարվելով, և մեզ հասկանալի կդառնա էնդեմիկ չղջիկների ներկայությունը օվկիանոսային կղզիների վրա, երբ բացակայում են մյուս բոլոր ցամաքային կաթնասունները։
Կա ևս մի հետաքրքրական հարաբերակցություն, այն է՝ կղզիները միմյանցից կամ ամենամոտ մայր ցամաքից բաժանող ծովի խորության և նրանց կաթնասունների ունեցած նմանության աստիճանի միջև։ Մ-ր Ուինդզոր Իրլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 248] այդ տեսակետից կատարել է մի քանի կարևոր դիտողություններ, որոնք դրանից հետո զգալիորեն ընդարձակվել են մ-ր Ուոլլեսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] հիանալի հետազոտություններով Մալայան արշիպելագի վրա, որը Ցելեբեսի մոտերքը խոր օվկիանոսի մի շերտով բաժանվում է կաթնասունների երկու խիստ տարբեր կենդանական աշխարհների։ Նեղուցի յուրաքանչյուր կողմում կղզիները տեղավորված են համեմատաբար ոչ խոր ստորջրյա հիմքի վրա և բնակված են միևնույն կամ շատ մերձավոր կաթնասուններով։ Ես դեռ ժամանակ չեմ ունեցել հետամուտ լինելու այդ հարաբերակցությանը աշխարհի բոլոր երկրներում, բայց որքան ես այդ կարողացել եմ անել, այդպիսի հարաբերակցություն կա։ Օրինակ, Բրիտանիան Եվրոպայից բաժանված է մի նեղ ջրանցքով, որի երկու կողմում ապրում են միևնույն կաթնասունները. նույնը ճիշտ է նաև Ավստրալիայի ափերի շուրջը գտնվող բոլոր կղզիների վերաբերմամբ։ Մյուս կողմից, Վեստ Ինդիայի կղզիները, որոնք նստած են ստորջրյա խոր, մոտավորապես 1000 ֆաթոմ խորությամբ, հիմքի վրա, բնակված են միանգամայն տարբեր տեսակների և նույնիսկ սեռերի պատկանող ամերիկական ձևերով։ Քանի որ այն փոփոխությունների չափը, որոնց ենթարկվում են զանազան կենդանիներ, մասամբ կախված է ժամանակամիջոցից, իսկ կղզիները, որոնք մայր ցամաքից կամ մեկը մյուսից բաժանված են ծանծաղ ջրանցքով, ամենայն հավանականությամբ միացած են եղել ավելի նոր ժամանակաշրջանում, քան խոր ջրանցքներով բաժանված կղզիները, ապա մենք կարող ենք հասկանալ, թե ինչպիսի հարաբերակցություն գոյություն ունի կաթնասունների երկու ֆաունան բաժանող խոր ծովի և նրանց նմանության աստիճանի միջև, մի հարաբերակցություն, որը միանգամայն անբացատրելի է անկախ ստեղծագործական ակտերի տեսությամբ։
Օվկիանոսային կղզիների բնակիչների մասին արված դիտողությունները, այն է՝ որ նրանք ունեն տեսակների սահմանափակ թիվ, բայց մեծ քանակությամբ էնդեմիկ ձևեր, միևնույն դասի որոշ խմբերի անդամները փոփոխվել են, մյուսներինը՝ ոչ, ամբողջ կարգերի բացակայություն, ինչպիսիք են երկկենցաղները և ցամաքային կաթնասունները, չնայած չղջիկների ներկայությանը, բույսերի որոշ ընտանիքների յուրահատուկ զարգացում (խոտանման ձևերի զարգանալն ու ծառաբույսեր դառնալը) և այլն, այս բոլորը, ինձ թվում է, ավելի լավ է համաձայնվում այն ենթադրության հետ, որ երկար ժամանակաշրջանի ընթացքում տեղի են ունեցել տեղափոխության պատահական եղանակներ, քան այն ենթադրությանը, թե բոլոր օվկիանոսային կղզիները առաջներում կապված են եղել մայր ցամաքի հետ։ Վերջին ենթադրության դեպքում զանազան դասերի ներկայացուցիչներ հավանորեն կգաղթեին ավելի հավասարաչափորեն, իսկ քանի որ մեծ քանակությամբ գաղթելիս նրանց փոխհարաբերությունները զգալի չափով չէին խախտվի, ապա սրանից հետևում է, որ նրանք կամ բոլորովին չէին փոփոխվի, կամ էլ բոլոր տեսակները կփոփոխվեին միանման չափով։
Ես չեմ ժխտում, որ շատ լուրջ դժվարություններ կան հասկանալու համար այն, թե ինչ կերպով ամենահեռավոր կղզիների բնակիչներից շատերը՝ կամ նախկին տեսակային ձևը պահպանողները, կամ արդեն փոփոխվածները հասել են իրենց այժմյան բնակավայրերը։ Բայց չպետք է մոռանալ այլ կղզիների գոյության հավանականությունը, որոնք մի ժամանակ կայանների դեր են կատարել և բոլորովին անհետ կորել են։ Ես ուզում եմ պարզաբանել մի դժվարին դեպք։ Գրեթե բոլոր օվկիանոսային կղզիները, նույնիսկ ամենից ավելի մեկուսացածները և փոքրագույն մեծություն ունեցողները բնակված են ցամաքային կակղամորթներով, սովորաբար էնդեմիկ տեսակներով, երբեմն ամենուրեք տարածված տեսակներով, որի զարմանալի օրինակները Խաղաղ օվկիանոսի համար տվել է դոկտոր Ա. Ա. Գուլդը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 249] Միաժամանակ պետք է նկատել, որ ծովի ջուրը հեշտությամբ սպանում է ցամաքային կակղամորթներին․ նրանց ձվերը, գոնե փորձի համար իմ վերցրածները, սուզվում են նրա մեջ և փչանում են։ Հետևաբար, պետք է լինեն որևէ անհայտ, բայց առիթի դեպքում իրական միջոցներ նրանց տեղափոխության համար։ Արդյոք չե՞ն կարող երբեմն հենց նոր ծնված անհատները կպչել գետնի վրա ման եկող թռչունների ոտներին և տեղափոխվել նրանց միջոցով։ Ինձ թվացել է նաև այն, որ ձմեռող կակղամորթները խեցու անցքը թաղանթով ծածկված վիճակում կարող են տեղափոխվել ոչ այնքան լայն ծովային նեղուցով լողացող ծառերի ճեղքերում։ Ես գտել եմ, որ այդ վիճակում զանազան տեսակներ առանց վնասի 7 օր դիմանում են ծովի ջրի մեջ սուզվելուն. իսկ Helix pomatia-ի մի խեցի նման մի փորձից հետո ձմեռային հանգստի միևնույն վիճակում իջեցվել է քսան օրով ծովի ջրի մեջ և հիանալի կերպով դիմացել է այդ ստուգմանը։ Իսկ այդ ժամանակամիջոցի ընթացքում խեցին կարող է միջին արագություն ունեցող ծովային հոսանքով տեղափոխվել 660 աշխարհագրական մղոն հեռավորության վրա։ Քանի որ Helix-ը հաստ կրային կափարիչ ունի, ես հեռացրի այն, իսկ երբ գոյացավ նոր, թաղանթանման ծածկոցը, ես նրան տասնչորս օրով նորից սուզեցի ծովային ջրի մեջ, որից հետո նա կրկին կազդուրվեց և կարող էր սողալ։ Ինձնից հետո նման փորձեր կատարել է բարոն Աուկեպտենը.[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 250] նա զետեղել է տասը տեսակների պատկանող հարյուր ցամաքային կակղամորթներ ծակոտկեն պատերով արկղի մեջ և այն սուզել է երկու շաբաթով ծովի մեջ։ Հարյուրից կենդանի են մնացել քսանյոթը։ Կափարիչի ներկայությունը, ամենայն հավանականությամբ, այդ դեպքում նշանակություն ունի, որովհետև այդ կափարիչն ունեցող Cyclostoma elegans-ի տասներկու անհատներից տասնմեկը կենդանի են մնացել։ Ուշագրավ է այն, որ այն ժամանակ, երբ իմ փորձի համար վերցրած Helix pomatia-ն լավ դիմադրել է ծովի ջրի ազդեցությանը, Աուկեպտենի վերցրած Helix-ի այլ չորս տեսակներին պատկանող հիսունչորս անհատներից ոչ մեկը չի դիմացել ստուգմանը։ Սակայն ամենևին չի կարելի հավանական համարել, որ ցամաքային կակղամորթները հաճախակի են փոխադրվել այդ ձևով. թռչունների ոտները հանդիսանում են ամենահավանական եղանակը։
ԿՂԶԻՆԵՐԻ ԲՆԱԿԻՉՆԵՐԻ ԱՌՆՉՈՒԹՅՈՒՆԸ ԱՄԵՆԱՄԵՐՁԱՎՈՐ ՄԱՅՐ ՑԱՄԱՔԻ ԲՆԱԿԻՉՆԵՐԻ ՀԵՏ
Ամենից ավելի զարմանալի և մեզ համար կարևոր փաստ է ներկայացնում կղզիների վրա բնակվող տեսակների նմանությունը ամենամոտ մայր ցամաքի տեսակների հետ՝ չնայած նրանց միջև նույնություն չկա։ Դա կարելի է հաստատել բազմաթիվ օրինակներով։ Հասարակածի մոտերքն ընկած Գալապագոսյան արշիպելագը գտնվում է Հարավային Ամերիկայի ափերից 500—600 մղոն հեռավորության վրա, և նրա ցամաքի կամ ջրերի գրեթե յուրաքանչյուր արտադրանք իր վրա կրում է Ամերիկայի մայր ցամաքի անկասկածելի դրոշմը։ Այնտեղ բնակվում են քսանվեց ցամաքային թռչուններ, որոնցից քսանմեկը կամ գուցե քսաներեքը համարվում են լավ առանձնացած տեսակներ և ընդհանրապես կարող էին դիտվել որպես այստեղ ծնվածներ, սակայն այդ թռչունների մեծամասնության մոտ ազգակցությունը ամերիկական տեսակների հետ արտահայտվում է ամեն բանում՝ սովորությունների, շարժումների, ձայնի մեջ։ Նույն բանն է նաև թե այլ կենդանիների և թե մեծ քանակությամբ բույսերի նկատմամբ, ինչպես այդ ցույց է տվել դոկտոր Հուկերն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] իր Գալապագոսյան արշիպելագի բուսականության մասին գրած հիանալի աշխատության մեջ։ Մայր ցամաքից մի քանի հարյուր մղոն հեռու Խաղաղ օվկիանոսի մեջ ընկած այդ հրաբխային կղզիների բնակիչների հետ ծանոթացող բնախույզն զգում է այնպես, թե ինքը գտնվում է ամերիկական հողի վրա։ Ինչո՞ւ է այդ այդպես։ Ինչո՞ւ այն տեսակները, որոնք, ինչպես ենթադրում են, ստեղծված են Գալապագոսյան արշիպելագի վրա և ոչ մի այլ տեղ, կրում են Ամերիկայում ստեղծված տեսակների հետ նմանության այնպիսի անկասկածելի դրոշմ։ Կղզիների ո՛չ կենսապայմաններում, ո՛չ երկրաբանական կառուցվածքում, ո՛չ բարձրության և կլիմայի մեջ, ո՛չ էլ առանձին դասերի միմյանց հետ ունեցած հարաբերակցության մեջ ոչինչ չկա, որ շատ նման լինի Հարավային Ամերիկայի մայր ցամաքի պայմաններին. իրականում բոլոր տեսակետներից մեծ տարբերություն կա։ Մյուս կողմից, հողի հրաբխային բնույթով, կլիմայով, բարձրությամբ և մեծությամբ Գալապագոսյան արշիպելագի կղզիները մեծ նմանություն ունեն Կանաչ հրվանդանի կղզիների հետ. բայց ինչպիսի լիակատար ու բացարձակ տարբերություն կա նրանց բնակիչների միջև։ Կանաչ հրվանդանի կղզիների բնակիչները պատկանում են Աֆրիկայի բնակիչներին, ինչպես որ Գալապագոսյան բնակիչները ամերիկականներին։ Նման փաստերը բացատրվել չեն կարող անկախ ստեղծագործություն ընդունող տեսակետով, մինչդեռ այստեղ ընդունված տեսակետից ակներև է այն, որ Գալապագոսյան կղզիներն ամենայն հավանականությամբ իրենց գաղթականներն ստացել են Ամերիկայից կամ գաղթի պատահական եղանակի շնորհիվ, կամ նրանք մի ժամանակ միացած են եղել ցամաքի հետ առանց ընդհատման (թեև այս վերջինին ես չեմ հավատում), իսկ Կանաչ հրվանդանի կղզիները՝ Աֆրիկայից, այդպիսի գաղթականները կարող էին փոփոխություններ կրել, բայց ժառանգականության սկզբունքը դեռևս մատնանշում է նրանց ծագման վայրը։
Կարելի է համանման շատ փաստեր բերել, որոնց գրեթե ընդհանուր կանոնն այն է, որ կղզիների էնդեմիկ արտադրանքները պատկանում են ամենամոտ մայր ցամաքի կամ ամենամոտ ընդարձակ կղզու արտադրանքներին։ Բացառությունները քիչ են և նրանց մեծամասնությունը կարելի է բացատրել։ Այսպես, թեև Կերգուելեն կղզին ավելի մոտ է Աֆրիկային, քան Ամերիկային, սակայն նրա բույսերը նման են և այն էլ մեծ չափով, ինչպես մենք այդ գիտենք դ-ր Հուկերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] աշխատություններից, Ամերիկայի բույսերին. բայց ունենալով այն տեսակետը, որ այդ կղզին ամբողջովին բնակեցվել է գերազանցապես այն սերմերով, որ բերվել են իշխող հոսանքների հետ շարժվող լողացող սառցակույտերի վրա եղած հողն ու քարերը, այդ անոմալիան չքանում է։ Նոր Զելանդիան իր էնդեմիկ բույսերով ավելի մոտ է Ավստրալիային՝ ամենամերձավոր ընդարձակ ցամաքին, քան որևէ մի այլ երկրի, և հենց այդ էլ կարելի էր սպասել. բայց նա մոտ է նաև Հարավային Ամերիկային, որը թեև Ավստրալիայից հետո մյուս մայր ցամաքներից ամենամոտն է, սակայն այնքան հեռու է ընկած, որ այդ փաստը դառնում է անոմալիա։ Այդ դժվարությունը մասամբ վերանում է, երբ ընդունում ենք այն տեսակետը, որ Նոր Զելանդիայի, Հարավային Ամերիկայի և այլ հարավային երկրների բնակիչները մասամբ եկել են միջանկյալ, թեև հեռավոր մի կետից, այն է՝ անտարկտիկական կղզիներից, երբ նրանք երրորդային ավելի տաք ժամանակաշրջանի ընթացքում, սառցադաշտային ժամանակաշրջանի սկսվելուց առաջ ծածկված են եղել բուսականությամբ։ Այդ նմանությունը, որ թեև թույլ, բայց գոյություն ունի, ըստ դ-ր Հուկերի վկայության, Ավստրալիայի և արավ-արևմտյան անկյան և Բարեհուսո հրվանդանի բուսական աշխարհների միջև, առավել ևս ուշագրավ դեպք է ներկայացնում, այդ նմանությունը, որ արտահայտվում է միայն բույսերի նկատմամբ, անկասկած երբևիցե կբացատրվի։
Միևնույն օրենքը, որը որոշում է կղզիների և նրանց ամենամոտ ընդարձակ ցամաքի բնակիչների միջև եղած ազգակցությունը, երբեմն արտահայտվում է ավելի փոքր չափերով, բայց վերին աստիճանի հետաքրքրական ձևով միևնույն արշիպելագի սահմաններում։ Այսպես, Գալապագոսյան արշիպելագի կղզիներից ամեն մեկն ունի մի քանի առանձին տեսակներ, մի բան, որն ինքնին զարմանալի փաստ է ներկայացնում. այդ տեսակներն անհամեմատ ավելի մոտ են միմյանց, քան Ամերիկայի մայր ցամաքի կամ աշխարհի որևէ այլ երկրի բնակիչներին։ Այդ էլ կարելի էր սպասել, որովհետև այդպես իրար մոտ գտնվող կղզիները իրենց գաղթականներն անպայման պիտի ստանային մի ընդհանուր աղբյուրից և մեկը մյուսից։ Բայց ինչպե՞ս է պատահել, որ գաղթականներից շատերը տարբեր կերպով են փոփոխվել, թեև միայն թույլ չափով, այն կղզիների վրա, որոնք գտնվում են իրար մոտ, ունեն միևնույն երկրաբանական կառուցվածքը, միևնույն բարձրությունը, միևնույն կլիման և այլև։ Ինձ համար դա երկար ժամանակ մնում էր որպես մեծ դժվարություն, բայց այդ բացատրվում էր գլխավորապես ամուր կերպով արմատացած այն մոլորությամբ, որ ֆիզիկական պայմաններն են համարվում ամենից ավելի կարևոր, մինչդեռ չի կարելի վիճել, որ այն տեսակների հատկությունները, որոնց հետ ամեն մեկը հարկադրված է մրցման մեջ մտնելու, ներկայացնում են հաջողության համար առնվազն նույնքան կարևոր և ընդհանրապես նույնիսկ ավելի կարևոր տարր։ Այժմ եթե մենք դառնանք այն տեսակներին, որոնք ապրում են Գալապագոսյան արշիպելագի վրա և գտնված են նաև աշխարհի այլ մասերում, մենք կտեսնենք, որ նրանք տարբեր կղզիների վրա շատ խիստ զանազանվում են։ Սակայն այդ տարբերությունը կարելի է սպասել, եթե կղզիները բնակեցվել են պատահական եղանակների օգնությամբ, օրինակ, եթե մի բույսի սերմ ընկել է մի կղզու վրա, իսկ մյուսի սերմը մի այլ կղզու վրա, թեև նրանք բոլորն էլ ծագում են առել միևնույն աղբյուրից։ Հետևաբար, երբ նախկին ժամանակներում գաղթականն ընկել է կղզիներից մեկը և հաջորդականորեն տարածվել է մեկից մյուսի վրա, նա իհարկե զանազան կղզիների վրա ընկել է տարբեր պայմանների մեջ, որովհետև նա պետք է մրցեր օրգանիզմների տարբեր խմբերի հետ. օրինակ, բույսը կարող էր զանազան կղզիների վրա գտնել, որ իրեն համար ամենապետքական գետինը զբաղված է մի քանի տարբեր տեսակներով և ինքը կարող էր ենթարկվել մի քանի տարբեր թշնամիների հարձակմանը։ Եթե նա սկսեր այստեղ փոփոխվել, ապա բնական ընտրությունը հավանորեն կսկսեր զանազան կղզիների վրա նպաստել զանազան տարատեսակների։ Սակայն որոշ տեսակներ կարող են տարաբնակվել, պահպանելով իրենց առանձնահատկությունները, այնպես, ինչպես որոշ տեսակներ լայն կերպով տարաբնակվում են մայր ցամաքի վրա և մնում են նույնը։
Իրոք, ինչպես Գալապագոսյան արշիպելագի վերաբերմամբ, այնպես էլ նվազ չափով այլ համանման դեպքերում զարմանալի է թվում այն փաստը, որ նոր տեսակներն առաջանալով մի կղզու վրա, հետո արագ կերպով չեն տարաբնակվել մյուսների վրա։ Այդ կղզիները թեև մեկը մյուսին մոտ են, բայց բաժանված են ծովի խոր բազուկներով, որոնք մեծ մասամբ Բրիտանական ջրանցքից ավելի լայն են, և հիմք չկա կարծելու, որ նրանք երբևիցե անմիջականորեն միացած են եղել։ Կղզիների միջև անցնող ծովային հոսանքներն արագ են, այստեղ փոթորիկներն արտասովոր հազվագյուտ են, և այդ պատճառով այդ կղզիներն իսկապես միմյանցից բաժանված են անհամեմատ ավելի իրականորեն, քան այդ թվում է քարտեզի վրա։ Այնուամենայնիվ, մի քանի տեսակներ, թե աշխարհի այլ մասերում գտնվածներից և թե արշիպելագին պատկանողներից, տարածված են մի քանի կղզիների վրա, և հիմնվելով նրանց այժմյան տարածման վրա, մենք կարող ենք կարծել, որ նրանք մի կղզուց տարաբնակվել են մյուսների վրա։ Սակայն ես կարծում եմ, որ մենք հաճախ սխալ տեսակետ ունենք մերձավոր տեսակների մեկը մյուսին պատկանող մարզերը փոխադարձաբար թափանցելու հավանականության մասին, երբ նրանց միջև ազատ հաղորդակցություն կա։ Կասկած չկա, որ եթե մի տեսակ ունի որևէ առավելություն մյուսի հանդեպ, նա կարճ ժամանակում իսպառ կամ մասամբ դուրս կմղի նրան. բայց եթե երկուսն էլ միանման լավ հարմարված են իրենց գրաված մարզերին, այդ դեպքում, հավանորեն, երկուսն էլ կպահպանեն իրենց տեղերը անկախ ժամանակի տևողությունից։ Լավ գիտենալով այն փաստը, որ մարդու միջոցով նատուրալիզացիայի ենթարկված շատ տեսակներ տարածվել են զարմանալի արագությամբ ընդարձակ տարածությունների վրա, մենք հակված ենք կարծելու, որ այդ բանին ընդունակ է տեսակների մեծ մասը. բայց մենք պետք է հիշենք, որ այն տեսակները, որոնք նատուրալիզացիայի են ենթարկվել նոր երկրներում, սովորաբար բոլորովին մոտ չեն բուն բնակիչներին, այլ շատ տարբերվում են նրանցից, մեծ մասամբ պատկանելով, ինչպես այդ ցույց է տվել Ա. դր-Կանդոլը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] զանազան սեռերի։ Գալապագոսյան արշիպելագի վրա նույնիսկ շատ թռչուններ, չնայած նրան, որ լավ հարմարված են կղզուց կղզի թռչելու համար, զանազան կղզիների վրա զանազան են. այսպես, նրանց մեջ կան երեք տեսակի կեռնեխներ, որոնցից ամեն մեկը հարմարված է իր կղզուն։ Այժմ ենթադրենք, թե քամին Չատամ կղզուց կեռնեխին քշել է Չարլզ կղզին, որն ունի իր կեռնեխը. նա ինչո՞ւ պիտի կարողանար հաջողությամբ հաստատվել այստեղ։ Մենք համարձակորեն կարող ենք կարծել, որ Չարլզ կղզին ամբողջովին բնակված է իր սեփական տեսակով, որովհետև ամեն տարի ավելի շատ ձու է ածվում և ավելի շատ ձագեր հանվում, քան իրականում կարելի է կերակրել, և մենք կարող ենք ենթադրել, որ Չարլզ կղզուն պատկանող կեռնեխը գոնե նույնքան լավ է հարմարված իր մարզին, ինչքան Չատամ կղզուն պատկանող տեսակը։ Սըր Չ. Լյաելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] և մ-ր Ուոլլաստոնը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 65] դրա մասին հաղորդել են ինձ հետևյալ ուշագրավ փաստը. Մադեյրա կղզին և նրան հարակից Պորտո-Սանտո կղզյակն ունեն ցամաքային կակղամորթների տարբեր, բայց միմյանց փոխարինող շատ տեսակներ, որոնցից մի քանիսն ապրում են քարերի ծերպերում. և թեև ամեն տարի Պորտո-Սանտոյից մեծ քանակությամբ քար են փոխադրում Մադեյրա, սակայն վերջին կղզին դեռևս չի բնակված Պորտո-Սանտոյի տեսակներով, այնուամենայնիվ, երկու կղզիներն էլ գրավել են եվրոպական ձևերը, որոնք հանկասկածելիորեն որոշ առավելություն ունեն տեղական տեսակների հանդեպ։ Այս նշումներից հետո, ես կարծում եմ, մենք առանձնապես չենք զարմանա, որ Գալապագոսյան արշիպելագի զանազան կղզիների վրա բնակվող էնդեմիկ տեսակներից ոչ բոլորն են տարածվել մի կղզուց մյուսը։ Մայր ցամաքի վրա տեղի ավելի վաղ գրավումը հավանորեն մեծ դեր է կատարում այն տեսակների գաղթականների տեղափոխման իրադրության մեջ, որոնք բնակվում են նման ֆիզիկական պայմաններ ունեցող այլ մարզերում։ Այսպես, Ավստրալիայի հարավ-արևելյան և հարավ-արևմտյան անկյուններն ունեն համարյա միևնույն ֆիզիկական պայմանները և միացած են իրար հետ չընդհատվող ցամաքով, բայց և այնպես նրանց վրա ապրում են մեծ թվով զանազան կաթնասուններ, թռչուններ և բույսեր, ըստ Բետսի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 181] նույնն է ստացվում նաև թիթեռների և այլ կենդանիների վերաբերմամբ, որոնք բնակվում են Ամազոնի լայնարձակ, բաց ու չընդհատվող հովտում։
Օվկիանոսային կղզիների բնակիչների ընդհանուր բնույթը որոշող սկզբունքը, այսինքն՝ կախումը այն աղբյուրից, որտեղից գաղթականները կարողացել են ամենից ավելի հեշտությամբ տարաբնակվել ու հետագայում փոփոխվել, իր լայն կիրառությունն է գտնում բնության մեջ։ Մենք նրան հանդիպում ենք յուրաքանչյուր լեռնագագաթի վրա, յուրաքանչյուր լճի և յուրաքանչյուր ճահճի մեջ։ Այդ պատճառով էլ ալպիական կենդանիները, բացառությամբ այն դեպքից, երբ տեսակը լայնորեն տարաբնակվել է սառցադաշտային ժամանակաշրջանում, մոտ են շրջապատող ցածրավայրերի կենդանիներին. այսպես, Հարավային Ամերիկայում մենք ունենք ալպիական կոլիբրիներ, ալպիական կրծողներ և ալպիական բույսերը, որոնք անկասկած պատկանում են ամերիկական ձևերին, դրա հետ միասին պարզ է, որ լեռն իր աստիճանական բարձրացման հետ զուգընթաց գաղութաբնակվել է շրջապատող ցածրավայրերից։ Նույնը վերաբերում է նաև լճերի ու ճահիճների բնակիչներին, բացառությամբ այնպիսիների, երբ տարածման հեշտությունը առաջ է բերել որոշ ձևերի տիրապետություն երկրագնդի մեծ մասի տարածության վրա։ Մենք այդ նույն սկզբունքը տեսնում ենք Ամերիկայի և Եվրոպայի քարանձավներում ապրող կույր կենդանիների վրա։ Կարելի է բերել և այլ նման դեպքեր։ Ես կարծում եմ, որ ընդհանրապես կարելի է որպես կանոն ընդունել, որ երբ երկու նույնիսկ շատ հեռավոր մարզերում հանդիպում են մեծ թվով մերձավոր կամ փոխարինվող տեսակներ, այստեղ էլ կգտնվեն նաև մի քանի ընդհանուր ձևեր, որոնց մի քանի բնախույզներ համարում են տեսակներ, մյուսները՝ միայն տարատեսակներ. այդ կասկածելի ձևերը մեգ ցույց են տալիս փոփոխականության զարգացման մեջ եղած զանազան աստիճանները։
Մի քանի տեսակների գաղթելու ընդունակության և նրանց գաղթումների չափերի միջև գոյություն ունեցող հարաբերությունը ինչպես ներկայումս, այնպես էլ ավելի վաղ ժամանակաշրջաններից որևէ մեկում, նաև շատ մերձավոր տեսակների գոյությունը երկրագնդի հեռավոր կետերում ապացուցվում են և մի այլ, ավելի ընդհանուր եղանակով։ Մ-ր Գուլդը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 89] դեռևս վաղուց ինձ ցույց է տվել, որ թռչունների այն սեռերում, որոնք տարածված են ամբողջ աշխարհում, շատ տեսակներ գրավում են ընդարձակ մարզեր։ Ես չեմ կարող կասկածել, որ այս կանոնն ընդհանրապես ճիշտ է, թեև դժվարությամբ է ապացուցվում։ Մենք տեսնում ենք, որ կաթնասունների մեջ այն ցայտուն կերպով արտահայտված է չղջիկների մոտ և ավելի փոքր չափով կատուների և շների ընտանիքներում։ Նույն կանոնը դրսևորվում է նաև թիթեռների և բզեզների տարածման մեջ։ Նույնն է քաղցրահամ ջրերում ապրողների մեծամասնության մոտ, որովհետև ամենատարբեր դասերի շատ սեռեր տարածված են ամբողջ երկրագնդի վրա և շատ տեսակներ էլ ունեն հսկայական տարածում։ Իհարկե, դա չի նշանակում, որ բոլորը, այլ լայն արեալ ունեցող սեռային խմբերի միայն մի քանի տեսակները տարածված են շատ լայն կերպով. չի ենթադրվում նաև, որ այդպիսի սեռերի տեսակներն ընդհանրապես ունենում են լայն տարածում, որովհետև դա մեծ լափով կախված է նրանից, թե ինչքան առաջ է գնացել փոփոխականության պրոցեսը։ Օրինակ, միևնույն տեսակի երկու տարատեսակներ ապրում են Ամերիկայում և Եվրոպայում, այդ պատճառով էլ տեսակը լայն տարածում ունի, բայց եթե փոփոխությունը փոքր ինչ առաջ գնա, այդ դեպքում երկու տարատեսակները կդառնան ինքնուրույն տեսակներ, և նրանց տարածման մարզերը զգալի չափով կկրճատվեն։ Մենք չենք պնդի, թե մի տեսակ, եթե միջոցներ ունի հաղթահարելու խոչընդոտները և տարածվելու լայն կերպով, ինչպես մենք այդ տեսնում ենք լավ թռչող թռչունների դեպքում, ապա և լայնորեն կտարածվի, որովհետև մենք երբեք չպետք է մոռանանք, որ լայն տարածվելու ընդունակությունը կախված է ոչ միայն խոչընդոտների վրայով անցնելու ընդունակությունից, այլև հեռավոր երկրում օտար մրցակիցների դեմ մղվող գոյության կռվում հաղթանակ տանելու ավելի կարևոր ընդունակությունից։ Սակայն համաձայնելով այն կարծիքին, որ մի սեռի բոլոր տեսակները, թեկուզև ցրված լինեն երկրագնդի ամենահեռավոր կետերում, առաջացել են մի ընդհանուր նախածնողից, ապա մենք պետք է գտնենք, և ես կարծում եմ, որպես կանոն, մենք գտնում ենք, որ այդ տեսակներից առնվազն մի քանիսն ունեն շատ ընդարձակ տարածում։
Մենք պետք է հիշենք, որ բոլոր դասերի մեջ շատ սեռեր շատ հին ծագում ունեն, և այդպիսի դեպքում նրանց տեսակները բավականաչափ ժամանակ են ունեցել տարաբնակման և հետագա փոփոխությունների համար։ Երկրաբանական տվյալներն իրենց հերթին հնարավորություն են տալիս ենթադրելու, որ յուրաքանչյուր խոշոր դասի սահմաններում ցածրակարգ օրգանիզմները ավելի դանդաղ են փոփոխվում, քան բարձրակարգ օրգանիզմները․ հետևաբար, նրանք ավելի շատ շանսեր են ունեցել լայն տարածման համարի պահպանելով միևնույն տեսակային առանձնահատկություններրը։ Այդ փաստը այն հանգամանքի հետ միասին, որ ամենից ավելի ցածր կազմվածք ունեցող ձևերի սերմերն ու ձվերը շատ փոքր են և ավելի հարմար են հեռավոր տարածություններ տեղափոխվելու համար, հավանորեն բացատրում է արդեն վաղուց հայտնի այն օրենքը, որը վերջերս քննարկել է Ա. դը-Կանդոլը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] բույսերի վերաբերմամբ, որ որքան ցածր է կանգնած օրգանիզմների խումբը, այնքան լայն է նրա տարածումը։
Հենց նոր բերված հարաբերակցությունները, այսինքն, որ ցածրակարգ օրգանիզմներն ավելի լայն կերպով են տարածված, քան բարձրակարգները, որ լայնորեն տարածված սեռերի մի քանի տեսակներն իրենց հերթին ունեն լայն տարածում. այնպիսի փաստերը, այն է՝ որ ալպիական, լճային և ճահճային արտադրանքներն ընդհանրապես մոտ են հարակից ցածր ու չոր վայրերում ապրողներին. ուշագրավ առնչությունը կղզիների և ամենամոտ մայր ցամաքի բնակիչների միջև․ միևնույն արշիպելագի կղզիների բնակիչների ավելի ես սերտ հարաբերակցությունը մեկը մյուսի նկատմամբ, ահա այս ամենը անբացատրելի է այն սովորական տեսակետով, որն ընդունում է յուրաքանչյուր տեսակի անկախ ստեղծագործությունը, բայց բացատրելի է, եթե մենք ընդունենք, որ գաղթականները եղել են ամենամոտ և ամենահարմար մի աղբյուրից՝ հետագայում հարմարվելով նոր բնակավայրին։
ՆԱԽՈՐԴ ԵՎ ՆԵՐԿԱ ԳԼՈՒԽՆԵՐԻ ԲՈՎԱՆԴԱԿՈՒԹՅԱՆ ՀԱՄԱՌՈՏ ԱԿՆԱՐԿ
Այս գլուխներում ես աշխատել եմ ցույց տալ, որ եթե մենք անհրաժեշտորեն ընդունենք մեր անտեղյակությունը կլիմայի և ցամաքի մակերևույթի այն փոփոխությունների բազմակողմանի ազդեցության մասին, որոնք անկասկած տեղի են ունեցել վերջին երկրաբանական ժամանակաշրջանի ընթացքում, նմանապես և այլ փոփոխությունների ազդեցության մասին, որ հավանորեն տեղի են ունեցել, եթե մենք մտաբերենք մեր անծանոթությունը տարածման շատ հետաքրքրական պատահական եղանակների հետ, եթե ըմբռնենք (որը վերին աստիճանի կարևոր է), թե ինչքան հաճախ տեսակները գրավել են լայնարձակ, անընդմեջ մի մարզ ու հետո մահաջնջվել են միջակա մարզերում, ապա մեր առջև անհաղթահարելի խոչընդոտ չի լինի, որպեսզի ընդունենք այն դրույթը, որ մի տեսակի բոլոր անհատները, որտեղ էլ նրանք գտնված լինեն, ծագում են առել ընդհանուր նախածնողներից։ Այդ եզրակացությանը, որին եկել են նաև ստեղծագործության կենտրոնի միասնականության գաղափարը պաշտպանող շատ բնախույզներ, մեզ բերում են զանազան ընդհանուր կշռադատություններ, առավելապես բազմապիսի արգելքների նշանակությունը և ենթասեռերի, սեռերի և ընտանիքների նույնանման տարածումը։
Ինչ վերաբերում է մի սեռի պատկանող առանձին տեսակներին, որոնք մեր տեսության համաձայն պետք է դուրս եկած ու տարաբնակված լինեին մի հիմնական մարզից, ապա եթե մենք խոստովանենք, ինչպես այդ արեցինք հիմա, մեր տեղեկությունների անբավարարությունը և հիշենք, որ մի քանի ձևեր փոփոխվել են շատ դանդաղ կերպով, որի հետևանքով գաղթելու համար նրանց տրամադրության տակ հսկայական ժամանակամիջոց է եղել, այն ժամանակ դժվարություններն անհաղթահարելի չեն թվա, թեև այդ դեպքում, ինչպես նաև մի տեսակի անհատների դեպքում նրանք հաճախ մեծ են լինում։
Օրգանիզմների տարածման վրա կլիմայական փոփոխությունների ազդեցությունը օրինակի վրա պարզելով, ես աշխատել եմ ցույց տալ, թե ինչպիսի կարևոր դեր է կատարել վերջին սառցադաշտային ժամանակաշրջանը, որն իր ազդեցությունը տարածել է նույնիսկ հասարակածային մարզերի վրա և հյուսիսում ու հարավում ցուրտ ժամանակաշրջանի հերթափոխման ընթացքում թույլ է տվել տարբեր կիսագնդերի արտադրանքների խառնում, ըստ որում նրանցից մի քանիսը աշխարհի բոլոր երկրներում մնացել են լեռնագագաթների վրա։ Ցույց տալու համար թե որքան մեծ է տեղափոխության պատահական եղանակների բազմազանությունը, ես մի քիչ ավելի երկար եմ կանգ առել քաղցրահամ ջրերում ապրող օրգանիզմների տեղափոխության եղանակների վրա։
Եթե անհաղթահարելի դժվարությունների չի հանդիպում այն ենթադրությունը, որ մի տեսակի պատկանող բոլոր անհատները կամ մի սեռի պատկանող բոլոր տեսակները երկար ժամանակամիջոցի ընթացքում դուրս են եկել մի աղբյուրից ու տարաբնակվել, այդ դեպքում աշխարհագրական տարածման բոլոր ղեկավար փաստերը բացատրվում են գաղթերի տեսությամբ, դրանց հաջորդական փոփոխությամբ և նոր ձևերի բազմացմամբ։ Այստեղից մեզ հասկանալի է դառնում արգելքների կամ անջրպետների — լինեն նրանք ջուր կամ ցամաք — հսկայական նշանակությունը ոչ միայն որպես սահմանազատիչների, այլև որպես բուսաբանական ու կենդանաբանական մարզեր առաջացնողների։ Այստեղից էլ մեզ հասկանալի է դառնում մերձավոր տեսակների կենտրոնացումը մի մարզի սահմաններում, ինչպես նաև այն, թե ինչու տարբեր լայնությունների տակ, օրինակ, Հարավային Ամերիկայում, հարթավայրերի և լեռների, անտառների, ճահիճների և անապատների բնակիչները միմյանց հետ կապված են այնքան խորհրդավոր կապերով և հավասարապես կապված են մահաջնջված օրգանիզմների հետ, որոնք առաջ ապրել են այդ մայր ցամաքի վրա։ Ընդունելով, որ օրգանիզմների փոխհարաբերությունները հսկայական նշանակություն ունեն, մենք կարող ենք հասկանալ, թե ինչու համարյա միանման ֆիզիկական պայմաններ ունեցող երկու երկրներ հաճախ բնակված են լինում շատ տարբեր ձևերով։ Կախված այն ժամանակամիջոցից, որ անցել է այն ժամանակից սկսած, երբ գաղթականները գրավել են մեկ կամ երկու մարզ, կախված հաղորդակցության այն եղանակից, որով պայմանավորվել է առավել կամ պակաս չափով այս ձևերի, այլ ոչ թե մյուս ձևերի տեղափոխությունը. կախված կամ անկախ նրանից, թե գաղթականներն ինչ չափով են ստիպված եղել շատ թե քիչ ուղղակի բախվելու միմյանց հետ և բնիկների հետ և վերջապես կախված նրանից, թե գաղթականներն ինչ արագությամբ կարողացել են փոփոխվել երկու կամ ավելի մարզերում, անկախ նրանց ֆիզիկական պայմաններից,— այս բոլորից կախված կարող էին ստացվել անսահմանորեն բազմազան կենսապայմաններ,— այստեղ կարող է լինել օրգանական ազդեցությունների ու հակազդեցությունների անվերջ բազմազանություն, և մենք կարող ենք սպասել, որ մի քանի խմբեր կգտնենք շատ խիստ, մյուսները միայն թույլ կերպով փոփոխված, մի քանիսը մեծ, մյուսները փոքր քանակությամբ, որը և մենք իսկապես էլ տեսնում ենք երկրագնդի աշխարհագրական մի քանի մեծ մարզերում։
Նույն սկզբունքների հիման վրա մենք կարող ենք հասկանալ, ինչպես ես աշխատել եմ ցույց տալ, թե ինչու օվկիանոսային կղզիների վրա բնակիչները քիչ են, բայց նրանց մեջ էնդեմիկ կամ տեղական ձևերը շատ են, թե ինչու տեղափոխության եղանակներից կախված էակների մի խումբը կազմված է միայն բնորոշ տեսակներից, մինչդեռ մյուսը, որը նույնիսկ միևնույն դասից է, կարող է ունենալ հարևան աշխարհամասի հետ նույն տեսակները։ Մենք կարող ենք նաև հասկանալ, թե ինչու օրգանիզմների ամբողջ խմբեր, ինչպիսիք են երկկենցաղները և ցամաքային կաթնասունները, բոլորովին բացակայում են օվկիանոսային կղզիների վրա, մինչդեռ ամենահեռավոր օվկիանոսային կղզիներն անգամ կարող են ունենալ իրենց օդային կաթնասունների սեփական բնորոշ տեսակները, այսինքն՝ չղջիկները։ Հասկանալի է դառնում, թե ինչու որոշ կապ կա կղզիների վրա ավելի կամ պակաս չափով փոփոխված կաթնասունների ներկայության և այդպիսի կղզիները մայր ցամաքից բաժանող ծովի խորության միջև։ Մեզ համար պարզ է դառնում, թե ինչու արշիպելագի բնակիչները զանազան կղզիների վրա զանազան տեսակներով ներկայացված լինելով հանդերձ, այնուամենայնիվ իրար մոտ են և ավելի նվազ չափով մոտ են նաև այն ամենամերձավոր մայր ցամաքի կամ մի այլ տեղի բնակիչներին, որտեղից գաղթականները կարող էին դուրս եկած լինել։ Կարելի է հասկանալ, թե ինչու երկու մարզերում, անկախ նրանց բաժանող տարածությունից, եթե կան միմյանց մոտ կամ փոխարինող տեսակներ, ապա գրեթե միշտ կգտնվեն նաև մի քանի ընդհանուր տեսակներ։
Կյանքը ղեկավարող օրենքները հանդես են բերում ժամանակի և տարածության տեսակետից ուշագրավ զուգահեռություն, որի վրա հաճախ մատնանշել է հանգուցյալ Էդվարդ Ֆորբսը․[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 88] այն օրենքները, որոնք անցյալ ժամանակներում որոշել են ձևերի հաջորդումը, գրեթե նույնն են, ինչ որ այժմ զանազան աշխարհագրական մարզերում ձևերի տարբերությունը որոշող օրենքները։ Մենք այդ տեսնում ենք բազմաթիվ փաստերի վրա։ Յուրաքանչյուր տեսակի կամ տեսակների խմբի գոյությունն անընդհատ է ժամանակի տեսակետից. թվացող բացառություններն այդ կանոնից այնքան քիչ են, որ մենք կարող ենք դրանք վերադրել այն հանգամանքին, որ միջանկյալ շերտերում մենք դեռ չենք գտել այն ձևերը, որոնք գոյություն ունեն վերևում և ներքևում։ Ճիշտ նույնը նաև տարածության վերաբերմամբ պետք է որպես կանոն ընդունել, որ յուրաքանչյուր տեսակի կամ մի քանի տեսակների մարզն անընդմեջ է, իսկ բացառությունները, թեև նրանք հազվագյուտ չեն, կարող են բացատրվել, ինչպես ես աշխատել եմ ցույց տալ, կամ տարբեր պայմաններում կատարված նախկին գաղթերով, կամ տարածման պատահական եղանակներով, կամ տեսակի մահաջնջմամբ միջանկյալ մարզերում։ Տեսակները և տեսակների խմբերն ունեն առավելագույն զարգացման կետեր հավասարապես ինչպես ժամանակի, այնպես էլ տարածման տեսակետից։ Տեսակների խմբերը, որոնք ապրում են միևնույն ժամանակաշրջանի ընթացքում կամ մի մարզում, հաճախ բնորոշվում են նույնիսկ չնչին ընդհանուր գծերով, ինչպիսիք են նախշերը կամ գույները։ Անցյալ դարերի երկար հաջորդականության կամ երկրագնդի վրա միմյանցից հեռու գտնվող մարզերի մասին խոսելով, մենք գտնում ենք, որ մի քանի դասերի տեսակներ քիչ են տարբերվում իրարից, մինչդեռ մի այլ դասի մեջ կամ նույն խմբի ստորաբաժանման մեջ նրանց միջև գոյություն ունի մեծ տարբերություն։ Յուրաքանչյուր դասի ցածր կազմվածք ունեցող ներկայացուցիչները սովորաբար բարձր կազմվածք ունեցողներից ավելի քիչ են փոփոխվում ինչպես ժամանակի, այնպես էլ տարածության մեջ. բայց երկու դեպքում էլ նկատված են այդ կանոնից բացառություններ։ Մեր տեսության համաձայն ժամանակի և տարածության միջև եղած հարաբերակցությունները հասկանալի են, որովհետև, երբ մենք խոսում ենք այն մերձավոր ձևերի մասին, որոնք փոփոխվել են հաջորդական դարերի ընթացքում, կամ այն ձևերի մասին, որոնք փոփոխվել են հեռավոր մարզեր գաղթելուց հետո, միևնույնն է, երկու դեպքում էլ նրանք միմյանց հետ կապված են հաջորդող սերունդների սովորական կապերով, երկու դեպքում էլ փոփոխականության օրենքները նույնն են, և փոփոխությունները կուտակվել են բնական ընտրության միշտ նույն եղանակով։
ԳԼՈՒԽ XIV։ ՓՈԽԱԴԱՐՁ ԱԶԳԱԿՑՈՒԹՅՈՒՆԸ ՕՐԳԱՆԻԶՄՆԵՐԻ ՄԻՋԵՎ. ՄՈՐՖՈԼՈԳԻԱ. ՍԱՂՄՆԱԲԱՆՈՒԹՅՈՒՆ. ԹԵՐԱՃ ՕՐԳԱՆՆԵՐ.
ԴԱՍԱԿԱՐԳՈՒՄԸ, ԽՄԲԵՐԻ ՀԱՄԱՍՏՈՐԱԴԱՍՈՒՄԸ։— ԲՆԱԿԱՆ ՍԻՍՏԵՄ։— ԴԱՍԱԿԱՐԳՄԱՆ ԿԱՆՈՆՆԵՐՆ ՈՒ ԴԺՎԱՐՈՒԹՅՈՒՆՆԵՐԸ, ՈՐՈՆՔ ԲԱՑԱՏՐԵԼԻ ԵՆ ԾԱԳՄԱՆ ՄԻԱՍՆԱԿԱՆՈՒԹՅԱՆ ԵՎ ՀԱՋՈՐԴԱԿԱՆ ՓՈՓՈԽՈՒԹՅՈՒՆՆԵՐԻ ՏԵՍՈՒԹՅԱՆ ՀԻՄԱՆ ՎՐԱ։— ՏԱՐԱՏԵՍԱԿՆԵՐԻ ԴԱՍԱԿԱՐԳՈՒՄԸ։— ԴԱՍԱԿԱՐԳՄԱՆ ՀԱՄԱՐ ՄԻՇՏ ՕԳՏԱԳՈՐԾՈՒՄ ԵՆ ԾԱԳՈԻՄԸ։— ԱՆԱԼՈԳԻԱԿԱՆ ԿԱՄ ՀԱՐՄԱՐՈՂԱԿԱՆ ՆՄԱՆՈՒԹՅՈՒՆՆԵՐ։— ԸՆԴՀԱՆՈՒՐ, ԲԱՐԴ ԵՎ ՏԱՐԱՄԻՏՎՈՂ ԱԶԳԱԿՑՈՒԹՅՈՒՆ։— ՄԱՀԱՋՆՋՈՒՄԸ ՄԵԿՈՒՍԱՑՆՈՒՄ ՈՒ ՍԱՀՄԱՆԱԶԱՏՈՒՄ Է ԽՄԲԵՐԸ։— ՄՈՐՖՈԼՈԳԻԱ. ՄԻԵՎՆՈՒՅՆ ԽՄԲԻ ԱՆԴԱՄՆԵՐԻ ԵՎ ՄԻԵՎՆՈՒՅՆ ՕՐԳԱՆԻԶՄԻ ՄԱՍԵՐԻ ՀԱՄԵՄԱՏՈՒԹՅՈՒՆԸ։— ՍԱՂՄՆԱԲԱՆՈՒԹՑՈՒՆ, ՆՐԱ ՕՐԵՆՔՆԵՐԸ, ՆՐԱՆՑ ԲԱՑԱՏՐՈՒԹՅՈՒՆԸ ՓՈՓՈԽՈՒԹՅՈՒՆՆԵՐՈՎ, ՈՐՈՆՔ ԵՐԵՎԱՆ ԵՆ ԳԱԼԻՍ ՈՉ ՎԱՂ ՀԱՍԱԿՈՒՄ Եվ ԺԱՌԱՆԳՎՈՒՄ ԵՆ ՀԱՄԱՊԱՏԱՍԽԱՆ ՀԱՍԱԿՈՒՄ։— ԹԵՐԱՃ ՕՐԳԱՆՆԵՐ. ՆՐԱՆՑ ԾԱԳՄԱՆ ԲԱՑԱՏՐՈՒԹՅՈՒՆԸ։— ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ։
ԴԱՍԱԿԱՐԳՈՒՄ
Մենք գիտենք, որ երկրագնդի պատմության ամենահեռավոր ժամանակաշրջաններից սկսած օրգանիզմների միջև նմանությունն արտահայտվում է զանազան վայրընթաց աստիճաններով, այնպես որ նրանց կարելի է դասավորել ստորադաս նշանակության խմբերով։ Այդ դասակարգումը կամայական չէ, ինչպես կամայական է, օրինակ, աստղերի խմբավորումը համաստեղություններում։ Խմբերի գոյությունը պարզ նշանակություն կունենար, եթե մի խումբը հարմարված լիներ բացառապես ցամաքային կյանքի համար, իսկ մյուսը՝ ջրային, մեկը մսեղեն սնունդ գործածելու համար, մյուսը՝ բուսական նյութեր և այլն. բայց իրականության մեջ դրությունը բոլորովին այլ է, որովհետև լավ հայտնի է, թե ինչպես հաճախ նույնիսկ մի ենթախմբի անդամները տարբերվում են միմյանցից իրենց կենցաղով։ Երկրորդ և չորրորդ գլուխներում խոսելով փոփոխականության և բնական ընտրության մասին, ես աշխատել եմ ցույց տալ, որ յուրաքանչյուր երկրի սահմաններում ամենից ավելի փոփոխական են լայն կերպով տարածված ու սովորական ձևերը, այսինքն յուրաքանչյուր դասի սեռային ընդարձակ խմբերի գերիշխող տեսակները։ Այս կերպ առաջացող տարատեսակները կամ սկզբնավորվող տեսակները վերջիվերջո դառնում են նոր, առանձնացած տեսակներ, վերջիններս ժառանգականության օրենքով ձգտում են առաջացնել նոր և գերիշխող տեսակներ։ Հետևաբար, այժմյան ընդարձակ խմբերը, որոնք ընդգրկում են գերիշխող շատ տեսակներ, ձգտում են զարգանալու, ծավալով շարունակ ավելի ու ավելի մեծանալու։ Այնուհետև ես աշխատել եմ ցույց տալ, որ յուրաքանչյուր տեսակի փոփոխված հաջորդները ձգտում են բնության տնտեսության մեջ գրավել հնարավորին չափ մեծ թվով բազմազան տեղեր և դրա հետևանքով նրանք երևան են հանում հատկանիշների տարամիտման հակում։ Վերջին եզրակացությունը հաստատվում է նրանով, որ նկատվում է ձևերի մեծ բազմազանություն, որոնք ամեն մի փոքր շրջանում մտնում են ամենամոտ մրցության մեջ, ինչպես նաև մի քանի տվյալներով, որ վերաբերում են նատուրալիզացիային։
Ես աշխատել եմ նաև ցույց տալ, որ թվով հետզհետե ավելացող և հատկանիշներով տարամիտվող ձևերն ունեն մշտական ձգտում ճնշելու և մահաջնջման հասցնելու այն ավելի հին ձևերին, որոնք հատկանիշներով ավելի քիչ են տարամիտվում և ավելի քիչ են կատարելագործվում։ Խնդրում եմ ընթերցողին ուշադրություն դարձնի այն դիագրամին, որը, ինչպես այդ առաջներն արդեն պարզաբանված է, լուսաբանում է այդ զանազան սկզբունքների ազդեցությունը, և նա կտեսնի, որ անխուսափելի հետևանքը կլինի ընդհանուր նախածնողի փոփոխված հաջորդների առանձնանալը, ստորադաս նշանակություն ունեցող խմբերի վերածվելը։ Դիագրամի վերին գծի վրա գտնվող յուրաքանչյուր տառ նշանակում է մի սեռ, որ պարփակում է մի քանի տեսակներ. այդ վերին գծի վրա տեղավորված բոլոր սեռերը միասին կազմում են մի դաս, որովհետև առաջացել են մի ընդհանուր նախահորից և այդ պատճառով նրանից ժառանգել են մի ինչ-որ ընդհանուր բան։ Բայց նույն հիման վրա էլ ձախակողմյան երեք սեռերը, որոնք անհամեմատ ավելի շատ ընդհանուր գծեր ունեն միմյանց հետ, կազմում են մի ենթաընտանիք, տարբեր այն ենթաընտանիքից, որը բովանդակում է երկու աջակողմյան ամենամոտ սեռերը, որոնք իրենց հատկանիշներով հեռացել են ընդհանուր նախածնողից զարգացման հինգերորդ ստադիայում։ Այս հինգ սեռերը նույնպես շատ ընդհանուր բան ունեն միմյանց հետ, բայց ավելի պակաս, քան յուրաքանչյուր ենթաընտանիքի սեռերը և միասին կազմում են մի ընտանիք, որը տարբերվում է ավելի վաղ ժամանակաշրջանում հեռացած ավելի ևս աջակողմյան երեք սեռ ընդգրկող ընտանիքից։ A-ից առաջացած բոլոր սեռերը կազմում են կարգ, որը տարբերվում է նրանից, որն ընդգրկում է I-ից առաջացած սեռերը։ Այսպիսով մենք այստեղ ունեք տեսակներ, որոնք առաջացել են ընդհանուր նախահորից և խմբավորվում են սեռերի մեջ, սեռերը՝ ենթաընտանիքների, ընտանիքների և կարգերի մեջ, բոլորը միասին՝ մի մեծ դասի մեջ։ Իմ կարծիքով, սրանով է բացատրվում խմբերի մեջ օրգանիզմների բնական ստորադասման կարևոր փաստը, մի փաստ, որն իր սովորական լինելու պատճառով միշտ չէ, որ իր վրա բավականաչափ ուշադրություն է գրավում։ Ոչ մի կասկած չկա, որ օրգանիզմները այլ առարկաների նման կարող են դասակարգվել զանազան կերպով, այն է՝ կամ արհեստականորեն, առանձին հատկանիշների հիման վրա, կամ ավելի բնականորեն, հատկանիշների գումարի հիման վրա։ Մենք գիտենք, օրինակ, որ այդ ձևով կարելի է դասակարգել հանքերը և պարզ մարմինները։ Այդ դեպքում, իհարկե, ոչ մի առնչություն չկա դասակարգման և ծննդաբանական հաջորդականության միջև, և ներկայումս անհայտ է, թե ինչի հիման վրա են նրանք բաժանվում խմբերի։ Բայց օրգանիզմների վերաբերմամբ բանն այլ կերպ է, որովհետև վերևում շարադրված տեսակետը լիովին համաձայնվում է օրգանիզմները ստորադաս նշանակություն ունեցող խմբերով դասավորելու հետ, և երբեք փորձ չի արվել այլ բացատրություն տալու այդ փաստին։
Ինչպես մենք տեսանք, բնախույզներն աշխատում են յուրաքանչյուր դասի տեսակները, սեռերը և ընտանիքները դասավորել, այսպես կոչված, բնական սիստեմում։ Բայց ի՞նչ է հասկացվում այդ սիստեմի տակ։ Մի քանի հեղինակներ նրա վրա նայում են որպես մի պարզ սխեմայի, որով կարելի է միասին դասավորել ամենից ավելի միանման օրգանիզմները և բաժանել ամենից ավելի չնմանվողները, կամ այն համարում են մի արհեստական եղանակ ընդհանուր գրույթները հնարավորին չափ համառոտակի շարադրելու համար, այսինքն մեկ նախադասության մեջ տալ ընդհանուր հատկանիշները, օրինակ, բոլոր կաթնասունների համար, մյուսի մեջ՝ բոլոր գիշատիչների համար, մի ուրիշի մեջ էլ շների սեռի համար ընդհանուր հատկանիշները, իսկ հետո ավելացնելով մասնավոր սահմանում՝ տալ արդեն շների յուրաքանչյուր տեսակի լրիվ նկարագրությունը։ Այդպիսի սիստեմի հարմարությունը և օգտակարությունը իհարկե չի կարելի վիճարկել։ Բայց շատ բնախույզներ կարծում են, որ բնական սիստեմի տակ պետք է ինչ-որ ավելի մեծ բան հասկանալ. նրանք հավատում են, որ նրա մեջ արտահայտվում է արարչի պլանը․ սակայն քանի դեռ չի որոշված, թե ինչ պետք է հասկանալ արարչի պլան ասելով արդյոք որոշ կարգ ժամանակի ու տարածության մեջ, կամ թե մեկը և թե մյուսը միասին կամ էլի որևէ այլ բան, ինձ թվում է, որ այդ ձևով մեր գիտելիքների վրա ոչինչ չի ավելանում։ Այնպիսի արտահայտությունները, որոնք նման են ավելի կամ պակաս ծածուկ ձևով կրկնվող այն հայտնի խոսքերին, որ ասել է Լիննեյը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 72] թե հատկանիշները չեն որոշում սեռը, այլ սեռն է որոշում հատկանիշները, ըստ երևույթին ցույց են տալիս, որ մեր դասակարգումները ենթադրում են ավելի խոր կապ, քան պարզ նմանություն։ Ես կարծում եմ, որ այդ այդպես էլ է և որ ծագման ընդհանրությունը, օրգանիզմների մերձավոր նմանության այդ միակ հայտնի պատճառը, հենց այն կապն է նրանց միջև, որը մասամբ բացվում է մեր առջև փոփոխությունների զանազան աստիճանները նկատող դասակարգումների միջոցով։
Այժմ ծանոթանանք դասակարգության կանոններին և այն դժվարություններին, որոնք առաջանում են այն տեսակետի պատճառով, որ դասակարգությունը կամ բացահայտում է ստեղծագործության մի ինչ֊որ անիմանալի պլան, կամ պարզապես ներկայացնում է մի սխեմա ընդհանուր դրույթներ շարադրելու համար և միմյանց հետ ամենից ավելի նման ձևերը զետեղելու համար։ Կարելի է կարծել, և, նախկին ժամանակում իսկապես այդպես էլ կարծում էին, որ դասակարգության մեջ ամենամեծ նշանակություն ունեն կառուցվածքի այն առանձնահատկությունները, որոնք իրենցով որոշում են յուրաքանչյուր էակի կենցաղը և գրաված ընդհանուր տեղը բնության տնտեսության մեջ։ Ոչինչ չի կարող ավելի սխալ լինել, քան այդ տեսակետը։ Ոչ ոք չի համարի, որ մկան և սրնչակի, դյուգոնի և կետի, կետի և ձկան արտաքին նմանությունն ունի որևէ նշանակություն։ Այդպիսի նմանությունը, ինչքան էլ նա սերտորեն կապված է այդ էակների ամբողջ կյանքի հետ, միայն որպես «հարմարողական կամ անալոգիական հատկանիշների» նշանակություն ունի և մենք դեռ կվերադառնանք այդպիսի նմանության վերլուծությանը։ Նույնիսկ կարելի է որպես ընդհանուր կանոն ընդունել, որ օրգանիզմի մի որևէ մաս որքան քիչ է կապված կենցաղի առանձնահատկությունների հետ, այնքան ավելի մեծ նշանակություն է ստանում դասակարգության համար։ Օրինակ, Օուենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] խոսելով դյուգոնի մասին, նկատում է. «Քանի որ վերարտադրողական օրգանները շատ հեռավոր առնչություն ունեն կենդանու կենցաղի և կերի հետ, ես նրանց միշտ համարել եմ այնպիսի օրգաններ, որոնք պարզորոշ ցուցում են տալիս կենդանու իսկական ազգակցության մասին։ Քիչ հավանական է, օր այդ օրգանների ձևափոխությունների մեջ մենք խառնենք պարզապես հարմարողական հատկանիշների ու էական հատկանիշների առանձնահատկությունները»։ Ինչքա՜ն զարմանալի է բույսերի նկատմամբ, որ նրանց վեգետատիվ օրգանները, որոնցից կախված է թե նրանց սննդառությունը և թե կյանքը, փոքր նշանակություն ունեն դասակարգման համար, մինչդեռ նրանց վերարտադրության օրգաններն իրենց արտադրած սերմի և սաղմի հետ միասին հսկայական կարևորություն ունեն։ Նույնն է նաև վերևում քննված մի քանի մորֆոլոգիական առանձնահատկությունների նկատմամբ, որոնք ֆունկցիոնալ տեսակետից կարևոր չեն, բայց, ինչպես մենք տեսանք, հաճախ մեծ ծառայություն են մատուցում դասակարգությանը։ Դա պայմանավորվում է նրանց կայունությամբ մերձավոր շատ խմբերի մոտ, իսկ նրանց կայունությունն իր հերթին կախված է նրանից, որ բնական ընտրությունը չի պահպանում և չի ուժեղացնում նրանց շեղումները, որովհետև նրա ազդեցությունը տարածվում է միայն սպասարկու առանձնահատկությունների վրա։
Այն, որ օրգանի միմիայն ֆիզիոլոգիական դերը չի որոշում նրա նշանակությունը դասակարգության համար, ավելի կամ պակաս չափով ապացուցվում է նրանով, որ այս կամ այն օրգանի դասակարգման համար ունեցած նշանակությունը, որը մերձավոր ազգակից խմբերի մեջ, ինչպես մենք բոլոր հիմքերն ունենք ենթադրելու, կատարում է մոտավորապես միևնույն ֆիզիոլոգիական դերը, չափազանց տարբեր է լինում։ Չկա մի բնախույզ, որը երկար ժամանակ աշխատելով որևէ մի խմբի վրա, զարմացած չլինի այդ փաստի վրա և դա նշված է գրեթե բոլոր հեղինակների աշխատություններում։ Բավական է վկայակոչել մի այնպիսի բարձր հեղինակություն, ինչպիսին է Ռոբերտ Բրոունը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 251] որը խոսելով Proteaceae-ի մի քանի օրգանների մասին, նկատում է, որ նրանց սեռային նշանակությունը «բոլոր մյուս մասերի նշանակության նման ոչ միայն այդ, այլ, ինչպես ինձ թվում է, ամեն մի բնական ընտանիքում չափազանց տարբեր և որոշ դեպքերում կարծես բոլորովին կորցված է»։ Մի այլ աշխատանքի մեջ նա ասում է, որ Connaraceae սեռի խմբերը «միմյանցից զանազանվում են իրենց ունեցած մեկ կամ մի քանի սերմնարաններով, էնդոսպերմի ներկայությամբ կամ բացակայությամբ, փեղկավոր կամ թեփուկածածկ բողբոջակազմությամբ։ Այս առանձնահատկություններից յուրաքանչյուրն առանձին հաճախ ունենում է ավելի քան սեռային նշանակություն, բայց այդ խմբում, նույնիսկ բոլորը միասին վերցրած, նրանք բավարար չեն լինում Cnestis սեռը Connarus սեռից տարբերելու համար»։ Վերցնենք մի օրինակ միջատների դասից։ Թաղանթաթև միջատների մեծ ստորաբաժանումներից մեկում բողկուկները, համաձայն Ուեստվուդի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 71] դիտողության, իրենց կառուցվածքով շատ անփոփոխ են, մյուսում՝ շատ խիստ փոփոխվում են, բայց այդ փոփոխությունները միանգամայն երկրորդական նշանակություն ունեն դասակարգության համար. բայց և այնպես ոչ ոք չի ասի, որ միևնույն կարգին պատկանող միջատների երկու տարբեր ստորաբաժանումների մոտ բողկուկների ֆիզիոլոգիական դերը տարբեր է։ Կարելի էր ուզած քանակությամբ օրինակներ բերել, որոնք ապացուցում են, որ իր ֆունկցիայով մի որոշ կարևոր օրգան խիստ տարբեր նշանակություն ունի դասակարգման համար օրգանիզմների միևնույն խմբի սահ մսաններում։
Այնուհետև, ոչ ոք չի ասի, որ թերաճ կամ ատրոֆիայի ենթարկված օրգաններն ունեն բարձր ֆիզիոլոգիական նշանակություն կամ մեծ դեր են կատարում օրգանիզմների կյանքում. սակայն ոչ մի կասկած չկա, որ նման վիճակում գտնվող օրգանները հաճախ մեծ նշանակություն ունեն դասակարգման համար։ Ոչ ոք չի վիճարկի, որ մատղաշ որոճողի վերին ծնոտում թերաճ ատամները և նրանց վերջավորությունների մի քանի թերաճ ոսկորները էական են որոճողների և հաստակաշիների մոտ ազգակցությունը ապացուցելու համար։ Ռոբերտ Բրոունը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 251] խիստ պնդել է, որ թերաճ ծաղիկների դիրքադրությունը շատ կարևոր է հացազգիների դասակարգման համար։
Կարելի էր բերել բազմաթիվ օրինակներ, որ այն հատկանիշները, որոնք վերցված են շատ կասկածելի ֆիզիոլոգիական նշանակություն ունեցող մասերից, բոլորի գնահատությամբ շատ պիտանի են ամբողջ խմբերի բնութագրման համար։ Օրինակ, քթանցքներից դեպի բերանի խոռոչը տանող բաց անցքի ներկայությունը կամ բացակայությունը, ըստ Օուենի,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] միակ հատկանիշն է, որով անպայման զատվում են ձկները սողուններից․ այդպիսին են՝ պարկավորների ստորին ծնոտի անկյան շեղումը․ միջատների թևերը ծալելու եղանակը․ որոշ ջրիմուռների գույնը․ հացազգիների ծաղկի որոշ մասերի վրա մազմզուկները. ողնաշարավորների մաշկային գոյացումների՝ փետուրների և մազերի բնույթը։ Եթե բադակտուցը մազերի փոխարեն ծածկված լիներ փետուրներով, ապա այդ արտաքին և քիչ նշանակություն ունեցող հատկանիշը բնախույզների կողմից կհամարվեր մի կարևոր միջոց այդ տարօրինակ էակի և թռչունների ազգակցության աստիճանը որոշելու համար։
Դասակարգման համար անկարևոր առանձնահատկությունների այդպիսի նշանակությունը կախված է գերազանցապես նրանց հարաբերակցությունից ավելի կամ պակաս կարևոր այլ առանձնահատկությունների հետ։ Իսկ առանձնահատկությունների կամ հատկանիշների ամբողջ գումարի նշանակությունը բնապատմության մեջ ինքնըստինքյան ակներև է։ Ուրեմն, ինչպես այդ հաճախ նշվել է, տեսակը կարող է տարբերվել իր մերձավորներից շատ հատկանիշներով, որոնք թե ֆիզիոլոգիական տեսակետից կարևոր են և թե ընդհանրապես ցայտուն կերպով են արտահայտված, բայց և այնպես մեզ թողել են կասկածանքի մեջ այն հարցի վերաբերմամբ, թե սիստեմի մեջ որտեղ պետք է զետեղել նրան։ Այստեղից էլ հայտնի է դարձել, որ մի հատկանիշի վրա հիմնված դասակարգությունը, ինչքան էլ հատկանիշը կարևոր լինի, միշտ սխալական է լինում, որովհետև չկա կազմվածքի մի այնպիսի գիծ, որը շարունակ անփոփոխ լիներ։ Հատկանիշների գումարի նշանակությունը, նույնիսկ այն դեպքում, երթ նրանց մեջ ոչ մեկը էական չէ, բացատրում է մեզ Լիննեյի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 72] արտահայտած այն աֆորիզմը, որ ոչ թե հատկանիշներն են որոշում սեռը, այլ սեռը՝ հատկանիշները, որովհետև, ինչպես թվում է, այդ աֆորիզմը բխում է նմանության այնպիսի անկարևոր շատ գծերի նշանակության գնահատումից, որոնք չափազանց թույլ են, որպեսզի կարողանային սահմանման ենթարկվել։ Malpighiaceae-ներին պատկանող մի քանի բույսեր տալիս են լրիվ և թերազարգացած ծաղիկներ․ վերջինների մոտ, ինչպես նկատում է Ա. դը-Ժյուսյեն,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 140] «տեսակի, սեռի, ընտանիքի և դասի հատկանիշների մեծ մասն անհետանում է, և այսպիսով նրանք ծիծաղում են մեր դասակարգության վրա»։ Բայց Aspicarpa-ն թեև մի քանի տարի շարունակ Ֆրանսիայում տվել է միայն այնպիսի թերազարգացած ծաղիկներ, որոնք բազմաթիվ ամենակարևոր առանձնահատկություններով զարմանալիորեն այնպես տարբերվում էին այն ընտանիքի իսկական տիպից, որին նա պատկանում է։ Ռիշարը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 252] համաձայն Ժյուսյեի դիտողության, այնուամենայնիվ խորաթափանց կերպով եկել է այն եզրակացության, որ այդ սեռը դարձյալ պետք է մնա Malpighiaceae-ների մեջ։ Այս դեպքը լավ պարզաբանում է մեր դասակարգումների ոգին։
Գործնականում բնախույզները, երբ աշխատում են, հոգ չեն տանում այն հատկանիշների ֆիզիոլոգիական նշանակության մասին, որոնց օգտագործում են խմբի բնութագրման համար կամ որևէ յուրահատուկ տեսակ բնորոշելու համար։ Եթե նրանք գտնում են քիչ թե շատ միանման հատկանիշ, որն ընդհանուր է որոշ ձևերի մեծ թվի համար և չի հանդիպում մյուսների մոտ, նրանք օգտագործում են այդ հատկանիշը՝ մեծ նշանակություն տալով նրան, իսկ եթե նա հատուկ է քիչ թվով ձևերի, այդ դեպքում նրան տալիս են երկրորդական նշանակություն։ Մի քանի բնախույզներ, լայն կերպով ընդունել են այդ սկզբունքը, և ոչ ոք այդ այնպես պարզորոշ կերպով չի արտահայտել, ինչպես հիանալի բուսաբան Ավգուստ Սենտ-Իլերը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 144] Եթե զանազան ոչ էական հատկանիշներ միշտ միասին են հանդիպում, ապա թեկուզ նրանց միջև կապ գտնված չլինի էլ, նրանց առանձին նշանակություն է վերագրվում։ Քանի որ կենդանիների խմբերի մեծամասնության մոտ զանազան կարևոր օրգանները, ինչպիսիք են արյան շրջանառության, շնչառության և բազմացման օրգանները, լինում են թե քիչ միանման, նրանց շատ պիտանի են համարում դասակարգության նպատակների համար․ բայց մի քանի խմբերում կենդանու կյանքի համար առաջնակարգ նշանակություն ունեցող այդ բոլոր օրգանները, ինչպես նկատված է, հանդիսանում են միանգամայն երկրորդական նշանակություն ունեցող հատկանիշներ։ Այսպես, վերջերս Ֆրից Մյուլլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] ցույց է տվել, որ խեցեմորթների միևնույն խմբում Cypridina-ն սիրտ ունի, մինչդեռ երկու շատ մերձավոր սեռերը՝ Cypris-ը և Cytherea-ն այդ օրգանը չունեն. Cypridina-ի մի տեսակը ունի լավ զարգացած խռիկներ, մյուսը զուրկ է դրանցից։
Քանի որ բնական դասակարգումը, իհարկե, ընդգրկում է բոլոր հասակները, ապա մեզ համար հասկանալի է, թե ինչու սաղմից վերցրած հատկանիշները նույնպես կարևոր են, ինչպես որ նրանք, որոնք վերցվում են չափահաս կենդանուց։ Բայց դրա փոխարեն սովորական տեսակետից ամենևին պարզ չէ, թե ինչու սաղմի կառուցվածքն ավելի կարևոր է դասակարգման նպատակների համար, քան չափահաս օրգանիզմի կառուցվածքը, որն էլ հենց որոշակի տեղ է գրավում բնության տնտեսության մեջ։ Սակայն այն, որ սաղմնային հատկանիշները բոլորից ամենակարևորներն են, վճռականորեն պաշտպանել են այնպիսի մեծ բնախույզներ, ինչպես Միլն Էդվարդսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 86] և Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] և այդ ուսմունքն ընդհանուրի կողմից ընդունվել է և համարվում է որպես ճշմարտություն։ Այնուամենայնիվ սաղմնային հատկանիշների նշանակությունը երբեմն չափազանցվել է, որովհետև նրանցից չեն բացառվել թրթուրների հարմարեցման առանձնահատկությունները, և որպեսզի ապացուցի այդ, Ֆրից Մյուլլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] բացառապես այդ հատկանիշների վրա է կառուցել խեցեմորթների ընդարձակ դասի սիստեմատիկան, որը և անբնական է ստացվել։ Ընդհանրապես կասկած չի կարող լինել, որ սաղմնային հատկանիշները, բացառությամբ թրթուրայինների, հսկայական նշանակություն ունեն ոչ միայն կենդանիների, այլև բույսերի դասակարգության համար։ Ծաղկավոր բույսերի գլխավոր ստորաբաժանումները հիմնված են սաղմի տարբերությունների, շաքիլների թվի և դիրքադրության և արմատիկի ու մատաղ առանցքի զարգացման եղանակի վրա։ Մենք հենց հիմա կտեսնենք, թե ինչու այդ հատկանիշները դասակարգության մեջ ունեն այդպիսի մեծ նշանակություն, եթե միայն ուշադրության առնենք, որ բնական սիստեմները կառուցված են ծննդաբանական սկզբունքի վրա։
Մեր դասակարգությունները հաճախ լիովին գտնվում են ազգակցության շարքերի ազդեցության տակ։ Ոչ մի բան այնպես հեշտ չէ, ինչպես բոլոր թռչունների համար ընդհանուր հատկանիշների գումարը տալը, բայց խեցեմորթների վերաբերմամբ այդպիսի փորձը մինչև այժմ անհնարին է եղել։ Այն խեցեմորթները, որոնք գրավում են շարքի հակադիր ծայրերը, դժվար թե ունենան որևէ ընդհանուր հատկանիշ, բայց ծայրային տեսակները, մոտ կանգնած լինելով մյուսներին, որոնք էլ իրենց հերթին մոտ են հետևյալներին և այլն, անվիճելիորեն պետք է համարվեն այդ դասին պատկանողներ, այլ ոչ թե Articulata-ի մյուս դասին։
Աշխարհագրական տարածումը նույնպես հաճախ օգտագործել են, թեև գուցե ոչ բոլորովին տրամաբանորեն, երբ դասակարգել են առանձնապես ընդարձակ խմբեր, որոնք ընդգրկում են բազմաթիվ շատ մոտ ձևեր։ Տեմմինկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 253] պնդում է այդ եղանակի օգտակարության և նույնիսկ անհրաժեշտության վրա՝ թռչունների որոշ խմբերի վերաբերմամբ, և նրան են հետևել շատ միջատաբաններ և բուսաբաններ։
Վերջապես, ինչ վերաբերում է տեսակների զանազան խմբերի համեմատական նշանակությանը, ինչպիսիք են կարգերը, ենթակարգերը, ընտանիքները, ենթաընտանիքները և սեռերը, ըստ երևույթին, գոնե ներկայումս, բոլորովին կամայական են։ Մի քանի ականավոր բուսաբաններ, ինչպես մ-ր Բենտամը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 64] և ուրիշներ, համառորեն պնդել են այդ խմբերի կամայական նշանակության վրա։ Բույսերի և միջատների համար կարելի է բերել այն օրինակները, որ պրակտիկ բնախույզների կողմից սկզբում միայն որպես սեռային ճանաչված խումբը հետո բարձրացվել է մինչև ենթաընտանիքի և ընտանիքի նշանակության․ և այդ տեղի է ունեցել ոչ թե այն պատճառով, որ հետագա հետազոտությունները հայտնաբերել են սկզբից բաց թողնված կառուցվածքային կարևոր առանձնահատկություններ, այլ այն պատճառով, որ ժամանակի ընթացքում հայտնաբերվել են բազմաթիվ տեսակներ, որոնք ունեն թույլ կերպով զանազանվող տարբերությունների աստիճաններ։
Դասակարգելիս բոլոր նախորդ կանոնները և դժվարությունները, ինչպես և օժանդակ միջոցները, եթե ես առանձնապես չեմ սխալվում, կարելի է բացատրել, ընդունելով՝ որ բնական սիստեմը բխում է փոփոխություններով ուղեկցվող ընդհանուր ծագման փաստից. որ հատկանիշները, որոնք բնախույզների կողմից համարվում են որպես ցուցում երկու կամ ավելի տեսակների իսկական հարաբերակցության մասին, մի ընղհանուր նախածնողից ժառանգված առանձնահատկություններ են և որ ամեն մի իսկական դասակարգություն ծննդաբանական է. որ այն թաքուն կապը, որը անգիտակցաբար որոնում են բնախույզները, գտնվում է հատկապես ծագման ընդհանրության մեջ, այլ ոչ բոլորովին արարչագործության ինչ֊որ անիմանալի պլանի մեջ կամ ընդհանուր դրույթների արտահայտության եղանակի և առավել կամ պակաս նման իրերի մոտեցման և անջատման ձևի մեջ։
Սակայն ես պետք է իմ միտքն արտահայտեմ ավելի լրիվ կերպով։ Ես կարծում եմ, որ մի դասի սահմաններում միմյանց հետ համապատասխան ստորադասման ու հարաբերակցության մեջ գտնվող խմբերի կառուցումը բնական լինելու համար՝ պետք է լինի խիստ ծննդաբանական. բայց տարբերությունների չափերը զանազան ճյուղերի և խմբերի մեջ, որոնք չնայած գտնվում են իրենց ընդհանուր արտադրողի նկատմամբ միևնույն ազգակցության աստիճանի վրա, կարող են մեծ չափով տարբերվել, որովհետև այդ չափերը կախված են տվյալ խմբերի անցած փոփոխությունների զանազան աստիճաններից. և դա արտահայտվում է ձևերի տեղաբաշխմամբ զանազան սեռերի, ընտանիքների, ենթակարգերի և կարգերի մեջ։ Ընթերցողին ավելի հասկանալի կլինի, թե դրա տակ ինչ է հասկացվում, եթե նեղություն հանձն առնի դիմելու այն դիագրամին, որը կցված է չորրորդ գլխին։ Ենթադրենք, թե A—L տառերը նշանակում են ազգակից սեռերը, որոնք գոյություն են ունեցել սիլուրյան դարաշրջանի ընթացքում և ծագում են առել է՛լ ավելի հին ձևերից։ Այդ սեռերից երեքի (A, F և I) մեջ մի տեսակը պահպանել է իր փոփոխված հաջորդներին մինչև մեր օրերը, և այդ հաջորդները ներկայացված են տասն և հինգ սեռերով (Չհաջողվեց վերլուծել (բառական սխալ):
), որոնք նշված են ամենավերին հորիզոնական գծի վրա։ Մեր վերցրած յուրաքանչյուր տեսակի բոլոր այդ փոփոխված հաջորդները արյունակցությամբ կամ ծագմամբ գտնվում են ազգակցության մի աստիճանի վրա, և փոխաբերաբար նրանց կարելի է անվանել միլիոներորդ աստիճանի հորեղբորորդիներ. բայց նրանք ուժեղ կերպով և շատ տարբեր աստիճանով զանազանվում են միմյանցից։ A-ից առաջացած ձևերը բաժանվել են այժմ երկու կամ երեք ընտանիքի, որոնք կազմում են մի որոշակի կարգ՝ բաժանված նրանից, որը առաջացել է I-ից, նույնպես բաժանվելով երկու ընտանիքի։ A-ից առաջացած գոյություն ունեցող տեսակները չեն կարող զետեղել մի սեռի մեջ նախածնողական A-ի հետ միասին, ճիշտ այնպես, ինչպես I-ից առաջացած տեսակները չեն կարող զետեղվել մի սեռի մեջ I-ի հետ միասին։ Բայց գոյություն ունեցող սեռը, ինչպես կաrելի է ենթադրել փոփոխվել է միայն թույլ աստիճանով և կարող է համատեղվել F նախածնողական սեռի հետ, որի օրինակ են հանդիսանում դեռ գոյություն ունեցող սակավաթիվ սիլուրյան սեռերը։ Այսպիսով, մեկը մյուսի հետ միևնույն արյունակցական ազգակցության աստիճանի վրա գտնվող օրգանիզմների միջև եղած տարբերությունների համեմատական նշանակությունը չափազանց տարբեր է դարձել։ Սակայն նրանց ծննդաբերական հարաբերակցությունը մնացել է միանգամայն միանման ոչ միայն ներկայումս, այլև նրանց զարգացման յուրաքանչյուր հաջորդական ժամանակաշրջանում։ A-ի փոփոխված բոլոր հաջորդները ժառանգել են մի ինչ որ ընդհանուր բան իրենց ընդհանուր նախածնողից, ճիշտ այնպես, ինչպես որ I-ի հաջորդները, և այդպես կլինի հաջորդների յուրաքանչյուր ստորադաս ճյուղի հետ ամեն մի հաջորդական ստադիայի վրա։ Սակայն, եթե մենք ենթադրենք, թե A-ի կամ I-ի որևէ հաջորդ այնքան է փոխվել, որ կորցրել է իր ազգակցության բոլոր հետքերը, այդ դեպքում նրա տեղն էլ բնական սիստեմում կկորչեր, որն ըստ երևույթին իրոք տեղի է ունեցել այժմ ապրող էակներից մի քանիսի հետ։ F սեռի բոլոր հաջորդներն իրենց զարգացման ամբողջ գծի ընթացքում, ինչպես ենթադրվում է, շատ քիչ են փոփոխվել և կազմում են մի սեռ։ Բայց այդ սեռը, թեև խիստ մեկուսի է կանգնած, դեռևս գրավում է իր իսկական միջակա դրությունը։ Խմբերի պատկերը այն ձևով, ինչպես այդ տրված է դիագրամի հարթության վրա, չափազանց պարզ է, որովհետև ճյուղերը տարածվել են բոլոր ուղղություններով։ Եթե խմբերի անունները գրելու լինենք պարզապես մի շարքում, այդ դեպքում խմբերի մասին պատկերացումն ավելի ևս պակաս բնական կլիներ, և հանրահայտ է, որ հարթության վրա մի շարքի մեջ անհնարին է ներկայացնել նույնիսկ այն հարաբերակցությունները, որոնց մենք հանդիպում ենք բնության մեջ միևնույն խմբի էակների միջև։ Այսպիսով, բնական սիստեմը ներկայացնում է էակների ծննդաբանական դասավորումը, ինչպես տոհմաբանական ծառի մեջ, բայց զանազան խմբերի անցած փոփոխությունների չափն արտահայտվում է նրանց զանազան, այսպես կոչված, սեռերի, ենթաընտանիքների, ընտանիքների, ենթակարգերի, կարգերի և դասերի վրա տեղաբաշխությամբ։
Դասակարգության մասին այդ տեսակետը արժանի է, որպեսզի պարզաբանված լինի լեզուների բնագավառից վերցված մի օրինակով։ Եթե մենք ունենայինք մարդկային ցեղի լիակատար ծննդաբանությունը, ապա զանազան տոհմերի տոհմաբանական դասավորումը կտար նաև աշխարհի բոլոր երկրներում գործածվող զանազան լեզուների լավագույն դասակարգումը. դա կլիներ միակ հնարավոր սիստեմը բոլոր անհետացած լեզուները և բոլոր անցումային, թեթևակի տարբերվող բարբառները մտցնելու համար։ Բայց կարող է պատահել, որ հին լեզուներից մի քանիսը շատ քիչ են փոփոխվել և սկիզբ են տվել սակավաթիվ նոր լեզուների, մինչդեռ մյուսները շատ ուժեղ կերպով են փոփոխվել՝ նայած միմյանց հետ ծագման տեսակետից կապված զանազան տոհմերի տարաբնակմանը, մեկուսացմանը և քաղաքակրթության աստիճանին, և բազմաթիվ նոր բարբառների ու լեզուների սկիզբ են տվել։ Միևնույն արմատից առաջացած լեզուների միջև տարբերության զանազան աստիճանները կարող են արտահայտվել ստորադաս նշանակության խմբեր սահմանելով. բայց և այնպես կառուցման ճիշտ կամ միակ հնարավոր սիստեմը պետք է լինի ծննդաբանականը. և նա կլիներ ամենախիստ իմաստով բնական, որովհետև, նա կարող էր միմյանց հետ կապել բոլոր լեզուները՝ ինչպես անհետացածները, այնպես էլ արդիականները, հիմնվելով նրանց սերտ ազգակցության վրա, կներկայացնի յուրաքանչյուր լեզվի տոհմագրությունը և ծագումը։
Այս տեսակետը հաստատելու համար վերցնենք այն տարատեսակների դասակարգումը, որոնց մասին մենք գիտենք կամ ենթադրում ենք, որ նրանք առաջացել են մի տեսակից։ Նրանք խմբավորվում են, կազմելով տեսակներ ճիշտ այնպես, ինչպես ենթատարատեսակները մտնում են բուն տարատեսակների կազմի մեջ, իսկ որոշ դեպքերում, օրինակ ընտանի աղավնու նկատմամբ, կարելի է սահմանել տարբերությունների նաև այլ աստիճաններ, այսինքն այդ ժամանակ հետևում են մոտավորապես հենց նույն կանոններին, ինչ որ տեսակների դասակարգման դեպքում։ Հեղինակները պնդել են, որ անհրաժեշտ է տարատեսակները դասավորել բնական, այլ ոչ թե արհեստական սիստեմով. օրինակ, մեզ նախազգուշացնում են չմիացնել անանասի երկու տարատեսակները միայն այն հիման վրա, որ երկուսի պտուղները շատ մեծ նմանություն ունեն, թեև պտուղն է ամենակարևոր մասը. ոչ ոք չի միացնի շվեդական և սովորական գոնգեղները, թեև նրանց հաստացած ուտելի ցողուններն այնքան նման են միմյանց։ Ինչ մաս որ ամենից ավելի կայուն է լինում, այն էլ օգտագործում են տարատեսակների դասակարգման համար։ Այսպես, հայտնի գյուղատնտես Մարշալը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 59] ասում է, որ եղջյուրներն այդ տեսակետից շատ օգտակար են, երբ գործ ունենք խոշոր եղջերավոր անասունների հետ, որովհետև նրանք ավելի քիչ փոփոխական են, քան մարմնակազմը, գույնը և այլն․ ընդհակառակը, երբ գործ ունենք ոչխարների հետ, եղջյուրները քիչ պիտանի են, որովհետև քիչ կայուն են։ Տարատեսակների դասակարգման վերաբերմամբ ես համոզված եմ, որ եթե մենք ունենայինք նրանց տոհմագրությունը, այդ դեպքում ծննդաբանական դասակարգությունը ամենուրեք մյուսներից գերադասելի կլիներ, և որոշ դեպքերում այդ ուղղությամբ փորձեր իրոք արվել են։ Որովհետև մենք կարող ենք վստահ լինել, որ փոփոխությունները մեծ լինեն, թե փոքր, մենք ժառանգականության սկզբունքի ուժով կմիացնենք այնպիսի ձևեր, որոնք ամենամեծ թվով շփման կետերով միմյանց մոտ են։ Այսպես, ինչ վերաբերում է թուրման աղավնիներին, թեև նրանց ենթատարատեսակներից մի քանիսը տարբերվում են մեկը մյուսից այնպիսի կարևոր հատկանիշով, ինչպիսին է կտուցի երկարությունը, սակայն բոլորը միասին միանում են իրենց գլխկոնձի տալու սովորության հետևանքով․ բայց կարճակտուց ձևը համարյա կամ բոլորովին կորցրել է այդ սովորությունը, չնայած դրան, մենք այսու հանդերձ առանց տատանվելու նրան թողնում ենք միևնույն խմբում, որովհետև նա մոտ է մնացածներին արյունակցությամբ և մի քանի տեսակետներից նրանց նման է։
Ինչ վերաբերում է բնական վիճակում եղած տեսակներին, ապա նրանց վերաբերմամբ յուրաքանչյուր բնախույզ իրականում իր դասակարգության մեջ մտցրել է նրանց ծագումը, որովհետև սիստեմի հենց առաջին աստիճաններում՝ տեսակների դասակարգության մեջ մտցված են երկու սեռեր, իսկ այն, թե ինչքան խիստ կերպով նրանք տարբերվում են իրարից նույնիսկ էական գծերով, լավ հայտնի է ամեն մի բնախույզի․ հազիվ թե կարելի է որևէ ընդհանուր բան գտնել մի քանի փնջոտնյա խեցգետինների չափահաս արուների և հերմաֆրոդիտների միջև, բայց ոչ ոք չի էլ մտածի նրանց անջատման մասին։ Հենց որ հայտնի է դարձել, որ խոլորձիների երեք ձևերը՝ Monachanthus-ը, Myanthus-ը և Catasetum-ը, որոնք մինչ այդ վերագրում էին երեք տարբեր սեռերի, երբեմն զարգանում են միևնույն բույսի վրա, անմիջապես սկսել են նրանց համարել տարատեսակներ, իսկ ներկայումս ես կարողացա ապացուցել, որ դրանք միևնույն տեսակի արական, իգական և հերմաֆրոդիտ ձևերն են։ Բնախույզը մի տեսակի է վերագրում միևնույն անհատի թրթուրային զանազան ստադիաները, ինչքան էլ որ նրանք ուժեղ կերպով տարբերվեն միմյանցից ու չափահաս կենդանուց, և ճիշտ այդպես էլ վարվում են, այսպես կոչված, Ստենստրուպի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 254] հերթափոխող սերունդների հետ, որոնք որոշ տեխնիկական իմաստով միայն կարող են ընդունվել որպես մի անհատ։ Նա որոշ տեսակի մեջ մտցնում է թե այլանդակությունները և թե շեղումները ոչ թե ծնողական ձևերի հետ նրանց ունեցած մասնակի նմանության հիման վրա, այլ այդ ձևից նրանց ունեցած ծագման հիման վրա։
Քանի որ ծագումն ընդհանրապես օգտագործում են միևնույն տեսակի պատկանող անհատներին մի ամբողջության մեջ միացնելիս, թեև արուները, էգերը և թրթուրները երբեմն չափազանց տարբեր են լինում, և քանի որ նրան օգտագործում են, երբ դասակարգում են տարատեսակները, որոնք կրել են որոշ և մի քանի դեպքերում էլ զգալի փոփոխություններ, ապա ծագումն էլ արդյոք անգիտակցաբար չե՞ն օգտագործել թե տեսակները սեռերի մեջ և թե սեռերը բարձր կարգի խմբերի մեջ խմբավորելու ժամանակ, այսպես կոչված, բնական դասակարգության մեջ։ Ես կարծում եմ, որ այն օգտագործել են անգիտակցաբար, և միայն այդ կերպով ես կարող եմ հասկանալ զանազան կանոններն ու եղանակները, որոնց հետևում են մեր ամենալավ սիստեմատիկները։ Չունենալով դրված ծննդաբանություններ, մենք հարկադրված ենք ընդհանուր ծագման հետքերը որոնել զանազան ձևի նման առանձնահատկությունների մեջ։ Դրա համար էլ մենք ընտրում ենք այնպիսի հատկանիշներ, որոնց մասին ամենաքիչ հավանականություն կա, որ նրանք փոփոխվել են գոյության այն պայմաններից կախված, որոնց մեջ վերջին ժամանակներս դրված է եղել յուրաքանչյուր տեսակ։ Այդ տեսակետից սաղմնային կառուցվածքի առանձնահատկությունները նույնքան պիտանի են կամ նույնիսկ էլ ավելի պիտանի են, քան կազմվածքի մյուս մասերը։ Մենք չենք հոգում այն մասին, թե ինչքան նվազ կարևոր կարող են լինել այդ հատկանիշները,— լինի դա միայն ստորին ծնոտի անկյան փոփոխությունը, միջատի թևերը ծալելու եղանակը, մազերով կամ փետուրներով ծածկված մաշկը. բայց եթե այդպիսի հատկանիշ ունեն տարբեր շատ տեսակներ, մանավանդ այնպիսիները, որոնք շատ ուժեղ կերպով զանազանվում են իրենց կենցաղով, այդ դեպքում նա մեծ նշանակություն է ստանում, որովհետև չափազանց տարբեր սովորություններ ունեցող ձևերի մոտ նրա ներկայությունը մենք կարող ենք բացատրել միայն մի ընդհանուր նախածնողից ժառանգված լինելու միջոցով։ Կազմվածքի առանձին առանձնահատկությունների վերաբերմամբ մենք կարող ենք այդ ուղղությամբ սխալվել. բայց եթե մի քանի, թեկուզև նվազ կարևոր առանձնահատկություններ, արտահայտված են օրգանիզմների մեծ խմբի մեջ, որոնք ունեն զանազան սովորություններ, այդ դեպքում ծագման տեսության հիման վրա մենք համարյա կարող ենք վստահ լինել, որ այդ առանձնահատկությունները ժառանգված են մի ընդհանուր նախահորից. և մենք գիտենք, որ զուգորդված հատկանիշներն առանձնապես նշանակություն ունեն դասակարգության մեջ։
Մենք կարող ենք հասկանալ, թե ինչու տեսակը կամ տեսակների խումբը կարող է զանազան շատ էական առանձնահատկություններով շեղվել մերձավոր ձևերից և այնուամենայնիվ պահպանել իր դրությունը սիստեմի մեջ նրանց կողքին։ Դա կարելի է հանգիստ կերպով ընդունել, և դա հաճախ ընդունվում է, քանի դեռ նույնիսկ անկարևոր հատկանիշների մի բավարար քանակություն դրսևորում է ծագման ընդհանուր, լինելու թաքնված կապը։ Թող երկու ձևերն էլ չունենան ոչ մի ընդհանուր հատկանիշ, բայց եթե նրանց ծայրային ձևերը մեկը մյուսի հետ միանում են միջակա խմբերի շղթայով, մենք իսկույն կարող ենք ընդունել նրանց ծագման ընդհանրությունը և նրանց բոլորին էլ զետեղում ենք մի դասի մեջ։ Գտնելով, որ ֆիզիոլոգիորեն շատ կարևոր օրգանները, այսինքն այնպիսիք, որոնք ծառայում են գոյության ամենատարբեր պայմաններում կյանքի պահպանության համար, սովորաբար շատ կայուն են, մենք նրանց առանձին նշանակություն ենք տալիս․ բայց եթե նույն օրգանները մի այլ խմբում կամ խմբի ստորաբաժանման մեջ գտնվում են խիստ փոփոխված դրությամբ, մենք իսկույն ավելի փոքր նշանակություն ենք տալիս նրանց մեր դասակարգության ժամանակ։ Հիմա մենք կհասկանանք, թե ինչու սաղմնաբանական հատկանիշները այդքան մեծ նշանակություն ունեն դասակարգելու ժամանակ։ Աշխարհագրական տարածումը նույնպես կարող է երբեմն օգուտ բերել ընդարձակ սեռերի դասակարգման ժամանակ, որովհետև բոլոր այն տեսակները, որոնք բնակվում են մի որևէ հեռավոր ու մեկուսացած մարզում, ամենայն հավանականությամբ առաջացել են միևնույն նախածնողներից։
ԱՆԱԼՈԳ ՆՄԱՆՈՒԹՅՈՒՆՆԵՐ
Վերևում շարադրվածի հիման վրա մեզ հասկանալի պիտի լինի այն վերին աստիճանի կարևոր տարբերությունը, որ կա իսկական նմանության և անալոգ կամ հարմարվողությամբ պայմանավորված նմանության միջև։ Առաջին անգամ Լամարկը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 2] ուշադրություն է դարձրել դրա վրա և գտել է իր տաղանդավոր հետևորդին՝ հանձինս Մեկլեյի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 255] և ուրիշների։ Դյուգոնների և կետերի մարմնի և լողաթևերի նմանվող առջևի վերջավորությունների ձևի մեջ եղած նմանությունը, ինչպես նաև կաթնասունների այդ երկու կարգերի և ձկների միջև նմանությունը անալոգիա է։ Այդպես է նաև մկան և սրնչակի (Sorex) միջև եղած նմանությունը, որոնք պատկանում են տարբեր կարգերի, կամ էլ ավելի մոտ նմանությունը, որի վրա պնդում է մ-ր Մայվարտը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 100] մկան և Ավստրալիայի մի փոքրիկ պարկավոր գազանիկի (Antechinus) միջև։ Այդպիսի նմանությունները, իմ կարծիքով, կարող են լինել որպես հարմարեցումներ թփուտների և խոտերի խիտ բուսականության միջով միանման շարժումներ կատարելու և դրա հետ միասին թշնամիներից թաքնվելու համար։
Միջատների մեջ այդպիսի անթիվ օրինակներ կան և այդ պատճառով էլ Լիննեյը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 72] խաբվելով արտաքին նմանությունից, Homoptera-ներից մի միջատի ցեցի տեղ է ընդունել։ Նման մի բան մենք տեսնում ենք նույնիսկ մեր ընտանի ցեղերի մեջ, օրինակ, տարբեր տեսակներից առաջացած չինական բուծարանային խոզի և սովորական խոզի անհատների մարմնի ձևի զարմանալի նմանությունը, կամ միմյանցից որպես տեսակներ տարբերվող սովորական գոնգեղի և շվեդական կերաշաղգամի ցողունների միանման հաստությունը։ Բարակ շների և արշավաձիու միջև եղած նմանությունը հազիվ թե ավելի երևակայական է, քան այն անալոգիաները, որ մի քանի հեղինակներ նշել են խիստ տարբեր կենդանիների միջև։
Համաձայն այն տեսակետի, որ դասակարգման համար հատկանիշների իսկական նշանակությունը չափվում է նրանով, թե նրանք ինչ չափով են ցույց տալիս ծագումը, կարելի է հեշտությամբ հասկանալ թե ինչու անալոգ հատկանիշները կամ հարմարեցման առանձնահատկությունները, կենդանի էակների բարօրության համար շատ կարևոր լինելով հանդերձ, գրեթե ոչ մի նշանակություն չունեն սիստեմատիկի աչքում։ Այն կենդանիները, որոնք պատկանում են ծագումով խիստ տարբեր երկու գծերի, կարող են հարմարվել միանման պայմանների և այդ պատճառով ձեռք բերել արտաքին մեծ նմանություն, բայց այդպիսի նմանությունը չի ցույց տալիս արյունակցական ազգակցություն, այլ ավելի շուտ քողարկում է այն։ Այդ պատճառով մենք կարող ենք հասկանալ նաև այն թվացող պարադոքսը, որ միևնույն հատկանիշները անալոգ են լինում, երբ համեմատում ենք միմյանց հետ տարբեր խմբեր, և ցույց են տալիս իսկական հարաբերակցությունը, երբ համեմատում ենք միևնույն խմբի անդամները միմյանց հետ. այսպես, մարմնի ձևը և լողաթևերի նմանվող վերջավորությունները միայն անալոգ հատկանիշներ են, եթե համեմատում ենք կետերին ձկների հետ, որովհետև նրանք երկու դասի մեջ էլ ջրում լողալու համար հարմարեցման առանձնահատկություններ են. բայց երբ համեմատում ենք կետերի ընտանիքի առանձին անդամները միմյանց հետ, այդ դեպքում մարմնի ձևը և լողաթևերի նմանվող վերջավորությունները ներկայացնում են այնպիսի հատկանիշներ, որոնք ցույց են տալիս իսկական նմանությունը, որովհետև այդ առանձնահատկությունները ամբողջ ընտանիքում որոշ սահմաններում այնքան միանման են, որ մենք չենք կարող կասկածել, որ նրանք ժառանգված են մի ընդհանուր նախահորից։ Նույնը վերաբերում է նաև ձկներին։ Կարելի է բերել զարմանալի նմանության բազմաթիվ դեպքեր առանձին մասերի կամ օրգանների վերաբերյալ, որոնք միանգամայն տարբեր օրգանիզմների մոտ հարմարված են միևնույն գործունեության համար։ Լավ օրինակ է ներկայացնում շան և թասմանիական գայլի (Thylacinus) ծնոտների մերձավոր նմանությունը, կենդանիների, որոնք բնական սիստեմում մեկը մյուսից շատ հեռու են կանգնած։ Բայց այդ նմանությունն արտահայտված է, ընդհանուր գծերով, դուրս ցցված ժանիքներում և սեղանատամների կտրող եզրում, որովհետև իրականության մեջ այդ կենդանիների ատամները շատ խիստ կերպով տարբերվում են. այսպես, շունը վերին ծնոտի յուրաքանչյուր կողմից ունի չորս առջևի և միայն երկու ետևի սեղանատամներ, մինչդեռ թասմանիական գայլը՝ երեք առջևի և չորս ետևի սեղանատամներ։ Բացի դրանից, երկու կենդանիների ետևի սեղանատամները խիստ տարբերվում են հարաբերական մեծությամբ և կազմությամբ, չափահաս կենդանու ատամներին նախորդում են նրանցից միանգամայն տարբերվող կաթնատամները։ Կարող են, իհարկե, ժխտել, որ երկու դեպքում էլ այդ կենդանիների ատամները հաջորդական փոփոխությունների բնական ընտրության միջոցով հարմարվել են միսը պատառոտելու համար. բայց եթե այդ ընդունենք մի դեպքում, ապա ինձ համար անհասկանալի է, որ այդ կարելի է ժխտել մյուս դեպքում։ Եվ ես ուրախ եմ, լսելով, որ այնպիսի ականավոր հեղինակությունը, ինչպիսին է պրոֆ. Ֆլոուերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 161] եկել է միևնույն եզրակացության։
Նախորդ գլուխներից մեկում բերված զարմանալի դեպքերը, որ խիստ տարբեր ձկներ ունեն էլեկտրական օրգաններ, խիստ տարբեր միջատներ՝ լուսատու օրգաններ, խոլորձիները և ասկալիպիադները՝ կպչուն սկավառակներով օժտված ծաղկափոշու զանգվածներ, այս բոլոր դեպքերը վերաբերում են անալոգ, նման առանձնահատկությունների միևնույն կարգին։ Բայց նրանք այն աստիճանի զարմանալի են, որ բերվել են որպես դժվարություններ կամ խոչընդոտներ մեր տեսության համար։ Այս բոլոր դեպքերում հաջողվում է բացահայտել հիմնական տարբերությունը մասերի աճման կամ զարգացման մեջ և սովորաբար նրանց կառուցվածքում և զարգացած վիճակում։ Վերջը նույնն է ստացվում, բայց նրան հասնելու ուղիները, թեև արտաքին տեսքով միևնույնն են թվում, էականորեն տարբեր են։ Այն սկզբունքը, որ նախապես նշվել է անալոգ փոփոխություններ անվան տակ, հավանորեն, հաճախ արտահայտվում է այդ դեպքերում, այսինքն միևնույն դասին պատկանող անդամները, որոնք թեև կապված են միմյանց հետ միայն հեռավոր ազգակցությամբ, իրենց կառուցվածքում այնքան շատ ընդհանուր բան են ժառանգել, որ ընդունակ են նման պատճառների ազդեցության տակ նաև նման կերպով փոփոխվելու. իսկ դա երևի նպաստում է բնական ընտրության միջոցով այնպիսի մասեր կամ օրգաններ ձեռք բերելուն, որոնք շատ նման են իրար՝ անկախ մի ընդհանուր նախահորից անմիջականորեն ստացած ժառանգությունից։
Քանի որ տարբեր դասերի տեսակները հաջորդական թույլ փոփոխությունների միջոցով հարմարվում են մոտավորապես միանման պայմաններում ապրելուն, օրինակ այս երեք միջավայրերից մեկում՝ ցամաքի վրա, օդի կամ ջրի մեջ ապրելուն, ուստի կարող է պատահել, որ մեզ հաջողվի դրա հիման վրա հասկանալ, թե ինչու երբեմն թվական զուգահեռություն է նկատվում տարբեր դասերի ենթախմրերի միջև։ Բնախույզը հանդիպելով այդպիսի զուգահեռության, կարող է զանազան դասերում խմբերի նշանակությունը կամայական կերպով ավելացնելով կամ պակասեցնելով (իսկ մեր փորձը մեզ ասում է, որ այդ նշանակության սահմանումը դեռևս միանգամայն կամայական է), հեշտությամբ ընդարձակել զուգահեռության երևույթները մինչև լայն սահմանների, սրանից էլ, ամենայն հավանականությամբ, առաջացել են դասակարգումների զանազան սիստեմները, որոնց հիմքում դրված են յոթ, հինգ, չորս, երեք և այլ զուգահեռ բաժանումներ։
Կա դեպքերի մի այլ հետաքրքրական կատեգորիա ևս, որտեղ արտաքին մոտ նմանությունը կախված է ոչ թե միանման կենցաղի հարմարվելուց, այլ ձեռք է բերվում պաշտպանողական նպատակներով։ Ես նկատի ունեմ այն դեպքը, երբ որոշ թիթեռներ, ինչպես առաջին անգամ այդ նկարագրել է Բետսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 181] նմանվում են իրենցից բոլորովին տարբեր այլ տեսակների։ Այդ հիանալի դիտողը ցույց է տվել, որ Հարավային Աֆրիկայի որոշ մասերում, որտեղ, օրինակ, Ithomia-ն հանդիպում է, հսկայական խմբերով, մի այլ թիթեռ՝ Leptalis-ը հաճախ հանդիպում է այդ խմբերի հետ խառը. Leptalis-ը գունավոր զոլերով և երանգներով, ինչպես նաև իր թևերի ձևով այնքան նմանվում է Ithomia-ին, որ Բետսը, որի աչքերը տասն և մեկ տարվա ընթացքում վարժեցված էին կոլեկցիաներ հավաքելու, շարունակ սխալվել է, թեպետև միշտ զգույշ է եղել։ Թե նմանվող ձևերը և թե նրանք, որոնց նրանք նմանվում են, բռնելով ու համեմատելով միմյանց հետ, պարզվել է, որ նրանք իրենց կառուցվածքի հիմնական գծերով խիստ տարբեր են լինում և պատկանում են ոչ միայն զանազան սեռերի, այլ հաճախ նույնիսկ զանազան ընտանիքների։ Եթե այդպիսի նմանողությանը հանդիպեր մեկ-երկու անգամ, ապա այն կարելի էր զարմանալի զուգադիպության վերագրել։ Բայց դիտողություններն այն մարզից, որտեղ մի Leptalis-ը նմանվում է Ithomia-ին, փոխադրելով մի այլ մարզ, կարելի է գտնել այլ նմանվող տեսակներ և այլ թիթեռներ, որոնց նմանվում են նրանք, որոնք պատկանում են նույն երկու սեռերին և հավասարապես մոտ նմանություն ունեն միմյանց հետ։ Ընդհանրապես հաշվում են ոչ պակաս քան տասը սեռ, որոնք բովանդակում են այդ թիթեռներին նմանվող տեսակներ։ Թե նմանվողները և թե նրանք, որոնց նմանվում են, միշտ ապրում են միևնույն մարզում. մենք երբեք չենք գտնում նմանվող ձևը մի այլ մարզում ապրելիս, որտեղ չկա այն ձևը, որին նա նմանվում է։ Նմանվողները գրեթե միշտ հազվագյուտ միջատներ են. նմանողություն հարուցող ձևերը գրեթե միշտ հանդիպում են մեծ խմբերով։ Նույն մարզում, որտեղ Leptalis-ի մի տեսակը շատ ճշտությամբ նմանվում է մի Ithomia-ի, երբեմն հանդիպում են նաև այլ թիթեռներ, որոնք նմանվում են նույն Ithomia-ին. այսպիսով, միևնույն վայրում թիթեռների երեք սեռերի տեսակները և նույնիսկ մի ցեց շատ մոտ նմանություն ունեն այն թիթեռի հետ, որ պատկանում է չորրորդ սեռին։ Առանձին հիշատակության արժանի է այն հանգամանքը, որ նմանվող Leptalis-ի ձևերից շատերը, ճիշտ այդպես էլ այն ձևերը, որոնց նմանվում են, ինչպես այդ ապացուցված է աստիճանական անցումների շարքով, միևնույն տեսակի միայն տարատեսակներն են, մինչդեռ մյուսները անկասկածելիորեն առանձին տեսակներ են։ Բայց կարելի է հարցնել, թե ինչո՞ւ որոշ ձևեր դիտվում են որպես նմանողություն հարուցողներ իսկ մյուսները՝ նմանվողներ։ Մ-ր Բետսը բավարար պատասխան է տալիս այդ հարցին, ցույց տալով, որ նմանողություն հարուցող ձևը պահպանում է այն խմբի ընդհանուր տեսքը և գույնը, որին պատկանում է, մինչդեռ «խաբեբաները» փոխել են իրենց գույնն ու ձևը և նման են իրենց ամենամոտ ազգականներին։
Ամենից առաջ մենք ստիպված ենք հարցնել, թե ինչի՞ վերագրել այն, որ որոշ թիթեռներ և ցեցը այդպես հաճախ հագնում են ուրիշ և բոլորովին տարբեր ձևի զգեստ, ինչո՞ւ շփոթության մեջ գցելով բնախույզներին, բնությունը բարեհաճել է իջնել մինչև այդպիսի թատերական արարքների։ Մ-ր Բետսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 181] անկասկած, գտել է ճիշտ բացատրությունը։ Այն ձևերը, որոնց նմանվում են, միշտ աչքի են ընկնում իրենց թվով և պետք է վերին աստիճանի ապահովված լինեն ոչնչացումից, որովհետև այլապես նրանք չէին կարողանա այդպիսի բազմությամբ հանդիպել. ներկայումս հավաքված են շատ ապացույցներ, որ նրանք զզվելի են թռչունների և այլ միջատակեր կենդանիների համար։ Մյուս կողմից, նմանվող ձևերը, որոնք բնակվում են նույն մարզում, համեմատաբար սակավ են և պատկանում են հազվագյուտ խմբերին. այստեղից պետք է եզրակացնել, որ նրանք պետք է տուժած լինեն որևէ պատճառից, որովհետև այլապես, եթե ուշադրության առնենք բոլոր թիթեռների ածած ձվերի քանակությունը, նրանք երեք կամ չորս սերունդների ընթացքում կտարածվեին ամբողջ երկրում։ Այնուհետև եթե այդ հետապնդվող ու հազվագյուտ խմբերից մեկի անդամը ձեռք է բերել զգեստ, որն այնքան նման է լավ պաշտպանված տեսակի զգեստին, որ իր նմանությամբ միշտ խաբում է միջատաբանի փորձված աչքը, նա պետք է խաբի նաև իրեն հետապնդող թռչուններին ու միջատներին և այս կերպով հաճախ խուսափի ոչնչացումից։ Ընդհանրապես կարելի է ասել, որ մ-ր Բետսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 181] լիովին ըմբռնել է այն պրոցեսը, որի միջոցով նմանվող ձևերը դառնում են այնքան նման այն ձևերին, որոնց նրանք նմանվում են, որովհետև նա գտել է, որ Leptalis-ի մի քանի ձևերը, որոնք այնքան բազմազան ձևով ընդօրինակում են այլ թիթեռների, չափազանց փոփոխական են։ Մի մարզում հանդիպել են մի քանի տարատեսակներ, որոնցից մեկը որոշ չափով նմանվել է Ithomia-ին, որը սովորական է նույն մարզում։ Մյուս մարզում հանդիպում են երկու կամ երեք տարատեսակներ, որոնցից մեկը զգալիորեն ավելի սովորական է մյուսներից և դրա հետ միասին ճիշտ կերպով նման է Ithomia-ի մյուս ձևին։ Այսպիսի փաստերի հիման վրա մ-ր Բետսը գալիս է այն եզրակացության, որ Leptalis-ը ամենից առաջ փոփոխական է, իսկ եթե նրա որևէ տարատեսակին հաջողվում է որոշ չափով նմանվել միևնույն մարզի որևէ սովորական թիթեռի, ապա այդ տարատեսակը բարեկեցիկ և քիչ հետապնդվող ցեղի հետ ունեցած նմանության շնորհիվ ավելի լավ շանսեր ունի, որպեսզի խուսափի գիշատիչ թռչունների և միջատների հետապնդումից և, հետևաբար, ավելի հաճախ է պահպանվում, «նմանության ավելի քիչ կատարյալ աստիճանները սերնդե սերունդ ոչնչանում են, և միայն մյուսներին է վիճակվում իրենցից հետո սերունդ թողնելու հնարավորությունը»։ Այսպիսով, այս դեպքը բնական ընտրության հիանալի օրինակ է։
Մ-րներ Ուոլլեսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] և Տրայմենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 256] իրենց հերթին նկարագրել են նմանողության մի քանի նույնքան զարմանալի դեպքեր Մալայան արշիպելագի և Աֆրիկայի թիթեռների և մի քանի այլ միջատների մոտ։ Մ-ր Ուոլլեսը հայտնաբերել է նաև նման մի դեպք թռչունների մոտ, բայց ավելի խոշոր կաթնասունների համար այդպիսի օրինակ մենք չգիտենք։ Այլ կենդանիների համեմատությամբ միջատների մեջ նմանողության մեծ տարածվածությունը հավանորեն նրանց փոքրության հետևանքն է. միջատները չեն կարող պաշտպանվել, բացառությամբ խայթոց ունեցող տեսակների, և ես երբեք չեմ լսել, որ այդպիսի միջատները նմանվեին ուրիշներին, մինչդեռ նրանց նմանվում են ուրիշները. միջատները չեն կարող նաև թռիչքի կամ փախուստի միջոցով ազատվել նրանց որսացող խոշոր կենդանիներից. ուրեմն, փոխաբերաբար ասած, թույլ էակների մեծամասնության նման, նրանք պետք է դիմեն խաբեության ու կեղծիքի։
Կարելի է նկատել, որ նմանողության պրոցեսը հավանորեն երբեք չի սկսվել այնպիսի երկու ձևերի միջև, որոնք իրենց գույնով շատ են տարբերվում. բայց սկսվելով որոշ չափով արդեն նման տեսակների մոտ, նա կարող է վերոհիշյալ եղանակով հանգել շատ լիակատար նմանության, եթե այդ օգտակար է։ Եվ եթե այն ձևը, որին նմանվում են, որևէ ներգործության ազդեցության տակ հաջորդականորեն ու աստիճանաբար փոփոխվում է, ապա նաև նմանվող ձևը կարող է ընթանալ միևնույն ուղիով և փոփոխվել այն աստիճանի, որ կարող է վերջ ի վերջո ձեռք բերել այնպիսի ձև ու գույն, որ ամենևին նման չէ այն ընտանիքի այլ անդամների ձևին ու գույնին, որին ինքն է պատկանում։ Սակայն այդ տեսակետից մեզ հանդիպում է որոշ դժվարություն, որովհետև մի քանի դեպքերում անհրաժեշտ է ընդունել, որ մի քանի տարբեր խմբերի հեռավոր անդամները՝ նախքան նրանց տարամիտման մինչև այժմյան աստիճանին հասնելը, պատահականորեն նմանվել են մի այլ հովանավորյալ խմբի անդամներին այնքան բավարար չափով, որպեսզի դրանով որոշ պաշտպանություն գտնեն․ դա էլ պետք է հիմք ծառայեր հետագա ավելի կատարյալ նմանություն ձեռք բերելու համար։
ՕՐԳԱՆԻԶՄՆԵՐԸ ՇԱՂԿԱՊՈՂ ԱԶԳԱԿՑՈՒԹՅԱՆ ԲՆՈՒՅԹԻ ՄԱՍԻՆ
Քանի որ ընդարձակ սեռերին պատկանող տիրապետող տեսակների փոփոխված հետնորդները հակված են ժառանգելու այն առավելությունները, որոնց շնորհիվ նրանց խմբերը դարձել են ընդարձակ և նրանց ծնողները տիրապետող, ապա ղրանով իսկ նրանք հնարավորություն են ստացել լայնորեն տարածվելու և բնության տնտեսության մեջ ավելի ու ավելի մեծ քանակությամբ տեղ գրավելու համար։ Յուրաքանչյուր դասի սահմաններում ավելի տարածված ու տիրապետող խմբերը ձգտում են ավելանալ ծավալով և դրա հետևանքով ճնշել ավելի փոքր ու թույլ խմբերին։ Դրանով մենք կարող ենք բացատրել այն, որ բոլոր օրգանիզմները, ինչպես այժմյանները, այնպես էլ մահաջնջվածները, մտնում են սակավաթիվ կարգերի և ավելի ևս փոքրաթիվ դասերի մեջ։ Ցույց տալու համար, թե որքան քիչ է խոշոր խմբերի թիվը և որքան նրանք լայնորեն են տարածված երկրագնդի վրա, բերենք այն ուշագրավ փաստը, որ Ավստրալիայի հայտնագործումը չի ավելացրել ոչ մի միջատ, որը պատկանի մի նոր մեծ խմբի, և ինչ վերաբերում է բույսերին, ապա, ինչպես ասում է դ-ր Հուկերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 28] տվել է միայն երկու կամ երեք փոքրիկ ընտանիքներ։
Երկրաբանական հաջորդականության վերաբերյալ գլխում, ելնելով այն դրույթից, որ յուրաքանչյուր խումբ իր փոփոխության երկարատև ժամանակաշրջանի ընթացքում ընդհանրապես շատ խիստ տարամիտվում է հատկանիշներով, ես աշխատել եմ պարզել, թե ինչու հին օրգանիզմները հաճախ ունեն հատկանիշներ, որոնք որոշ չափով անցումային են գոյություն ունեցող խմբերի և նրանց միջև։ Քանի որ հին և միջակա քիչ ձևեր միայն ներկա ժամանակին փոխանցել են շատ փոքր չափով փոփոխված հաջորդներ, ուստի վերջիններս կազմում են, այսպես կոչված, շեղվող կամ միջակա հատկանիշներ ունեցող մերձավոր տեսակներ։ Որքան որևէ ձև ավելի ուժեղ կերպով է շեղվում, այնքան ավելի շատ պիտի լինի այն ձևերի թիվը, որոնք շաղկապում են նրան արդեն մահաջնջված և իսպառ անհետացած այլ ձևերի հետ։ Որ այդպիսի միջին հատկանիշներ ունեցող խմբերը շատ դաժանորեն ենթարկվել են մահաջնջման, դրա համար մենք ունենք որոշ ապացույց այն բանում, որ նրանք գրեթե միշտ ներկայացված են չափազանց սակավաթիվ տեսակներով. և բացի այդ, այդպիսի տեսակները սովորաբար շատ ուժեղ կերպով զանազանվում են միմյանցից, որն իր հերթին ցույց է տալիս նրանց մահաջնջումը։ Օրինակ, բադակտուցի (Ornithorhynchus) և լեպիդոսիրենի (Lepidosiren) սեռերը կլինեին ոչ պակաս շեղվող, եթե նրանք ունենային մի դյուժին տեսակներ, փոխանակ ունենալու մեկ, երկու կամ երեք տեսակ, ինչպես այդ տեսնում ենք այժմ։ Ես կարծում եմ, մենք կարող ենք այդ փաստը բացատրել միայն ընդունելով, որ շեղվող խմբերի ձևերը պարտություն են կրել ավելի օժտված մրցակիցներից և դեռ պահպանել են իրենց սակավաթիվ ներկայացուցիչները բացառիկ բարենպաստ պայմաններում։
Մ-ր Ուոտերհաուզը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 87] նկատել է, որ եթե կենդանիների որևէ խմբի անդամներից մեկը նմանություն է հանդես բերում բոլորովին մի այլ խմբի հետ, ապա շատ դեպքում այդ նմանությունն ընդհանուր է, այլ ոչ թե մասնահատուկ։ Այսպես, ըստ Ուոտերհաուզի, բոլոր կրծողներից վիսկաշան ամենից ավելի մոտ է պարկավորներին. բայց նրա շփման կետերն այդ կարգի հետ ընդհանուր են, այսինքն՝ պարկավորների մի այնպիսի տեսակ չկա, որին նա ավելի մոտ լիներ, քան մի ուրիշին։ Քանի որ կառուցվածքի նման առանձնահատկությունները բացատրվում են իսկական ազգակցությամբ, այլ ոչ թե պարզ հարմարեցմամբ, ուստի նրանք, մեր տեսակետների համաձայն, պետք է ժառանգված լինեն մի ընդհանուր նախածնողից։ Այդ պատճառով մենք կամ պետք է ենթադրենք, որ բոլոր կրծողները և այդ թվում էլ վիսկաշան անջատվել են որևէ հին պարկավոր ձևից, որը բնականաբար իր հատկանիշներով պետք է դառնար քիչ թե շատ անցումային բոլոր գոյություն ունեցող պարկավորների վերաբերմամբ, կամ թե, որ կրծողներն ու պարկավորները ծագում են առել մի ընդհանուր նախածնողից և, շարունակ ավելի ու ավելի հեռանալով իրարից, մինչև հիմա մեծ փոփոխություններ են կրել։ Այս տեսակետներից որ մեկին էլ հետևենք, մենք պետք է ենթադրենք, որ վիսկաշան ժառանգականության միջոցով իր հին նախածնողից ավելի շատ առանձնահատկություններ է պահպանել, քան մյուս կրծողները. այդ պատճառով էլ նա առանձին առնչություն չունի այժմյան պարկավորներից որևէ մեկի հետ, այլ ուղղակի կապված է բոլոր կամ գրեթե բոլոր պարկավորների հետ, մասամբ պահպանելով նրանց ընդհանուր նախածնողի կամ խմբի հին անդամներից մեկի հատկանիշները։ Մյուս Կողմից, ըստ Ուոտերհաուզի դիտողության, բոլոր պարկավորներից վոմբատը (Phascolomys) ամենից ավելի նման է ոչ թե կրծողներից որևէ մեկին, այլ ընդհանրապես բոլոր կրծողներին։ Բայց այս դեպքում ամենայն իրավամբ կարելի է ենթադրել, որ այդ նմանությունը միայն անալոգիական է, որը ցույց է տալիս, որ վոմբատը հարմարվել է կրծողների կենցաղին։ Դը-Կանդոլ ավագը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 67] մոտավորապես նույնպիսի դիտողություններ է արել բույսերի զանազան ընտանիքների միջև եղած նմանության ընդհանուր բնույթի մասին։
Մի ընդհանուր նախածնողից առաջացած տեսակների բազմացման և հատկանիշների աստիճանական տարամիտման օրենքի և դրա հետ միասին ժառանգականության միջոցով նրանց մեջ ընդհանուր հատկանիշների պահպանման հիման վրա մենք կարող ենք հասկանալ, թե ինչքան արտասովոր կերպով բարդ ու բազմակողմանի են այն ընդհանուր գծերը, որոնցով շաղկապվում են միևնույն ընտանիքի անդամները կամ ավելի ևս ընդարձակ խմբերը միմյանց հետ։ Մահաջնջման միջոցով այժմ զանազան խմբերի և ենթախմբերի բաժանված ամբողջ ընտանիքի ընդհանուր նախածնողը պետք է փոխանցեր իր հատկանիշներից մի քանիսը բոլոր տեսակներին, որոնք տարբեր ուղղությամբ ու տարբեր աստիճանի փոփոխված են, և այդ պատճառով էլ տեսակները պետք է միմյանց հետ կապված լինեն ազգակցության շրջանաձև գծերով, որոնք ունեն զանազան երկարություն (այդ կարելի է տեսնել դիագրամի վրա, որը մենք այնպես հաճախ վկայակոչում ենք) և սկիզբ են առնում բազմաթիվ նախնիների միջից։ Քանի որ ծննդաբանական ծառի օգնությամբ նույնիսկ դժվար է և առանց նրան գրեթե անհնարին է ապացուցել որևէ հին նշանավոր ընտանիքի բազմաթիվ հաջորդների միջև եղած արյունակցական ազգակցությունը, ուստի հեշտ է հասկանալ այն արտասովոր դժվարությունները, որոնց հանդիպում է բնախույզը առանց դիագրամի օգնության զանազան հարաբերակցություններ նկարագրելիս, որոնք նկատվում են որևէ մեծ բնական դասի ապրող և մահաջնջված անդամների միջև։
Մահաջնջումը, ինչպես մենք տեսանք չորրորդ գլխում, կարևոր դեր է կատարել յուրաքանչյուր դասի զանազան խմբերի միջև եղած տարածությունների առաջացման ու լայնացման գործում։ Դրա հիման վրա մենք կարող ենք բացատրել մի ամբողջ դասի մեկուսացածությունը մի ուրիշից, օրինակ՝ թռչուններին մյուս բոլոր ողնաշարավորներից, ենթադրելով, որ հին շատ ձևեր բոլորովին անհետացել են, օրինակ նրանք, որոնց միջոցով թռչունների հեռավոր նախնիները սկզբում շաղկապվել են այլ ողնաշարավորների դեռ այն ժամանակ ավելի քիչ դիֆերենցված դասերի նախնիների հետ։ Անհամեմատ ավելի քիչ է եղել այն ձևերի մահաջնջումը, որոնք մի ժամանակ ձկներին կապել են երկկենցաղների հետ։ Է՛լ ավելի փոքր է մահաջնջումը որոշ դասերի, օրինակ խեցեմորթների մեջ, որովհետև այստեղ զարմանալիորեն տարբեր ձևեր դեռևս կապվում են միմյանց հետ մերձավոր ձևերի երկար և միայն մասամբ ընդհատվող շղթայով։ Մահաջնջումը միայն պարագծել է խմբերը, բայց բնավ չի ստեղծել նրանց, որովհետև, եթե նախկին ժամանակներում երկրի վրա ապրած բոլոր ձևերը հանկարծ կենդանանայինք միանգամայն անհնարին կշիներ ցույց տալ առանձին խմբերի սահմանները, բայց բնական դասակարգություն կամ առնվազն բնական խմբավորում հնարավոր կլիներ։ Մենք այդ կտեսնենք, դիմելով դիագրամին։ A-ից մինչև L տառերը պետք է պատկերեն սիլուրիական տասնմեկ սեռերը, ըստ որում նրանցից մի քանիսից առաջացել են փոփոխված հաջորդների մեծ խմբեր, որոնք ունեն դեռևս ապրող միացուցիչ օղակներ գլխավոր ու երկրորդական ճյուղերի միջև․ այդ կապակցող օղակները ավելի չեն նրանցից, որոնք կան այժմ գոյություն ունեցող տարատեսակների միջև։ Այդ դեպքում միանգամայն անհնարին է տալ այնպիսի սահմանումներ, որոնցով զանազան խմբերի զանազան անդամները կարելի կլիներ տարբերել նրանց ավելի մերձավոր նախածնողներից և հետնորդներից։ Սակայն դիագրամում խմբավորումը այնուամենայնիվ պետք է պահպանվի և լինի բնական, որովհետև ժառանգականության օրենքով բոլոր ձևերը, որ առաջացել են, օրինակ, A-ից, պետք է մի ինչ որ ընդհանուր բան ունենան։ Մենք կարող ենք ծառի վրա տարբերել այս կամ այն ճյուղը, թեև երկճյուղավորման տեղում նրանք մոտենում են ու միաձուլվում։ Ինչպես ես ասել եմ, մենք չենք կարող սահմանումներ տալ զանազան խմբերի համար, բայց մենք կարող ենք ցույց տալ տիպերը կամ այնպիսի ձևեր, որոնք իրենց մեջ միացնում են յուրաքանչյուր մեծ կամ փոքր խմբի հատկանիշների ամենամեծ քանակություն և այդպիսով մեղ տալիս են ընդհանուր պատկերացում նրանց միջև եղած տարբերությունների նշանակության մասին։ Դա հենց այն է, որին մենք կհանգեինք, եթե մեզ երբևիցե հաջողվեր հավաքել մի որոշ դասի բոլոր ձևերը, որոնք երբևիցե գոյություն են ունեցել ժամանակի և տարածության մեջ։ Իհարկե, մենք այդպիսի հաջողության երբեք չենք հասնի. այնուամենայնիվ որոշ դասերում մենք ձգտում ենք այդ նպատակին, և Միլն Էդվարդսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 86] վերջերս իր մի հետաքրքրական հոդվածում ապացուցում է տիպերի ուսումնասիրության կարևոր նշանակությունը անկախ նրանից, թե կարելի է բաժանել ու որոշել այն խմբերը, որոնց պատկանում են այդ տիպերը։ Վերջապես, մենք տեսանք, որ բնական ընտրությունը, որ բխում է գոյության կռվից և գրեթե անխուսափելիորեն հանգում է ձևերի մահաջնջման և մի նախածնողական տեսակի հաջորդների մեջ հատկանիշների տարամիտման, բացատրում է բոլոր օրգանիզմների հարաբերակցության մեջ եղած մի մեծ ընդհանուր առանձնահատկության գոյությունը, այսինքն՝ նրանց տեղաբաշխումն ըստ ստորադաս նշանակություն ունեցող խմբերի։ Մենք ծագումն օգտագործում ենք երկու սեռի և ամեն հասակի անհատներին մի տեսակի մեջ զետեղելու համար, թեև նրանց միջև, կարող է պատահել, շատ քիչ ընդհանուր բան կա․ մենք ծագումը օգտագործում ենք մեզ լավ հայտնի տարատեսակների տեղը որոշելու համար, ինչքան էլ որ նրանք ուժեղ կերպով տարբերվելիս լինեն ծնողներից․ և ես կարծում եմ, որ ծագումն է այն գաղտնի կապը, որ հասկանում են բնախույզները բնական սիստեմ անվան տակ։ Օրգանիզմների բնական սիստեմի գաղափարը միայն, որքան նա կատարյալ է և խմբավորման հիմքում բովանդակում է արյունակցական ազգակցությունը՝ նմանության տարբեր աստիճաններով միասին, որոնք արտահայտվում են սեռեր, ընտանիքներ, կարգեր և այլ տերմիններով, մեզ հնարավորություն է տալիս հասկանալու այն կանոնները, որոնցով պետք է ղեկավարվել մեր դասակարգության ընթացքում։ Մենք կարող ենք նաև հասկանալ, թե ինչու որոշ նմանվող առանձնահատկությունների նշանակությունն ավելի կարևոր է, քան մյուսներինը․ թե ինչու մենք օգտագործում ենք թերաճ ու անօգուտ օրգանները կամ կասկածելի ֆիզիոլոգիական նշանակություն ունեցող այլ օրգանները․ թե ինչու զանազան խմբերի միջև ազգակցություն որոնելիս մենք ընդհանրապես մի կողմ ենք նետում անալոգ կամ հարմարողական հատկանիշները և օգտագործում ենք դրանք, երբ որոշում ենք միևնույն խմբի ձևերի միջև եղած ազգակցությունը։ Վերջապես մենք կարող ենք հեշտությամբ տեսնել, թե ինչու բոլոր ապրող և մահաջնջված ձևերը կարելի է համախմբել փոքրաթիվ խոշոր խմբերում և թե ինչու յուրաքանչյուր դասի առանձին անդամները շաղկապված են միմյանց հետ նմանության վերին աստիճանի բարդ և բազմակողմանի գծերով։ Ամենայն հավանականությամբ մեզ երբեք չի հաջողվի բաց անել ազգակցական այն կապերի աներևակայելի կերպով խճճված ցանցը, որոնք շաղկապում են որևէ դասի անդամներին միմյանց հետ. բայց նկատի ունենալով մի որոշակի խնդիր և բացատրության համար չդիմելով ստեղծագործության որևէ անիմանալի պլանի վկայակոչությանը, մենք կարող ենք անկասկածելի, թեև դանդաղ, հաջողության հույս ունենալ։
Պրոֆ. Հեկկելը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 257] իր «Generelle Morphologie»[73]. և այլ աշխատություններում վերջերս իր ընդարձակ դիտելիքներն ու տաղանդը նվիրել է այն հարցի ուսումնասիրությանը, որ նա անվանում է ֆիլոգենիա, կամ տոհմագրություն, որը շաղկապում է բոլոր օրգանական էակներին միմյանց հետ։ Զանազան ծննդաբանական շարքեր կազմելիս նա հենվում է գլխավորապես սաղմնաբանական առանձնահատկությունների վրա, բայց օգտագործում է նաև հոմոլոգ և թերաճ օրգանները, ինչպես և այն դարաշրջանների հաջորդականությունը, որոնց ընթացքում կյանքի զանազան ձևերը, ինչպես կարծում են, առաջին անգամ երևան են եկել զանազան երկրաբանական ֆորմացիաներում։ Նա դրանով համարձակորեն խիզախել է մի մեծ նախաձեռնության և մեզ ցույց է տվել, թե ապագայում ինչպես պետք է մշակել դասակարգման հարցը։
ՄՈՐՖՈԼՈԳԻԱ
Մենք տեսանք, որ միևնույն դասի անդամները, անկախ իրենց վարած կենցաղից, կազմվածքի ընդհանուր պլանով իրար նման են։ Այդ նմանությունը հաճախ արտահայտում են «պլանի միասնականություն» տերմինով կամ բառերով, որ միևնույն դասի զանազան տեսակների որոշ մասերը և օրգանները հոմոլոգ են։ Սրան վերաբերող բոլոր հարցերը պատկանում են մորֆոլոգիային, որը ներկայացնում է բնապատմության ամենահետաքրքիր բաժիններից մեկը և, առհասարակ կարելի է ասել, նրա էական մասը։ Ի՞նչը կարող է ավելի հետաքրքրական լինել նրանից, որ բռնելու համար նախանշված մարդու ձեռքը, խլուրդի փորող թաթը, ձիու ոտքը, դելֆինի լողաթաթը, չղջիկի թևը կառուցված են միևնույն օրինակով և պարունակում են միանման ոսկորներ, միևնույն հարաբերական դասավորությամբ։ Ինչքան հետաքրքրական են ավելի պակաս կարևոր, բայց և այնպես այն զարմանալի փաստերը, որ կենգուրուի ետևի ոտները այդքան լավ հարմարված են արձակ հարթավայրերում ցատկում կատարելու համար․ մագլցող, տերևներով սնվող կոալի ոտքերը հավասարապես լավ հարմարված են ծառերի ճյուղերը գրկելու համար. գետինը փորող, միջատներով ու արմտիքով սնվող բենդիկուտի և ավստրալիական մի քանի այլ պարկավորների թաթերը կառուցված են միևնույն վերին աստիճանի զարմանալի տիպի համաձայն, որտեղ երկրորդ և երրորդ մատների ոսկորները չափազանց բարակ են և ծածկված են ընդհանուր մաշկային ծածկոցով, այնպես որ նմանվում են երկու ճիրան ունեցող մեկ մատի։ Չնայած կառուցվածքի նմանության, ակնհայտ է, որ այդ զանազան կենդանիների ետևի ոտները ծառայում են ամենատարբեր նպատակների համար, ինչպիսիք միայն կարելի է պատկերացնել։ Բայց փաստերի այս ամբողջ խումբը ավելի ևս զարմանալի է դառնում այն պատճառով, որ ամերիկյան պարկամկները, որոնք մոտավորապես նույն կենցաղն են վարում, ինչ որ նրանց ավստրալիական ազգակիցները, ունեն սովորական ձևով կառուցված վերջավորություններ։ Պրոֆ. Ֆլոուերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 161] որից մենք վերցրել ենք այս փաստերը, վերջում այսպիսի դիտողություն է անում. «Մենք կարող ենք այդ անվանել տիպի պահվածություն՝ չմտնելով ավելի խոր բացատրության մեջ, և ավելացնում է. «Արդյոք դա իրո՞ք ցույց չի տալիս իսկական ազգակցություն, մի ընդհանուր ծնողից ժառանգական փոխանցում»։
Ժոֆֆրուա Սենտ-Իլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 16] վճռականորեն պնդում էր հոմոլոգ մասերի հարաբերական դրության կամ հարաբերակցության կարևոր նշանակության վրա. նրանք կարող են իրենց ձևով ու չափերով ծայրահեղորեն փոփոխվել, բայց միշտ միացած են միևնույն անփոփոխ եղանակով։ Օրինակ, մենք երբեք չենք գտնի, որ բազկի և նախաբազկի կամ ազդրի և սրունքի ոսկրները տեղափոխվեն։ Ուստի միևնույն անունները կարելի է տալ չափազանց տարբեր կենդանիների հոմոլոգ ոսկիներին։ Նույն կարևոր օրենքը մենք տեսնում ենք միջատների բերանի մասերի կառուցվածքում. ի՞նչը կարող է ավելի տարբեր լինել, քան գիշերաթիթեռի ծայր աստիճան երկար, պարուրաձև ոլորված կնճիթը, մեղվի կամ փայտոջիլի յուրահատուկ ձևով կառուցված կնճիթը և բզեզի մեծ ծնոտները։ Բայց և այնպես այդ բոլոր օրգանները, որոնք ծառայում են շատ տարբեր նպատակների համար, առաջացել են վերին շրթունքի, վերին ծնոտների և երկու զույգ ստորին ծնոտների անսահման փոփոխությունների միջոցով։ Միևնույն օրենքն է ղեկավարում խեցեմորթների բերանի և վերջավորությունների մասերի կառուցվածքը։ Նույնը ճիշտ է նաև բույսերի ծաղիկների վերաբերմամբ։
Ոչինչ չի կարող ավելի անհուսալի լինել, քան միևնույն դասի անդամների մոտ եղած կառուցվածքի այդ ընդհանրությունը օգտակարության, կամ վախճանական պատճառների ուսմունքի տեսակետից բացատրելու փորձը։ Այդպիսի փորձի անհուսալիությունը պարզ կերպով մատնանշել է Օուենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] իր «Վերջավորությունների բնույթը» վերին աստիճանի հետաքրքրական աշխատության մեջ։ Հետևելով այն սովորական տեսակետին, որ յուրաքանչյուր էակ անկախ կերպով է ստեղծվել, մենք կարող ենք միայն ընդունել փաստերը և ասել, որ արարչին հաճո է եղել յուրաքանչյուր մեծ դասի, բոլոր կենդանիներին ու բույսերին կառուցել մի ընդհանուր պլանով, բայց դա գիտական, բացատրություն չէ։ Դրա փոխարեն՝ շատ պարզ բացատրություն է տալիս հաջորդական թույլ փոփոխությունների ընտրության տեսությունը, որոնցից յուրաքանչյուրը որոշ չափով օգտակար է փոփոխվող ձևին, բայց հաճախ պայմանավորվում է կազմվածքի մյուս մասերի հետ ունեցած կոռելյացիայով։ Նման փոփոխությունների ժամանակ հիմնական պլանի փոփոխության և մասերի տեղափոխության հակումը կամ քիչ է, կամ բոլորովին չի լինում։ Վերջավորությունների ոսկրները կարող են որոշ չափով կարճանալ ու տափակել, միաժամանակ ծածկվելով փափուկ մասերի ավելի հաստ շերտերով և այդ դեպքում փոխարկվում են լողաթևի. կամ թե թաղանթով օժտված վերջավորության բոլոր կամ մի քանի ոսկրները կարող են որոշ չափով երկարել, որի հետ միասին մեծանում է նրանց կապող թաղանթը և վերջավորությունը կարող է դառնալ թև. բայց այդ բոլոր փոփոխությունները չեն ձգտում փոփոխել ոսկրների հիմնական կառուցվածքը կամ մասերի հարաբերակցությունը։ Եթե մենք ենթադրենք, որ բոլոր կաթնասունների, թռչունների և սողունների ամենահին նախածնողի, որին կարելի է անվանել նախատիպ, վերջավորությունները կառուցված են եղել այժմ գոյություն ունեցող հիմնական պլանի համաձայն, անկախ նրանց կատարած ֆունկցիաներից, մենք իսկույն կըմբռնենք բոլոր դասերում վերջավորությունների հոմոլոգ կառուցվածքի ամբողջ իմաստը։ Ճիշտ նույնպես միջատների բերանի մասերի վերաբերմամբ մեզ հարկավոր է միայն ենթադրել, որ նրանց նախածնողն ունեցել է վերին շրթունք, վերին ծնոտներ և երկու զույգ ստորին ծնոտներ, ընդ որում թերևս այդ մասերը եղել են շատ պարզ կազմությամբ, և բնական ընտրությունը կբացատրի մեզ միջատների բերանի մասերի կազմության և կատարած դերերի ամբողջ անսահման բազմազանությունը։ Սակայն հասկանալի է, որ օրգանի կառուցվածքի հիմնական պլանը կարող է այն աստիճանի թաքնված լինել, որ բոլորովին կանհետանա կամ թերազարգացման, կամ որոշ մասերի լիակատար ռեդուկցիայի, մյուսների միաձուլման, երրորդ մասերի կրկնապատկման և ընդհանրապես թվի ավելացման հետևանքով՝ բոլորն էլ այնպիսի փոփոխություններ են, որոնք, ինչպես մենք գիտենք, գտնվում են հնարավորության սահմաններում։ Մահաջնջված հսկայական ծովային մողեսների լողաթևերի և մի քանի խեցեմորթների ծծող բերանամասերի մեջ հիմնական պլանը մասամբ թվում է մի քիչ քողարկված։
Կա էլի մի հավասար չափով հետաքրքրական հարց, որը դարձյալ սրան է վերաբերում, այն է՝ սերիական հոմոլոգիաները կամ միևնույն անհատի զանազան մասերի ու օրգանների համեմատությունը, այլ ոչ թե միևնույն դասի զանազան անդամների միևնույն մասերի և օրգանների համեմատությունը։ Ֆիզիոլոգների մեծամասնությունը կարծում է, թե գանգի ոսկրները հոմոլոգ են, այսինքն՝ թվով և հարաբերական դասավորությամբ համապատասխանում են բրոշ թվով ողերի հիմնական մասերին։ Բոլոր բարձրակարգ ողնաշարավորների խմբերում առջևի և հետևի վերջավորությունները հոմոլոգ են։ Ճիշտ այդպես էլ հոմոլոգ են խեցեմորթների զարմանալի բարդ ծնոտները և վերջավորությունները։ Գրեթե ամեն մեկին հայտնի է, որ ծաղկի մեջ բաժակաթերթերի, պսակաթերթերի, առէջքների և վարսանդների հարաբերական դիրքը, ինչպես նաև նրանց կառուցվածքի մանրամասնությունները հասկանալի են դառնում միայն այն դեպքում, երբ մենք նրանց համարում ենք պարուրաձև դասավորություն ունեցող փոփոխված տերևներ։ Այլանդակ բույսերի վրա մենք հաճախ ուղղակի ցուցումներ ենք գտնում մի օրգանը մյուսի փոխարկվելու հնարավորության մասին, և մենք կարող ենք նկատել ծաղիկների, ինչպես նաև խեցեմորթների և այլ կենդանիների զարգացման վաղ, կամ սաղմնային ստադիաներում, որ զարգացման հետ զուգընթաց չափազանց տարբերվող օրգանները սկզբում բոլորովին նման են իրար։
Ինչքա՜ն անբացատրելի են սերիական հոմոլոգիաների դեպքերը արարչագործական գործունեության նկատմամբ եղած սովորական տեսակետով։ Ինչո՞ւ գլխուղեղը պարփակված է տուփի մեջ, որը կազմված է այնքան բազմաթիվ և յուրահատուկ ոսկրներից, որոնք ըստ երևույթին ներկայացնում են ողեր։ Ըստ Օուենի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] դիտողության՝ այն օգուտը, որ առաջանում է կաթնասունների ծննդաբերության ժամանակ առանձին ոսկիների որոշ շարժականությունից, երբեք չի բացատրում թռչունների և սողունների գանգի նույն կառուցվածքը։ Ինչո՞ւ միևնույն ոսկրները ստեղծվել են չղջիկի թևի և հետևի վերջավորության կառուցման համար, որոնք գործածվում են այնքան տարբեր նպատակների համար, ինչպես թռչելն ու ման գալը։ Ինչո՞ւ բազմաթիվ հավելուկներով չափազանց բարդ կառուցված բերան ունեցող խեցեմորթները դրա հետ միասին միշտ ավելի քիչ ոտներ են ունենում, և ընդհակառակը, նրանք, որոնք շատ ոտներ ունեն, բերանի մասերի ավելի պարզ կառուցվածք են ունենում։ Ինչո՞ւ յուրաքանչյուր ծաղկի մեջ բաժակաթերթերը, պսակաթերթերը, առէջքները և վարսանդները, թեև բոլորովին տարբեր դեր են ունենում, կառուցված են մի հիմնական տիպարով։
Բնական ընտրության տեսության միջոցով մենք որոշ չափով կարող ենք պատասխանել այդ հարցերին։ Մենք կարիք չունենք խորհելու, թե ինչպես որոշ կենդանիների մարմինը բաժանվել է սկզբում մի շարք հատվածների կամ թե ինչպես նրանց մոտ առաջացել են աջ և ձախ կողմերը համապատասխան օրգաններով միասին, որովհետև այդպիսի հարցերը գրեթե դուրս են գալիս հետազոտության սահմաններից։ Սակայն հավանական է, որ որոշ դեպքերում սերիական կառուցվածքը հանդիսանում է բջիջների բաժանման միջոցով բազմանալու արդյունք, որն իր հերթին պայմանավորում է այդպիսի բջիջներից զարգացող մասերի շատացում։ Մեզ համար բավական է իմանալ, որ միևնույն մասերի կամ օրգանների անսահման կրկնությունը, ըստ Օուենի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] դիտողության, բնորոշում է բոլոր ստորակարգ կամ քիչ մասնագիտացած ձևերին. այդ պատճառով ողնաշարավորների անհայտ նախածնողը հավանորեն ունեցել է շատ ողեր, հոդվածոտանիների նախածնողը՝ շատ հատվածներ, ծաղկավոր բույսերի նախածնողը՝ շատ տերևներ, որոնք դասավորված են եղել մեկ կամ ավելի պարուրներով։ Ճիշտ այդպես էլ մենք արդեն տեսանք, որ մի քանի անգամ կրկնվող մասերը մեծ հակում ունեն փոփոխվելու ոչ միայն թվով, այլև ձևով։ Հետևաբար, այդպիսի մասերը, որոնք արդեն գոյություն ունեն մեծ թվով և լինելով չափազանց փոփոխական, բնականաբար նյութ են հանդիսանում շատ բազմազան նպատակների հարմարվելու համար, սակայն, ընդհանրապես նրանք պետք է ժառանգականության ուժով պահպանեին իրենց նախասկզբնական կամ հիմնական նմանության ակնհայտ հետքերը։ Եվ նրանք կարող էին պահպանել այդ նմանությունը, մանավանդ, որ այն վարիացիաները, որոնք բնական ընտրության ազդեցությամբ նրանց հետագա փոփոխությունների համար հիմք են կազմել, հենց սկզբից հակում են ունեցել իրար նմանվելու, որովհետև այդ մասերը զարգացման վաղ ստադիաներում նման են եղել և գտնվել են մոտավորապես միանման պայմաններում։ Այդպիսի մասերը ավելի կամ պակաս չափով փոփոխվելով, բայց դեռ չկորցնելով իրենց ընդհանուր ծագման հետքերը, դարձել են սերիական հոմոլոգ մասեր։
Կակղամորթների ընդարձակ դասի մեջ, չնայած տարբեր տեսակների մոտ հոմոլոգ մասերի գոյությանը, կարելի է շարքային հոմոլոգիաների միայն քիչ օրինակներ ցույց տալ, ինչպիսիք են, օրինակ, խիտոնների խեցիները, այսինքն մենք միայն հազվագյուտ դեպքերում կարող ենք ասել, որ միևնույն անհատի մի մասը հոմոլոգ է նրա մյուս մասին։ Եվ դա մեզ հասկանալի է, որովհետև կակղամորթների մեջ, նույնիսկ այդ դասի ստորակարգ ներկայացուցիչների մեջ, մենք բոլորովին չենք գտնում որևէ մասի այնպիսի անսահմանափակ կրկնություն, որին մենք հանդիպում ենք կենդանական ու բուսական աշխարհի մյուս մեծ խմբերում։
Բայց մորֆոլոգիայի խնդիրներն անհամեմատ ավելի բարդ են, քան թվում է առաջին հայացքից, ինչպես վերջերս ակնառու կերպով այդ ցույց է տվել մ-ր Ռեյ Լանկեստերն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 134] իր մի ուշագրավ հոդվածում, որտեղ նա մատնանշել է էական տարբերություններ դեպքերի զանազան կատեգորիաների միջև, որոնք մինչև այդ բնախույզների կողմից միանման ձևով համարվել են հոմոլոգներ։ Նա առաջարկում է հոմոգեն անվանել այն մասերը, որոնք նման են մի ընդհանուր նախածնողից փոփոխությունների միջոցով առաջացած տարբեր կենդանիների մոտ, և հոմոպլաստիկ անվանել նրանց, որոնք այդ ձևով չի կարելի բացատրել։ Օրինակ, նա կարծում է, որ թռչունների և կաթնասունների սիրտը միանգամայն հոմոգեն է, որովհետև ժառանգված է մի ընդհանուր նախածնողից, բայց սրտի խոռոչները երկու դասերում միայն հոմոպլաստիկ են, այսինքն զարգացել են անկախ կերպով։ Մ-ր Լանկեստերն այնուհետև նշում է միևնույն անհատի մարմնի աջ և ձախ կողմերի մասերի և իրար հաջորդող հատվածների մերձավոր նմանությունը, այստեղ մենք ունենք այնպիսի մասերի օրինակ, որոնց ընդհանրապես անվանում են հոմոլոգ, բայց սրանք ոչ մի առնչություն չունեն մի նախածնողից զանազան տեսակների առաջացման հետ։ Կառուցվածքի հոմոպլաստիկ առանձնահատկությունները նույնն են, որոնց ես խմբավորել եմ, թեպետ և շատ անկատար ձևով, անալոգ փոփոխությունների կամ նմանության անվան տակ։ նրանց առաջացումը կարելի է վերագրել մասամբ տարբեր օրգանիզմների, մասամբ միևնույն օրգանիզմի տարբեր մասերի անալոգ փոփոխություններին, մասամբ էլ նրանք բացատրվում են նման փոփոխություններով, որոնք հարմարված են նույն ընդհանուր նպատակին կամ ֆունկցիային, որի մասին բերված են բազմաթիվ օրինակներ։
Բնախույզները հաճախ ասում են, որ գանգը կազմված է ձևափոխված ողերից, խաչափառի ծնոտները ներկայացնում են ձևափոխված վերջավորություններ, ծաղիկների առէջքները և վարսանդները ձևափոխված տերևներ են, բայց ըստ պրոֆ. Հյոքսլիի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 27] դիտողության, շատ դեպքերում ավելի ճիշտ կլիներ թե՛ գանգի, թե՛ ողերի, թե՛ ծնոտների և թե՛ վերջավորությունների մասին ասել, որ նրանք առաջացնել են ոչ թե մեկը մյուսի փոխարկվելու միջոցով՝ նրանց արդի վիճակի վերածվելով, այլ առաջացել են ավելի պարզ ընդհանուր սաղմից։ Սակայն բնախույզների մեծամասնությունը այդ արտահայտությունը գործ է ածում միայն փոխաբերական իմաստով. նրանք հեռու են այն մտքից, որ զարգացման երկարատև ընթացքում որևէ սկզբնական օրգան, մի դեպքում ողերը, մյուս դեպքում վերջավորությունները, իսկապես դարձել են գանգ կամ ծնոտներ։ Բայց այդպիսի պրոցեսի հետ նմանությունն այնքան մեծ է, որ բնախույզները չեն կարողանում իրենց զսպել այդ արտահայտությունից, որի ուղղակի նշանակությունը հենց այդպիսին է։ Իսկ այստեղ արտահայտված տեսակետների համաձայն այդպիսի արտահայտությունները պետք է գործածել բառացի իմաստով. այն ժամանակ, օրինակ, այն ուշագրավ դեպքը խաչափառի ծնոտների նկատմամբ, որոնք ունեն հավանորեն ժառանգականության միջոցով պահպանված բազմաթիվ առանձնահատկություններ, եթե այդ օրգանները իրոք զարգացել են իսկական, թեկուզ չափազանց պարզ վերջավորություններից, մասամբ գտնում է իր բացատրությունը։
ԶԱՐԳԱՑՈՒՄ ԵՎ ՍԱՂՄՆԱԲԱՆՈՒԹՅՈՒՆ
Ամբողջ բնապատմության մեջ սա ամենակարևոր բաժիններից մեկն է։ Միջատների կերպարանափոխությունները, որ այնքան լավ հայտնի են մեզանից ամեն մեկին, սովորաբար կարծես հանկարծակի են կատարվում՝ սակավաթիվ ստադիաներ անցնելու միջոցով, իրականության մեջ այդ ձևափոխությունները բազմաթիվ են ու հաջորդական, թեև թաքնված են։ Միօրյաներից (Chlöeon) մի միջատ իր զարգացման ընթացքում, ինչպես ցույց է տվել սըր Ջ. Լեբբոկը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 60] մաշկափոխվում է մոտավորապես քսան անգամ և յուրաքանչյուր անգամ որոշ փոփոխություններ է կրում. այս դեպքում մենք տեսնում ենք, որ կերպարանափոխությունը տեղի է ունենում պարզ ձևով և աստիճանաբար։ Շատ միջատներ և առանձնապես որոշ խեցեմորթներ օրինակ են ծառայում, թե ինչպիսի զարմանալի փոփոխություններ են տեղի ունենում կաոուցվածքի մեջ զարգացման ժամանակ։ Բայց այդ փոփոխություններն իրենց գագաթնակետին են հասնում մի քանի ստորակարգ կենդանիների, այսպես կոչված, հերթափոխվող բազմացման մեջ։ Օրինակ, այն զարմանալի փաստը, որ նրբակազմ ճյուղավորվող հիդրոիդը, որը ծածկված է պոլիպներով ու կպած է ստորջրյա ծովային ժայռին, կարող է բողբոջներով և հետո բաժանվելով առաջացնել բազմաթիվ լողացող խոշոր մեդուզաներ, վերջիններս արտադրում են ձվեր, որոնցից զարգանում են լողացող թրթուրներ, այնուհետև թրթուրներն ամրանում են ժայռին, զարգանում են, դառնում ճյուղավորվող հիդրոիդ և այսպես անվերջ։ Այն կարծիքը, թե հերթափոխ բազմացման և սովորական կերպարանափոխության պրոցեսները էապես նման են միմյանց, իր ծանրակշիռ ապացույցը գտավ Վագների[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 85] արած հայտնագործության մեջ, որ մի ճանճի, այն է՝ Cecidomyia-ի թրթուրները անսեռ ճանապարհով առաջացնում են ուրիշ թրթուրներ, որոնք էլ վերջապես դառնում են միանգամայն զարգացած արուներ և էգեր. վերջիններս արդեն բազմանում են սովորական եղանակով, այսինքն՝ ձվերով։
Հիշատակության արժանի է այն, որ երբ Վագների[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 85] ուշագրավ հայտնագործությունը առաջին անգամ հայտնի դարձավ, ինձ հարց տրվեց, թե ինչպե՞ս կարելի է բացատրել, որ այդ ճանճի թրթուրները ձեռք են բերել անսեռ բազմացման ընդունակություն։ Քանի դեռ այդ դեպքը միակն էր, ոչ մի պատասխան տալ չէր կարելի։ Բայց հետո Գրիմմը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 258] ցույց տվեց, որ մի ուրիշ երկթևանի՝ Chironomus-ը բազմանում է գրեթե նույն ձևով, և կարծիք հայտնեց, որ այդ դեպքերը երկթև միջատների կարգում բավական հաճախակի են։ Chironomus-ի մոտ նշված ընդունակությամբ օժտված են հարսնյակները, այլ ոչ թե թրթուրները և Գրիմմը նկատում է, որ այդ դեպքը որոշ չափով «միացնում է ցեցիդոմիաների բազմացումը Coccidae-ի կուսածնության հետ». «կուսածնություն» տերմինը արտահայտում է այն, որ Coccidae-ի չափահաս էգերը կարող են առանց արուների մասնակցության արտադրել զարգանալու ընդունակ ձվեր։ Ներկայումս մեզ հայտնի է, որ զանազան դասերի կենդանիներ ընդունակություն ունեն բազմանալու անսովոր վաղ հասակում, և բավական է միայն ընդունել, որ կուսածին բազմացումը կարող է անցնել ավելի ու ավելի վաղ հասակի, Chironomus-ը լինում է միջակա մի ստադիա, այսինքն հարսնյակի ստադիան,— և մենք թերևս կգտնենք Cecidomyia-ի ուշագրավ երևույթի բացատրությունը։
Ինչպես արդեն նշվել է, որ միևնույն անհատի զանազան մասերը, որոնք միանգամայն նման են իրար վաղ սաղմնային շրջանում, չափահաս կենդանու մոտ դառնում են բոլորովին տարբեր և նախանշվում բոլորովին տարբեր նպատակների համար։ Ճիշտ այդպես էլ նշվել է, որ միևնույն դասին պատկանող ամենատարբեր տեսակների սաղմերն առհասարակ շատ նման են իրար, բայց հասնելով լրիվ զարգացման՝ շատ խիստ զանազանվում են միմյանցից։ Որպես վերջինիս ապացույց ամենալավը, որ կարելի է բերել, դա ֆոն-Բերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 25] խոսքերն են, որ «կաթնասունների թռչունների, մողեսների, օձերի, հավանորեն նաև կրիաների սաղմերը վերին աստիճանի նման են միմյանց իրենց սկզբնական ստադիաներում ինչպես ամբողջությամբ, այնպես էլ առանձին մասերի զարգացման եղանակով․ այդ նմանությունը իսկապես այնքան մեծ է, որ հաճախ մենք կարող ենք սաղմերը զանազանել միայն նրանց մեծությամբ։ Ինձ մոտ սպիրտի մեջ պահվում են երկու փոքրիկ սաղմեր, որոնց անունները ես մոռացել էի նշանակել, և այժմ ես բոլորովին ի վիճակի չեմ ասելու, թե նրանք ինչ խմբի են պատկանում։ Գուցե դրանք մողեսներ են, գուցե փոքրիկ թռչուններ, կամ գուցե թե շատ փոքրիկ կաթնասուններ, այնքան մեծ է այդ կենդանիների գլխի և իրանի կառուցվածքի նմանությունը։ Ասենք՝ այդ սաղմերը վերջավորություններ դեռ չունեն։ Բայց եթե վերջավորություններ նույնիսկ լինեին, գտնվելով զարգացման ամենավաղ ստադիաներում, ապա այն ժամանակ էլ մենք բան չէինք իմանա նրանցից, որովհետև մողեսների թաթերը և կաթնասունների վերջավորությունները, թռչունների թևերն ու ոտները, ինչպես նաև մարդու ձեռքերն ու ոտները զարգանում են միևնույն հիմնական ձևից»։ Խեցեմորթների մեծամասնության թրթուրները զարգացման համապատասխան ստադիաներում շատ նման են իրար, ինչքան էլ տարբեր լինեին չափահաս կենդանիները, և այդպես է ուրիշ շատ կենդանիների մոտ։ Սաղմնային նմանության օրենքի հետքերը երբեմն պահպանվում են մինչև համեմատաբար ուշ հասակը. այսպես, միևնույն և մերձավոր սեռերին պատկանող թռչունները հաճախ իրար նմանվում են մատղաշ հասակի փետրավորությամբ, որը մենք տեսնում ենք, օրինակ, մատղաշ կեռնեխների բծավոր փետրավորման մեջ։ Կատուների ընտանիքում տեսակների մեծամասնությունը չափահաս հասակում բծավոր են կամ զոլավոր, և գույնի այդ առանձնահատկությունը լավ կարելի է զանազանել կորյունների և պումայի ձագերի վրա։ Երբեմն, թեև շատ հազվադեպ, նման մի բան նկատվում է նաև բույսերի վրա. ֆիլլոդիաներ ունեցող Ulex փշոտ դրոկի և ակացիայի առաջին տերևները փետրաձև են կամ բաժանատերև՝ նման թիթեռնածաղկավորների սովորական տերևներին։
Կազմվածքի բոլոր առանձնահատկությունները, որոնցով միևնույն դասին պատկանող խիստ տարբեր կենդանիների սաղմերը նմանվում են իրար, հաճախ անմիջական առնչություն չունեն գոյության պայմանների հետ։ Մենք չենք կարող, օրինակ, կարծել, որ թե մոր արգանդում զարգացող կաթնասունի, թե թռչունի բնում թխսված ձվի մեջ և թե ջրի տակ գորտի ձուղպի մեջ խռիկային ճեղքերի շրջանում զարկերակների հանգույցների գոյացումը կապված է միանման պայմանների հետ։ Մենք այդպիսի առնչության հավատալու ավելի առիթ չունենք, քան հավատալու այն բանին, որ մարդու ձեռքում, չղջիկի թևում և դելֆինի լողաթևում եղած ոսկրների նմանությունը կապված է կյանքի միանման պայմանների հետ։ Ոչ ոք չի կարծում, որ կորյունի զոլավորությունը կամ սև կեռնեխի ձագուկի բծավորությունը գոնե որևէ կերպ օգտակար են կենդանուն։
Բայց բոլորովին այլ բան է, եթե կենդանին իր զարգացման շրջանի որևէ մասի ընթացքում գրոծունյա է լինում և հարկ եղած չափով հոգում է իր մասին։ Գործունեության շրջանը կարող է վրա հասնել վաղ կամ ուշ, բայց երբ էլ նա սկսվելու լինի, թրթուրների հարմարվածությունն իրենց կենսապայմաններին նույնքան կատարյալ են ու հիացմունքի արժանի, որքան չափահաս կենդանու մոտ։ Թե ինչ աստիճանի է հասնում այդ, պարզ կերպով ցույց է տվել վերջերս սըր Ջ. Լեբբոկն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 60] իր դիտողություններում, թե ինչպես կենսապայմանների հետ կապված՝ շատ տարբեր կարգերի մի քանի միջատների թրթուրները մերձավոր նմանություն ունեն միմյանց հետ և միևնույն կարգին պատկանող այլ միջատների թրթուրները իրար նման չեն։ Այդպիսի հարմարեցումների ազդեցության տակ մերձավոր կենդանիների թրթուրների նմանությունը երբեմն խիստ քողարկված է շինում․ հատկապես դա լինում է այն դեպքերում, երբ աշխատանքի բաժանումը տեղի է ունենում ըստ զարգացման տարբեր աստիճանների, ինչպես, օրինակ, եթե միևնույն թրթուրը մի որոշ ստադիայում զբաղվում է կեր որոնելով, իսկ մյուս ստադիայում՝ ամրանալու համար տեղ որոնելով։ Կարելի է նույնիսկ օրինակներ ցույց տալ, որ մերձավոր տեսակների կամ տեսակների խմբերի թրթուրները ավելի շատ են զանազանվում իրարից, քան չափահաս կենդանիները։ Բայց շատ դեպքում թրթուրները, թեև գործունյա են, ավելի կամ պակաս չափով ենթարկվում են ընդհանուր սաղմնային նմանության օրենքին։ Փնջոտնյա խեցեմորթները դրա շատ լավ օրինակ են ներկայացնում, նույնիսկ հռչակավոր Կյուվիեն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 133] չի կասկածել, որ այդ կենդանիները պատկանում են խեցեմորթներին, բայց թրթուրների վրա մի հայացք գցելը բավական է, որպեսզի դրանում ոչ մի կասկած չմնա։ Փնջոտնյաների նույնիսկ երկու գլխավոր ստորաբաժանումները՝ ցողունավորներն ու նստածները իրենց արտաքին տեսքով ինչքան էլ խիստ են տարբերվում միմյանցից, բայց և այնպես ունեն թրթուրներ, որոնք իրենց բոլոր ստադիաներում հազիվ են տարբերվում իրարից։
Սաղմի զարգացման ժամանակ նրա կազմվածքն ավելի կատարյալ է դառնում․ ես այդ արտահայտությունը գործ եմ ածում, թեև գիտեմ, որ հազիվ թե հնարավոր է պարզորոշ կերպով որոշել, թե ինչ է հասկացվում բարձր կամ ավելի ցածր կազմվածք ասելով։ Այնուամենայնիվ, ամենայն հավանականությամբ, ոչ ոք չի վիճարկի, որ թիթեռը իր կազմվածքով ավելի բարձր է թրթուրից։ Սակայն որոշ դեպքերում, ինչպես, օրինակ, մի քանի մակաբույծ խեցեմորթների մոտ էլ չափահաս կենդանուն կարելի է դիտել որպես թրթուրից ավելի ցած կանգնած։ Մի անգամ էլ վերադառնանք փնջոտնյաներին։ Նրանց թրթուրներն առաջին ստադիայում օժտված են շարժողության երեք զույգ օրգաններով, ունեն մի պարզ աչք և կնճիթանման բերան, որի օգնությամբ նրանք առատորեն սնվում են, որովհետև նրանք զգալիորեն մեծանում են։ Երկրորդ ստադիայում, որը համապատասխանում է թիթեռների հարսնյակին, նրանք ունեն վեց զույգ հիանալի կառուցված լողաոտեր, մի զույգ սքանչելի բարդ աչքեր և չափազանց բարդ շոշափուկներ․ բայց բերանը փակ է, անկատար և պիտանի չէ սնունդը կլանելու համար․ այդ ստադիայում նրանց խնդիրն է իրենց լավ զարգացած զգայարանների միջոցով որոնել և լողալու լավ միջոցների շնորհիվ հասնել մի հարմար տեղ ամրանալու համար, որտեղ էլ նրանք ավարտում են իրենց կերպարանափոխությունները։ Երբ կերպարանափոխություններն ավարտված են, նրանք ամրանում են մինչև իրենց կյանքի վերջը, այդ ժամանակ նրանց վերջավորությունները դառնում են բռնելու օրգաններ․ բերանը նորից դառնում է լավ կառուցված, բայց շոշափուկներ չունեն, իսկ երկու աչքերը ձևափոխվել, դարձել են մի փոքրիկ պարզ ակնային բիծ։ Այս վերջին ստադիայում փնջոտանին կարելի է համարել և ավելի բարձր, և ավելի ցածր կազմակերպված, քան նա եղել է թրթուրային ստադիայում։ Բայց մի քանի սեռերի թրթուրները զարգանում, դառնում են սովորական կառուցվածքով հերմաֆրոդիտ անհատներ, կամ այնպիսիներ, որոնց ես անվանել եմ լրացուցիչ արուներ․ վերջիններիս զարգացումը անկասկած ետ է գնացել, որովհետև այդպիսի արուն իրենից ներկայացնում է պարզապես մի պարկ, որը կարճատև գոյություն ունի և զուրկ է բերանից, ստամոքսից և բոլոր այլ կարևոր օրգաններից, բացի վերարտադրության օրգաններից։
Մենք այնքան սովորել ենք սաղմի և չափահաս կենդանու միջև եղած տարբերությանը, որ աշխատում ենք այդ տարբերությունը կախման մեջ դնել աճումից։ Բայց այդ դեպքում հիմք չէր լինի նրա համար, օրինակ, որպեսզի չղջիկի թևը կամ դելֆինի լողաթևը իրենց բոլոր մասերով հիմնադրված չլինեին՝ պահպանելով որոշ հարաբերական չափեր, հենց որ որևէ մասը նկատելի կդառնա։ Այդ լինում է որոշ խմբերում ամբողջությամբ և այլ խմբերում տարբեր անդամների մոտ, ըստ որում սաղմն իր ոչ մի ստադիայում զգալիորեն չի տարբերվում չափահաս կենդանուց. այսպես, թանաքաձկան մասին Օուենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] նկատել է, որ «այստեղ կերպարանափոխություն չկա. գլխոտանի կակղամորթի բնույթն արտահայտված է դեռ շատ ավելի վաղ, քան սաղմի բոլոր մասերը կարողանում են ձևավորվել»։ Ցամաքային կակղամորթները և քաղցրահամ ջրերի խեցեմորթները ծնվում են՝ ունենալով իրենց բնորոշ ձևը, մինչդեռ նույն երկու մեծ խմբերի ծովային անդամներն իրենց զարգացման ժամանակ ենթարկվում են բավական կարևոր և հաճախ նույնիսկ մեծ փոփոխությունների։ Սարդերն, ընդհակառակը, հազիվ թե որևէ կերպարանափոխության են ենթարկվում։ Միջատների մեծամասնության թրթուրները անցնում են որդանման ստադիայով, միևնույնն է թե նրանք գործունյա են և հարմարված զանազան պայմանների կամ անգործունյա, երբեմն գտնվելով ուղղակի սնունդ ծառայող միջավայրի մեջ, երբեմն կերակրվելով ծնողների կողմից․ բայց մի քանի սակավաթիվ դեպքերում, օրինակ խոտային լվիճների մոտ, եթե նայենք այդ միջատների զարգացման սքանչելի նկարները, որ արել է պրոֆ. Հյոքսլին,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 27] մենք հազիվ նշմարենք որդանման ստադիայի որևէ հետք։
Երբեմն դուրս են ընկնում զարգացման միայն վաղ ստադիաները։ Այսպես. Ֆրից Մյուլլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] արել է մի ուշագրավ հայտնագործություն, որ Penaeus-ին մերձավոր մի քանի խեցեմորթներ սկզբում երևան են գալիս նաուպլիուսի ձևով, հետո անցնում են Zoea-ի երկու կամ ավելի ստադիաները, այնուհետև միզիս ստադիան և վերջապես ստանում են չափահասի կառուցվածք. ամբողջ Malacostraca ընդարձակ կարգի մեջ, որին պատկանում են այդ խեցեմորթները, չկա խմբի մի այլ անդամ, որը երևան գար Nauplius-ի ստադիայում, թեև շատերը երևան են գալիս Zoea-ի ձևով. այնուամենայնիվ Մյուլլերը հիմքեր է բերում իր կարծիքի համար, որ եթե այդ բոլոր դեպքերում ճնշված զարգացում չլիներ, ապա այդ բոլոր խեցեմորթները կծնվեին նաուպլիուսների ձևով։
Իսկ ինչպե՞ս մենք կարող ենք բացատրել սաղմնաբանության բնագավառից վերցված այս զանազան փաստերը, այն է՝ շատ տարածված, բայց ոչ ընդհանրական տարբերությունը սաղմի և չափահաս կենդանու կառուցվածքի միջև. միևնույն սաղմնային անհատի զարգացման վաղ ստադիաներում զանազան մասերի նմանությունը, որոնք վերջ ի վերջո խիստ, տարբերվում են ու ծառայում զանազան նպատակների համար. միևնույն դասի ամենատարբեր տեսակների սաղմերի և թրթուրների միջև եղած ընդհանուր, թեև տատանվող նմանությունը. այն, որ ձվի կամ արգանդի մեջ գտնվող սաղմը հաճախ ունենում է կառուցվածքի այնպիսի առանձնահատկություններ, որոնք նրան օգուտ չեն բերում կյանքի ոչ այդ, ոչ էլ հետագա ստադիաներում, մինչդեռ իրենց սեփական ուժերին թողնված թրթուրները հիանալի կերպով հարմարված են շրջապատող պայմաններին, և վերջապես այն օրինակները, որ որոշ թրթուրներ իրենց կազմվածքով ավելի բարձր են կանգնած, քան այն չափահաս կենդանին, որպիսին նրանք պիտի լինեն զարգանալուց հետո։ Ես կարծում եմ, որ բոլոր այս տվյալները կարելի է բացատրել հետևյալ կերպով։
Սովորաբար ընդունում են, կարող է պատահել այն հիման վրա, որ այլանդակությունները սաղմի վրա արտահայտվում են շատ վաղ շրջանում, որ թույլ փոփոխությունները կամ անհատական տարբերությունները անհրաժեշտորեն պետք է ճիշտ նույնպես էլ հայտնվեն շատ վաղ հասակում։ Մենք այդ ուղղությամբ քիչ տվյալներ ունենք, բայց այն, որ ունենք, պարզորոշ կերպով մեզ բերում են այլ եզրակացության․ այսպես, ուշադրության արժանի է այն, որ եղջերավոր անասուններ, ձիեր և ընտանեցված զանազան կենդանիներ բուծող անձինք չեն կարող նախապես որոշակի ասել, թե ինչ արժանիքներ կամ թերություններ ունի նորածին կենդանին, մինչև որ նրա ծնվելուց հետո մի որոշ ժամանակ չանցնի։ Միանգամայն նույնը մենք տեսնում ենք մեր զավակների վրա, որովհետև մենք չենք կարող ասել, թե արդյոք երեխան բարձրահասակ կլինի, թե ցածրահասակ, և իսկությամբ ինչպես կլինեն նրա դիմագծերը։ Հարցը նրանում չէ, թե ինչ շրջանում են դրսևորվում այդ փոփոխության հետևանքները։ Մի որոշ հանգամանք կարող է ներգործել և, ես կարծում եմ, հաճախ ներգործում է ծնողների մեկի կամ երկուսի վրա էլ մինչև ծննդաբերությունը։ Պետք է հիշատակել, որ մատղաշ կենդանու համար քանի դեռ նա մնում է ձվի մեջ կամ մոր արգանդում և կերակրվում ու պահպանվում է ծնողներից մեկի կողմից, ամենևին կարևոր չէ, թե ինքը իր առանձնահատկությունների մեծ մասը ձեռք կբերի քիչ ավելի վաղ կամ ուշ։ Այսպես, օրինակ, շատ ծռված կտուցով կեր հայթայթող թռչունի համար ամենևին կարևոր չէ, թե արդյոք ձագուկն ունի այդպիսի կտուց, քանի դեռ այդ ձագուկը կերակրվում է ծնողների միջոցով։
Առաջին գլխում ես պարզել եմ, որ փոփոխությունը ծնողի մոտ ինչ հասակում էլ երևան գա, նա ձգտում է հաջորդի մոտ երևալ համապատասխան հասակում։ Որոշ փոփոխություններ կարող են երևան գալ միայն համապատասխան հասակում, օրինակ՝ շերամի թրթուրի, հարսնյակի և չափահաս ստադիայի, կամ եղջերավոր անասունների բոլորովին զարգացած եղջյուրների առանձնահատկությունները։ Բայց այն փոփոխություններն էլ, որոնք, որքան մենք կարող ենք դատել, կարող էին երևալ ավելի վաղ կամ ավելի ուշ, հավասարապես ձգտում են երևալ ծնողի և հաջորդի մոտ համապատասխան հասակում։ Ես հեռու եմ այն մտքից, որ միշտ այդպես է լինում, և կարող էի բերել փոփոխությունների (հասկանալով այդ բառը ամենալայն իմաստով) մի քանի բացառիկ դեպքեր, երբ փոփոխությունները ձագի վրա երևացել են ավելի վաղ հասակում, քան ծնողների վրա։
Այս երկու սկզբունքները, այսինքն՝ որ թույլ փոփոխությունները սովորաբար երևան են գալիս ոչ ամենավաղ հասակում, և ժառանգաբար փոխանցվում են համապատասխան ոչ վաղ հասակում, ինչպես ես կարծում եմ, բացատրում են սաղմնաբանության բնագավառից վերցրած ամենից ավելի աչքի ընկնող փաստերը։ Բայց նախ ծանոթանանք մի քանի անալոգ դեպքերի հետ մեր ընտանի ցեղերի նկատմամբ։ Մի քանի հեղինակներ, որոնք գրել են շների մասին, հաստատում են, որ գամփռը և բուլդոգը, թեև այդքան տարբեր են թվում, իրականության մեջ ներկայացնում են շատ մոտ ցեղեր, որոնք ծագել են միևնույն վայրենի նախածնողից. այդ պատճառով ինձ չափազանց հետաքրքրել է, թե որչափ են տարբերվում նրանց լակոտները միմյանցից․ և ես իմացա շնաբույծներից, որ այդ ցեղերի լակոտները իրարից տարբերվում են նույնչափ, որչափ որ չափահաս շները, որը ճշմարիտ է՝ աչքաչափով դատելով. բայց չափելով ծեր շներին և նրանց վեցօրյա լակոտներին, ես գտել եմ, որ վերջիններս հարաբերական չափերով լրիվ տարբերություն չեն ունենում։ Ճիշտ այդպես էլ ինձ ասել են, որ ծանրաքաշ ձիու և արշավաձիու մտրուկները, այսինքն ընտանեցված կենդանիներից ընտրության միջոցով ստացված երկու ցեղերի մտրուկներն իրարից տարբերվում են նույն չափով, ինչ չափով որ չափահաս անհատները, բայց ես մանրազնին կերպով չափելով արշավաձիու և ծանրաքաշի զամբիկներին և երեք օրական մտրուկներին, գտել եմ, որ այդպիսի տարբերություն բնավ չկա։ Ունենալով իմ առջև վճռական ապացույցներ այն մասին, որ աղավնիների բոլոր ցեղերը ծագում են առել մի վայրի տեսակից, ես զբաղվել եմ նրանց ձագուկների համեմատությամբ ձվից դուրս գալուց 12 ժամ հետո ես մանրազնին կերպով չափել եմ (բայց այստեղ չեմ բերում մանրամասնությունները) կտուցը, բերանի բացվածքը, քթանցքների և կոպերի, մատների և ամբողջ ոտքի երկարությունը թե վայրի նախածնողական տեսակի և թե փքունի, սիրամարգանմանի, թուրմանի, բերբերիականի, փոստատար և այլ ցեղերի աղավնիների վրա։ Այս թռչուններից մի քանիսը չափահաս վիճակում այնքան ուժեղ կերպով են տարբերվում իրարից իրենց կտուցի երկարությամբ ու ձևով, ինչպես նաև այլ առանձնահատկություններով, որ եթե նրանք գտնվեին վայրի վիճակում, ապա նրանց անշուշտ կդասեին տարբեր սեռերի շարքում։ Բայց երբ այդ թռչունների ձագուկները զետեղվեցին մի չարքում, ապա թեև նրանց մեծ մասը կարելի էր ճանաչել, սակայն վերոհիշյալ մասերում եղած հարաբերական տարբերությունը անհամեմատ ավելի քիչ էր, քան այդ նկատելի է չափահաս թռչունների մոտ։ Տարբերության մի քանի բնորոշ գծեր, օրինակ բերանի բացվածքը, ձագուկների մոտ հազիվ կարող են նկատվել։ Բայց այս կանոնից կար մի ուշագրավ բացառություն, այն է՝ կարճակտուց թուրմանի ձագուկները վայրի աղավնու և այլ ցեղերի ձագուկներից գրեթե նույնքան են տարբերվում, որքան տարբերվում են չափահաս թռչունները միմյանցից։
Այս փաստերը իրենց բացատրությունն են գտնում վերևում բերված սկզբունքներում։ Անասնաբույծները, առանձին ցեղեր ստեղծելու համար, զույգընտրում են շներին, ձիերին, աղավնիներին և այլն, երբ կենդանիներն արդեն հասել են լրիվ զարգացման, նրանց համար միևնույն է, թե որոնելի հատկությունները արդյոք ավելի վաղ կամ ավելի ուշ ձեռք կբերվեն, միայն թե նրանք լինեն չափահաս կենդանու մոտ։ Եվ բերված օրինակները, առանձնապես աղավնիների վրա կատարված դիտողություններից վերցրածները, ապացուցում են, որ այն բնորոշ տարբերությունները, որոնք առաջացել են մարդու զույգընտրության միջոցով և իրենցով պայմանավորում են ցեղերի առանձնահատկությունները, երևան են գալիս ոչ վաղ և ժառանգաբար փոխանցվում են համապատասխան ոչ վաղ հասակում։ Բայց տասներկու օրական հասակում բնորոշիչ հատկանիշներ ունեցող կարճակտուց թուրմանի օրինակն ապացուցում է, որ դա ընդհանուր կանոն չէ, որովհետև կամ այդ դեպքում բնորոշ առանձնահատկությունները սովորականից ավելի շուտ են երևացել, կամ եթե այդ չկա, ապա այդ առանձնահատկությունները ժառանգաբար փոխանցվել են ոչ համապատասխան, այլ ավելի վաղ հասակում։
Այժմ փորձենք այս երկու սկզբունքները կիրառել բնական պայմաններում գտնվող տեսակների վերաբերմամբ։ Վերցնենք մի խումբ թռչուններ, որոնք ծագել են մի որոշ հին ձևից և բնական ընտրության ազդեցության տակ փոփոխվել են՝ հարմարվելով տարբեր կենցաղի։ Քանի որ հաջորդական թույլ փոփոխությունները զանազան տեսակների մոտ երևան են եկել ոչ վաղ հասակում և ժառանգաբար փոխանցվել են համապատասխան հասակում, ուստի մատղաշները կարող են շատ քիչ փոփոխվել և պետք է իրար նմանվեն անհամեմատ ավելի շատ, քան չափահասները, հենց այն, ինչ մենք տեսանք աղավնիների ձագուկների վրա։ Այդ տեսակետը մենք կարող ենք տարածել նաև շատ տարբեր օրգանների և ամբողջ դասերի վրա։ Օրինակ, առջևի վերջավորությունները, որոնք հեռավոր նախածնողին ծառայել են որպես ոտքեր, կարող էին երկարատև փոփոխությունների միջոցով հարմարվել որոշ հաջորդների մոտ գործելու որպես ձեռքեր, մյուսների մոտ՝ որպես լողաթևեր, երրորդների մոտ՝ որպես թևեր․ բայց վերոհիշյալ սկզբունքների հիման վրա զանազան ձևերի սաղմերի առջևի վերջավորությունները պետք է շատ չտարբերվեն իրարից, թեև յուրաքանչյուր ձևի մեջ առջևի վերջավորությունը խիստ տարբեր է չափահաս հասակում։ Ինչպիսին էլ որ լինի երկարատև վարժեցման կամ ոչ վարժեցման ազդեցությունը որևէ տեսակի վերջավորությունների կամ այլ մասերի փոփոխության վրա, գերադասորեն կամ բացառապես նա կարող է արտահայտվել միայն բոլորովին չափահաս հասակում, երբ օրգանիզմը կարող է իր ամբողջ ուժերից օգտվել իր գոյությունը պահպանելու համար․ և դրա հետևանքները կարող են փոխանցվել հետնորդներին համապատասխան գրեթե հասուն հասակում։ Այսպիսով, մարմնամասերի հարաճուն վարժեցման կամ ոչ վարժեցման ազդեցության տակ մատղաշ կենդանին կամ բոլորովին չի փոփոխվում կամ միայն թույլ չափով է փոփոխվում։
Մի քանի կենդանիների մոտ հաջորդական փոփոխությունները կարող են սկսվել շատ վաղ շրջանում, կամ համապատասխան ստադիաները կարող են ժառանգաբար փոխանցվել ավելի վաղ հասակում, քան առաջին անգամ երևացել են։ Թե այս և թե այն դեպքում մատղաշ կենդանին կամ սաղմը մոտ նմանություն կունենա մայրական ձևի հետ, ինչպես մենք տեսնում ենք կարճակտուց թուրմանի վրա։ Զարգացման այդպիսի ընթացքը կանոն է ինչպես մի քանի մեծ խմբերի համար նրանց ամբողջ կազմով վերցրած, այնպես էլ այլ խմբերի ստորաբաժանումների համար, օրինակ գլխոտանիների և ցամաքային կակղամորթների, քաղցրահամ ջրերի խեցեմորթների, սարդերի և միջատների մեծ դասի մի քանի ստորաբաժանումների համար։ Ինչ վերաբերում է այն բանի հիմնական պատճառին, որ այդ խմբերում ծնված անհատները կերպարանափոխություններ չեն կրում, ապա այդ մենք կարող ենք բացատրել հետագա երևույթներով, այսինքն նրանով, որ մատղաշ անհատը դեռ իր շատ վաղ հասակում թողնվում է սեփական ուժերին և վարում է միևնույն կենցաղը, ինչ որ ծնողները. այդ դեպքում մատղաշ անհատների գոյության համար անպայման անհրաժեշտ է, որպեսզի նրանք փոփոխվեին այնպես, ինչպես իրենց ծնողները։ Ինչ վերաբերում է այն տարօրինակ փաստին, որ ցամաքային և քաղցրահամ ջրերի շատ կենդանիներ կերպարանափոխության չեն ենթարկվում, մինչդեռ նույն խմբերի ծովային անդամները անցնում են զանազան կերպարանափոխություններով, ապա Ֆրից Մյուլլերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] հայտնել է այն ենթադրությունը, որ կենդանու դանդաղ փոփոխվելու և ցամաքի վրա կամ ծովի փոխարեն քաղցրահամ ջրում ապրելուն հարմարվելու պրոցեսը կպարզանար նրանով, եթե կենդանիները չանցնեին թրթուրային ստադիայով, որովհետև միանգամայն անհավատալի կլիներ, որ թե թրթուրային և թե չափահաս ստադիաների համար պիտանի տեղերը, նրանց կյանքի այնքան նոր և փոփոխված կենսապայմաններում, սովորաբար գրավված չլինեին, կամ վատ գրավված լինեին այլ օրգանիզմների կողմից։ Այդ դեպքում աստիճանաբար ավելի ու ավելի վաղ հասակում չափահասի կառուցվածքի ձեռք բերումը օժանդակություն կգտներ բնական ընտրության կողմից, և վերջապես նախկին կերպարանափոխության բոլոր հետքերը կկորչեին։
Մյուս կողմից, եթե բնական ընտրությունը նպաստում է մատղաշ կենդանուն, որպեսզի վարի ծնողական ձևի կենցաղից քիլ տարբեր կենցաղ, և հետևաբար քիչ տարբերվի իր կառուցվածքով, կամ եթե նպաստում է այն բանին, որպեսզի ծնողներից արդեն տարբերվող թրթուրը ավելի ևս փոփոխվի, այդ դեպքում, համապատասխան հասակում առանձնահատկությունների ժառանգական փոխանցման սկզբունքի հիման վրա, մատղաշ անհատները, նամ թրթուրները բնական ընտրության ազդեցության տակ կարող են ավելի ու ավելի տարբերվել ծնողներից ուզած չափով։ Թրթուրների միջև եղած տարբերությունները կարող են նաև որոշ հարաբերակցության մեջ լինել իրենց զարգացման հաջորդական ստադիաների հետ, և առաջին ստադիայում գտնվող թրթուրը կարող է շատ խիստ կերպով տարբերվել երկրորդ ստադիայի թրթուրից, որը իրոք նկատվում է շատ կենդանիների մոտ։ Չափահաս կենդանիները կարող են պիտանի դառնալ նստակյաց կամ մի այնպիսի կյանքի համար, որի ժամանակ շարժողության օրգանները, զգայարանները և այլն դառնում են անօգուտ, այդ դեպքում կերպարանափոխությունը պետք է ընթանա ռեգրեսիվ ճանապարհով։
Արված դիտողություններից կարելի է եզրակացնել, թե ինչպես մատղաշ անհատների կազմվածքի մեջ կատարված փոփոխությունների, դրա հետ միասին կենցաղի փոփոխության և համապատասխան հասակներում զանազան առանձնահատկությունների ժառանգման միջոցով կենդանիները կարող են անցնել զարգացման այնպիսի ստադիաներ, որոնք բոլորովին տարբեր են նրանց չափահաս նախածնողների սկզբնական վիճակից։ Մեր ականավոր հեղինակների մեծամասնությունը համոզված է այն բանում, որ միջատների թրթուրների և հարսնյակների զանազան ստադիաները ձեռք են բերվել հարմարեցման միջոցով, այլ ոչ թե որևէ հին ձևից ժառանգաբար են ստացվել։ Sitaris-ի, մի բզեզի, որն անցնում է զարգացման մի քանի անսովոր ստադիաներ, հետաքրքրական դեպքը լավ բացատրություն է ծառայում այն բանի համար, թե այդ ինչպես կարող էր տեղի ունենալ։ Առաջին թրթուրային ձևը, որ նկարագրել է Ֆաբրը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 78] ներկայացնում է փոքրիկ շարժուն մի միջատ, օժտված վեց ոտներով, երկու երկար բողկուկներով և չորս աչքերով։ Այդ թրթուրները դուրս են գալիս մեղուների բներում, և երբ բոռերը գարնանը էգերից ավելի շուտ են դուրս գալիս բներից, թրթուրները բարձրանում են նրանց վրա, իսկ հետո նրանց և էգերի զուգավորման ժամանակ անցնում են էգերի վրա։ Հենց որ էգերը ձվեր են դնում մեղրահացի բջիջներում լցված մեղրի մակերեսի վրա, Sitaris-ի թրթուրները հարձակվում են ձվերի վրա և լափում նրանց։ Դրանից հետո նրանք լրիվ փոփոխություն են կրում. նրանց աչքերն անհետանում են, ոտները և բողկուկները դառնում են ռուդիմենտար, և այժմ սկսում են սնվել մեղրով, դրանով հանդերձ ավելի նմանվելով միջատների սովորական թրթուրներին. վերջապես, նրանք ենթարկվում են վերջնական կերպարանափոխության և վերջ ի վերջո դառնում չափահաս բզեզներ։ Պարզ է, որ եթե Sitaris-ի նման կերպարանափոխության ենթարկվող մի միջատ դառնար միջատների մի ամբողջ նոր խմբի նախածնող, ապա նոր խմբի զարգացման ընթացքը խիստ տարբեր կլիներ գոյություն ունեցող միջատների զարգացումից, և առաջին թրթուրային ստադիան անշուշտ չէր ներկայացնի որևէ հին չափահաս ձևի վաղ ստադիան։
Մյուս կողմից, վերին աստիճանի հավանական է, որ շատ կենդանիների սաղմնային կամ թրթուրային ստադիաները շատ թե քիչ պարզ կերպով մեզ ցույց են տալիս, թե ինչպիսի կառուցվածք է ունեցել ամբողջ խմբի նախածնողն իր չափահաս վիճակում։ Խեցեմորթների ընդարձակ դասի մեջ միմյանցից խիստ տարբերվող ձևերը, այն է՝ ծծող մակաբույծները, փնջոտնյաները, էնտոմոստրակաները և նույնիսկ մալակոստրակաները, ծնվում են թրթուրի ձևով, որը հայտնի է նաուպլիուս անունով․ և որովհետև այդ թրթուրները ապրում են և իրենց կերը հայթայթում են բաց ծովում, այլ ոչ թե հարմարված են որևէ առանձին կենցաղի համար, ինչպես նաև այլ նկատառումով, որ բերում է Ֆրից Մյուլլերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 61] հավանական է, որ մի ինչ որ շատ հեռավոր ժամանակաշրջանում գոյություն է ունեցել Nauplius-ին նման մի կենդանի, որը հաջորդականորեն փոփոխվելով, հետնորդների մի քանի տարամիտվող գծեր առաջացնելու միջոցով առաջ է բերել խեցեմորթների վերոհիշյալ սիստեմատիկական խմբերը։ Ճիշտ այդպես էլ հիմնվելով այն բոլորի վրա, ինչ որ մեզ հայտնի է կաթնասունների, թռչունների, սողունների և ձկների սաղմերի մասին, կարելի է հավանական համարել, որ այդ կենդանիները ներկայացնում են մի այնպիսի ընդհանուր հին նախածնողի փոփոխված հաջորդներ, որը չափահաս վիճակում ունեցել է խռիկներ, լողափամփուշտ, երկու զույգ լողաթևանման վերջավորություններ, երկար պոչ և հարմարված է եղել ջրային կենցաղի։
Քանի որ բոլոր կազմակերպված էակները, թե մահաջնջվածները և թե այժմ ապրողները, հետևաբար՝ երբևիցե գոյություն ունեցածները, կարող են բաժանվել սակավաթիվ դասերի և ամեն մի դասի սահմաններում, ըստ մեր տեսության, շաղկապվում են միմյանց հետ աստիճանական անցումներով, ուստի ամենալավ և միակ հնարավոր դասակարգությունը, եթե մեր հավաքածուները գոնե մոտավորապես լրիվ լինեին, կլիներ ծննդաբանականը․ ընդհանուր ծագումը ներկայացնում է այն թաքնված կապերը, որոնք բնախույզն աշխատում է հայտնաբերել, նշանակելով դրանք «բնական սիստեմ» տերմինով։ Այդ տեսակետից մենք կարող ենք հասկանալ, թե ինչու բնախույզների մեծամասնության աչքում սաղմի կառուցվածքը դասակարգության համար նույնիսկ ավելի մեծ նշանակություն ունի, քան չափահաս կենդանու կառուցվածքը։ Կենդանիների երկու կամ ավելի խմբեր ինչքան էլ չափահաս վիճակում խիստ կերպով տարբերվեն միմյանցից իրենց կազմվածքով ու կենցաղով, եթե զարգացման ընթացքում անցնում են մերձավոր ստադիաներ, մենք կարող ենք վստահ լինել, որ նրանք ծագում են առել մի ընդհանուր նախածնողական ձևից և հետևաբար իրար հետ մերձավոր ազգակցության մեջ են։ Այսպիսով սաղմնային կառուցվածքի ընդհանրությունը կապված է ծագման ընդհանրության հետ. բայց սաղմնային զարգացման ոչ միանմանությունը չի ցույց տալիս, որ նրանք տարբեր ծագում ունեն, որովհետև երկու խմբերից մեկի զարգացման մի քանի ստադիաները կամ կարող են ճնշված լինել, կամ կյանքի նոր պայմաններին հարմարվելու հետևանքով այնքան փոփոխված, որ դառնում են անճանաչելի։ Նույնիսկ այնպիսի խմբերում, որոնց մեջ չափահաս ձևերը չափազանց տարբեր են, ծագման ընդհանուր լինելը դրսևորվում է թրթուրային կառուցվածքում. օրինակ, մենք տեսանք, որ փնջոտնյա խեցեմորթները, չնայած կակղամորթների հետ իրենց ունեցած արտաքին նմանությանը, իսկույն ճանաչվում են իրենց թրթուրների ձևով ու կառուցվածքով, որ խեցեմորթների մեծ դասին են պատկանում։ Քանի որ սաղմը հաճախ մեզ ցույց է տալիս խմբի նվազ փոփոխված հեռավոր նախածնողի շատ թե քիչ լրիվ կառուցվածքը, ուստի մենք կարող ենք հասկանալ, թե ինչու հին մահաջնջված ձևերն իրենց չափահաս վիճակում այդքան հաճախ նմանվում են միևնույն դասին պատկանող այժմ ապրող տեսակների սաղմերին։ Ագասսիսը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 97] կարծում է, որ դա բնության ընդհանուր օրենքն է, և մենք կարող ենք հուսալ, որ երբևիցե այդ օրենքը կապացուցվի։ Սակայն այդ օրենքի ճշտությունը կարելի է ապացուցել միայն այն դեպքում, երբ նախածնողական խմբի հին ձևը իսպառ չի անհետացել կամ շատ վաղ շրջանի ընթացքում հաջորդական փոփոխությունների ազդեցության տակ, կամ շնորհիվ այդպիսի փոփոխությունների ժառանգական փոխանցման ավելի վաղ հասակում, քան նրանք սկզբում երևան են եկել։ Պետք է նաև նկատի ունենալ, որ նշված օրենքը իսկապես կարող է գոյություն ունենալ, բայց քանի որ երկրաբանական տարեգրությունը դեպի ետ հեռավոր ժամանակների վրա չի տարածվում, ուստի երկար ժամանակ և նույնիսկ ընդմիշտ այդ օրենքը կմնա չապացուցված։ Այդ օրենքը առանձնապես խստորեն կիրառելի չէ այն դեպքերում, երբ հին ձևն իր թրթուրային վիճակում հարմարվել է որևէ հատուկ կենսապայմանների և այդ թրթուրային վիճակը փոխանցել է ամբողջ խմբի հետնորդներին, որովհետև այդպիսի թրթուրները նմանություն չունեն ավելի հեռավոր ձևի հետ նրա չափահաս վիճակում։
Այսպիսիք են, ինչպես ինձ թվում է, սաղմնաբանության ղեկավարող փաստերը, որոնք ամենևին չկորցնելով իրենց նշանակությունը, բացատրվում են նրանով, որ մի հեռավոր նախածնողի հետնորդների փոփոխությունները երևան են գալիս կյանքի ոչ շատ վաղ շրջանում և ժառանգաբար փոխանցվում են համապատասխան հասակում։ Սաղմնաբանությունը հետաքրքրության տեսակետից մեծ չափով կշահի, եթե մենք սաղմի մեջ տեսնենք մի մեծ խմբի բոլոր անդամների ընդհանուր նախածնողի ավելի կամ պակաս աղոտ պատկերը նրա չափահաս կամ թրթուրային վիճակում։
ԹԵՐԱՃ, ԱՏՐՈՖԻԱՅԻ ԵՆԹԱՐԿՎԱԾ ԵՎ ԹԵՐԱԶԱՐԳԱՑԱԾ ՕՐԳԱՆՆԵՐ
Այն օրգանները կամ օրգանիզմի մասերը, որոնք գտնվում են իրենց անօգտակարությունը ամբողջապես դրսևորող այս զարմանալի վիճակում, շատ սովորական են և ամենուրեք տարածված են բնության մեջ։ Անհնարին է անվանել բարձրակարգ կենդանիներից որևէ մեկին, որի այս կամ այն մասը թերաճ վիճակում չլինի։ Օրինակ, կաթնասունների արուներն ունեն թերաճ պտուկներ, օձերի թոքերի մի բլթակը թերաճ է, թռչունների «թևիկը» կարելի է համոզված կերպով դիտել որպես թերաճ մատ, իսկ մի քանի տեսակների թևը այն աստիճան թերաճ է, որ թռչելու համար չի կարող ծառայել։ Ի՞նչը կարող է ավելի հետաքրքրական լինել, քան ատամների ներկայությունը կետերի սաղմերի մոտ, որոնք չափահաս վիճակում ատամներ բոլորովին չունեն, կամ այն, որ դեռ չծնված հորթը վերին ծնոտի վրա ունենում է երբեք դուրս չեկող կտրիչ ատամներ։
Թերաճ օրգանները պարզորոշ ու տարբեր ձևերով երևան են հանում իրենց ծագումն ու նշանակությունը։ Այսպես, կան մերձավոր տեսակների կամ նույնիսկ միևնույն տեսակին պատկանող բզեզներ, որոնցից մի քանիսն ունեն մեծ, միանգամայն զարգացած թևեր, մինչդեռ մյուսների թևերը ներկայացնում են միայն թաղանթների թերաճիկներ, որոնք հաճախ գտնվում են սերտաճած վերնաթևերի տակ. այսպիսի դեպքերում անհնարին է կասկածել, որ թերաճիկները թևեր են։ Թերաճ օրգանները երբեմն պահպանում են իրենց գործունեության ընդունակությունը․ այսպես, պատահում է, որ կաթնասունների արուների պտուկները լավ զարգացած են լինում և կաթ են արտաթորում։ Bos սեռի ներկայացուցիչների կուրծը նորմալ կերպով ունի չորս լավ զարգացած և երկու թերաճած պտուկներ, բայց վերջիններս մեր ընտանի կովերի մոտ երբեմն լինում են լավ զարգացած և կաթ են արտաթորում։ Ինչ վերաբերում է բույսերին, ապա միևնույն տեսակի տարբեր անհատների մոտ պսակաթերթիկները լինում են մերթ թերաճ, մերթ՝ լավ զարգացած։ Մի քանի բաժանասեռ բույսերի մոտ Կյոլռեյտերը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] գտել է, որ երբ խաչաձևվում է այն տեսակը, որի արական ծաղիկներն ունեն վարսանդի թերաճիկ, լավ զարգացած վարսանդ ունեցող հերմաֆրոդիտ տեսակի հետ, այդ դեպքում հիբրիդի մոտ վարսանդը շատ մեծանում է, որը պարզ կերպով ցույց է տալիս, որ թերաճ և նորմալ վարսանդներն իրենց բնույթով էապես նման են։ Կենդանու զանազան մասերը կարող են լավ զարգացած լինել, բայց որոշ իմաստով թերաճ են իրենց անօգտակարության պատճառով։ Այսպես, սովորական սալամանդրայի շերեփուկը, ինչպես նկատում է միստեր Ջ. Հ. Լյուիսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 135] «ունի խռիկներ և ապրում է ջրի մեջ. բայց սև սալամանդրան, որն ապրում է բարձրում, լեռների վրա, ծնում է բոլորովին կազմավորված մատղաշ անհատ։ Այս կենդանին ջրում երբեք չի ապրում։ Սակայն եթե հերձենք հղի էգին, մենք նրա մեջ կգտնենք շատ զարգացած փետրաձև խռիկներ ունեցող շերեփուկներ, և եթե նրանց թողնենք ջրի մեջ, ապա կտեսնենք, որ նրանք լողում են սովորական սալամանդրայի շերեփուկների նման։ Ակներև է, որ ջրի մեջ ապրելու համար հարմարված այդ կազմվածքը ոչ մի առնչություն չունի կենդանու ապագա կյանքի հետ և ամենաչնչին հարմարեցում չի ներկայացնում սաղմնային գոյության համար․ նա հարաբերակցության մեջ է միայն հեռավոր հարմարեցումների հետ, իրմով կրկնելով իր նախածնողի զարգացման մեջ եղած մի որոշակի փուլը»։
Երկու նպատակների համար ծառայող օրգանը կարող է դառնալ թերաճ և նույնիսկ բոլորովին թերազարգացած մի ավելի կարևոր նպատակի համար և միանգամայն պիտանի մնալ մյուսի համար։ Այսպես, օրինակ, բույսերի մոտ վարսանդի դերն այն է, որ փոշեհատիկի խողովակներն անցկացնի դեպի սերմնարանի մեջ գտնվող սերմնաբողբոջը։ Վարսանդը կազմված է սպիից, որը հենվում է սռնակի վրա. բայց մի քանի բարդածաղիկների արական ծաղիկները, որոնք իհարկե չեն կարող բեղմնավորվել, ունեն թերաճ վարսանդ, որի վրա սպի չկա, մինչդեռ սռնակը լավ զարգացած է և սովորականի նման ծածկված է մազմզուկներով, որոնք ծառայում են նրա համար, որպեսզի դուրս նետեն ծաղկափոշին իրեն շրջապատող և իրար հես։ Խողովակի ձևով սերտաճած փոշանոթներից։ Բացի դրանից, օրգանը կարող է թերաճ դառնալ իրեն հատուկ դերի համար և գործածվել մի այլ նպատակի համար․ որոշ ձկների մոտ լողափամփուշտը թերազարգացած է որպես հիդրոստատիկ ապարատ, բայց դարձել է շնչառության նոր սկզբնավորվող մի օրգան կամ թոք։ Կարելի է բերել շատ նաև այլ նման օրինակներ։
Օգտակար օրգանները, ինչքան էլ նրանք քիչ զարգացած լինեն, չեն կարող թերաճ հարմարվել մինչև այնքան ժամանակ, քանի դեռ մենք հիմք չունենք կարծելու, որ նրանք առաջ եղել եկ ավելի զարգացած։ Նրանք կարող են ներկայացնել միայն առաջին ստադիաներ, որոնք դեռ հետագա զարգացում պիտի ստանան։ Մյուս կողմից թերաճ օրգանները կամ բոլորովին անօգուտ են, ինչպես, օրինակ, լնդերքի միջից երբեք դուրս չեկող ,ատամները, կամ քիչ օգտակար են, ինչպես ջայլամի թևերը, որոնք միայն որպես առագաստներ են ծառայում։ Քանի որ այդպիսի օրգաններն իրենց նախկին վիճակում, երբ դեռ քիչ զարգացած են եղել, նույնքան քիչ են գործածվել, ինչքան որ այժմ, ուստի նրանք իհարկե չէին կարող փոփոխության միջոցով զարգանալ բնական ընտրության ազդեցության տակ, որովհետև այդ ազդեցությունն արտահայտվում է միայն օգտակար ձևափոխությունների պահպանման մեջ։ Այդպիսի օրգանները մասամբ պահպանվել են ժառանգականության միջոցով և վերաբերում են իրերի նախկին դրությանը։ Սակայն հաճախ դժվար է լինում տարբերություն գտնել թերազարգացած և նոր երևացող օրգանների միջև, որովհետև մենք միայն անալոգիայով կարող ենք դատել, թե արդյոք օրգանն ընդունակ է հետագա զարգացման, և միայն այդ դեպքում նա արժանի է նոր երևացող կամ սկբնավորվող օրգան անվանվելու։ Այդպիսի դրության մեջ գտնվող օրգանները բավական քիչ են հանդիպում, որովհետև դրանցով օժտված օրգանիզմները սովորաբար դուրս են մղվում կամ փոխարինվում են իրենց հաջորդներով, որոնք ունեն նույն օրգաններն ավելի զարգացած վիճակում, և հետևաբար մահաջնջվում են։ Պինգվինի թևը, գործելով որպես լողաթև, մեծ նշանակություն ունի, հետևապես նա կարող է ներկայացնել որպես թևի առաջացման վաղ ստադիա, թեև ես այդ չեմ կարծում․ բայց ավելի հավանական է, որ դա թերաճած օրգան է, որը փոփոխվել է նոր ֆունկցիայի համար. մյուս կողմից, կիվիի թևը բոլորովին անօգուտ է և իսկապես թերաճ է։ Օուենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] լեպիդոսիրեն ձկան պարզ թելանման վերջավորությունները համարում է «այնպիսի օրգանների սկզբնավորումներ, որոնք լրիվ ֆունկցիոնալ զարգացման են հասնում բարձրակարգ ողնաշարավորների մոտ», բայց համաձայն դոկտոր Գյունտերի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 154] վերջերս պաշտպանած հայացքների, ամենայն հավանականությամբ, այդ մնացորդներն են, որոնք բաղկացած են լողաթևի պահպանված առանցքից, որը կորցրել է կողմնային ճառագայթները կամ ճյուղերը։ Բադակտուցի կաթնագեղձերը կովի կրծի համեմատությամբ կարելի է դիտել որպես նոր երևացող օրգան։ Մի քանի փնջոտնյաների ձվային սանձիկները, որոնք դադարել են ձվերը պահելուց և թույլ են զարգացած, սկզբնավորվող խռիկներ են։
Թերաճ օրգանները միևնույն տեսակին պատկանող տարբեր անհատների մոտ մեծ հակում ունեն փոփոխվելու ինչպես իրենց զարգացման աստիճանի, այնպես էլ այլ տեսակետներից։ Ճիշտ այդպես էլ միևնույն օրգանի կրճատման չափերը շատ մերձավոր տեսակների մոտ երբեմն խիստ զգալիորեն են փոփոխվում։ Վերջինս լավ լուսաբանվում է միևնույն ընտանիքին պատկանող ցեցերի էգերի թևերի վիճակով։ Թերաճ օրգանները, վերջապես, կարող են բոլորովին անհետանալ, և այդ մեզ բացատրում է, թե ինչու մի քանի կենդանիների և բույսերի մոտ բացակայում են այնպիսի մասեր, որոնք անալոգիայով մենք կարող ենք սպասել գտնելու նրանց մոտ և որոնք երբեմն երևան են գալիս այլանդակ անհատների մոտ։ Խլածաղիկների (Scrophulariaceae) մեծամասնության հինգերորդ առէջքը սովորաբար բոլորովին թերազարգացած է. սակայն դատելով նրանից, որ այդ օրգանի թերաճիկը հանդիպում է տվյալ ընտանիքի շատ տեսակների մոտ, մենք կարող ենք գալ այն եզրակացության, որ հինգերորդ առէջքը առաջներում գոյություն է ունեցել․ պատահելիս նրա թերաճիկը լինում է նույնիսկ շատ լավ զարգացած, ինչպես այդ երբեմն կարելի է տեսնել «առյուծաբերանի» (Antirrhinum) ծաղկի վրա։ Միևնույն ղասին պատկանող զանազան անդամների մոտ որևէ հոմոլոգիա հայտնաբերելու համար չկա ավելի սովորական և զանազան մասերի փոխհարաբերությունը հասկանալու տեսակետից ավելի օգտակար միջոց, քան թերաճ օրգանների հայտնագործումը։ Այդ լավ ցույց է տվել Օուենը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 15] ձիու, ցուլի և ռնգեղջյուրի վերջավորությունների ոսկորների նկարների վրա։
Չափազանց կարևոր է այն, որ զանազան օրգաններ, ինչպիսիք են, օրինակ, կետերի և որոճողների վերին ծնոտի վրա ատամները, հաճախ կարող են սաղմի մոտ գտնվել, բայց հետո բոլորովին անհետանում են։ Ես կարծում եմ, կարելի է նաև որպես ընդհանուր կանոն ընդունել այն, որ սաղմի մոտ թերաճ օրգանը ունենում է հարաբերականորեն ավելի մեծ չափեր, քան չափահաս կենդանու մոտ, այնպես որ այդպիսի օրգանն իր զարգացման վաղ ստադիաներում ավելի քիչ թերաճած է լինում և թերևս նույնիսկ արժանի չէ այդպես անվանվելու։ Այդ հիման վրա հաճախ ասում են, որ թերաճիկ օրգանները չափահաս կենդանու մոտ պահպանում են իրենց սաղմնային կառուցվածքը։
Այսպիսով, ես բերեցի թերաճ օրգանների վերաբերյալ ղեկավարող փաստերը։ Խորհելով նրանց վրա, յուրաքանչյուր ոք կզարմանա, որովհետև միևնույն դատողությունը, որը հարկադրում է մեզ օրգանների մեծամասնության մեջ տեսնել հարմարեցում որոշ նպատակների համար, միաժամանակ և բերում է մեզ այն եզրակացության, որ այդ թերաճ կամ ատրոֆիայի ենթարկված օրգաններն անկատար են և անօգուտ։ Բնապատմության վերաբերյալ աշխատություններում թերաճ օրգանները սովորաբար դիտվում են որպես «համաչափության նպատակներով» ստեղծվածներ կամ նրա համար են, որպեսզի «լրացնեն բնության պլանը»։ Բայց դա բացատրություն չէ, այլ միայն փաստի պարզ կրկնություն։ Նա մինչև անգամ իր մեջ պարունակում է ներքին հակասություն, այսպես, բոա-կոնստրիկտորն ունի հետևի վերջավորությունների և կոնքի թերաճիկներ, և եթե ասենք, թե այդ ոսկորներն այդտեղ պահպանվել են «բնության պլանը լրացնելու համար», այդ դեպքում ինչո՞ւ, հարցնենք մենք էլ պրոֆ. Վեյսմանի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 32] հետ միասին, նրանք չեն պահպանվել մյուս օձերի մոտ, որոնք այդ ոսկորների հետքն անգամ չունեն։ Մենք ի՞նչ կմտածենք այն աստղագետի մասին, որը կսկսի պնդել թե արբանյակները մոլորակների շուրջը էլիպսներով պտտվում են «համաչափության նպատակով», որովհետև մոլորակներն էլ նույն ձևով պտտվում են արևի շուրջը։ Մի ականավոր ֆիզիոլոգ թերաճ օրգանների գոյությունը բացատրում է այն ենթադրությամբ, որ նրանք ծառայում են օրգանիզմի մեջ եղած ավելորդ կամ վնասակար նյութերը հեռացնելու համար, բայց արդյոք կարելի՞ է ընդունել, որ մի աննշան պտկիկ, որպիսին հաճախ իրենից ներկայացնում է արական ծաղկի մեջ եղած վարսանդը և կազմված է պարզ բջջային հյուսվածքից, կարող է այդպիսի նշանակություն ունենալ։ Կարելի՞ է արդյոք ընդունել, որ թերաճ ատամները, որոնք հետագայում ատրոֆիայի են ենթարկվում, բարենպաստ են հորթի սաղմի արագ աճման համար այն պատճառով, որ հեռացնում են այնքան թանկարժեք մի նյութ, ինչպիսին է կալցիում ֆոսֆատը։ Մարդու մատը անդամահատելուց հետո երբեմն մատի վրա երևան է գալիս եղունգի թերաճիկ, և մենք նույնպիսի իրավունքով կարող ենք ընդունել, որ եղունգի այդ նշաններն այստեղ զարգանում են եղջերանյութը արտադրելու համար, ինչպես որ մանատի[74] լողաթևերի վրա առաջանում են եղունգների թերաճիկներ։
Հաջորդական փոփոխությունների միջոցով ձևերի զարգացման տեսակետից թերաճ օրգանների գոյությունը բացատրվում է համեմատաբար ավելի պարզ կերպով և հնարավորություն է տալիս ըմբռնելու այն օրենքների լայն նշանակությունը, որոնք ղեկավարում են անկատար զարգացումը։ Մենք ունենք թերաճ օրգանների գոյության շատ դեպքեր մեր ընտանի օրգանիզմների մեջ, օրինակ անպոչ ցեղերի մոտ պոչի թերաճիկը, անականջ ոչխարների մոտ ականջների հետքերը, փոքրիկ, ոչ ամուր կերպով նստած եղջյուրիկները անեղջյուր ցեղերի մոտ, մանավանդ, ինչպես ասում է Իուետը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 50] մատղաշ անհատների մոտ, վերջապես ամբողջ ծաղկի ընդհանուր վիճակը ծաղկակաղամբի մեջ։ Մենք հաճախ տեսնում ենք զանազան օրգանների սաղմերը նաև այլանդակության դեպքերում, բայց ես կասկածում եմ, որ որևէ նման մի դեպք կարողանա լույս սփռել թերաճ օրգանի ծագման վրա բնական դրության մեջ և տար որևէ բան ավելի, քան այն պարզ ցուցումը, որ հնարավոր է այդպիսի թերաճիկների գոյացումը, որովհետև հասարակ ակներևությունը պարզ կերպով ասում է մեզ, որ տեսակները բնական վիճակում մեծ և հանկարծակի փոփոխությունների չեն ենթարկվում։ Բայց մեր ընտանի օրգանիզմների ոաումնասիրությունը մեզ ցույց է տալիս, որ օրգանների անգործածությունը տանում է դեպի նրանց փոքրացումը և անգործածության հետևանքները փոխանցվում են ժառանգաբար։
Հավանական է թվում, որ անգործածությունը գլխավոր գործոնն է հանդիսանում այն բանում, որ օրգանը դառնում է թերաճ։ Թույլ փոփոխությունները կարող են աստիճանաբար առաջացնել օրգանի ավելի ու ավելի մեծ թերազարգացում, մինչև որ վերջապես նա դառնում է թերաճ, ինչպես, օրինակ, քարանձավներում ապրող կենդանիների աչքերի դեպքում և այն թռչունների թևերի նկատմամբ, որոնք բնակվում են օվկիանոսային կղզիների վրա և որոնք շատ քիչ են հարկադրված լինում գիշատիչների հետապնդման պատճառով օդ բարձրանալու, որի հետևանքով վերջապես բոլորովին կորցնում են թռչելու ընդունակությունը։ Կամ թե՝ որոշ պայմաններում օգտակար օրգանը կարող է այլ պայմաններում դառնալ վնասակար, ինչպես, օրինակ, փոքրիկ բաց կղզիների վրա ապրող բզեզների թևերը, այդ դեպքում բնական ընտրությունը մասնակցություն է ունենում օրգանի թերազարգացման մեջ, մինչև որ նրան դարձնում է նշանակություն չունեցող ու թերաճ։
Կառուցվածքի և ֆունկցիայի մեջ աստիճանաբար կատարվող յուրաքանչյուր փոփոխություն գտնվում է բնական ընտրության ներգործության սահմաններում. այսպիսով, այն օրգանը, որ կենցաղի փոփոխության հետևանքով որոշ նպատակների համար դարձել է անօգուտ և նույնիսկ վնասակար, կարող է փոփոխվել և գործածման մեջ դրվել այլ նպատակների համար։ Ճիշտ այդպես էլ օրգանը կարող է պահպանվել իր նախկին ֆունկցիաներից որևէ մեկի համար։ Այն օրգանները, որոնք սկզբնապես ձևավորվել են բնական ընտրության ազդեցության տակ, դառնալով անօգուտ՝ կարող են շատ անկայուն լինել, որովհետև նրանց փոփոխությունները այլևս չեն ամրապնդվում բնական ընտրությամբ։ Այս բոլորը միանգամայն համընկնում են բնական պայմաններում մեր տեսածի հետ։ Բացի դրանից, անգործածությունը կամ ընտրությունը կյանքի ինչ շրջանում էլ օրգանը դարձնեն թերաճ,— սովորաբար այդ տեղի է ունենում, երբ օրգանիզմը հասել է հասունության և ձեռք է բերել իր ընդունակությունների լիակատար զարգացում,— փոփոխությունները համապատասխան հասակում ժառանգաբար փոխանցելու օրենքի համաձայն օրգանը հակում է ցուցաբերում հասուն հասակում թերաճ լինելու և հազվագյուտ դեպքում այդպես է լինում սաղմնային վիճակում։ Սրանից մեզ հասկանալի են դառնում թերաճ օրգանների մեծ չափերը սաղմի մոտ՝ համեմատած հարակից մասերի հետ, և նրանց հարաբերական փոքրությունը չափահաս կենդանու մոտ։ Եթե, օրինակ, չափահաս կենդանու մատը կենցաղի որոշ փոփոխությունից կախված մի քանի սերունդների ընթացքում շարունակ ավելի ու ավելի քիչ գործածվի կամ եթե որևէ օրգան կամ գեղձ շարունակ ավելի ու ավելի քիչ աշխատի, մենք կարող ենք սպասել, որ նրանց մեծությունը որոշ կենդանու հաջորդների մեջ չափահաս վիճակում կփոքրանա և մոտավորապես իր նորմալ չափերը կպահպանի սաղմի մեջ։
Սակայն մնում է դեռ հետևյալ դժվարությունը։ Երբ օրգանի գործածությունն արդեն դադարել է և այդ պատճառով նա զգալիորեն կրճատվել է, արդյոք անգործածությո՞ւնն է հարկադրում նրան ավելի ևս փոքրանալու այնքան, մինչև, որ նրանից աննշան հետքեր կմնան, և արդյոք այդ նրան կհասցնի՞ լիակատար ոչնչացման։ Հազիվ թե կարելի է ընդունել, որ անգործածությունը կարող է որևէ փոփոխություն ևս առաջացնել, երբ օրգանն արդեն դադարել է գործելուց։ Այդ դեպքում անհրաժեշտ է որոշ լրացուցիչ բացատրություն, որը տալ ես չեմ կարող։ Եթե, օրինակ, ապացուցվեր, որ օրգանիզմի յուրաքանչյուր մաս հակում ունի ավելի փոքրանալու, քան մեծանալու ուղղությամբ փոփոխվելու, այն ժամանակ կարելի կլիներ հասկանալ, թե ինչպես անօգուտ դարձած օրգանը, անգործածության ազդեցությունից անկախ կերպով դարձել է թերաճ և վերջապես իսպառ անհետացել է, որովհետև այն փոփոխությունները, որոնք տանում են դեպի օրգանի փոքրացումը, այդ դեպքում կդադարեն բնական ընտրության ազդեցությամբ պահպանվելուց։ Նախորդ գլխում պարզաբանված աճման խնայողության սկզբունքը, որի շնորհիվ տիրոջ համար որևէ օրգանի անօգտակարության դեպքում նրա կազմող նյութերը պահպանվում են հնարավորին չափ երկար ժամանակ, գուցե իր ազդեցությունն է ունենում չգործող օրգանը թերաճ օրգան դառնալու գործում։ Բայց այդ սկզբունքը գրեթե անհրաժեշտորեն պետք է կիրառվի ռեդուկցիայի պրոցեսի միայն սկզբնական ստադիաներում, որովհետև մենք չենք կարող կարծել, որ, օրինակ, այն փոքրիկ պտկիկը, որն արական ծաղկի մեջ ներկայացնում է պտղաբերող ծաղկի վարսանդը և կազմված է պարզապես բջջային հյուսվածքից, կարող է էլ ավելի փոքրանալ կամ նույնիսկ անհետանալ սննդի խնայողության նպատակով։
Վերջապես, քանի որ օրգանները, ինչ ճանապարհով էլ որ նրանք փոխարկված լինեն իրենց այժմյան անօգտակար վիճակին, որպես ցուցում են ծառայում իրերի ավելի վաղ դրության մասին և պահպանվել են բացառապես ժառանգականության շնորհիվ, ուստի մենք կարող ենք հասկանալ, հետևելով ծննդաբանական դասակարգությանը, թե ինչու սիստեմատիկները, օրգանիզմները զետեղելով բնական սիստեմի մեջ նրանց պատշաճ տեղերում, հաճախ թերաճ օրգանները համարել են նույնքան կամ նույնիսկ ավելի օգտակար, քան ֆիզիոլոգիական մեծ նշանակություն ունեցող օրգանները։ Թերաճ օրգանները կարելի է համեմատել այն տառերի հետ, որոնք ուղղագրության մեջ պահպանվում են, բայց արտասանության մեջ դարձել են անօգուտ, ծառայելով որպես բանալի նրա ծագումը բացատրելու համար։ Կողմնակից լինելով օրգանիզմների փոփոխության միջոցով նրանց ծագման տեսակետին, մենք տեսնում ենք, որ թերաճ, անկատարյալ, անօգուտ և նույնիսկ բոլորովին թերազարգացած օրգանների գոյության բացատրությունը բնավ այնպիսի հսկայական դժվարություն չի ներկայացնում, ինչպես անկասկած ներկայացրել է անկախ ստեղծագործության հին ուսմունքի ժամանակ, և կարող է լիովին համաձայնեցվել այստեղ շարադրված տեսակետների հետ։
ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆՆԵՐ
Այս գլխում ես աշխատել եմ ցույց տալ, որ՝ բոլոր դարաշրջանների բոլոր օրգանիզմների դասավորումը ստորադաս խմբերի մեջ. այն ազգակցական կապերի բնույթը, որոնցով բոլոր գոյություն ունեցող և մահաջնջված օրգանիզմները կապվում են միմյանց հետ բարդ, տարամիտվող ու շրջանաձև գծերով մի քանի ընդարձակ դասերի մեջ. այն կանոնները, որոնց հետևում են բնախույզները, և այն դժվարությունները, որոնց հանդիպում են նրանք իրենց դասակարգությունները կառուցելիս, զանազան հատկանիշներին տրվող նշանակությունը, այսինքն՝ նրանց կայունությունը, գերակշռությունը, խոշոր կամ չնչին նշանակությունը կամ, ինչպես թերաճ օրգանների դեպքում, նշանակության լիակատար բացակայությունը. այն հսկայական տարբերությունը, որ գոյություն ունի անալոգ կամ հարմարողական առանձնահատկությունների և աներկբայելի ազգակցության դեպքերի միջև և այլ նման տվյալներ,— այս բոլորը բնական հետևանք են հանդիսանում, երբ ընդունում ենք մերձավոր ձևերի ընդհանուր ծագումը, նրանց զարգացումը փոփոխության և բնական ընտրության միջոցով, ինչպես նաև նրանց մահաջնջումը և հատկանիշների տարամիտումը։ Դասակարգության վերաբերմամբ այս տեսակետն ունենալով հանդերձ, չպետք է մոռանալ, որ ծագումն ընդհանրապես օգտագործվում է զանազան սեռեր, հասակներ, դիմորֆ ձևեր միացնելիս, ինչպես նաև մի տեսակի պատկանող տարատեսակները որոշելիս, ինչքան էլ նրանք իրենց կառուցվածքով խիստ կերպով տարբերվեն միմյանցից։ Ընդարձակելով օրգանիզմների նմանության միակ հայտնի պատճառ հանդիսացող ընդհանուր ծագման գաղափարի օգտագործումը, մենք կհասկանանք նաև այն, թե ինչ է հասկացվում բնական սիստեմ ասելով․ դա ծննդաբանական դասակարգության մի փորձ է, որի մեջ ձեռք բերված տարբերության զանազան աստիճաններն արտահայտվում են տարատեսակներ, տեսակներ, սեռեր, ընտանիքներ, կարգեր և դասեր տերմիններով։
Հաջորդական փոփոխությունների միջոցով զարգացման վերաբերյալ տեսակետի հիման վրա մեզ հասկանալի է դառնում կարևոր մորֆոլոգիական տվյալների մեծ մասը, երթ մենք կանգ ենք առնում ինչպես մի դասի պատկանող զանազան տեսակների հոմոլոգ օրգանների կառուցվածքի ընդհանուր պլանի վրա՝ անկախ այդ օրգանների կատարած դերից, այնպես էլ կենդանական ու բուսական յուրաքանչյուր անհատի շարքային և կողմնային[75] հոմոլոգիաների վրա։
Այն օրենքի համաձայն, որ հաջորդական փոքրիկ փոփոխությունները երևան են գալիս ոչ անպայման և ոչ էլ բոլորի մոտ կյանքի շատ վազ հասակում և նրանց ժառանգական փոխանցումը տեղի է ունենում համապատասխան հասակում, մեզ բացատրում են սաղմնաբանության առաջատար փաստերը, այսինքն՝ սաղմի մոտ հոմոլոգ մասերի մեծ նմանությունը, որոնք հասունացած հասակում իրենց կառուցվածքով ու կատարած դերով դառնում են խիստ տարբեր․ միմյանց հետ մերձավոր, բայց տարբեր տեսակների հոմոլոգ մասերի կամ օրգանների նմանությունը, թեև նրանք հասուն վիճակում հարմարվում են չափազանց տարբեր նպատակների։ Թրթուրները գործունյա սաղմեր են, որոնք ավելի կամ պակաս չափով փոփոխվել են՝ նայած իրենց կենցաղին և համապատասխան վաղ հասակում փոփոխությունների ժառանգմանը։ Այս սկզբունքների օգնությամբ և ուշադրություն առնելով նաև այն, որ օրգաններն իրենց չափերով փոքրանում են անգործածության կամ բնական ընտրության հետևանքով սովորաբար կյանքի այնպիսի շրջանում, երբ օրգանիզմը ինքն է հոգում իր մասին, ինչպես նաև հիշելով, թե որքան մեծ է ժառանգականության ուժը, մենք ի վիճակի կլինենք նույնիսկ բացատրելու թերաճ օրգանների գոյությունը։ Սաղմնաբանական տվյալների և թերաճ օրգանների նշանակությունը հասկանալի է, եթե մենք հետևենք այն տեսակետին, որ բնական դասակարգությունը պետք է լինի ծննդաբանական։
Վերջապես, այս գլխում քննարկված փաստերի զանազան խմբերը, ըստ իս, այնքան պարզորոշ կերպով են ցույց տալիս այն, որ երկրագնդի վրա բնակվող անհամար տեսակները, սեռերը և ընտանիքները, յուրաքանչյուրն իր դասի կամ խմբի սահմաններում, ծագել են ընդհանուր նախածնողներից և այնուհետև փոփոխվել են հաջորդական զարգացման ընթացքում, որ ես առանց տատանվելու կընդունեի այդ տեսակետը, եթե նույնիսկ այն չհաստատվեր այլ փաստերով և այլ ապացույցներով։
ԳԼՈՒԽ XV։ ՀԱՄԱՌՈՏ ԿՐԿՆՈՒԹՅՈՒՆ ԵՎ ԸՆԴՀԱՆՈՒՐ ԵԶՐԱԿԱՑՈՒԹՅՈՒՆ
ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՏԵՍՈՒԹՅԱՆ ԴԵՄ ԵՂԱԾ ԱՌԱՐԿՈՒԹՅՈՒՆՆԵՐԻ ՀԱՄԱՌՈՏ ԿՐԿՆՈՒԹՅՈՒՆ։— ՆՐԱ ՕԳՏԻՆ ԽՈՍՈՂ ԸՆԴՀԱՆՈՒՐ Եվ ՄԱՍՆԱՎՈՐ ՀԱՆԳԱՄԱՆՔՆԵՐԻ ՀԱՄԱՌՈՏ ԿՐԿՆՈՒԹՅՈՒՆ։— ՏԵՍԱԿՆԵՐԻ ԱՆՓՈՓՈԽԵԼԻՈՒԹՅԱՆ ՄԱՍԻՆ ԵՂԱԾ ԸՆԴՀԱՆՈՒՐ ՀԱՎԱՏԻ ՊԱՏՃԱՌՆԵՐԸ։— ԲՆԱԿԱՆ ԸՆՏՐՈՒԹՅԱՆ ՏԵՍՈՒԹՅԱՆ ԿԻՐԱՌՄԱՆ ՍԱՀՄԱՆՆԵՐԸ։— ԱՅԴ ՏԵՍՈՒԹՅԱՆ ԸՆԴՈՒՆՄԱՆ ՀԵՏԵՎԱՆՔՆԵՐԸ ԲՆԱՊԱՏՄՈՒԹՅԱՆ ՈՒՍՈՒՄՆԱՍԻՐՈՒԹՅԱՆ ԳՈՐԾՈՒՄ։— ԵԶՐԱՓԱԿՈՒՄ։
Քանի որ այս ամբողջ գիրքը ծայրից ծայր միայն ապացույցներ է, ուստի ընթերցողի համար թերևս անօգուտ չի լինի նրա մեջ եղած ղեկավար փաստերի և ընդհանուր եզրակացությունների համառոտ կրկնությունը։
Ես չեմ ժխտում, որ կան բազմաթիվ և ծանրակշիռ առարկություններ, որոնք կարող են առաջադրվել փոփոխության և բնական ընտրության միջոցով տեսակների ծագման տեսության դեմ։ Ես աշխատել եմ շարադրել դրանք, պահպանելով նրանց ամբողջ ուժը։ Առաջին հայացքից գրեթե անհնարին է թվում, որ ամենաբարդ օրգաններն ու բնազդները կարող էին առաջանալ ու կատարելագործվել ոչ թե այնպիսի ներգործությունների միջոցով, որոնք թեև նման են մարդկային բանականությանը, բայց անչափելի կերպով գերազանցում են նրան, այլ այնպիսի անթիվ, աննշան փոփոխությունների աստիճանական կուտակման միջոցով, որոնցից յուրաքանչյուրը օգտակար է եղել դրանցով օժտվածներին։ Այնուամենայնիվ այդ դժվարությունը, թեև նա մեր երևակայությանը անհաղթահարելի է թվում, չի կարելի իրական համարել, եթե միայն ընդունենք հետևյալ դրույթները, որ կազմվածքի բոլոր մասերը, ինչպես նաև բնազդները համենայն դեպս ներկայացնում են անհատական տարբերություններ. որ կա գոյության կռիվ, որի հետևանքով պահպանվում են կառուցվածքի կամ բնազդի օգտակար փոփոխությունները և, վերջապես, որ գոյություն են ունեցել յուրաքանչյուր օրգանի կատարելագործման աստիճաններ, որոնցից ամեն մեկն ինքն ըստ ինքյան օգտակար է եղել։ Այս դրույթների ճշտությունը, ես կարծում եմ, չի կարող վիճարկվել։
Կասկածի ենթակա չէ, որ չափազանց դժվար է պատկերացնել, թե շատ կառուցվածքներ ինչպիսի աստիճաններով կարող էին անցնել դեպի կատարելագործություն տանող ճանապարհը, մանավանդ օրգանական էակների մանրատված և թերի խմբերում, որոնք շատ են տուժել մահաջնջումից. բայց մենք բնության մեջ հանդիպում ենք այնպիսի ամենատարօրինակ անցումների, որ պետք է չափազանց շրջահայաց լինենք, պնդելով՝ թե այս կամ այն օրգանը կամ բնազդը, կամ ամբողջ օրգանիզմը չէին կարող հասնել արդի վիճակին հաջորդական մի շարք աստիճաններով։ Պետք է ընդունենք, որ կան դեպքեր, որոնք առանձին դժվարություններ են ներկայացնում բնական ընտրության տեսության համար․ նման դեպքերի մեջ ամենահետաքրքրականներից մեկն է մրջյունների միևնույն համայնքում աշխատավորների կամ անպտուղ էգերի երկու կամ երեք որոշակի կաստաների գոյությունը, ես աշխատել եմ ցույց տալ, թե ինչպես կարելի է այդ դժվարությունները վերացնել։
Ինչ վերաբերում է այն գրեթե ընդհանուր կանոնին, որի համաձայն տեսակներն իրենց առաջին խաչաձևման ժամանակ անպտղաբեր են լինում որով նրանք խիստ տարբերվում են տարատեսակներից, որոնք այդ պայմաններում գրեթե միշտ պտղաբեր են, ես պետք է ընթերցողներին հանձնարարեմ դիմելու իններորդ գլխի վերջում բերված փաստերի ընդհանուր ամփոփմանը։ Այդ ամփոփումը, ինձ թվում է, համոզեցուցիչ կերպով ապացուցում է, որ անպտղաբերությունը տեսակների համար նույնքան քիչ յուրահատուկ հատկություն է, որքան որ երկու տարբեր ծառատեսակների փոխադարձաբար պատվաստվելու անընդունակությունը. թե՛ մեկ և թե՛ մյուս դեպքում մենք պետք է տեսնենք զուգընթաց երևույթներ, որոնք կախված են խաչաձևվող տեսակների վերարտադրողական սիստեմների տարբերության աստիճանից։ Այդ եզրակացության ճշտությունը երևան է գալիս նույն տեսակների փոխադարձ խաչաձևումների արդյունքների խոր տարբերության մեջ, այսինքն այն դեպքում, երբ միևնույն տեսակը մերթ հոր, մերթ մոր դեր է կատարում։ Նույն եզրակացությանն է բերում նաև անալոգիան դիմորֆ և տրիմորֆ բույսերի նկատմամբ, երբ այդ ձևերը ենթարկվում են ապօրինի բեղմնավորման, նրանք տալիս են քիչ սերմեր կամ բոլորովին չեն տալիս, իսկ դրանից առաջացած սերունդը լինում է ավելի կամ պակաս չափով անպտղաբեր։ Իսկ մինչդեռ այդ ձևերը անկասկածելիորեն պատկանում են միևնույն տեսակին և ոչնչով չեն տարբերվում մեկը մյուսից, բացի իրենց վերարտադրության օրգանների կառուցվածքի և գործունեության մեջ եղած առանձնահատկություններից։
Թեպետ և տարատեսակների պտղաբերությունը նրանց խաչաձևման ժամանակ, ինչպես նաև նրանցից ծագում առած խառնացեղերի խաչաձևման պտղաբերությունը շատ հեղինակներ համարում են որպես ընդհանուր կանոն, այնուամենայնիվ այդ տեսակետը չի կարելի լիովին ճիշտ համարել նկատի ունենալով բերված փաստերը, որոնք հիմնվում են այնպիսի բարձր հեղինակությունների վրա, ինչպիսիք են Գերտները[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 83] և Կյոլռեյտերը։[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 33] Այն տարատեսակների մեծ մասը, որոնց վրա կատարվել են փորձերը, ստացված է եղել ընտանեցման կամ կուլտուրական վիճակում։ Բայց քանի որ ընտանեցումը (ես դրա տակ բացառապես միայն նեղ գերություն չեմ հասկանում) գրեթե միշտ վերացնում է այն անպտղաբերությունը, որը անալոգիայի հիման վրա, հավանորեն, պետք է երևան գար նախահայր տեսակների խաչաձևման հետևանքով, ուստի մենք հիմք չունենք սպասելու, որ ընտանեցման հետևանքը կարող էր լինել նրանց փոփոխված հաջորդների անպտղարերությունը խաչաձևման ժամանակ։ Անպտղաբերության այդ վերացումը, հավանորեն, տեղի է ունենում նույն պատճառներից, որոնք թույլ են տալիս մեր ընտանի կենդանիներին ազատ կերպով բազմանալ ամենաբազմազան պայմաններում, իսկ դա իր հերթին, ըստ երևույթին, բխում է նրանից, որ նրանք աստիճանաբար ընտելացել են իրենց կենսապայմանների հաճախակի փոփոխություններին։
Փաստերի կրկնակի և զուգահեռ շարքը, ըստ երևույթին, նոր լույս է սփռում տեսակների առաջին խաչաձևման ժամանակ ստացվող անպտղաբերության և այդ խաչաձևումից առաջացող հիբրիդների անպտղաբերության հարցի վրա։ Մի կողմից, մենք լիակատար հիմք ունենք ենթադրելու, որ կենսապայմանների մեջ տեղի ունեցած աննշան փոփոխությունները նպաստում են բոլոր օրգանական էակների ուժերի և պտղաբերության ավելացմանը։ Մենք գիտենք նաև, որ միևնույն տարատեսակի տարբեր անհատների կամ տարբեր տարատեսակների միջև կատարված խաչաձևումը շատացնում է նրանց հաջորդների թիվը և անկասկած դարձնում է նրանց ավելի խոշոր և ուժեղ։ Դա գլխավորապես կախված է նրանից, որ խաչաձևման ենթարկվող ձևերը նախապես դրվել են փոքր ինչ տարբեր կենսապայմանների մեջ, որովհետև ինձ հաջողվել է մի շարք ամենամանրազնին փորձերով ապացուցել, որ եթե միևնույն տարատեսակի անհատները մի քանի սերունդների ընթացքում ենթարկվել են միանման պայմանների ներգործությանը, այդ դեպքում խաչաձևումից ստացվող օգուտը հաճախ նվազում է կամ նույնիսկ իսպառ կորչում։ Դա հարցի մի կողմն է։ Մյուս կողմից, մենք գիտենք, որ երկար ժամանակամիջոցի ընթացքում գրեթե միանման պայմաններում գտնվող տեսակները երբ ընկել են գերության մեջ միանգամայն նոր ու խիստ փոփոխված պայմանների մեջ, ապա նրանք կամ ոչնչացել են կամ էլ, եթե ողջ են մնացել, դարձել են անպտղաբեր, միաժամանակ մնալով բոլորովին առողջ։ Մեր ընտանի ցեղերի հետ, որոնք երկար ժամանակ ենթարկվել են փոփոխական պայմանների, այդ տեղի չի ունենում և եթե տեղի է ունենում, ապա շատ թույլ չափով։ Ուստի, երբ մենք հանդիպում ենք այն փաստին, որ երկու առանձին տեսակների խաչաձևման միջոցով առաջացած հիբրիդները շատ սակավաթիվ են, կամ այն պատճառով, որ նրանք մեռնում են սաղմնավորվելուց կարճ ժամանակ հետո, կամ ամենավաղ հասակում, կամ թե այն պատճառով, որ ողջ մնալով, նրանք ավելի կամ պակաս չափով անպտուղ են լինում, մեզ վերին աստիճանի հավանական է թվում, որ այդ արդյունքը կենսապայմանների այն խոր փոփոխության հետևանք է, որ տեղի է ունենում, երբ մի էակի մեջ միաձուլվում են երկու տարբեր կազմվածքներ։ Նա, ով կկարողանար որոշակիությամբ բացատրել, թե ինչու փիղը և աղվեսը չեն բազմանում գերության մեջ նույնիսկ իրենց հայրենիքում[76], մինչդեռ ընտանի խոզը կամ շունը բազմանում են ամենաբազմազան պայմաններում, կարող էր նաև որոշակի պատասխան տալ այն հարցին, թե ինչու երկու տարբեր տեսակներ խաչաձևվելիս, ինչպես նաև նրանց հիբրիդները, սովորաբար ավելի կամ պակաս չափով լինում են անպտղաբեր, մինչդեռ երկու ընտանի տարատեսակներ, ինչպես նաև նրանց խառնացեղերը խաչաձևվելիս միանգամայն պտղաբեր են լինում։
Անցնելով աշխարհագրական տարածմանը, մենք տեսնում ենք, որ փոփոխության միջոցով ծագման տեսությունը հանդիպում է բավական լուրջ դժվարությունների։ Միևնույն տեսակին պատկանող բոլոր անհատները, ինչպես նաև միևնույն սեռի տեսակները կամ նույնիսկ ավելի ընդարձակ խմբեր առաջացել են ընդհանուր ծնողներից, և հետնաբար, ինչքան էլ մեկուսացած ու մեկը մյուսից հեռացած լինեն աշխարհի այն մասերը, որտեղ նրանք այժմ հանդիպում են, նրանք հաջորդական սերունդների ընթացքում պետք է տեղափոխվեին մի որևէ կետից դեպի մնացած բոլոր վայրերը։ Մենք նույնիսկ ի վիճակի չենք կռահելու, թե այդ ինչպես կարող էր կատարվել։ Այնուամենայնիվ, քանի որ մենք լիակատար հիմք ունենք ենթադրելու, որ մի քանի տեսակներ պահպանել են նույն տեսակային ձևերը երկարատև ժամանակաշրջանների ընթացքում, հսկայական ժամանակաշրջանների, եթե դրանք տարիներով չափենք, ապա չպետք է առանձին նշանակություն տանք միևնույն տեսակի լայն տարածման պատահած դեպքերին, որովհետև այդպիսի երկարատև ժամանակաշրջանների ընթացքում կարող էին գտնվել հանգամանքներ, որոնք այս կամ այն ճանապարհով նպաստել են տարաբնակմանը։ Տարաբնակման մարզի մասերի բաժանված կամ ընդհատուն լինելը կարող է հեշտությամբ բացատրվել միջակա գոտիներում եղած տեսակների անհետացմամբ։ Չի կարելի ժխտել, որ մենք դեռևս շատ քիչ բան գիտենք վերջին ժամանակաշրջանների ընթացքում երկրի երեսին տեղի ունեցած կլիմայական և աշխարհագրական փոփոխությունների մասին, իսկ այդպիսի փոփոխությունները հաճախ կարող էին նպաստել տարաբնակմանը։ Որպես օրինակ ես աշխատել եմ ցույց տալ, թե ինչքան մեծ է եղել սառցադաշտային ժամանակաշրջանի ազդեցությունը միևնույն կամ ազգակից տեսակների տարածման վրա երկրագնդի երեսին։ Մենք ոչինչ չգիտենք օրգանիզմների տարածման պատահական շատ եղանակների մասին։ Քանի որ հեռավոր ու իրարից անջատված երկրներում ապրող, միևնույն սեռին պատկանող տարբեր տեսակների վերաբերմամբ փոփոխության պրոցեսը անհրաժեշտորեն շատ դանդաղ է կատարվել, ապա այդպիսի երկարատև ժամանակաշրջանում կարող էին հնարավոր լինել տարածման ամեն եղանակներ. այսպիսով միևնույն սեռին պատկանող տեսակների լայն տարաբնական բացատրության դժվարությունը որոշ չափով նվազում է։
Նկատի ունենալով այն, որ բնական ընտրության տեսության հիման վրա, մենք պետք է ընդունենք անհամար միջակա ձևերի գոյությունը, որոնք շաղկապել են միևնույն խմբի տեսակները միմյանց հետ նույնքան աննկատելի անցումներով, որքան որ աննկատելի են անցումները գոյություն ունեցող տարատեսակների միջև, ապա կարող են հարցնել, իսկ ինչո՞ւ այդ շաղկապող օղակները ամենուրեք չեն շրջապատում մեզ։ Ինչո՞ւ բոլոր օրգանական էակները չեն միաձուլվում մի ընդհանուր անլուծելի քաոսի մեջ։ Ապրող ձևերի վերաբերմամբ մենք պետք է հիշենք, որ մենք իրավունք չունենք սպասելու (բացառությամբ հազվագյուտ դեպքերի), որ կհայտնաբերենք ոչ թե այնպիսի օղակներ, որոնք ուղղակի շաղկապում են նրանց իրար հետ, այլ միայն այնպիսիները, որոնք շաղկապում են նրանց որևէ դուրս մղված ու մահաջնջված ձևերի հետ։ Նույնիսկ ընդարձակ տարածությունների վրա, որոնք երկարատև ժամանակաշրջանների ընթացքում պահպանել են իրենց անընդհատությունը և որտեղ կլիմայական ու այլ կենսապայմանները մի տեսակի գրաված մարզից նրան մերձավոր տեսակի գրաված մի ուրիշ մարզ անցնելիս փոփոխվել են անզգալիորեն, նույնիսկ այդտեղ էլ մենք հիմք չունենք հաճախ հուսալու, որ կհանդիպենք միջակա գոտում միջակա տարատեսակների։ Այնուամենայնիվ մենք հիմք ունենք ենթադրելու, որ միևնույն սեռին պատկանող տեսակներից միայն քչերն են ենթարկվում փոփոխությունների, իսկ մնացած տեսակները մահաջնջվում են, իրենցից հետո չթողնելով սերունդներ։ Փոփոխվող տեսակների թվից էլ միայն քչերն են փոփոխվում նույն երկրում, միևնույն ժամանակ, և բոլոր փոփոխությունները չափազանց դանդաղ են կատարվում։ Ես ցույց եմ տվել նաև, որ միջակա տարատեսակներին, որոնք սկզբում հավանորեն գոյություն են ունեցել միջակա գոտիներում, հեշտությամբ կարող են դուրս մղել նրանց հետ այս ու այն կողմից շփման մեջ մտնող ազգակից ձևերը, որովհետև այս վերջինները որպես անհատների մեծ թիվ ունեցողներ սովորաբար փոփոխվում և կատարելագործվում են ավելի արագ կերպով, քան սակավաթիվ միջակա տարատեսակները, այնպես որ այդ միջակա տարատեսակները վերջ ի վերջո պիտի դուրս մղվեն և բնաջնջվեն։
Բայց եթե ճիշտ է այդ ուսմունքը, որի համաձայն բնաջնջվել են այն անթիվ օղակները, որոնք շաղկապում են գոյություն ունեցող ձևերը անհետացածների հետ, ինչպես նաև իրար հաջորդող յուրաքանչյուր ժամանակաշրջանի սահմաններում անհետացած ձևերը ավելի հին ձևերի հետ, ապա ինչո՞ւ ամեն մի երկրաբանական ֆորմացիա լիքը լցված չէ այդպիսի օղակներով։ Ինչո՞ւ բրածոների յուրաքանչյուր հավաքածու չի ներկայացնում օրգանական ձևերի աստիճանական անցումների և փոխարկումների ակներև հետքեր։ Թեև երկրաբանական հետազոտությունը անկասկածն լիորեն երևան է բերել, որ անցյալում գոյություն են ունեցել բազմաթիվ օղակներ, որոնք շաղկապել են միմյանց հետ զգալի թվով կենդանի էակներ, բայց և այնպես նա երևան չի հանել գոյություն ունեցած և այժմյան տեսակների միջև այն բազմաթիվ նուրբ անցումները, որոնք հարկավոր են մեր տեսությանը, և հենց այդ է ամենաակընհայտը այն բազմաթիվ առարկություններից, որ կարող են ներկայացվել նրա դեմ։ Այնուհետև հարց է առաջանում, թե ինչո՞ւ իրար ազգակից տեսակների ամբողջ խմբեր կարծես հանկարծակի են երևան գալիս երկրաբանական ասպարեզում, թեև հանկարծակի երևան գալու մասին պատկերացումը հաճախ կեղծ է լինում։ Թեպետ մենք այժմ գիտենք, որ օրգանական կյանքը մեր մոլորակի վրա երևան է եկել մեզանից անչափելի հեռավոր ժամանակաշրջանում, կեմբրիական սիստեմի ամենաստորին շերտերի կուտակումից շատ առաջ, հապա ինչո՞ւ այդ սիստեմի կուտակված շերտերի տակ չենք հանդիպում դարձյալ այնպիսի շերտերի, որոնք լիքն են կեմբրիական բրածոների նախնիների մնացորդներով։ Չէ՞ որ այս տեսության հիման վրա շերտերը պետք է որ գոյություն ունենային մեր երկրի այն հեռավոր, մեզ բոլորովին անհայտ պատմական դարաշրջաններում։
Այդ բոլոր հարցերին և առարկություններին ես կարող եմ պատասխանել միայն ենթադրությամբ, որ երկրաբանական տարեգրությունը զգալիորեն ավելի թերի է, քան այդ ենթադրում է երկրաբանների մեծամասնությունը։ Մեր բոլոր թանգարաններում գտնվող օրինակների թիվը միանգամայն չնչին է այն անհամար սերունդների, անթիվ տեսակների համեմատությամբ, որոնք անկասկած գոյություն են ունեցել։ Երկու կամ մի քանի տեսակների նախահայր ձևը չի կարող անմիջականորեն միջակա հանդիսանալ իր փոփոխված հաջորդների միջև, ճիշտ այնպես, ինչպես ժայռաղավնին իր պոչի և կտնառքի ձևով միջակա չի հանդիսանում իր երկու հաջորդների՝ սիրամարգանմանի և փքունի միջև։ Մենք ի վիճակի չենք մի տեսակ համարել որպես մի այլ փոփոխված տեսակի նախահայր, ինչքան էլ որ մենք ուշադիր կերպով նրանց հետազոտենք, եթե մեր տրամադրության տակ չի գտնվում միջակա օղակների մեծ մասը, իսկ երկրաբանական տարեգրության անկատարության հետևանքով մենք ոչ մի իրավունք չունենք հույս ունենալ գտնելու օղակների այդպիսի բազմություն։ Եթե գտնվեն էլ երկու, երեք կամ նույնիսկ ավելի թվով շաղկապող ձևեր, այդ դեպքում շատ բնախույզներ նրանց համապատասխանորեն կդասեին նոր շատ տեսակների շարքը, մանավանդ եթե նրանք գտնվել են տարբեր երկրաբանական հարկերում, թեկուզև այդ ձևերի տարբերության աստիճանը չափազանց փոքր լիներ։ Կարելի էր այժմյան կասկածելի տեսակների շատ դեպքեր նշել, որոնք իրականում, ամենայն հավանականությամբ, միայն տարատեսակներ են, բայց ո՞վ կարող է պնդել, թե ապագայում կհայտնաբերվեն այնքան բրածո օղակներ, որ բնախույզներն ի վիճակի կլինեն լուծելու այն հարցը, թե այդ կասկածելի ձևերը արդյոք պե՞տք է տարատեսակներ համարել, թե ոչ։ Մինչև այժմ երկրաբանորեն հետազոտված է երկրագնդի միայն մի աննշան մասը։ Միայն որոշ դասերի օրգանական էակները կարող են բրածո վիճակում գոնե մի քիչ զգալի քանակությամբ պահպանվել։ Շատ տեսակներ իրենց առաջացումից հետո չեն ենթարկվում հետագա փոփոխության, բայց մահաջնջվում են, չթողնելով իրենցից հետո փոփոխված հաջորդներ. այն ժամանակաշրջանները, որոնց ընթացքում տեսակները փոփոխություններ են կրել, թեև շատ երկար են հղել, եթե նրանց չափենք տարիներով, հավանորեն շատ կարճ են եղել համեմատած այն ժամանակաշրջանների հետ, որոնց ընթացքում տեսակները պահպանել են իրենց կայուն ձևը։ Փոփոխականության ամենից ավելի ենթակա են տիրապետող և լայնորեն տարաբնակված տեսակները, իսկ տարատեսակները հաճախ սկզբում լինում են տեղական,— երկու հանգամանքներ, որոնք ավելի ևս քիչ հավանական են դարձնում շաղկապող օղակների հայտնաբերումը մի ֆորմացիայի սահմաններում։ Տեղական տարատեսակները չեն ներթափանցի այլ և այն էլ հեռավոր երկրներ, քանի դեռ չեն կարողացել զգալիորեն փոփոխվել ու կատարելագործվել․ իսկ երբ նրանք տարածվել են և հայտնաբերվում են մի երկրաբանական ֆորմացիայում, այդ դեպքում նրանք տեղում հանկարծակի ստեղծվածների տպավորություն են թողնում և պարզապես կդասակարգվեն որպես նոր տեսակներ։ Ֆորմացիաների մեծ մասի գոյացումը եղել է ընդմիջվող, իսկ նրանց գոյության տևողությունը, ամենայն հավանականությամբ, ավելի կարճ է եղել, քան տեսակների գոյության միջին տևողությունը։ Հաջորդող ֆորմացիաները շատ դեպքերում բաժանված են մեկը մյուսից հսկայական ժամանակամիջոցներով, որովհետև բրածոներ պարունակող, հետագա քայքայման դիմանալու համար բավականաչափ հաստ շերտերը, որպես ընդհանուր կանոն, կարող էին առաջանալ միայն այնտեղ, որտեղ մեծ նստվածքներ են գոյացել ծովի մշտապես իջնող հատակին։ Հաջորդող բարձրացման կամ մշտական մակարդակի պահպանման ժամանակաշրջաններում երկրաբանական տարեգրությունը պիտի ընդհատվեր, թողնելով դատարկ տեղեր։ Այս վերջին ժամանակաշրջաններում, ամենայն հավանականությամբ, տեղի է ունեցել օրգանական ձևերի գործունյա փոփոխություն. ցած իջնելու ժամանակաշրջաններում գերակշռել է նրանց մահաջնջումը։
Ինչ վերաբերում է կեմբրիական ֆորմացիայից ավելի ցած բրածոներով հարուստ շերտերի բացակայությանը, ապա ես կարող եմ միայն կրկնել տասերորդ գլխում ասածս հիպոթեզը, այսինքն, որ թեև մեր մայր ցամաքները և օվկիանոսները հսկայական ժամանակաշրջանների ընթացքում պահպանվել են գրեթե ներկայիս իրենց ունեցած հարաբերական դրությամբ, այնուամենայնիվ մենք հիմք չունենք ենթադրելու, որ այդ միշտ այդպես է եղել. հետևաբար, այն ֆորմացիաները, որոնք շատ ավելի հին են, քան մեզ հայտնի ֆորմացիաները, կարող են թաղված լինել մեր մեծ օվկիանոսների տակ։ Իսկ ինչ վերաբերում է օրգանական ձևափոխությունների ենթադրյալ շարքի համար, այն ժամանակամիջոցի, որն անցել է մեր մոլորակի պնդանալուց հետո, իբր թե անբավարար լինելուն, ապա այդ առարկությունը, որ ներկայացրել է սըր Վիլյամ Թոմսոնը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 211] հավանորեն մինչև այժմ ներկայացված ամենալուրջ առարկություններից մեկն է։ Ես կարող եմ միայն ասել, նախ՝ որ մենք չգիտենք, թե ինչքան արագ է կատարվում տեսակների ձևափոխությունը, եթե այդ ժամանակը չափենք տարիներով, և երկրորդ՝ որ շատ գիտնականներ մինչև այժմ հնարավոր չեն համարում ընդունել, որ մեր տեղեկությունները տիեզերքի և մեր մոլորակի ընդերքի կառուցվածքի մասին այնքան բավարար են, որպեսզի մենք կարողանայինք վստահելի կերպով դատել նրա անցած գոյության տևողության մասին։
Որ երկրաբանության տարեգրությունն անկատար է, ամեն ոք հոժարությամբ կընդունի, բայց որ նա անկատար է այն չափով, ինչ չափով պահանջվում է մեր տեսության համար, դրա հետ, հավանորեն, շատ քչերը կհամաձայնեն։ Եթե մենք նկատի ունենք բավարար տևողությամբ ժամանակամիջոցներ, այդ դեպքում երկրաբանությունը մեզ կընձեռի պարզ ապացույց, որ բոլոր տեսակները փոփոխվել են և այն էլ այնպես, ինչպես այդ պահանջում է տեսությունս, այսինքն՝ դանդաղ ու աստիճանաբար։ Մենք այդ պարզ կերպով նկատում ենք այն փաստից, որ իրար հաջորդող ֆորմացիաների բրածո մնացորդները միշտ իրար ավելի շատ են նմանվում, քան մեկը մյուսից հեռու գտնվող ֆորմացիաների բրածոները։
Այս է այն ամենագլխավոր առարկությունների և դժվարությունների հանրագումարը, որոնք կարող են իրավացիորեն առաջ բերել իմ տեսության դեմ. ես համառոտակի կրկնեցի նաև այն պատասխաններն ու պարզաբանությունները, որոնք, ինչպես ինձ թվում է, կարող են տրվել։ Երկար տարիների ընթացքում ես խորապես գիտակցել եմ այդ դժվարությունները և դրա համար էլ չեմ կասկածում նրանց ծանրակշռության վրա։ Առանձին ուշադրության արժանի է այն, որ ամենաէական առարկությունները վերաբերում են այն հարցերին, որոնց նկատմամբ մենք պետք է խոստովանենք մեր անտեղյակությունը, մենք նույնիսկ ի վիճակի չենք գնահատելու այդ անտեղյակության ամբողջ խորությունը։ Մեզ անհայտ են ամենապարզ ու ամենաբարդ օրգանի միջև եղած բոլոր հնարավոր միջանկյալ աստիճանները։ Մենք չենք կարող պնդել, թե գիտենք օրգանիզմների տարածման բոլոր բազմազան եղանակները երկար ժամանակաշրջանների ընթացքում, չգիտենք նաև երկրաբանական տարեգրության անկատարության աստիճանը։ Ինչքան էլ էական լինեն այս բոլոր առարկությունները, իմ կարծիքով, նրանք անբավարար են նրա համար, որպեսզի հերքեն իմ տեսությունը, որը վերաբերում է օրգանիզմների ընդհանուր ծագմանը՝ հաջորդական փոփոխությունների միջոցով։
Այժմ դառնանք փաստարկի մյուս կողմին։ Ընտանեցման վիճակում մենք նկատում ենք մեծ չափով փոփոխականություն, որ պատճառում են կամ առնվազն հարուցում են փոփոխված կենսապայմանները, բայց հաճախ այդ փոփոխությունները երևան են գալիս այնպիսի աղոտ ձևով, որ մենք հակվում ենք դրանք ինքնակամ համարելու։ Փոփոխականությունը ղեկավարվում է բազմաթիվ ու բարդ օրենքներով՝ զարգացման հարաբերակցությամբ (կոռելյացիայով), փոխհատուցմամբ (կոմպենսացիայով), մարմնամասերի ուժեղացրած վարժեցմամբ կամ վարժեցման բացակայությամբ և շրջապատող պայմանների որոշակի ներգործությամբ։ Չափազանց դժվար է համոզվել, թե ինչպիսի խոր փոփոխություններ են տեղի ունեցել մեր ընտանի ցեղերի մեջ, բայց մենք համարձակորեն կարող ենք ընդունել, որ այդ փոփոխությունների չափերը մեծ են եղել և որ փոփոխություններն իրենք կարող են ժառանգաբար փոխանցվել երկար ժամանակաշրջանների ընթացքում։ Քանի դեռ կենսապայմանները չեն փոխվում, մենք լիակատար հիմք ունենք ենթադրելու, որ մի քանի սերունդների ժառանգած փոփոխությունները կարող են փոխանցվել գրեթե անսահման թվով սերունդների։ Մյուս կողմից, մենք ապացույցներ ունենք, որ մեր ընտանի ցեղերի մեջ մի անգամ ի հայտ եկած փոփոխականությունը չի դադարում երկար ժամանակաշրջանի ընթացքում. մեզ նույնիսկ անհայտ է, թե արդյոք նա ընդհանրապես դադարում է, որովհետև նոր տարատեսակները դեռ շարունակում են երևան գալ նաև մեր ամենահին ցեղերի մեջ։
Իրականության մեջ մարդը չի ստեղծում փոփոխականություն. նա միայն առանց դիտավորության օրգանական էակներին ենթարկում է նոր կենսապայմանների և այն ժամանակ բնությունը ներգործում է նրանց կազմվածքի վրա և հարուցում փոփոխություններ։ Բայց մարդը կարող է ընտրել և իրոք ընտրում է բնության տրամադրած փոփոխությունները և այդպիսով կուտակում է դրանք իրեն ցանկալի որևէ ուղղությամբ։ Նա այս կերպով հարմարեցնում է կենդանիներին և բույսերին իր պահանջներին կամ քմահաճույքներին։ Նա կարող է այդ անել մեթոդիկորեն կամ անգիտակցաբար, վերջին դեպքում պահելով իրեն համար ամենաօգտակար կամ հաճելի անհատներին՝ առանց ցեղը փոփոխելու որևէ նպատակի։ Կասկածի ենթակա չէ, որ նա կարող է ուժեղ կերպով ազդել ցեղի բնույթի վրա, ընտրելով յուրաքանչյուր հաջորդական անհատական շեղումները, այնքան թույլ շեղումները, որ միայն վարժված աչքը կարող է նկատել դրանք։ Անգիտակցական ընտրության այդ պրոցեսը խոշոր գործոն է հանդիսացել ամենացայտուն կերպով արտահայտված և օգտակար ընտանի ցեղերի առաջացման գործում։ Որ մարդու ստեղծած շատ ցեղեր մեծ չափով բնական տեսակների բնույթ են կրում, ապացուցվում է այն չլուծված կասկածներով, թե նրանցից շատերը արդյոք տարատեսակներ են, թե նախնադարյան ինքնուրույն տեսակներ։
Ոչ մի հիմք չկա կասկածելու, որ ընտանի օրգանիզմների վերաբերմամբ այնպես գործունյա եղած սկզբունքը կարող էր իր ազդեցությունը գործել նաև բնական վիճակում։ Անընդհատ վերսկսվող գոյության կռվում ամենից ավելի բարենպաստյալ անհատների և ցեղերի գերապրման մեջ մենք տեսնում ենք ընտրության հզոր և անընդհատ ներգործող ձևը։ Գոյության կռիվն անխուսափելիորեն բխում է բազմացման արագ աճող երկրաչափա՛կան պրոգրեսիայից, որը հատուկ է բոլոր օրգանական էակներին։ Բազմացման այդ արագ պրոգրեսիան ապացուցվում է հաշիվներով, շատ կենդանիների և բույսերի արագ բազմացմամբ որոշ կլիմայական պայմանների հաջորդական կրկնության դեպքում և նոր երկրներում նատուրալիզացիայի ժամանակ։ Ծնվում են ավելի շատ անհատներ, քան կարող են ապրել։ Բնության կշեռքի վրա մի ձավարահատիկը կարող է որոշել մի անհատի կյանքը և մյուսի մահը, որոշել, թե ո՛ր տեսակը կամ տարատեսակը թվով պիտի ավելանա և որոնք են նվազելու կամ վերջնականապես անհետանալու։ Քանի որ միևնույն տեսակին պատկանող անհատները բոլոր տեսակետներից ամենասերտ մրցության մեջ են մտնում իրար հետ, ուստի նրանց միջև եղած կռիվը ամենից ավելի դաժան կլինի։ Նա գրեթե նույնքան դաժան կլինի միևնույն տեսակին պատկանող տարատեսակների միջև և արդեն փոքր ինչ ավելի թույլ՝ միևնույն սեռին պատկանող տեսակների միջև։ Մյուս կողմից, կռիվը հաճախ դաժան կլինի նաև այն էակների միջև, որոնք բնության սանդուխքի վրա գրավում են միմյանցից հեռու տեղեր։ Մի քանի անհատների ամենաթույլ առավելությունը որոշ հասակում կամ տարվա որոշ ժամանակին նրանց հանդեպ, որոնց հետ նրանք մտնում են մրցության մեջ, կամ ավելի լավ հարմարվածությունը շրջապատող ֆիզիկական պայմաններին, թեկուզ չնչին չափով, կարող է ժամանակի ընթացքում կշեռքը թեքել նրանց կողմը։
Բաժանասեռ կենդանիների արուները շատ դեպքերում կռվում են էգերին տիրանալու համար։ Ամենաուժեղ արուները կամ նրանք, որոնք ամենից ավելի հաջող կերպով են պայքարում կենսապայմանների դեմ, սովորաբար ամենաշատ սերունդներ են թողնում։ Բայց հաջողությունը հաճախ կախված է նաև այն բանից, որ արուներն օժտված են լինում հատուկ զենքով կամ պաշտպանական միջոցով, կամ էլ առանձնապես հրապուրիչ են լինում, ամենաթույլ առավելությունն անգամ կարող է հաղթանակի հասցնել։
Քանի որ երկրաբանությունը պարզ կերպով ցույց է տալիս, որ ամեն մի երկիր ենթարկվել է խոշոր ֆիզիկական փոփոխությունների, ապա մենք իրավունք ունենք սպասելու, որ օրգանական էակները բնական վիճակում փոփոխվել են այնպես, ինչպես որ նրանք փոփոխվել են ընտանեցված վիճակում։ Իսկ երբ բնական վիճակում երևան է եկել փոփոխականությունը, ապա անբացատրելի փաստ կլիներ, եթե բնական ընտրությունը երևան չհաներ իր ներգործությունը։ Հաճախ պնդել են, թեև այդ պնդումը ապացուցման չի ենթարկվում, որ փոփոխականությունը բնական վիճակում սահմանափակվում է շատ նեղ սահմաններում։ Մարդը թեև ներգործում է մի այն արտաքին հատկանիշների վրա և հաճախ առաջնորդվում է քմահաճույքներով, այնուամենայնիվ կարողանում է կարճ ժամանակամիջոցում հասնել զգալի արդյունքների, կուտակելով իր ընտանի ցեղերի մեջ միայն անհատական տարբերությունները. և ոչ ոք իհարկե չի կարող բացասել, որ բնական տեսակներն էլ ունենում են անհատական տարբերություններ։ Սակայն բացի այդ տարբերություններից, բոլոր բնախույզներն ընդունում են նաև բնական տարատեսակների գոյությունը, որոնք այնքան են տարբերվում իրարից, որ նրանց հիշատակության արժանի են համարում սիստեմատիկական աշխատություններում։ Դեռևս ոչ ոք պարզորոշ սահման չի հաստատել անհատական տարբերությունների և աննշան տարատեսակների միջև կամ ցայտուն կերպով արտահայտված տարատեսակների և ենթատեսակների ու տեսակների միջև։ Մի՞թե չկան առանձին մայր ցամաքների վրա կամ միևնույն մայր ցամաքի միմյանցից զանազան անջրպետներով բաժանված տարբեր մասերում և հեռավոր կղզիների վրա բազմաթիվ ձևեր, որոնց փորձված բնախույզներից ոմանք համարում են տարատեսակներ, մյուսները՝ աշխարհագրական ցեղեր կամ ենթատեսակներ, իսկ երրորդները ինքնուրույն, թեև միմյանց հետ մերձավոր տեսակներ։
Իսկ եթե կենդանիներն ու բույսերը փոփոխվում են, թեև չափազանց դանդաղորեն ու աննշան չափով, հապա ինչո՞ւ տարատեսակները կամ որևէ չափով օգտակար անհատական տարբերությունները չպետք է պահպանվեն ու կուտակվեն բնական ընտրության ներգործության կամ ամենահարմարվածների գերապրելու շնորհիվ։ Եթե մարդն իր համբերության շնորհիվ ընտրում է իրեն օգտակար փոփոխություններ, հապա ինչո՞ւ կյանքի բարդ և փոփոխվող պայմաններում չեն կարող հաճախակի առաջանալ ու պահպանվել կամ ընտրության ենթարկվել այնպիսի փոփոխություններ, որոնք օգտակար են հենց բնության կենդանի արտադրանքների համար։ Ի՞նչ սահման կարող է դրվել այդ սկզբունքին, որը ներգործում է երկար դարերի ընթացքում և խստապահանջ կերպով ստուգում է յուրաքանչյուր էակի ընդհանուր կենցաղաձևը, կառուցվածքը և սովորությունները, նպաստելով ամեն մի օգտակար բանի և դեն շպրտելով վնասակարը։ Ես ոչ մի սահման չեմ տեսնում այդ սկզբունքի գործունեության համար, որը դանդաղ ու հիանալի կերպով հարմարեցնում է յուրաքանչյուր ձև ամենաբարդ կենսապայմաններին։ Բնական ընտրության տեսությունը, եթե նույնիսկ մենք սահմանափակվեինք այդ կշռադատություններով, վերին աստիճանի հավանական է թվում։ Ես ամենայն հնարավոր բարեխղճությամբ տվեցի այս տեսության դեմ արված առարկությունների և դժվարությունների հանրագումարը։ Այժմ դառնանք այն հատուկ փաստերին և ապացույցներին, որոնք խոսում են հօգուտ այս տեսության։
Դիտելով տեսակները միայն որպես ցայտուն կերպով նշագծված և կայուն տարատեսակներ և համարելով, որ յուրաքանչյուր տեսակ մի ժամանակ եղել է տարատեսակ, մենք կարող ենք ըմբռնել, թե ինչու խիստ սահմանագծեր չկան տեսակների, որոնք առաջացել են, ինչպես սովորաբար ենթադրում են, հատուկ արարչագործական ակտի շնորհիվ, և տարատեսակների միջև, որոնց ծագումը վերագրվում է երկրորդային օրենքների ներգործությանը։ Այդ տեսակետով մենք կարող ենք մեզ բացատրել, թե ինչու մի որևէ երկրում, որտեղ առաջացել են միևնույն սեռին պատկանող շատ տեսակներ, որոնք շարունակում են բարգավաճել, այդ տեսակներն ունեն շատ տարատեսակներ. այնտեղ, որտեղ տեսակների արտադրումը եղել է շատ գործունյա, մենք կարող ենք սպասել, որպես ընդհանուր կանոն, որ նրան դեռ գործողության մեջ կգտնենք, և իրոք այդ այդպես է, որովհետև տարատեսակները հանդիսանում են սկզբնավորվող տեսակներ։ Բացի դրանից, ամենախոշոր սեռերի տեսակները, որոնք ներկայացնում են մեծ թվով տարատեսակներ կամ սկզբնավորվող տեսակներ, որոշ չափով դեռ պահպանում են տարատեսակների բնույթը, նրանք միմյանցից զանազանվում են ավելի քիչ տարբերություններով, քան փոքր սեռերի տեսակները։ Իրար մոտ կանգնած և միաժամանակ ավելի ընդարձակ սեռերի պատկանող տեսակներն ըստ երևույթին ավելի սահմանափակ տարածում ունեն և իրենց ազգակցության հիման վրա խմբվում են այլ տեսակների շուրջը՝ կազմելով փոքրիկ խմբեր, երկու տեսակետներից էլ նմանվելով տարատեսակներին։ Այդ փոխհարաբերությունները շատ տարօրինակ կլինեին, եթե տեսակներն ստեղծված լինեին մեկը մյուսից անկախ կերպով, բայց նրանք միանգամայն հասկանալի են դառնում, երբ յուրաքանչյուր տեսակ սկզբում գոյություն է ունեցել որպես տարատեսակ։
Քանի որ յուրաքանչյուր տեսակ բազմացման երկրաչափական պրոգրեսիայի շնորհիվ ձգտում է անսահմանորեն ավելացնել իր թիվը և քանի որ յուրաքանչյուր տեսակի փոփոխված սերունդներն այնքան ավելի հեշտությամբ կարող են բազմանալ, որքան ավելի բազմազան են նրանց սովորություններն ու կառուցվածքը, որոնք հնարավորություն են տալիս նրանց գրավելու բնության տնտեսության մեջ ամենաբազմազան վայրեր, ուստի բնական ընտրությունը շարունակ ձգտում է պահպանել ամեն մի տեսակի մեջ այն հաղորդներին, որոնք ամենից ավելի տարբերվում են իրարից։ Հետևաբար, փոփոխությունների երկար շարքի ընթացքում միևնույն տեսակին պատկանող տարատեսակները բնորոշող թույլ տարբերությունները ձգտում են աճել ու դառնալ ցայտուն տարբերություններ, որոնք հատուկ են միևնույն սեռին պատկանող տեսակներին։ Նոր և կատարելագործված տարատեսակները անշեղորեն փոխարինելու և ոչնչացնելու են հին, ավելի նվազ կատարյալ և միջակա տարատեսակներին. և այսպիսով տեսակները դառնում են և միմյանցից անջատված օբյեկտներ։ Յուրաքանչյուր դասի սահմաններում ավելի խոշոր խմբերի պատկանող տիրապետող տեսակները ձգտում են առաջացնել նոր և տիրապետող ձևեր․ այսպիսով ամեն մի մեծ խումբ կձգտի ավելի ևս մեծանալ և միևնույն ժամանակ հատկանիշներով ավելի ևս հեռանալ միմյանցից։ Սակայն բոլոր խմբերն այս ձևով չեն կարող աճել, քանի որ ամբողջ աշխարհը չէր տեղավորի դրանց, որի հետևանքով տիրապետող խմբերը կհաղթեն ավելի քիչ տիրապետող խմբերին։ Մեծ խմբերի աճելու և հատկանիշներով հեռանալու այդ ձգտումը, որը կապված է անխուսափելի մահաջնջման հետ, բացատրում է, թե ինչու բոլոր կենդանի էակները դասավորվում են խմբերի մեջ, որոնք ստորադասվելով այլ խմբերի՝ պարփակվում են բոլոր ժամանակներում տիրապետող դարձած սակավաթիվ դասերի սահմաններում։ Բոլոր օրգանական էակների այսպես կոչված բնական սիստեմի մեջ խմբավորման այս մեծ փաստը անբացատրելի է առանձին արարչագործական ակտերի տեսության տեսակետից։
Քանի որ բնական ընտրությունը ներգործում է բացառապես աննշան, հաջորդական, բարենպաստ փոփոխությունների կուտակման միջոցով, ուստի նա չի կարող առաջացնել մեծ ու անակնկալ ձևափոխություններ, նա առաջ է շարժվում միայն փոքր ու դանդաղ քայլերով։ Դրա համար էլ «Natura non facit saltum» կանոնը, որը մեր գիտելիքների ընդարձակման հետ զուգընթաց ավելի ու ավելի է հաստատվում, այս տեսության հիման վրա հասկանալի է դառնում։ Մենք նաև ըմբռնում ենք այն, թե ինչու բնության մեջ ամենուրեք միևնույն նպատակը ձեռք է բերվում բազմազան ճանապարհներով, որովհետև յուրաքանչյուր առանձնահատկություն, որ մի անգամ ձեռք է բերվել, երկար ժամանակ ժառանգվում է, և շատ տարբեր ուղղություններով փոփոխված կազմվածքները պետք է հարմարվեն մի ընդհանուր նպատակի։ Մի խոսքով, մենք կարող ենք տեսնել, թե ինչու բնությունը բազմազանության նկատմամբ շռայլ է, թեև ժլատ է նորամուծությունների նկատմամբ։ Բայց թե ինչու պիտի գոյություն ունենար բնության այդպիսի օրենքը, եթե տեսակներն ստեղծվել են մեկը մյուսից անկախ կերպով, ոչ ոք չի կարող բացատրել։
Էլի շատ այլ փաստեր, ինչպես ինձ թվում է, բացատրվում են այս տեսությամբ։ Որքա՜ն տարօրինակ է այն փաստը, որ փայտփորի ընդհանուր կերպարանքով թռչունը գետնի վրա միջատներ է որսում, որ լեռնային սագը, որը հազվագյուտ դեպքում է լողում կամ երբեք չի լողում, մատների միջև ունի թաղանթներ. որ կեռնեխի նման մի թռչուն սուզվում է ու սնվում ջրային միջատներով և որ մրրկահավն ունի այնպիսի սովորություններ ու կառուցվածք, որոնցով հարմարվել է պինգվինի կենցաղին։ Եվ այսպես անթիվ այլ դեպքեր։ Սակայն ըստ այն տեսակետի, որ յուրաքանչյուր տեսակ անընդհատ ձգտում է ավելացնել իր թիվը և բնական ընտրությունը մշտապես գործում է, միշտ պատրաստ լինելով հարմարեցնել նրանցից յուրաքանչյուրի դանդաղ փոփոխվող հաջորդներին բնության մեջ դեռ չգրավված կամ վատ գրավված ամեն մի վայրի, այդ փաստերը դադարում են տարօրինակ լինելուց և կարող են նույնիսկ կանխագուշակվել։
Մենք կարող ենք որոշ չափով հասկանալ, թե ինչու բնության մեջ այդքան շատ գեղեցկություն կա, որովհետև այդ ևս մասամբ կարող է վերագրվել ընտրության գործունեությանը։ Որ գեղեցկությունը, նրա մասին մեր ունեցած հասկացողության համաձայն, համաընդհանրական երևույթ չի ներկայացնում, կընդունի ամեն ոք, ով մի հայացք կգցի մի քանի թունավոր օձերի, մի քանի ձկների և մի քանի այլանդակ չղջիկների վրա, որոնք ներկայացնում են մարդկային կերպարանքի աղավաղված նմանությունը։ Սեռական ընտրությունը բազմաթիվ թռչունների, թիթեռների և այլ կենդանիների արուներին, իսկ երբեմն էլ երկու սեռերին հաղորդել է ամենափայլուն գույներ, ամենանուրբ ձևեր և այլ զարդարանքներ։ Նա թռչունների արուներին տվել է այնպիսի ձայն, որը երաժշտական է էգերի համար, ինչպես նաև մեր լսողության համար։ Ծաղիկներն ու պտուղները դարձել են տարբերվող իրենց վառ գույնով, որով նրանք աչքի են ընկնում սաղարթի կանաչ գույնի միջից, որպեսզի միջատները հեշտությամբ նկատեն ծաղիկները, այցելեն նրանց ու փոշոտեն, իսկ սերմերը տարածվեն թռչունների միջոցով։ Թե ինչպես է պատահել, որ շատ գույներ, ձայներ և ձևեր հաճույք են պատճառում թե՛ մարդուն և թե՛ ստորակարգ կենդանիներին, այլ խոսքով ասած՝ ինչպե՞ս է առաջացել գեղեցկության զգացմունքն իր ամենապարզ ձևով։ Մենք այդ չգիտենք, ինչպես չգիտենք նաև այն, թե ինչու որոշ հոտեր և համեր հաճելի են դարձել։
Քանի որ բնական ընտրությունը ներգործում է մրցության միջոցով, ուստի նա հարմարեցնում և կատարելագործում է յուրաքանչյուր երկրի բնակիչների միայն նրանց հետ համակեցության մեջ գտնվող բնակիչների նկատմամբ. մենք առիթ չունենք զարմանալու այն փաստի վրա, որ որևէ տվյալ երկրի տեսակները, որոնք թեև սովորական տեսակետի համաձայն ստեղծված ու հատկապես հարմարված են այդ երկրի համար, երբեմն հաղթվում են ու փոխարինվում այլ երկրներից բերված նատուրալիզացիայի ենթարկված էակների կողմից։ Մենք չպետք է զարմանանք նաև այն բանի վրա, որ բնության մեջ հարմարեցումները, որքան մենք կարող ենք դատել, բացարձակապես կատարյալ չեն, ինչպես այդ նկատվում է նույնիսկ մարդկային աչքի վերաբերմամբ, որ նրանցից մի քանիսը չեն համապատասխանում հարմարեցման մասին մեր ունեցած պատկերացմանը։ Մենք չպետք է զարմանանք նաև, որ մեղվի խայթոցը, որն օգտագործվում է թշնամու դեմ, մեղվին իրեն մահ է պատճառում. որ բոռերն արտադրվում են այդպիսի մեծ թվով մի գործողության համար, իսկ հետո կոտորվում են իրենց անպտուղ քույրերի կողմից. որ զարմանալի քանակությամբ ծաղկափոշի է ծախսվում, որը նկատվում է սոճիի մոտ. չպետք է զարմանանք այն բնազդային ատելության վրա, որ մայր մեղուն տածում է դեպի իր սեփական պտղաբեր դուստրերին. այն բանի վրա, որ իխնեվմոնիդները կերակրվում են թրթուրի կենդանի մարմնի ներսում, կամ այլ նման դեպքերի վրա։ Բնական ընտրության տեսության հիման վրա ավելի շատ զարմանալի է թվում այն, որ բացարձակ կատարելության բացակայության նման դեպքեր է՛լ ավելի մեծ թվով չեն հայտնաբերված։
Տարատեսակների առաջացումը ղեկավարող բարդ և մեզ քիչ հայտնի օրենքները, որքան մենք կարող ենք դատել, նույնն են, ինչ որ ղեկավարել են նաև առանձին տեսակների առաջացումը։ Երկու դեպքում էլ ֆիզիկական պայմանները, ըստ երևույթին, ունեցել են ուղղակի և որոշակի ներգործություն, բայց թե որքան մեծ է այդ ներգործությունը, մենք չենք կարող ասել։ Այսպես, երբ տարատեսակները տեղափոխվում են մի նոր բնակավայր, նրանք երբեմն ստանում են այդ վայրի տեսակներին հատուկ հատկանիշներ։ Ինչպես՝ տարատեսակների, այնպես էլ տեսակների վերաբերմամբ վարժեցման առկայությունը կամ բացակայությունը, ըստ երևույթին, ունեցել են զգալի ներգործություն․ այդ եզրակացությունից անհնարին է հրաժարվել, տեսնելով, օրինակ, էյտոնովյան բադին նրա այնպիսի թևերով, որոնք թռչելու համար գրեթե նույնքան անպետք են, ինչքան որ մեր ընտանի բադինը, կամ հողի մեջ թաղվող տուկու-տուկոն, որը երբեմն կույր է լինում, և մի քանի խլուրդներ, որոնք սովորաբար կույր են և ունեն մաշկով ծածկված աչքեր. կամ վերջապես տեսնելով այն կույր կենդանիները, որոնք ապրում են ամերիկական ու եվրոպական ստորերկրյա քարանձավներում։ Ինչպես տարատեսակների, այնպես էլ տեսակների վերաբերմամբ, ըստ երևույթին, էական դեր է կատարել հարաբերակցական փոփոխությունը, այնպես որ, երբ փոփոխվել է մի որևէ մաս, անհրաժեշտորեն փոփոխվել են նաև մյուսները։ Թե՛ տարատեսակները և թե՛ տեսակները երբեմն երևան են բերում վերադարձ դեպի վաղուց կորցրած հատկանիշները։ Առանձին արարչագործսկան ակտերի տեսակետից ինչքա՜ն անհասկանալի է ձիու սեռի զանազան տեսակների և նրանց հիբրիդների ուսերի և ոտների վրա զոլերի պատահական երևան գալը։ Եվ ինչքան պարզ կերպով է բացատրվում այդ փաստը, եթե մենք ընդունենք, որ այդ բոլոր տեսակները առաջացել են մի զոլավոր նախահորից, ճիշտ այնպես, ինչպես աղավնու ընտանի տարբեր ցեղերը առաջացել են լայնակի զոլեր ունեցող կապտագորշ ժայռաղավնուց։
Տեսակների անկախ ստեղծագործության մասին տարածված տեսակետի հիման վրա, ինչպե՞ս կարելի է բացատրել, թե ինչու տեսակային հատկանիշները կամ այն հատկանիշները, որոնցով միմյանցից տարբերվում են միևնույն սեռին պատկանող տեսակները, ավելի փոփոխական են, քան սեռային հատկանիշները, որոնցով նրանք բոլորն էլ իրար նման են։ Ինչո՞ւ, օրինակ, ծաղկի գույնի փոփոխականությունն ավելի հավանական է այն տեսակի մոտ, որը պատկանում է այնպիսի սեռի, որի տեսակներն ունեն զանազանագույն ծաղիկներ, քան այն տեսակի մոտ, որը պատկանում է միանման ծաղիկներով տեսակներ ունեցող մի սեռի։ Եթե տեսակները միայն լավ արտահայտված տարատեսակներ են, որոնց հատկանիշները ավելի կայունացած են, ապա մենք կարող ենք բացատրել այդ փաստը, որ նրանք արդեն փոփոխվել են այն պահից սկսած, երբ նրանք ճյուղավորվել են իրենց ընդհանուր նախահորից և փոփոխվել են հատկանիշներով, որոնք կազմում են նրանց տեսակային տարբերությունը, դրա համար էլ հատկապես այդ հատկանիշները պետք է ավելի փոփոխական լինեն, քան սեռային հատկանիշները, որոնք մշտապես հսկայական ժամանակաշրջանի ընթացքում ժառանգաբար փոխանցվել են։ Առանձին արարչագործության տեսության հիման վրա անբացատրելի է նաև այն, թե ինչու այն մասը, որն արտասովոր զարգացման է հասել տվյալ սեռի մի որևէ տեսակի մոտ, ուստի և, ինչպես որ մենք բնականաբար կարող ենք եզրակացնել, չափազանց կարևոր է տվյալ տեսակի համար, պիտի առանձնապես փոփոխական լիներ. բայց մեր տեսակետից այդ մասն արդեն զգալի աստիճանի փոփոխականության է ենթարկվել այն ժամանակից սկսած, երբ տարբեր տեսակները սկսել են ճյուղավորվել մի ընդհանուր նախահորից, այդ պատճառով էլ մենք կարող ենք սպասել, որ այդ մասը մինչև այժմ ընդհանրապես պահպանել է իր փոփոխվելու հակումը։ Սակայն մասը կարող է ամենաարտասովոր կերպով զարգացած լինել, ինչպես, օրինակ, չղջիկի թևը, և այնուամենայնիվ ավելի մեծ փոփոխականություն չցուցաբերել, քան որևէ այլ օրգան, երբ այդ մասն ընդհանուր է լինում իրար ստորադասված ձևերի ամբողջ խմբի համար, այսինքն այն դեպքում, երբ նա ժառանգաբար փոխանցվել է երկար ժամանակաշրջանի ընթացքում, որովհետև այդ դեպքում բնական ընտրության երկարատև ներգործության հետևանքով նա արդեն դարձել է կայուն։
Դառնում եմ բնազդներին․ ինչքան էլ զարմանալի են նրանցից մի քանիսը, բայց նրանք մարմնի կառուցվածքից ավելի մեծ դժվարություններ չեն ներկայացնում հաջորդական, աննշան, բայց օգտակար փոփոխությունների բնական ընտրության տեսության համար։ Այսպիսով, մենք կարող ենք հասկանալ, թե ինչու բնությունը միևնույն դասի տարբեր կենդանիների օժտելով նրանց հատուկ բնազդներով, առաջ է գնում աստիճանական քայլերով։ Ես աշխատել եմ ցույց տալ, թե աստիճանականության սկզբունքը ինչքան լույս է սփռում սովորական մեղվի զարմանալի ճարտարապետական ընդունակությունների վրա։ Սովորությունը, անկասկած, մասնակցում է բնազդների փոփոխությանը, բայց հավանորեն այդ անհրաժեշտ չէ, ինչպես այդ տեսնում ենք անսեռ միջատների օրինակի վրա, որոնք իրենցից հետո սերունդ չեն թողնում, որը կկարողանար ժառանգել երկարատև սովորությունների հետևանքները։ Ընդունելով, որ միևնույն սեռի բոլոր տեսակներն առաջացել են մի ընդհանուր նախահորից և ժառանգել են ընդհանուր շատ առանձնահատկություններ, մենք կարող ենք հասկանալ, թե ինչպես նման տեսակները, գտնվելով ամենատարբեր կենսապայմաններում, այնուամենայնիվ ղեկավարվում են գրեթե միևնույն բնազդներով․ թե ինչու, օրինակ, Հարավային Ամերիկայի արևադարձային և բարեխառն մասերում կեռնեխներն իրենց բները ծածկում են տիղմով, այնպես, ինչպես որ մեր բրիտանական տեսակները։ Բնական ընտրության միջոցով դանդաղորեն բնազդներ ձեռք բերելու տեսակետից մեզ զարմանալի չի թվում, եթե որոշ բնազդներ անկատար են և առիթ են տալիս սխալների, ինչպես նաև այն, որ շատ բնազդներ տանջանք են պատճառում այլ կենդանիների։
Եթե տեսակները միայն լավ արտահայտված ու կայուն տարատեսակներ են, ապա մեզ համար իսկույն պարզվում է, թե ինչու նրանց խաչաձևումից ստացված արդյունքներն իրենց ծնողների հետ ունեցած նմանության հաջորդականությամբ ենթարկվում են նույն բարդ օրենքներին, ինչ օրենքների որ ենթարկվում են ակնհայտ տարատեսակների խաչաձևման արդյունքները, այսինքն հաջորդական խաչաձևումների ժամանակ աստիճանաբար մեկը կլանում է մյուսին։ Այդ նմանությունը տարօրինակ կթվար, եթե տեսակներն իրենց ծագմամբ պարտական լինեին անկախ արարչագործական ակտերին, իսկ տարատեսակները առաջացած լինեին երկրորդային օրենքների ուժով։
Եթե մենք ընդունենք, որ երկրաբանական տարեգրությունը վերին աստիճանի անկատար է, այդ դեպքում նրա մատակարարած փաստերը ուժեղ հաստատություն են հանդիսանում հաջորդական փոփոխություններով ծագման միասնականության տեսության համար։ Նոր տեսակները հրապարակ են եկել դանդաղորեն և իրար հաջորդող ժամանակամիջոցներից հետո, ըստ որում հավասար ժամանակամիջոցների ընթացքում փոփոխականության չափը զանազան խմբերի համար խիստ տարբեր է եղել։ Տեսակների և նրանց ամբողջ խմբերի մահաջնջումը, որն այնպիսի աչքի ընկնող դեր է կատարել օրգանական աշխարհի պատմության մեջ, բնական ընտրության գրեթե անխուսափելի հետևանքն է, որովհետև հին ձևերը փոխարինվում են նոր և կատարելագործված ձևերով։ Ոչ առանձին տեսակներ, ոչ էլ տեսակների մի խումբ նորից երևան չեն գալիս, եթե բնական գոյացման շղթան կտրված է։ Տիրապետող ձևերի աստիճանական տարածումը, նրանց հաջորդների դանդաղ փոփոխության հետ միասին, այնպիսի տպավորություն է թողնում, որ կարծես թե օրգանական ձևերը մեծ ժամանակաշրջանների ընթացքում միաժամանակ են փոփոխվել ամբողջ երկրագնդի վրա։ Այն փաստը, որ յուրաքանչյուր ֆորմացիայի բրածո մնացորդները հատկանիշներով որոշ չափով միջակա են նրանից անմիջապես վերև և անմիջապես ներքև գտնվող ֆորմացիաների բրածոների միջև, շատ պարզ կերպով բացատրում է նրանց ունեցած միջակա դրությամբ ծննդաբանական շղթայի մեջ։ Այն հիմնական փաստը, որ բոլոր անհետացած էակները կարող են դասակարգվել այժմ ապրողների հետ միասին մի ընղհանուր սիստեմի մեջ, բնականաբար բխում է նրանից, որ թե՛ ապրող և թե՛ անհետացած էակներն առաջացել են ընդհանուր նախնիներից։ Քանի որ տեսակներն իրենց փոփոխության երկարատև ժամանակաշրջանի ընթացքում սովորաբար տարամիտվել են իրենց հատկանիշներով, ապա հասկանալի է դառնում, թե ինչու ամեն մի խմբի ամենահին ձևերը կամ ամենահին նախածնողները հաճախ գրավում են որոշ չափով միջակա դրության գոյություն ունեցող խմբերի միջև։ Այժմյան ձևերը սովորաբար համարվում են կազմվածքի ավելի բարձր աստիճանի վրա կանգնած, քան հին ձևերը. նրանք իսկապես էլ պետք է ավելի բարձր դիրք գրավեն, քանի որ հետագա և կատարելագործված ձևերը գոյության կռվում հաղթել են հին և ավելի քիչ կատարելագործված ձևերին․ նրանց օրգանները նույնպես ավելի մեծ չափով են մասնագիտացել զանազան ֆունկցիաների համար։ Այս փաստը միանգամայն համատեղվում է այն բազմաթիվ օրգանիզմների գոյության հետ, որոնք ունեն ավելի պարզ, քիչ կատարելագործված կառուցվածք, որը միանգամայն պիտանի է իրենց գոյության պարզ պայմանների համար․ նա միանգամայն համատեղվում է այն երևույթի հետ, որ մի քանի ձևեր իրենց կազմվածքով հետադիմել են, շարունակելով այդ հետադարձի ճանապարհի յուրաքանչյուր աստիճանի վրա ավելի լավ հարմարված լինել նոր և ցածրացած կենսապայմաններին։ Վերջապես, մի մայր ցամաքի վրա ազգակից ձևերի երկարատև պահպանման զարմանալի օրենքը,— պարկավորներն Ավստրալիայում, թերատամները Ամերիկայում և այլ նման դեպքեր,— միանգամայն հասկանալի է, քանի որ միևնույն երկրի սահմաններում գոյություն ունեցող և անհետացած օրգանիզմները իրենց ընդհանուր ծագմամբ սերտորեն կապված են իրար հետ։
Դառնում եմ աշխարհագրական տեղաբաշխմանը. եթե մենք ընդունենք, որ դարերի երկար շարքի ընթացքում, շնորհիվ նախկինում տեղի ունեցած կլիմայական և աշխարհագրական փոփոխությունների և բազմաթիվ պատահական ու մեզ անհայտ տարածման եղանակների, տեղի է ունեցել օրգանիզմների տարաբնակումը աշխարհի մի մասից մյուսը, այն ժամանակ մենք ի վիճակի կլինենք փոփոխությամբ ուղեկցվող ընդհանուր ծագման տեսության հիման վրա հասկանալու նաև այն հիմնական փաստերի մեծամասնությունը, որոնք վերաբերում են օրգանիզմների տեղաբաշխմանը։ Մենք կարող ենք բացատրել նաև այն զարմանալի զուգահեռությունը, որ գոյություն ունի տարածության մեջ օրգանական էակների տեղաբաշխման և ժամանակի մեջ նրանց երկրաբանական հաջորդականության միջև, որովհետև երկու դեպքում էլ այդ էակները միանման կերպով կապված են եղել միմյանց հետ ազգակից ծագմամբ և փոփոխության եղանակները եղել են նույնը։ Մեզ համար պարզ է այն զարմանալի փաստի լիակատար իմաստը, որ ապշեցրել է ամեն մի ճանապարհորդի, այն է՝ որ միևնույն մայր ցամաքի վրա ամենատարբեր պայմաններում, թե՛ տաք և ցուրտ կլիմայում, թե՛ լեռներում և թե՛ հարթավայրերում, թե՛ տափաստաններում և թե՛ ճահիճներում, միևնույն ղասին պատկանող բնակիչների մեծ մասը հանդես է բերում բացահայտ ազգակցական գծեր, որովհետև նրանք միևնույն նախահայրերի և հին գաղթականների սերունդներն են։ Առաջներում տեղի ունեցած և շատ դեպքում փոփոխությամբ ուղեկցվող տարաբնակման սկզբունքի հիման վրա՝ մենք կարողանում ենք բացատրել սառցադաշտային ժամանակաշրջանի միջոցով մի քանի սակավաթիվ բույսերի նույնությունը և ուրիշ շատ բույսերի մոտ ազգակցությունը, որոնք ապրում են իրարից շատ հեռու ընկած լեռների վրա և հյուսիսային ու հարավային բարեխառն գոտիներում. այդպես էլ մենք բացատրում ենք նաև հյուսիսային և հարավային բարեխառն գոտու ծովերի մի քանի բնակիչների մերձավոր ազգակցությունը, չնայած նրան, որ նրանք անջատված են միմյանցից օվկիանոսի այն ամբողջ մասով, որն ընկած է արևադարձների միջև։ Թեկուզ երկու երկրների ֆիզիկական պայմաններն այնքան նման լինեին իրար, որքան այդ կարող է պահանջել միևնույն տեսակը, ապա մենք զարմանալու առիթ չէինք ունենա, որ նրանց բնակիչները խիստ տարբերվում են միմյանցից, եթե միայն այդ երկրները երկարատև ժամանակաշրջանի ընթացքում լրիվ անջատված են եղել իրարից։ Քանի որ օրգանիզմների միջև եղած փոխհարաբերությունները բոլոր հարաբերություններից ամենակարևորն են և քանի որ երկու երկրներն էլ իրենց գաղթականները պիտի ստանային մի որևէ այլ երկրից կամ մեկը մյուսից տարբեր ժամանակներում, տարբեր հարաբերությամբ, ուստի և փոփոխության ուղղությունը երկու մարզերում անխուսափելիորեն պետք է տարբեր լիներ։
Գաղթի և նրան հետևող փոփոխության երևույթների նկատմամբ այս տեսակետից մենք կարող ենք բացատրել, թե ինչու օվկիանոսային կղզիները բնակված են միայն սակավաթիվ տեսակներով և ինչու այդ տեսակների մեծ մասը հատուկ է տվյալ վայրին, կամ էնդեմիկ ձևեր են։ Մեզ համար պարզ է, թե ինչու կենդանիների այն խմբերին պատկանող տեսակները, որոնք չեն կարողանում տեղափոխվել օվկիանոսի մեծ տարածությունների վրայով, ինչպիսիք են գորտերը և ցամաքային կաթնասունները, չեն հանդիպում օվկիանոսային կղզիների վրա. մյուս կողմից, թե ինչու չղջիկների նոր և բացառիկ տեսակները,— այսինքն կենդանիներ, որոնք կարող են թռչել օվկիանոսի վրայով,— հաճախ են հանդիպում որևէ մայր ցամաքից հեռու գտնվող կղզիների վրա։ Այդպիսի դեպքերը, ինչպես, օրինակ, օվկիանոսային կղզիների վրա չղջիկների բնորոշ տեսակների առկայությունը և ուրիշ ցամաքային կաթնասունների լիակատար բացակայությունը, փաստեր են, որոնք բոլորովին անբացատրելի են առանձին արարչագործական ակտերի տեսության տեսակետից։
Որևէ երկու մարզերում մոտ ազգակից կամ փոխարինող տեսակների գոյությանը փոփոխությամբ ուղեկցվող ընդհանուր ծագման տեսության հիման վրա ենթադրում է, որ նույն ծնողական ձևերը առաջները գոյություն են ունեցել երկու մարզերում. և մենք իրոք գրեթե միշտ տեսնում ենք, որ երբ բազմաթիվ ազգակից տեսակներ բնակվում են երկու մարզերում, ապա այդ տեսակների միջև գտնվում են այնպիսիները, որոնք նույնն են և տարածված են երկու մարզերում։ Այնտեղ, որտեղ հանդիպում են բազմաթիվ, միմյանց հետ մոտ, բայց և այնպես պարզորոշ տարբերվող տեսակներ, հանդիպում են նաև կասկածելի ձևեր և տարատեսակներ, որոնք պատկանում են նույն խմբերին։ Կարելի է որպես շատ լայն տարածված կանոն ընդունել այն, որ որևէ մարզի բնակչությունը ազգակցության մեջ է գտնվում այն ամենամոտ աղբյուրի բնակչության հետ, որտեղից կարող էր տեղի ունենալ ներգաղթը։ Այդ աչքի է ընկնում Գալապագոսյան արշիպելագի, Ժուան Ֆերնանդես կղզիների և ամերիկական այլ կղզիների գրեթե բոլոր բույսերի ու կենդանիների և հարևան ամերիկական մայր ցամաքի բույսերի ու կենդանիների միջև եղած ազգակցության վերաբերմամբ։ Նույնն է նկատվում նաև Կանաչ հրվանդանի արշիպելագի և այլ աֆրիկական կղզիների և աֆրիկական մայր ցամաքի վերաբերմամբ։ Պետք է ընդունել, որ այդ փաստերը առանձին արարչագործությունների տեսության հիման վրա ոչ մի բացատրություն չեն ստանում։
Այն փաստը, որ բոլոր գոյություն ունեցող և երբևիցե գոյություն ունեցած օրգանիզմները կարող են դասավորվել մի քանի մեծ դասերի սահմաններում եղած խմբերի մեջ, որոնք ստորադասվում են այլ խմբերի, ըստ որում անհետացած խմբերը հաճախ գրավում են միջակա դրություն գոյություն ունեցող խմբերի միջև, միանգամայն հասկանալի է բնական ընտրության տեսության և նրա անհրաժեշտ հետևանքների՝ մահաջնջման ու հատկանիշների տարամիտման հիման վրա։ Նույն սկզբունքների հիման վրա մեզ համար պարզ է, թե ինչու ձևերի փոխադարձ ազգակցությունը այդքան բարդ է և զարտուղի։ Մենք տեսնում ենք, թե ինչու որոշ հատկանիշներ դասակարգության նպատակների համար ավելի գերադասելի են, քան մյուսները․ թե ինչու հարմարեցման հետ կապված հատկանիշները, որ վերին աստիճանի կարևոր են դրանցով օժտված էակների համար, գրեթե ոչ մի նշանակություն ունեն դասակարգության գործում․ թե ինչու թերաճ մասերից առաջացած հատկանիշները, որոնք թեև անօգուտ են այդ մասերն ունեցող էակները համար, երբեմն այնքան արժեքավոր են դասակարգության մեջ և վերջապես, թե ինչու սաղմնաբանական հատկանիշները հաճախ ամենից ավելի արժեքավոր են։ Օրգանական էակների իսկական ազգակցությունը, ի տարբերություն նրանց այն նմանությունների, որոնք առաջանում են միանման հարմարեցումների հետևանքով, իր առաջացմամբ պարտական է ժառանգականությանը և ծագման միասնականությանը։ Բնական սիստեմը ոչ այլ ինչ է, եթե ոչ էակների ծննդաբանական դասավորումը, ըստ որում նրանց ձեռք բերած տարբերության աստիճանները որոշվում են տարատեսակներ, տեսակներ, սեռեր, ընտանիքներ և այլ տերմիններով։ Մեր խնդիրն է հայտնաբերել այդ ծննդաբանական կապերը ամենակայուն հատկանիշների միջոցով, ինչպիսին էլ որ լինեն այդ հատկանիշները և ինչքան էլ ուզում է աննշան լինի նրանց նշանակությունը այդ էակների կյանքի համար։
Մարդու ձեռքի, չղջիկի թևի, դելֆինի լողաթևի, ձիու ոտքի մեջ ոսկրների նման դասավորությունը, ընձուղտի և փղի պարանոցը կազմող ողերի միանման թիվը և այլ նման անթիվ փաստերը մեզ համար իսկույն հասկանալի են դառնում դանդաղ ու աստիճանական հաջորդական փոփոխությամբ ուղեկցվող ընդհանուր ծագման տեսության հիման վրա։ Չղջիկի թևի և ոտքի հիմնական կառուցվածքում եղած նմանությունը, թեև նրանք բոլորովին տարբեր դեր ունեն, խաչափառի ծնոտների ու ոտքերի, ծաղկի պսակաթերթերի, առէջքների և վարսանդների կառուցվածքում եղած նմանությունները նույնպես մեծ չափով հասկանալի են՝ համաձայն այն տեսակետի, որ մասերը կամ օրգանները ենթարկվել են աստիճանական ձևափոխման և նրանք սկզբում իրար նման են եղել այդ դասերից յուրաքանչյուրի հեռավոր նախահոր մոտ։ Այն կանոնի հիման վրա, որ իրար հաջորդող փոփոխությունները ոչ միշտ երևան են գալիս վաղ հասակում և ժառանգվում են կյանքի համապատասխան ոչ վաղ շրջանում, մենք կարող ենք բացատրել, թե ինչու կաթնասունների, թռչունների, սողունների և ձկների սաղմերը այդքան զարմանալիորեն նման են իրար, մինչդեռ չափահաս ձևերը այնքան տարբեր են։ Մեզ այլևս չի զարմացնի այն փաստը, որ օդով շնչող կաթնասունների կամ թռչունների սաղմերն ունեն խռիկների ճեղքեր և հանգույցներով դասավորված զարկերակներ, ինչպես ձկների մոտ է, որոնք իրենց միանգամայն զարգացած խռիկներով շնչում են ջրի մեջ լուծված օդով։
Վարժության բացակայությունը երբեմն բնական ընտրության աջակցությամբ հաճախ օժանդակել է այն օրգանների այլասերմանը, որոնք կենցաղի և կենսապայմանների փոփոխության դեպքում անօգուտ են դարձել. այստեղից մեզ համար հասկանալի է դառնում թերաճ օրգանների իմաստը։ Բայց վարժության բացակայությունը և ընտրությունը սովորաբար ներգործում են այն էակների վրա, որոնք հասել են լիակատար հասունության և մասնակցում են գոյության կռվին, և այսպիսով քիչ ազդեցություն են թողնում օրգանի վրա վաղ հասակում. այստեղից բխում է, որ օրգանը ռեդուկցիայի չի ենթարկվում կամ թերաճ չի դառնում վաղ հասակում։ Այսպես, օրինակ, հորթը ժառանգել է ատամներ, որոնք երբեք դուրս չեն գալիս վերին ծնոտի լնդերքի միջից, նա այդ ատամները ժառանգել է իր վաղ նախահորից, որն ունեցել է միանգամայն զարգացած ատամներ։ Մենք կարող ենք ենթադրել, որ չափահաս կենդանու ատամները մի ժամանակ ռեդուկցիայի են ենթարկվել վարժեցման բացակայության հետևանքով, որի պատճառը եղել է այն, որ լեզուն, քիմքը կամ շրթունքները բնական ընտրության շնորհիվ հիանալի կերպով հարմարվել են առանց ատամների մասնակցության խոտը պոկռտելու համար։ Մինչդեռ հորթի ատամները մնացել են անփոփոխ և համապատասխան հասակում ժառանգման օրենքի հիման վրա, ժառանգվել են հեռավոր ժամանակներից սկսած մինչև այսօր։ Ըստ այն տեսակետի, որ յուրաքանչյուր օրգանիզմ իր բոլոր մասերով միասին առանձին է ստեղծվել, վճռականապես անհասկանալի է, որ այդքան հաճախ կարող են հանդիպել այն օրգանների գոյության փաստերը, որոնց անօգտակարությունը այնպես ակնհայտ է, ինչպես հորթի սաղմերի մոտ ատամները կամ շատ բզեզների միաձուլված վերնաթևերի տակ գտնվող ճխլտված թևերը։ Բնությունը կարծես դիտմամբ հոգացել է այն մասին, որպեսզի թերաճ օրգանների, սաղմնաբանական և հոմոլոգիական նմանությունների միջոցով մեր առաջ բաց անի այն պլանը, որով նա ստեղծում է փոփոխություններ, բայց մենք չափազանց կույր ենք, որ ըմբռնենք, թե նա դրանով ինչ է արտահայտում։
Ես համառոտակի կրկնեցի այն կշռադատություններն ու փաստերը, որոնք ինձ միանգամայն համոզել են, որ տեսակները փոփոխվել են սերունդների երկար շարքի ընթացքում։ Այդ ձեռք է բերվել գլխավորապես բազմաթիվ, հաջորդական, աննշան, բարենպաստ փոփոխությունների բնական ընտրության միջոցով, որին էական աջակցություն են ցույց տվել, մի կողմից, օրգանի վարժեցման կամ ոչ վարժեցման ժառանգված հետևանքները և, մյուս կողմից, ոչ էական օժանդակություն են ցույց տվել արտաքին պայմանների անմիջական ներգործությունը թե՛ անցյալի և թե՛ ներկայի հարմարողական հատկանիշների վրա և այն փոփոխությունները, որոնք մեր անտեղյակության պատճառով մեզ թվում են ինքնակամ։ Ըստ երևույթին, առաջները ես անբավարար նշանակություն եմ տվել փոփոխականության այս վերջին ձևերի ուժին ու տարածվածությանը, որոնք բնական ընտրությունից անկախ առաջ են բերում հաստատուն փոփոխություններ կառուցվածքի մեջ։ Բայց քանի որ վերջերս իմ եզրակացությունները հաճախ թյուր կերպով են մեկնաբանել և պնդել են, թե ես տեսակների փոփոխությունը վերագրում եմ բացառապես բնական ընտրությանը, ուստի ես ինձ թույլ կտամ նկատելու, որ այս գրքի առաջին և հետագա հրատարակություններում ես զետեղել եմ շատ աչքի ընկնող տեղում, այն է՝ ներածության վերջում հետևյալ բառերը։ «Ես համոզված եմ, բնական ընտրությունը եղել է փոփոխություններ առաջացնող գլխավոր, բայց ոչ միակ միջոցը»։ Բայց դա չօգնեց։ Ուրիշների մտքերը համառորեն խեղաթյուրելու ուժը մեծ է, սակայն գիտության պատմությունը ցույց է, տալիս, որ բարեբախտաբար այդ ուժի ներգործությունը տևական չէ։
Անկարելի է ընդունել, որ մի սխալ տեսություն կարողանար այնպես գոհացուցիչ կերպով բացատրել, ինչպես այգ բացատրում են բնական ընտրության տեսության փաստերի այն մեծ խմբերը, որոնք հենց նոր հիշատակեցինք։ Վերջերս առարկություն է արվել, որ փաստարկման նման եղանակը վստահելի չէ, բայց այն շարունակ կիրառվում է կյանքում և կիրառվել է մեծագույն բնախույզների կողմից։ Այդպես է ստեղծվել լույսի ալիքաձև շարժման տեսությունը, և այն համոզվածությունը, որ երկիրը պտտվում է իր առանցքի շուրջը, մինչև վերջին ժամանակները գրեթե ոչ մի ուղղակի ապացույցի վրա չէր հենվում։ Այն առարկությունը, թե գիտությունը մինչև այժմ չի լուսաբանել շատ ավելի բարձր խնդիրներ կյանքի էության և սկզբնավորության մասին, նշանակություն չունի։ Ո՞վ հանձն կառնի բացատրելու տիեզերական ձգողականության էությունը։ Ոչ ոք այժմ չի առարկում այն եզրակացությունների դեմ, որոնք բխում են ձգողականության այդ անհայտ սկզբունքից, չնայած նրան, որ Լեյբնիցը[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 259] մի ժամանակ մեղադրել է Նյուտոնին[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 260] այն բանում, որ նա «փիլիսոփայության մեջ խորհրդավոր հատկություններ ու հրաշքներ» է մտցնում։
Ես բավականաչափ հիմք չեմ տեսնում, թե ինչու այս գրքում շարադրվող տեսակետները կարող են վիրավորել որևէ մեկի կրոնական զգացմունքը։ Ապացուցելու համար, թե ինչքան վաղանցիկ է նման տպավորությունը, կարելի է հիշել, որ մեծագույն հայտնագործությունը, որ երբևիցե կատարել է մարդը, այսինքն՝ տիեզերական ձգողականության հայտնագործությունը, հանդիպել է Լեյբնիցի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 259] հարձակումներին՝ որպես «բնական կրոնի, հետևապես և հայտնության հիմքերը ցնցող»։ Մի նշանավոր հեղինակ և միաժամանակ հոգևոր անձ գրել է ինձ. «Ես հետզհետե սովորեցի տեսնել, որ միանման համատեղելի է աստվածության մասին բարձր պատկերացումը ինչպես հավատ դեպի այն, որ նա ստեղծել է մի քանի նախասկզբնական ձևեր, որոնք ընդունակ են եղել ինքնազարգացման միջոցով սկիզբ տալու մյուս անհրաժեշտ ձևերին, այնպես էլ հավատ դեպի այն, որ նա ամեն անգամ նոր ստեղծագործական ակտի կարիք է ունեցել, որպեսզի լցնի բաց տեղերը, որ երևան ես եկել իր հաստատած օրենքների ներգործության հետևանքով»։
Սակայն գուցե հարցնեն, թե ինչո՞ւ դեռ ապրող բնախույզներից և երկրաբաններից գրեթե բոլոր ամենաականավորները մինչև վերջին ժամանակներս չեն հավատացել տեսակների փոփոխականությանը։ Չի կարելի պնդել, որ օրգանական էակները բնական վիճակում չեն ենթարկվում փոփոխականության. չի կարելի ապացուցել, որ փոփոխությունների գումարը դարերի երկար շարքի ընթացքում ներկայացնում է մի սահմանային մեծություն. ոչ մի պարզորոշ սահման չի առաջարկվել և չի կարող առաջարկվել տեսակների և պարզ արտահայտված տարատեսակների միջև։ Չի կարելի պնդել, որ տեսակներն իրենց խաչաձևման ժամանակ անպայման լինում են անպտղաբեր, իսկ տարատեսակները՝ անպայման պտղաբեր․ կամ պնդել, որ տեսակների անպտղաբերությունը նրանց յուրահատուկ հատկությունն է և անկախ ստեղծման նշանն է։ Տեսակների անշարժության վերաբերմամբ եղած հավատը գրեթե անխուսափելի էր մինչև այն ժամանակ, քանի դեռ գոյություն ուներ այն համոզմունքը, որ երկրի պատմությունը կարճ է։ Բայց այժմ, երբ մենք մոտավոր գաղափար ենք կազմել ժամանակների ընթացքի տևողության մասին, մենք հակված ենք առանց բավարար ապացույցների ընդունելու, որ երկրաբանական տարեգրությունն այնքան կատարյալ է, որ պետք է պահպանած լիներ տեսակների փոփոխության անկասկածելի հետքերը, եթե նրանք ենթարկվել են փոփոխության։
Դրա գլխավոր պատճառը, որ մենք իբնե չենք ցանկանում ընդունել, թե որևէ մի տեսակ կարող է սկիզբ տալ իրեն չնմանվող մի այլ տեսակի, նրանումն է, որ միշտ դժկամությամբ ենք ընդունում մեծ փոփոխությունների գոյությունը, որոնց առանձին ստադիաները մենք ի վիճակի չենք նշմարել։ Այդ դժվարությանը միանգամայն նման է այն դժվարությանը, որ զգացել են երկրաբաններն այն ժամանակ, երբ Լյաելն[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 29] առաջին անգամ սկսել է պնդել, որ մայր ցամաքների վրա եղած երկարաձիգ ժայռոտ լեռնաշարքերը և խոր հովիտները այն գործոնների գործունեության հետևանք են, որոնց մենք այժմ էլ տեսնում ենք գործողության մեջ։ Մեր բանականությունը չի կարող լիովին ըմբռնել նույնիսկ «միլիոն տարի» արտահայտության հետ կապված իմաստը. նա չի կարող գրեթե անհամար սերունդների ընթացքում կուտակված բազմաթիվ թեթև փոփոխությունների հանրագումարը տալ և տեսնել վերջնական արդյունքը։
Թեև ես լիովին համոզված եմ այն բոլոր հայացքների ճշտությանը, որ բերված են այս գրքի մեջ համառոտ ակնարկի ձևով, սակայն ես ոչ մի կերպ հույս չունեմ համոզելու այն փորձված բնախույզներին, որոնց մտքերը լի են մեծաքանակ փաստերով, որոնց նրանք երկար տարիների ընթացքում քննարկում են իմ տեսակետին ուղղակի հակառակ տեսակետից։ Այնքան հեշտ է մեր անտեղյակությունը թաքցնել այնպիսի արտահայտությունների քողի տակ, ինչպիսիք են «արարչագործության պլան», «գաղափարի միասնականություն» և այլն, և երևակայել, որ մենք բացատրություն ենք տալիս, մինչդեռ միայն այլ արտահայտություններով կրկնում ենք բուն փաստը։ Ամեն ոք, ով հակված է ավելի կշիռ տալու չլուծված դժվարություններին, քան որոշ փաստերի բավարար բացատրությանը, անշուշտ կժխտի իմ տեսությունը։ Սակավաթիվ բնախույզների վրա, որոնք օժտված են մտքի զգալի ճկունությամբ և նույնիսկ սկսում են կասկածել տեսակների անշարժությանը, այս գիրքը թերևս ազդեցություն կունենա։ Բայց ես վստահությամբ եմ նայում ապագային, բնախույզների նոր առաջացող երիտասարդ սերնդին, որը ի վիճակի կլինի անաչառ կերպով կշռադատել հարցի երկու կողմերը։ Նա, ով կհամոզվի, որ տեսակները փոփոխական են, լավ ծառայություն մատուցած կլինի, բարեխիղճ կերպով արտահայտելով իր համոզմունքը, միայն այդ ձևով կարելի է տեղից շարժել նախապաշարումների այն զանգվածը, որը ծանրանում է այդ հարցի վրա։
Մի քանի ականավոր բնախույզներ վերջերս մամուլի մեջ հայտնել են այն համոզմունքը, որ յուրաքանչյուր սեռի մեջ ընդունված տեսակներից շատերը իսկական տեսակներ չեն, բայց դրան հակառակ մյուսները իսկական են, այսինքն՝ ստեղծված են մեկը մյուսից անկախ կերպով։ Այդպիսի եզրակացությունը ինձ չափազանց տարօրինակ է թվում։ Նրանք ընդունում են, որ շատ ձևեր, որոնց մինչև վերջին ժամանակը իրենք համարել են առանձին արարչագործական ակտերի արդյունք, իսկ բնախույզների մեծամասնությունը մինչև այժմ էլ դրանց համարում է այդպես, որ այդ ձևերը, որոնք հետևաբար կրում են իսկական տեսակների բոլոր արտաքին հատկանիշները, առաջացել են փոփոխության միջոցով, և միաժամանակ նրանք հրաժարվում են տարածել այդ տեսակետը մյուս ձևերի վրա, որոնք միայն թեթևակի տարբերվում են առաջիններից։ Այնուամենայնիվ, նրանք հանձն չեն առնում տալու այնպիսի սահմանում կամ նույնիսկ առաջարկելու այնպիսի ենթադրություններ, որոնց հիման վրա կարելի կլիներ տարբերել ստեղծված ձևերը նրանցից, որոնք առաջացել են երկրորդային օրենքների ուժով․ նրանց համար փոփոխականությունը մի դեպքում vera causa[77] է, իսկ մյուս դեպքում նրանք կամայականորեն ժխտում են այն, չնշելով, թե ինչումն է այդ երկու դեպքերի տարբերությունը։ Կգա մի օր, երբ դա կհամարեն կանխակալ կարծիքից առաջացող կուրացման մի հետաքրքրական օրինակ։ Այդ հեղինակներին նույնքան քիչ է զարմացնում հրաշքային ստեղծագործական ակտը, որքան և սովորական ծնունդը։ Մի՞թե նրանք իսկապես ենթադրում են, որ մեր մոլորակի պատմության մեջ անթիվ անգամ որոշ տարրական ատոմների հանկարծակի կարգադրվել է կազմակերպվել և վերածվել կենդանի հյուսվածքների։ Արդյո՞ք նրանք կարծում են, որ ամեն մի այդպիսի ենթադրյալ արարչագործական ակտի ժամանակ գոյացել են մեկ, թե շատ անհատներ։ Բոլոր անհամար կենդանիներն ու բույսերը արդյո՞ք ստեղծվել են ձվերի կամ սերմերի, թե՞ չափահաս անհատների ձևով։ Եվ արդյո՞ք կաթնասուններն ստեղծվել են սննդառության կեղծ նշաններով դեռ մոր արգանդում։ Հասկանալի է, որ այդ հարցերից մի քանիսին չեն կարող պատասխան տալ նրանք, ովքեր հավատում են կյանքի քիչ ձևերի կամ միայն մի ձևի երևան գալուն կամ ստեղծմանը։ Մի քանի հեղինակներ առարկել են, թե միլիոնավոր էակների ստեղծմանը նույնքան հեշտ է հավատալ, ինչքան որ մեկի ստեղծմանը, բայց «նվազագույն ներգործության» փիլիսոփայական սկզբունքը, որ արտահայտել է Մոպերտյուին,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 261] ակամայից հակում է մեր միտքը փոքր թվի օգտին, և իհարկե մենք չպետք է հավատանք, որ յուրաքանչյուր մեծ դասի սահմաններում եղած անթիվ էակները ստեղծվել են միասնական ծնողներից ծագման ակներև, բայց խաբուսիկ հատկանիշներով։
Որպես հիշեցում իրերի նախկին դրության մասին ես նախորդ պարագրաֆներում և այլ տեղերում պահպանել եմ մի քանի դիտողություններ, որոնք մատնանշում են այն, որ բնախույզները հավատում են յուրաքանչյուր տեսակի առանձին ստեղծմանը, և ինձ դատապարտել են նրա համար, որ ես արտահայտվել եմ այդպես։ Սակայն անկասկած է, որ այդպես էր ընդհանուր համոզմունքը, երբ լույս տեսավ այս գրքի առաջին հրատարակությունը։ Անցյալում ես առիթ եմ ունեցել շատ բնախույզների հետ զրուցելու էվոլյուցիայի մասին և ոչ մի անգամ չեմ նկատել նրանց կողմից համակրական վերաբերմունք դեպի այդ տեսակետը։ Հնարավոր է, որ նրանցից մի քանիսը դեռ այն ժամանակ էլ հավատում էին էվոլյուցիայի գոյությանը, բայց նրանք կամ լռում էին, կամ այնպես երկդիմի ձևով էին արտահայտվում, որ դժվար էր կռահել նրանց խոսքերի իմաստը։ Այժմ դրությունը վերջնականապես փոխվել է, և այժմ գրեթե ամեն մի բնախույզ ընդունում է էվոլյուցիայի մեծ սկզբունքը։ Սակայն մի քանիսը դեռ շարունակում են կարծել, որ տեսակները միանգամայն անհասկանալի պատճառներով հանկարծակի առաջացրել են նոր և իրենցից բոլորովին տարբեր ձևեր, բայց ինչպես ես աշխատել եմ ցույց տալ, որ այդ հանկարծակի և խիստ ձևափոխություններ ընդունելու դեմ կարելի է ներկայացնել շատ ծանրակշիռ ապացույցներ։ Գիտական տեսակետից և հետագա հետազոտությունների համար որպես նյութ այդ ենթադրությանը, որի համաձայն նոր ձևերը հանկարծակի և անբացատրելի կերպով առաջացել են ավելի հին և նրանցից խիստ տարբերող ձևերից, նույնքան քիչ առավելություններ են ներկայացնում, ինչքան որ այն հին հավատալիքը, որ նոր տեսակներն ստեղծվել են երկրագնդի հողից։
Գուցե կհարցնեն, թե ես ինչքա՞ն հեռու եմ տարածում տեսակների փոփոխության մասին ուսմունքը. այդ հարցին պատասխանելը հեշտ չէ, որովհետև որքան մեծանում է մեր քննարկած ձևերի միջև եղած տարբերության աստիճանը, այնքան նվազում են թե՛ թվով և թե՛ իրենց համոզարարության աստիճանով այն ապացույցները, որոնք վկայում են ընդհանուր ծագման օգտին։ Սակայն մի քանի առանձնապես ծանրակշիռ ապացույցներ կարող են շատ հեռու տարածվել։ Ամբողջ դասերի բոլոր ներկայացուցիչները միմյանց հետ կապված են ազգակցության շղթայով և բոլորը կարող են դասակարգվել միևնույն սկզբունքի հիման վրա հաջորդականորեն մեկը մյուսից ստորադասվող խմբերի մեջ։ Բրածո մնացորդները երբեմն լցնում են գոյություն ունեցող կարգերի միջև եղած լայն տարածությունները։
Թերաճ վիճակում գտնվող օրգանները պարզ կերպով ցույց են տալիս, որ հեռավոր նախահայրն ունեցել է այդ օրգանը միանգամայն զարգացած վիճակում, իսկ այդ շատ դեպքերում հարկադրում է մեզ ենթադրել, որ փոփոխականությունը հաջորդների մեջ հսկայական աստիճանի է հասնում։ Ամբողջ դասերի սահմաններում տարբեր օրգաններ կառուցված են միևնույն օրինակով և վաղ հասակում սաղմերը մոտ նմանություն ունեն միմյանց հետ։ Այդ հիման վրա ես չեմ կասկածում, որ փոփոխությամբ ուղեկցվող ծագման միասնականության տեսությունն ընդգրկում է յուրաքանչյուր ընդարձակ դասի կամ թագավորության բոլոր ներկայացուցիչները. ես կարծում եմ, որ կենդանիները ծագել են ամենաշատը չորս կամ հինգ նախասկզբնական ձևերից, իսկ բույսերը՝ նույնքան կամ ավելի ևս պակաս։
Անալոգիան պետք է ինձ շարժի մի քայլ ևս առաջ, այն է՝ ընդունել, որ բոլոր կենդանիները և բոլոր բույսերը ծագել են մի նախատիպից։ Բայց անալոգիան կարող է երբեմն սխալ ուղեցույց լինել։ Այնուամենայնիվ բոլոր կենդանի էակներն իրենց քիմիական բաղադրությամբ, իրենց բջջային կառուցվածքով, իրենց աճման օրենքներով և վնասակար ազդեցությունների ենթարկվածությամբ շատ նման են միմյանց։ Մենք այդ տեսնում ենք այնպիսի, կարծես թե, աննշան փաստի մեջ, ինչպիսին է միևնույն թույնի միանման ներգործությունը թե՛ բույսերի և թե՛ կենդանիների վրա. կամ ինչպիսին է գխտորաստեղծի թույնի ներգործությունը, որ առաջ է բերում գխտորի այլանդակ գոյացում մասրենու և կաղնու վրա։ Բոլոր օրգանական էակների, թերևս բացառությամբ ստորակարգների, սեռական պրոցեսը էապես միանման է։ Բոլոր օրգանիզմների մոտ, որքան ներկայումս հայտնի է, սաղմնային փամփուշտը միևնույնն է, այդպես որ բոլոր օրգանիզմների մեկնակետը մի ընդհանուր սկզբնավորություն է։ Եթե մենք նույնիսկ կանգ առնենք երկու գլխավոր ստորաբաժանումների վրա, այսինքն կենդանիների և բույսերի վրա, ապա մի քանի ստորակարգ ձևեր այնքան միջակա բնույթ ունեն, որ բնախույզները բազմիցս վիճել են, թե այդ երկու խմբերից որի մեջ պետք է դասել նրանց։ Ինչպես նկատել է պրոֆ. Ազա-Գրեյը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 68] «ստորակարգ շատ ջրիմուռների սպորները և այլ վերարտադրողական օրգանները սկզբում դրսևորում են կենդանական, իսկ հետագայում անկասկած բուսական գոյություն»։ Ուստի, հատկանիշների տարամիտմամբ ուղեկցվող բնական ընտրության սկզբունքի հիման վրա անհավանական չէ, որ որևէ նման ստորակարգ կազմվածք ունեցող և միջակա մի ձևից կարող էին զարգանալ ինչպես կենդանիները, այնպես էլ բույսերը․ իսկ եթե մենք ընդունում ենք այդ, ապա մենք պետք է ընդունենք նաև այն, որ երկրագնդի վրա երբևիցե ապրած բոլոր օրգանական էակները կարող են ծագել մի նախնական ձևից։ Բայց այս եզրակացությունը հենվում է գլխավորապես անալոգիայի վրա, և էական չէ, թե նա կընդունվի, թե ոչ։ Իհարկե միանգամայն հնարավոր է, ինչպես մատնանշել է Ջ. Հ. Լյուիսը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 135] որ կյանքի սկզբնական ծագման ժամանակ երևացել են բազմաթիվ զանազան ձևեր․ բայց եթե այդ այդպես է, ապա մենք կարող ենք եզրակացնել, որ նրանցից միայն քչերն իրենցից հետո թողել են փոփոխված սերունդներ, որովհետև, ինչպես ես հենց նոր նշեցի, յուրաքանչյուր խոշոր բաժնի անդամների վերաբերմամբ, ինչպիսիք են ողնաշարավորները, հոդվածոտանիները և այլն, մենք ունենք պարզ ապացույցներ, որոնք բխում են նրանց սաղմնաբանական, հոմոլոգիական ե թերաճիկային կառուցվածքից, որ յուրաքանչյուր բաժնի սահմաններում եղած բոլոր անդամները ծագել են մի ընդհանուր նախահորից։
Երբ այս գրքում իմ կողմից, ինչպես նաև մ-ր Ուոլլեսի[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 10] կողմից արծարծած տեսակետները, կամ նման հայեցակետները տեսակների ծագման մասին ընդհանուր ընդունելություն կգտնեն, մենք կարող ենք աղոտ կերպով նախատեսնել խոր հեղաշրջում բնագիտության ասպարեզում։ Սիստեմատիկները կկարողանան առաջվա նման զբաղվել իրենց գործով, բայց նրանք ազատված կլինեն շարունակ ուրվականի պես նրանց հետապնղող կասկածից, թե արդյոք այս կամ այն ձևը իսկական տեսակ է, թե ոչ։ Եվ ես հավատացած եմ,— այս ասում եմ իմ փորձի հիման վրա,— որ դա մեծ թեթևություն կլինի։ Կդադարեն անվերջ վեճերն այն մասին, թե բրիտանական մոշի ինչ որ հիսուն տեսակներ արդյոք լավ տեսակներ են, թե ոչ։ Սիստեմատիկներին միայն մի հարց կմնա լուծելու (որը այնքան էլ հեշտ չէ), արդյոք բավականաչափ կայուն է այս կամ այն ձևը և արդյոք նա բավականաչափ տարբերվում է այլ ձևերից, որպեսզի նա որոշելու ենթարկվի. և եթե այդ որոշումը հնարավոր է, ապա տարբերությունը արդյոք բավականաչափ էական է, որպեսզի նա արժանանա առանձնահատուկ անվան։ Վերջին կետն անհամեմատ ավելի շատ ուշադրության է արժանի, քան այդ ընդունված է ներկայումս, որովհետև ժամանակակից բնախույզների մեծամասնությունն ընդունում է, որ ինչքան էլ չնչին լինի երկու որևէ ձևի միջև եղած տարբերությունը, եթե միայն նրանք միացած չեն միջակա աստիճաններով, այդ բավական է երկու ձևերը տեսակների աստիճանի բարձրացնելու համար։
Այնուհետև մենք հարկադրված ենք ընդունելու, որ տեսակի և լավ արտահայտված տարատեսակների միջև եղած միակ տարբերությունը միայն այն է, որ վերջինները, ինչպես հայտնի է կամ ենթադրվում է, միմյանց հետ ներկայումս կապված են միջակա աստիճաններով, մինչդեռ տեսակները նախկինում են կապված եղել նույնպիսի ձևով։ Հետևապես, չժխտելով երկու ձևերի միջև միջակա աստիճանների առկայության նշանակությունը, մենք հարկադրված կլինենք ավելի մանրազնին կերպով կշռադատելու և ավելի բարձր գնահատելու նրանց միջև եղած տարբերությունների իսկական գումարը։ Շատ հավանական է, որ այն ձևերը, որոնք ներկայումս սովորաբար ընդունվում են որպես տարատեսակներ, հետագայում արժանի համարվեն առանձին տեսակային անունների, և այդ դեպքում գիտական և առօրյա լեզուները մեծ համաձայնության կհասնեն։ Մի խոսքով, մենք պետք է տեսակներին վերաբերվենք այնպես, ինչպես որ սեռերին վերաբերվում են այն բնախույզները, որոնք ընդունում են, որ սեռերը միայն հարմարության համար հորինված արհեստական, խմբավորումներ են։ Գուցե թե սա այնքան էլ շատ մխիթարական հեռանկար չէ, բայց մենք ընդմիշտ կազատվենք «տեսակ» բառի մինչև այժմ չըմբռնված և անըմբռնելի էության ապարդյուն որոնումներից։
Բնապատմության մյուս ավելի ընդհանուր բաժինները ավելի մեծ հետաքրքրություն կստանան։ Բնախույզների գործածած արտահայտությունները՝ «ազգակցություն, ազգակցական կապ, տիպի ընդհանուր լինելը, հայրություն, մորֆոլոգիա, հարմարում, ռուդիմենտար և աբորտիվ (այլասերված) օրգաններ» և այլն, կդադարեն փոխաբերություններ լինելուց և կստանան հստակ իմաստ։ Երբ մենք կդադարենք օրգանական էակի վրա այնպես նայելուց, ինչպես վայրենին նայում է նավի վրա, այսինքն՝ որպես նրա ըմբռնողության համար բոլորովին անհասանելի ինչ-որ բանի վրա. երբ բնության յուրաքանչյուր արտադրանք մենք կդիտենք որպես մի բան, որն ունի երկար պատմություն, երբ ամեն մի բարդ կառուցվածքի կամ բնազդի մեջ մենք կտեսնենք այն բազմաթիվ հարմարեցումների հանրագումարը, որոնցից յուրաքանչյուրը օգտակար է իր տիրոջ համար, այնպես, ինչպես ամեն մի մեխանիկական մեծ գյուտ բազմաթիվ աշխատավորների աշխատանքի, փորձի, բանականության և մինչև անգամ սխալների հանրագումարն է. երբ մենք օրգանական էակների նկատմամբ կմշակենք այդպիսի տեսակետ,— այն ժամանակ որքա՜ն անչափելի կմեծանա,— ասում են անձնական փորձի հիման վրա,— հետաքրքրությունը դեպի բնապատմությունը։
Կբացվի փոփոխականության պատճառների և օրենքների, զարգացման հարաբերակցությունների, վարժեցման ու ոչ վարժեցման ներգործությունների, արտաքին պայմանների անմիջական ներգործության և այլոց վերաբերյալ ուսումնասիրությունների մի հսկայական և գրեթե կուսական դաշտ։ Հսկայական չափով կմեծանա այն հետաքրքրությունը, որ ներկայացնում է մեր ընտանի ցեղերի ուսումնասիրությունը։ Մարդու ստեղծած մի նոր տարատեսակը ուսումնասիրության ավելի հետաքրքրական և ավելի կարևոր առարկա կլինի, քան էլի մի տեսակի ավելացումը ցուցակի մեջ արդեն գրանցվածների անհամար թվի վրա։ Մեր դասակարգությունները կվերածվեն, որքան այդ հնարավոր է, ծննդաբանությունների, և այն ժամանակ նրանք իսկապես կներկայացնեն մեզ այն, ինչ որ իրավամբ կարելի է անվանել ստեղծագործության պլան։ Դասակարգման կանոնները անկասկած ավելի պարզ կդառնան այն վայրկյանից սկսած, երբ մենք նկատի կունենանք միանգամայն որոշակի խնդիր։ Մեզ մոտ չեն պահպանվել ոչ ծննդաբանական, ոչ էլ հերալդիկական ցուցումներ, և մենք պետք է երևան հանենք մեր բնական ծննդաբանությունների բազմաթիվ տարամիտվող գծերի հետքերը՝ առաջնորդվելով ամեն մի հատկանիշով, որը վաղուց ի վեր ժառանգաբար փոխանցվել է։ Թերաճ օրգաններն անսխալ կերպով կվկայեն վաղուց կորցված կառուցվածքների բնույթի մասին։ Այն տեսակները և տեսակների խմբերը, որոնք համարվել են անոմալիաներ և որոնց թերևս կարելի է կենդանի բրածոներ անվանել, կօգնեն մեզ կյանքի հին ձևերի մասին պատկերացում կազմելու։ Սաղմնաբանությունը հաճախ կհայտնաբերի յուրաքանչյուր խոշոր դասի նախատիպերի որոշ չափով քողարկված կառուցվածքը։
Երբ մենք կհամոզվենք, որ միևնույն տեսակի բոլոր անհատները և սեռերի մեծամասնության պատկանող միմյանց հետ մոտ ազգակից տեսակները ոչ այնքան հեռավոր ժամանակաշրջանի սահմաններում առաջացել են մի ընդհանուր ծնողից և իրենց մի ծննդավայրից տարաբնակվել են, երբ մենք ավելի լավ կծանոթանանք տարաբնակման բազմազան եղանակների հետ, այն ժամանակ երկրագնդի կլիմայի և ռելիեֆի մեջ եղած փոփոխությունների վրա երկրաբանության արդեն սփռած և դեռ սփռվելիք լույսի տակ մենք իհարկե ի վիճակի կլինենք հիանալի կերպով հետևելու նաև ամբողջ երկրագնդի բնակիչների տարաբնակման նախկին եղանակներին։ Նույնիսկ ներկայումս մայր ցամաքի հակադիր կողմերում գտնվող ծովերի բնակիչների և այդ մայր ցամաքի զանազան բնակիչների բնույթի տարբերության համեմատությունը նրանց տարածման հավան ական եղանակի կապակցությամբ արդեն որոշ լույս է սփռում հին աշխարհագրության վրա։
Երկրաբանությունը, այդ ընտիր գիտությունը, անշուշտ իր փառքի որոշ մասը կորցնում է իր տարեգրության չափազանց թերի լինելու պատճառով։ Երկրագնդի կեղևը իր մեջ ամփոփված օրգանական մնացորդներով չի կարող հարուստ թանգարան համարվել, այլ ավելի շուտ որպես աղքատ հավաքածու է, որը հավաքված է պատահական կերպով և ժամանակի երկարատև ընդմիջումներով։ Պետք է ընդունել, որ մեծ և բրածոներով հարուստ ֆորմացիաներն իրենց գոյացմամբ պարտական են բարենպաստ հանգամանքների արտասովոր զուգորդմանը և որ իրար հաջորդող հարկերի միջև եղած դատարկ տարածությունները համապատասխանում են հսկայական տևողություն ունեցող ժամանակաշրջաններին։ Սակայն մենք այդ ընդմիջումների տևողությունը ի վիճակի կլինենք գնահատելու նախորդող և հաջորդող օրգանական ձևերը համեմատելու միջոցով։ Մենք մեծ զգուշությամբ կարող ենք երկու ֆորմացիաներ խիստ ժամանակակից համարել, եթե նրանք իրենց մեջ չեն պարփակում բազմաթիվ նույնանման տեսակներ՝ շնորհիվ կյանքի ձևերի հաջորդականության։ Տեսակներն առաջանում և բնաջնջվում են շնորհիվ դեռ հիմա էլ գոյություն ունեցող պատճառների դանդաղ ներգործության, այլ ոչ թե շնորհիվ հրաշքային ստեղծագործական ակտերի։ Օրգանական փոփոխությունները հարուցող բոլոր պատճառներից ամենաէականներն են նրանք, որոնք գրեթե անկախ են փոփոխվող և երբեմն էլ հանկարծակի փոփոխվող ֆիզիկական պայմաններից, այսինքն՝ օրգանիզմների միջև եղած փոխհարաբերությունները, ըստ որում մի օրգանիզմի փոփոխությունն առաջ է բերում մյուսների կատարելագործում կամ ոչնչացում։ Այս երկու դրույթներից հետևում է այն, որ իրար հաջորդող ֆորմացիաների բրածոների օրգանական փոփոխությունների աստիճանը, ըստ երևույթին, լավ չափանիշ է ծառայում անցած ժամանակամիջոցի հարաբերական, բայց ոչ բացարձակ տևողության համար։ Այնուամենայնիվ, որոշ թվով տեսակներ համատեղ գոյություն ունենալիս կարող են երկար ժամանակ պահպանվել առանց փոփոխության, մինչդեռ նույն ժամանակաշրջանի ընթացքում այն տեսակներից մի քանիսը, որոնք գաղթելով նոր երկրներ և մրցության մեջ մտնելով օտարերկրացիների հետ, կարող են փոփոխվել․ հետևապես, մենք չափազանց մեծ նշանակություն չպետք է տանք օրգանական փոփոխություններին որպես ժամանակի չափանիշ ծառայելու տեսակետից։
Ապագայում, ես նախատեսում եմ, ասպարեզ կբացվի հետագա է՛լ ավելի կարևոր հետազոտությունների համար։ Հոգեբանությունը ամուր հենարան կստանա այն հիմքի վրա, որ հիանալի կերպով արդեն գրել է Հերբերտ Սպենսերը,[ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ 18] այսինքն՝ ամեն մի մտավոր հատկություն և ընդունակություն աստիճանական գրադացիաների միջոցով ձեռք բերելու անհրաժեշտությունը։ Շատ կլուսաբանվի մարդու ծագումը և նրա պատմությունը։
Ականավոր շատ հեղինակներ, ըստ երևույթին, միանգամայն բավարարված են այն տեսակետով, որ յուրաքանչյուր տեսակ անկախ է ստեղծվել։ Իմ հասկացողությամբ՝ արարչի կողմից նյութական բնության երևույթների հիմքում դրված օրենքներին ավելի համապատասխանում է այն, որ երկրագդնդի անցյալ ու ներկա բնակիչների առաջացումը և անհետացումը ղեկավարվում է նույնպիսի երկրորդային պատճառներով, ինչպես որ նրանք, որոնք որոշում են անհատների ծնունդն ու մահը։ Երբ ես նայում եմ բոլոր էակների վրա ոչ որպես առանձին արարչագործական ակտերի արդյունքների, այլ որպես կեմբրիական սիստեմի աոաջին շերտերի կուտակումից շատ առաջ ապրած սակավաթիվ էակների անմիջական հաջորդների վրա, ապա նրանք ինձ թվում են ազնվացրած։ Դատելով ըստ անցյալի, մենք կարող ենք վստահ կերպով եզրակացնել, որ այժմ ապրող ոչ մի տեսակ չի փոխանցի իր հեռավոր հաջորդներին իր չփոփոխված նմանությունը, իսկ այժմ գոյություն ունեցողներից միայն քչերը որևէ սերունդ կթողնեն մինչև հեռավոր ապագան, որովհետև բոլոր օրգանական էակների ընդհանուր խմբավորումը ցույց է տալիս, որ յուրաքանչյուր սեռի մեջ տեսակների մեծ մասը և շատ սեռերի բոլոր տեսակները սերունդներ չեն թողել, այլ վերջնականապես բնաջնջվել են։ Մենք կարող ենք այնքան մարգարեաբար նայել ապագային, որպեսզի կանխագուշակենք, որ ամենասովորական և լայնորեն տարածված տեսակները, որոնք յուրաքանչյուր դասի սահմաններում պատկանում են ընդարձակ ու տիրապետող խմբերին, վերջ ի վերջո կհաղթեն և ծնունդ կտան նոր տիրապետող տեսակների։ Քանի որ բոլոր ապրող ձևերը ընդհանուր ծննդաբանությամբ կապված են նրանց հետ, որոնք կեմբրիական դարաշրջանից շատ առաջ են ապրել, ուստի մենք կարող ենք համոզված լինել, որ սերունդների ընդհանուր հերթափոխությունը ոչ մի անգամ չի ընդհատվել և որ ոչ մի աղետ երբեք ավերածություններ չի տարածել ամբողջ աշխարհի վրա։ Այդ պատճառով մենք կարող ենք հանգիստ լինել անվտանգ ապագայի համար երկար ժամանակ։ Եվ քանի որ բնական ընտրությունը ներգործում է յուրաքանչյուր էակի օգտին, ապա բոլոր մարմնական ու մտավոր հատկությունները պետք է ձգտեն կատարելագործության։
Հետաքրքիր է դիտել խիտ բուսակալած մի ափ, որը ծածկված է բազմաթիվ բազմազան բույսերով, որտեղ թռչունները երգում են թփերի մեջ, շուրջդ ճախրում են միջատները, խոնավ հողում սողում են որդերը, և մտածել, որ այդ հիանալի կերպով կառուցված, այնքան միմյանցից տարբեր և այնքան բարդ կերպով մեկը մյուսից կախված ձևերը ստեղծվել են այն օրենքների շնորհիվ, որոնք դեռ հիմա էլ գործում են մեր շուրջը։ Այդ օրենքները ամենալայն իմաստով հետևյալներն են՝ աճումն ու վերարտադրումը, ժառանգականությունը, որը գրեթե անհրաժեշտորեն բխում է վերարտադրումից, փոփոխականությունը, որը կախված է կենսապայմանների ուղղակի կամ անուղղակի ներգործությունից կամ վարժեցումից և ոչ վարժեցումից․ բազմացման պրոգրեսիան, որն այնքան բարձր է, որ հանգում է գոյության կռվի ու նրա հետևանքին՝ բնական ընտրությանը, որն առաջ է բերում հատկանիշների տարամիտում և նվազ կատարյալ ձևերի մահաջնջում։ Այսպիսով, բնության մեջ այդ մոլեգնող պատերազմից, սովից ու մահից անմիջականորեն բխում է այն ամենաբարձր հետևանքը, որ մեր միտքը կարող է պատկերացնել, այն է՝ կենդանական կյանքի բարձրագույն ձևերի առաջացումը։ Կա վեհություն այդ տեսակետի մեջ կյանքի վերաբերմամբ՝ նրա զանազան ուժերով հանդերձ, որ սկզբից ի վեր արարիչը ներդրել է աննշան թվով ձևերի մեջ կամ միայն մեկի մեջ և մինչդեռ մեր մոլորակն իր ծիրն է կատարել և շարունակում է կատարել տարածության մեջ ձգողականության անփոփոխ օրենքների համաձայն, այդպիսի պարդ սկզբնավորությունից առաջացել են և շարունակում են զարգանալ զարմանալիորեն կատարյալ ու սքանչելի անհամար ձևեր։
- ↑ «Արիստոտելը իր «Physicae Auscultationes»-ում (գիրք 2, գլ. 8, էջ 2), այն դիտողությունից հետո, որ անձրևը գալիս է ոչ թե նրա համար, որպեսզի հացահատիկների բերքին նպաստի, ճիշտ այդպես էլ ոչ նրա համար, որպեսզի փչացնի այն հացահատիկը, որը կալսում են բաց երկնքի տակ, միևնույն փաստարկը կիրառում է նաև օրգանիզմների նկատմամբ. նա ավելացնում է (ինչպես այդտեղը թարգմանում է Կլեր Գրեսը, որն առաջին անգամ իմ ուշադրությունը դարձրեց նրա վրա). «Բնության մեջ ի՞նչն է խանգարում (մարմնի) զանազան մասերին, որ միմյանց նկատմամբ գտնվեն նույնպիսի պատահական հարաբերության մեջ։ Ատամները, օրինակ, աճում են ըստ անհրաժեշտության՝ առջևինները սուր են և հարմարված են սնունդը պատառ-պատառ անելու համար, իսկ սեղանատամները տափակ են, պիտանի են կերակուրը աղալու համար, բայց նրանք ստեղծված չեն դրա համար, այլ պատահականության արդյունք են։ Նմանապես և մյուս մասերի վերաբերմամբ, որոնք մեզ թվում են որևէ նպատակի համար հարմարված։ Այսպիսով, ամենուրեք, որտեղ առարկաներն ամբողջությամբ վերցրած (ինչպես, օրինակ, ամբողջության մասերը) մեզ այնպես են թվում, կարծես պատրաստված են ինչ-որ նպատակով, իրականում նրանք միայն պահպանվել են, որովհետև ինչ-որ ներքին ձգտման շնորհիվ համապատասխանորեն կառուցված են եղել. իսկ բոլոր այն առարկաները, որոնք այդպես կառուցված չեն եղել, ոչնչացել են և շարունակում են ոչնչանալ»։ Մենք այստեղ կարծես նկատում ենք բնական ընտրության ապագա սկզբունքի նշույլը, բայց թե որքան քիչ էր ըմբռնում Արիստոտելը այդ սկզբունքի էությունը, երևում է նրա դիտողություններից ատամների մասին։
- ↑ «Կենդանաբանության փիլիսոփայությունը»։— Խմբ.։
- ↑ «Անողնաշարավոր կենդանիների բնական պատմությունը»։— Խմբ.։
- ↑ Լամարկի առաջին աշխատության հրատարակության տարեթիվը ես վերցրել եմ Իսիդոր Ժոֆֆրուա Սենտ-Իլերից, որը իր գրքում (Hist. Nat. Génerale, t. II, p. 405, 1859 թ.) տվել է այդ հարցի վերաբերյալ հղած տարբեր հայացքների մասին հիանալի պատմական ակնարկ։ Այդ աշխատությունում կարելի է գտնել նաև Բյուֆոնի հայացքների մասին լրիվ ակնարկ։ Հետաքրքիր է, թե որքան լայն չափով իմ պապս՝ Էրազմ Դարվինը իր «Զոոնոմիայում» (հատոր 1, էջ 500—510), որը լույս է տեսել 1794 թվականին, նախապես կռահել է այն հայացքները և սխալ հիմունքները, որոնցով առաջնորդվել է Լամարկը։ Իսիդոր Ժոֆֆրուայի կարծիքով, կասկածի ենթակա չէ, որ Գյոթեն եղել է նման հայացքների ջերմ կողմնակից, ինչպես այդ հետևում է 1794 և 1795 թվականներին նրա գրած, բայց ավելի ուշ հրատարակած աշխատության առաջաբանից. նա միանգամայն որոշակի կերպով հայտնում է այն միտքը («Goethe als Naturforscher» Կարլ Մեդինգի, էջ 34), որ ապագայում բնախույզին պետք է զբաղեցնի այն հարցը, թե եղջերավոր անասունը ինչպես է ձեռք բերել իր եղջյուրները, այլ ոչ թե այն հարցը, թե նրանք ինչի համար են հարկավոր նրան։ Որպես հիանալի օրինակ այն բանի, թե ինչպես իրար նման գաղափարները կարող են միաժամանակ, բայց բոլորովին անկախ կերպով ծագել, ծառայում է այն փաստը, որ Գյոթեն Գերմանիայում, Է. Դարվինը Անգլիայում և Ժոֆֆրուա Սենտ-Իլերը (ինչպես հիմա կտեսնենք) Ֆրանսիայում 1794—1795 թվականներին՝ կարճ ժամանակամիջոցում եկան միևնույն եզրակացություններին տեսակների ծագման մասին։
- ↑ Շրջապատող աշխարհը։— Խմբ.։
- ↑ «... այդ պրոբլեմը պետք է ամբողջովին հանձնել ապագային, եթե միայն ապագայում նրանով կզբաղվեն»։— Խմբ.։
- ↑ «Պարտիզագործության վերաբերյալ աշխատություններ»։— Խմբ.։
- ↑ Էդինբուրգի փիլիսոփայական ամսագիր։— Խմբ.։
- ↑ «Լիննեյան ամսագիր»։— Խմբ.։
- ↑ «Պարտիզագործի լրատու»։— Խմբ.։
- ↑ «Կանարյան կղզիների ֆիզիկական նկարագրությունը»։— Խմբ.։
- ↑ Բնական գիտության բոստոնյան ամսագիր։— Խմբ.։
- ↑ «Ստեղծագործության հետքերը»։— Խմբ.։
- ↑ «Թագավորական ակադեմիայի բյուլետեն Բրյուսելում»։— Խմբ.։
- ↑ «Վերջավորությունների բնույթը»։— Խմբ.։
- ↑ Ողնաշարավորների անատոմիան։— Խմբ.։
- ↑ «Լանդոնյան հանդես»։— Խմբ.։
- ↑ «Կենդանաբանության ամսագիր»։— Խմբ.։
- ↑ «Նրանք հաստատուն են յուրաքանչյուր տեսակի համար, քանի դեռ նա բազմանում էր միևնույն պայմաններում. հատկանիշները փոխվում են, երբ սկսում են փոխվել շրջապատող պայմանները»։ Ընդհանուր առմամբ վայրի կենդանիների վրա կատարված դիտումը ակնառու կերպով ցույց է տաքիս տեսակների սահմանափակ փոփոխականությունը։ Ընտելացված վայրի կենդանիների և վայրենացած ընտանի կենդանիների վրա կատարված փորձերը ավելի ևս պարզորոշ կերպով ապացուցում են այդ դրույթը, դեռ ավելին՝ հենց այդ փորձերը ապացուցում են, որ ստեղծված տարբերությունները կարող են այնպիսի նշանակություն ունենալ, որը հատուկ է տարբեր սեռերի։
- ↑ «Ընդհանուր բնական պատմություն»։ Կա այդ գրքի ռուսերեն թարգմանությունը «Ընդհանուր բիոլոգիա» վերնագրով։ Մ. 1880, հատոր I։— Խմբ.։
- ↑ «Դուբլինյան բժշկական ամսագիր»։— Խմբ.։
- ↑ «Փորձեր»։— Խմբ.։
- ↑ «Պարտեզագործական տեղեկատու»։— Խմբ.։
- ↑ «Մուզեի նոր արխիվները»։ — Խմբ.։
- ↑ [«Խորհրդավոր, անորոշ ուժ. ոմանց համար ճակատագրականությունը, մյուսների համար նախախնամության կամքն է, որի անդադար ներգործությունը կենդանի էակների վրա աշխարհի գոյության բոլոր դարաշրջաններում որոշում է նրանցից յուրաքանչյուրի ձևը, ծավալը և երկարակեցությունը՝ համապատասխան նրա նշանակության այն իրերի կարգում, որոնց մասն է կազմում նա։ Հենց այդ ուժն է, որ յուրաքանչյուր անդամը համապատասխանեցնում է ամբողջին, հարմարեցնելով նրան այն դերին, որ նա պիտի կատարի բնության ընդհանուր օրգանիզմում, այն դերին, որ նրա գոյության իմաստն է»]։
Բրոննի «Untersuchungen über die Entwickelungs Gesetze»-ում ասածների հիման վրա պարզվում է, որ նշանավոր բուսաբան և հնէաբան Ունգերը 1852 թ. տպագիր կերպով հայտնել է իր համոզմունքը, որ տեսակները փոփոխվում և զարգանում են։ 1821 թ. նույն համոզմունքն է հայտնել Դալտոնը բրածո համրուկների մասին իր և Պանդերի կողմից կատարած ուսումնասիրության մեջ։ Նման հայացքներ, ինչպես հայտնի է, հայտնել է նաև Օկեանը իր միստիկական «Natur Philosophie»-ում։ Գոդրոնի «De l’Espèce» գրքում հանդիպող այլ վկայակոչումների հիման վրա պարզվում է, որ Բորի Սեն Վենսենը, Բուրդախը, Պուարեն և Ֆրիսն ընդունել են, որ նոր տեսակները շարունակ նորից առաջանում են։ Ես կարող եմ ավելացնել, որ այս պատմական ակնարկում հիշատակված և տեսակների փոփոխականության վերաբերմամբ իրենց համոզմունքը արտահայտած կամ, համենայն դեպս, առանձին ստեղծագործական ակտեր չընդունող երեսուն և չորս հեղինակներից քսանյոթը միևնույն ժամանակ եղել են բնապատմության և երկրաբանության զանազան բնագավառներում հատուկ հետազոտությունների հեղինակներ։ - ↑ «Երկրաբանական ընկերության բյուլետեն»։— Խմբ.։
- ↑ «Հռենոսյան մարզի բնախույզների ընկերության աշխատություններ»։— Խմբ.։
- ↑ «Ուրվագծեր բույսերի աշխարհագրության մասին»։— Խմբ.։
- ↑ «Այստեղից երևում է, որ տեսակների անփոփոխության կամ փոփոխականության մասին մեր որոնումները մեզ ուղղակի հանգեցնում են այն գաղափարներին, որ հռչակել են երկու իրավամբ նշանավոր մարդիկ՝ Ժոֆֆրուա Սենտ-Իլերը և Գյոթեն»։— Խմբ.։
- ↑ «Կենդանաբանական—մարդաբանական հետազոտություններ»։— Խմբ.։
- ↑ Տ՛ես 161 էջի ծանոթությունը։— Խմբ.։
- ↑ Դարվինի համաձայնելը Վեյսմանի հետ «օրգանիզմի բնույթի և պայմանների բնույթի» վերաբերմամբ թեպետև պատահական չէ, որովհետև կապի մեջ է գտնվում փոփոխությունների, որպես «ընտրության ուղղության նկատմամբ պատահական» լինելու մասին նրա տված սխալ մեկնաբանության հետ, սակայն իրականում դա ձևական «համաձայնություն» է։ Դարվինը դիտավորություն ունի ընդգծելու օրգանիզմի ժառանգական բնույթի յուրահատկության նշանակությունը տարբեր օրգանիզմների վրա միևնույն պայմանների ազդեցության հետևանքների յուրահատկության համար, իսկ Վեյսմանն ընդհանրապես ժխտում է այն փոփոխությունների ժառանգելիությունը, որոնք առաջանում են արտաքին պայմանների ազդեցությամբ։— Խմբ.։
- ↑ Այստեղից երևում է, որ Դարվինը այսպես կոչված, «անորոշ փոփոխությունները» դիտում է որպես որոշակի, բայց պոպուլյացիայի ներսում անհատների բազմազան փոփոխություններ, որոնք հակադրվում են տեսակի բոլոր անհատների կամ մեծ մասի բազմաթիվ հարաբերաբար միանման փոփոխություններին։ Անհատական փոփոխությունների բազմազանությունը պայմանավորված է նրանով, որ պայմանների ազդեցությանը ենթարկվող անհատները իրենց դրությամբ կամ կազմությամբ նույնը չեն, որից և առաջանում է ազդեցության հետևանքների ոչ նմանությունը։ Սակայն ամբողջությամբ վերցրած տեսակի համար այդ անորոշ փոփոխությունները միանգամայն որոշակի են նրանով, որ նրանցից յուրաքանչյուրը առաջանում է որոշակի պատճառից և իր ուղղությամբ միանգամայն որոշակի կերպով, որովհետև «անորոշ փոփոխությունները» գումարվում են սերունդներում։— Խմբ.։
- ↑ Նեկտարիններ կոչվում են հարթ, առանց աղվամազի մաշկով դեղձերը։— Խմբ.։
- ↑ Այստեղ Դարվինը իրոք անցնում է Վեյսմանի դիրքավորման կողմը, նահանջելով իր բացատրությունից, որ փոփոխականություն առաջանում է օրգանիզմի վրա կենսապայմանների գործած ազդեցությունից։ Բույսերի կարծես թե հանկարծակի փոփոխությունների, ինչպես քննարկվող «սպորտների» առաջացման ուղիների հարցը տեսականորեն պարզաբանված են Միչուրինի կողմից, իսկ նրանից հետո Տ. Դ. Լիսենկոյի կողմից օրգանիզմների գենետիկական տարբերորակության ուսմունքի մեջ։— Խմբ.։
- ↑ Բերված օրինակներով Դարվինը մատնացույց է անում այն, որ փոփոխությունների հարաբերակցականության իսկական պատճառն է նրանց համար ընդհանուր ֆիզիոլոգիական հիմքը. հարաբերակցությունների պատճառների որոնումների համար ընդհանուր ուղղության այդ ցուցումով Դարվինը արդեն վերացնում է նրանց «խորհրդավորությունը»։— Խմբ.։
- ↑ Քարային, ժայռային, թխագորշ կամ վայրի աղավնին — Columba livia, որը ներկայումս հանդիպում է Ղրիմի, Կովկասի, Միջին Ասիայի լեռներում, Ուրալի հարավային լեռնաճյուղերում, Ալթայում, Միջերկրական ծովի ափերում և կղզիների վրա, Իռլանդիայում և Շոտլանդիայում, Հյուսիսային Աֆրիկայում, Փոքր Ասիայում, Սիրիայում և Իրանում։ Ժայռաղավնուց տարբերում են դաշտային թխագորշ աղավնուն։ Այս ցեղը արտաքուստ շատ նման է ժայռաղավնուն և վարում է միանգամայն վայրի կենսակերպ, բայց և այնպես մարդուն մոտիկ տեղերում։— Խմբ.։
- ↑ Դարվինի այդ եզրակացությունը բոլորովին սխալ է։ «Տեսակ» տերմինը բնության մեջ համապատասխանում է կենդանի նյութի գոյության որոշակի ձևերի միջև եղած իրական որակական տարբերությանը, որը չի հանգում զուտ քանակական տարբերությունների, որոնցով զանազանվում են տարատեսակները տեսակի ներսում։ Այդ ապացուցված է նաև աղավնիների ցեղերի մասին տրված դարվինյան վերլուծությամբ։ Տե՛ս նաև «Հատկանիշների տարամիտումը» գլուխ 4, էջ 199։— Խմբ.։
- ↑ Դարվինը այստեղ ճիշտ կերպով է վկայակոչում Մալթուսին։ Բայց փորձելով բնական ընտրության վերաբերմամբ կիրառել Մալթուսի տեսությունը, Դարվինն իրեն համար աննկատելի կերպով ցույց է տալիս, որ Մալթուսի տեսությունը անկիրառելի է վայրի տեսակների կյանքի վերաբերմամբ։ Մարքսիզմի հիմնադիրները խիստ սահմանազատում էին Դարվինի ուսմունքի էությունը և մալթուսյան «գոյության կռիվը»։ Այսպես, Մարքսն ասում է. «Դարվինի աշխատության մեջ, օրինակ, տեսակների մահաջնջման պատճառների քննարկության մեջ կա նաև — չխոսելով նրա հիմնական սկզբունքի մասին — մալթուսյան տեսության մանրամասն բնապատմական հերքում»։ (Կ. Մարքս, Հավելյալ արժեքի տեսությունը, հատոր 2, մաս 1, էջ 209, Կուսհրատ, 1936 թ.)։— Խմբ.։
- ↑ «Հյուսիսային բրիտանական հանդես»։— Խմբ.։
- ↑ М. Вагнер. Дарвин и его закон переселения, թարգմ. գերմաներենից, Անդրեևսկու խմբագր.։ ՍՊԲ, 1870 թ., էջ 1—64։— Խմբ.։
- ↑ Այստեղ Դարվինն ընդհուպ մոտենում է տեսակների և տարատեսակների միջև գոյություն ունեցող տարբերությունների անհավասարարժեքության ճիշտ ըմբռնմանը և ճիշտ եզրակացություն է անում, որ նախահայրերի հատկանիշներից շեղումների պարզ շարունակումը դեռ բավական չէ հին տեսակի մի տարատեսակը նոր տեսակի փոխակերպվելու համար։ Տեսակների անջատվածության մասին հետագայում Դարվինի տված բացատրությունը դարձյալ հանգում է միջանկյալ ձևերի ոչնչացմանը։— Խմբ.։
- ↑ Հատկանիշների տարամիտումը հաճախ նշվում է «դիվերգենցիա» տերմինով։ Դրանով հասկացվում է մի ընդհանուր նախահոր հաջորդներում ավելի ու ավելի բազմազան էակների առաջացումը։
- ↑ Հատկանիշների մերձեցումը կամ մոտեցումը հաճախ նշվում է «կոնվերգենցիա» տերմինով։— Խմբ.։
- ↑ Կարնիոլիա կամ Կրայնա — Ավստրիայի ամենահարավային մարզերից մեկն է, Կենտուկկի — Հյուսիսային Ամերիկայի նահանգներից մեկն է։— Խմբ.։
- ↑ Իսկական պատճառը։— Խմբ.։
- ↑ Սարկոդա խոսքով Դարվինի ժամանակները հաճախ հասկանում էին պրոտոպլազմա։— Խմբ.։
- ↑ Բնությունը թռիչքներ չի անում։— Խմբ.։
- ↑ Բնությունը թռիչք չի անում։— Խմբ.։
- ↑ Այստեղ Դարվինը մոտենում է այսպես կոչված մուտացիաների պատճառների հայտնաբերմանը։ Դրանք օրգանիզմի վրա արտաքին պայմանների կրկնական ազդեցությունների կուտակման արդյունքներ են, որոնք սկզբում միայն արտահայտվում են օրգանիզմի բնույթի մեջ թաքնված փոփոխությունների ձևով։— Խմբ.։
- ↑ Փակ ծաղիկները սովորաբար կոչվում են կլեյստոգամներ։— Խմբ.։
- ↑ «Միջնապատը, որ քիչ թե շատ լայն է, տեղավորված է պտղապատյանի և սերմնակիրի միջև։— Խմբ.։
- ↑ «Այսպիսով, միևնույն անհատի վրա բներն ու սռնակը երբեմն ամրակցված են ուղղաձիգ առանցքի վրա, երբեմն էլ գինոբազայի (գինեցեի հիմքի) վրա։— Խմբ.։
- ↑ Մոտավորապես մի երրորդական դյույմ — 8—9 մմ. 0,14 դյույմը — 3,3 մմ։— Խմբ.։
- ↑ Գաճաճ շերտավորը ավելի փոքր է մյուս կետերից, նրա երկարությունը 10 մետրից փոքր է։— Խմբ.։
- ↑ Բժշկավիրահատական հանդես։— Խմբ.։
- ↑ Տե՛ս գլուխ XIII. «Աշխարհագրական տարածում»։— Խմբ.։
- ↑ «Բնութլունը թռիչք չի անում» — այս լեյբնիցյան սկզբունքը ներկայացնում է բնության միակողմանի, այդ պատճառով էլ սխալ պատկերում։ Որակական կտրուկ ձևափոխությունները՝ թռիչքաձև անցումները կամ պրոցեսի ընթացքում մի ուղղությունից դեպի մի այլ ուղղություն կատարվող վճռական շրջադարձերը բնության մեջ նույնքան օրինական են, որքան աստիճանական փոփոխությունները միևնույն ուղղությամբ։— Խմբ.։
- ↑ Cervulus՝ մունջակ սեռը միացնում է փոքր եղջերուների մի քանի տեսակներ, որոնք ապրում են հարավային և արևելյան Ասիայի լեռնային անտառներում։— Խմբ.։
- ↑ Միմյանց հետ։— Խմբ.։
- ↑ Ներկայումս խնձորենու բոլոր տեսակները դասվում են Malus սեռի մեջ, մինչդեռ տանձենու բոլոր շատ տարբեր տեսակները՝ Pyrus սեռի մեջ։- Խմբ.։
- ↑ Robinia-ն մեզ մոտ հայտնի է սպիտակ ակացիա անունով։— Խմբ.։
- ↑ Չ. Դարվինի այս եզրակացությունը օգտագործել է ակադեմիկոս Տ. Դ. Լիսենկոն ցորենների ներսորտային խաչաձևումների վրա կատարած իր փորձերում։— Խմբ.։
- ↑ Միամյաներ գնարբուկների ընտանիքից (Primulaceae, Anagallis arvensis L. A. coerula L.)։— Խմբ.։
- ↑ Լեյասը մեզոզոյի ստորին յուրայի նստվածքներն են։— Խմբ.։
- ↑ 66,0 66,1 Fathom = 2 յարդի, կամ 6 ոտնաչափի, այսինքն 1,829 մետրի։— Խմբ.։
- ↑ Սոլենհոֆենում 1869 թ. գտնված է թռչունի մի առանձին փետուր, 1861 թ. նույն տեղում գտել են Archeopteryx թռչունի ոչ լրիվ մի օրինակ, իսկ 1877 թվականին՝ այդ թռչունի լրիվ օրինակը։— Խմբ.։
- ↑ Ատտիկան միջին Հունաստանի հարավ-արևելյան մասն է՝ Աթենք գլխավոր քաղաքով։— Խմբ.։
- ↑ Դարվինի այս դրույթներում է պարփակվում բիոգենետիկական օրենքի էությունը, որի համաձայն օրգանիզմն իր զարգացման ընթացքում կարծես անցնում է այն ստադիաները, որոնք համապատասխանում են նրա նախնիների կառուցվածքում եղած հաջորդական փոփոխություններին։— Խմբ.։
- ↑ Այսպես է բնագրում։ Պետք էր ասել Կասպիական ծովի, քանի որ ըստ Լ. Բերգի վկայության (Աշխարհագր. ընկ. Թուրքեստ. բաժ. Տեղեկագիր, 5-րդ 1908, 451) Արալյան ծովում արալակասպիական կենդանիները գրեթե բացակայում են։— Խմբ.։
- ↑ Իսկական պատճառը։— Խմբ.։
- ↑ Անգլիական ունցիան կազմում է 28,55 գր., հետևաբար, 6¾ ունցիան կլոր հաշված հավասար է 191,4 գրամի։— Խմբ.։
- ↑ «Ընդհանուր մորֆոլոգիա» — Խմբ.։
- ↑ Կաթնասուն խոտակեր է, ապրում է հասարակածային գոտու գետերի մեջ. երկու տեսակ գտնվում են Հարավային Ամերիկայում և մեկ տեսակը արևմտյան Աֆրիկայում։— Խմբ.։
- ↑ Շարքային կամ սերիական հոմոլոգիան ընդգրկում է ժառանգական օրգանները վայրընթաց շարքերի անհատների մոտ (պապ, հայր, թոռ, ծոռ և այլն). կողմնային հոմոլոգիան ընդգրկում է մի սերնդի անհատների օրգանները, որոնք ծագումով համապատասխան են։— Խմբ.։
- ↑ ՍՍՌՄ-ի Կենդանաբանական այգում և գազանաբուծական ֆերմաներում այդ հարցը փղերի և մուշտակամորթ գազանների մասին դրական կերպով է լուծվում։— Խմբ.։
- ↑ Իսկական պատճառ։— Խմբ.։
ԱՆՈՒՆՆԵՐԻ ՑԱՆԿ
- ↑ Բյուֆոն (Buffon Georges-houis Leclerede)։— Հռչակավոր բնախույզ Ֆրանսիայում (1707—1788)։ Հսկայական ազդեցություն է ունեցել 18-րդ դարի բնագիտության զարգացման վրա։ Անդամ ֆրանսիական Գիտությունների ակադեմիայի։ Դիրեկտոր Փարիզի թագավորական այգու, որն իր բազմաթիվ հավաքածուների ստեղծման համար մեծ չափով պարտական է Բյուֆոնին։ Գրել է բնագիտության զանազան բնագավառների վերաբերյալ, այդ թվում նաև 36 հատորից բաղկացած նշանավոր «Բնագիտության պատմությունը», որի մեջ առաջին անգամ տվել է երկրագնդի կենդանական աշխարհի ընդհանուր տեսությունը՝ Երկրի պատմության ֆոնի վրա։ Կլիմայի, սննդի և ընտանեցման պայմանների փոփոխությունների ազդեցության տակ տեսակների ձևափոխության տեսության հեղինակ։
- ↑ 2,0 2,1 2,2 2,3 Լամարկ (Lamarck Jean-Baptiste-Pierre Antoine de Monet)։ Ականավոր բնախույզ Ֆրանսիայում (1744—1829)։ Նորարար բուսաբանության և կենդանաբանության ասպարեզում։ Տվել է էվոլյուցիոն ուսմունքի առաջին գիտական հիմնավորումը, որը շարադրված է նրա «Կենդանաբանության փիլիսոփայությունը» գրքում, որի լույս ընծայման տարին (1809 թ.) կարելի է համարել այդ ուսմունքի հիմնադրման տարին։ Հեղինակ այն օրենքի, որի համաձայն արտաքին միջավայրի ազդեցության տակ օրգանիզմների ձեռք բերած նոր հատկությունները ժառանգական են։ Կենդանաբանության և բուսաբանության վերաբերյալ նրա մյուս աշխատությունները, ինչպես օրինակ, «Անողնաշարավորների բնապատմությունը», «Ֆրանսիայի բուսականությունը» և այլն, հանդիսացել են իրենց ժամանակի համար առաջնակարգ երկասիրություններ։ Արտահայտել է մի շարք ուշագրավ գաղափարներ նյարդային սիստեմի դերի մասին, «ներքին միջավայրի» մասին և գլխուղեղի կեղևի հատուկ նշանակության մասին։
- ↑ 3,0 3,1 Ժոֆֆրուա Սենտ-Իլեր Էտիեն (ավագ) (Geoffroy Saint Hilaire-Etienne)։— Հռչակավոր կենդանաբան Ֆրանսիայում (1772—1844)։ Փարիզի Բուսաբանական այգու կաթնասունների բաժանմունքի վարիչ, 1794 թ. ստեղծել է ողնաշարավորների կենդանաբանության առաջին ամբիոնը Փարիզում։ Գրել է բազմաթիվ աշխատություններ, որոնց նպատակն է ապացուցել օրգանիզմների կառուցվածքի միասնականությունը և էվոլյուցիան։ Էվոլյուցիոնիստների մեջ առաջիններից մեկն էր, որն աշխատել է հիմնավորել այն օրինաչափությունը, որ բազմաթիվ սերունդների ընթացքում օրգանիզմների բնույթի մեջ կուտակվող էվոլյուցիոն փոփոխությունները վերածվում են ամբողջ կազմվածքի խիստ թռիչքաձև փոփոխությունների, որոնք տեղի են ունենում էմբրիոգենեզի մեջ, որի հետևանքով էլ երևան են գալիս կենդանիների նոր դասեր։ 1830 թ. բռնվեց հայտնի բանավեճի Կյուվյեի հետ, որը ժխտում էր օրգանիզմների էվոլյուցիան։ Այդ բանավեճը տեղի է ունեցել Ֆրանսիական գիտությունների ակադեմիայում և տևել է մի քանի տարի։ Է. Ժոֆֆրուայի գործունեության արդյունքները ամփոփված են նրա կենսագրության մեջ, որը կազմել է նրա որդի Իսիդորը։
- ↑ 4,0 4,1 4,2 Ուելս Վ.Ս.
- ↑ Ռոուլեյ։
- ↑ Բրես (Brace Charles-Loring)։— Գրող, Ամերիկայի Միացյալ Նահանգներում (1826—1890)։
- ↑ 7,0 7,1 7,2 7,3 7,4 7,5 7,6 Հերբերտ Վիլիամ (Herbert William)։— Մանչեստերի ավագ քահանա և միաժամանակ հայտնի պարտիզագործ (1778—1847)։ Ծաղկատու կոճղեզավոր բույսերի սիրահար, հեղինակ ամարիլիսներին մշակության նվիրված գրքի։ Հիմնական հիպոթեզն այն է, որ բիոլոգիական միավոր հանդիսանում է սեռը, իսկ տեսակները միայն սեռերի տարատեսակներն են, որ ավելի «բարձր կարգեր» են, քան տեսակների տարատեսակները, և ավելի կայուն։
- ↑ Գրանտ (Grant James-Augustus)։— Ճանապարհորդ Շոտլանդիայում (1827—1892)։ Մասնակցել է Արևելյան Արևադարձային Աֆրիկայի ուսումնասիրության մի շարք գիտարշավների։
- ↑ 9,0 9,1 Պատրիկ Մատյու (Patrik Matthew)։— Ծագումով Շոտլանդիայից (1790—1874)։ Հեղինակ «Նավաշինարարական անտառանյութ և ծառաբուծություն» գրքի (1831), որի մեջ Դարվինի տեսակետների նման մտքեր է հայտնել բնական ընտրությամբ տեսակների ծագման մասին։
- ↑ 10,00 10,01 10,02 10,03 10,04 10,05 10,06 10,07 10,08 10,09 10,10 10,11 10,12 10,13 10,14 Ուոլլես (Wallace Alfred-Russel)։— Հռչակավոր բնախույզ, մալթուսական և միստիկ Անգլիայում (1823—1913)։ Դարվինի հետ միաժամանակ եկել է այն եզրակացության, որ օրգանիզմների էվոլյուցիան պետք է դիտել որպես բնական ընտրության հետևանք, որը տեղի է ունենում շնորհիվ այն հանգամանքի, որ օրգանիզմները ձգտում են անսահմանորեն շեղվել նախասկզբնական տիպից։ Ուոլլեսը գրել է մի գիրք դարվինիզմի մասին, ինչպես նաև կենդանիների աշխարհագրական տարածման մասին։
- ↑ Բուխ Լեոպոլդ (Buch Leopold)։— Հայտնի երկրաբան Գերմանիայում։ (1774—1853)։ Ապացուցել է հրաբխային գործունեության նշանակությունը մայր ցամաքների ձևավորման գործում։ Հեղինակ Եվրոպայի զանազան մասերի, այդ թվում Կանարյան երկրաբանության վերաբերյալ աշխատության (1825)։ Հրատարակել է նաև մի շարք բուսաբանա-աշխարհագրական արժեքավոր աշխատություններ։ Տրանսֆորմիստ, կողմնակից է այն տեսության, որ օրգանիզմների վրա բնակավայրի փոփոխվող պայմանների և երկրի կլիմայի խիստ փոփոխությունների ազդեցության հետևանքով տեսակները առաջացել են մեկը մյուսից։
- ↑ Ռաֆինեսկ (Rafinesque Smaltz Samuel)։— Բուսաբան ԱՄՆ-ում (1783—1840)։ Գրել է «Հյուսիսային Ամերիկայի նոր բուսականությունը»։
- ↑ Հոլդման (Haldeman Samuel-Stehman)։— Բնախույզ և ֆիզիոլոգ (1812—1880)։ Պրոֆեսոր Պենսիլվանիայի համալսարանում։ Գրել է մի շարք աշխատություններ կենդանաբանության վերաբերյալ, նրանցից առանձնապես հայտնի են կակղամորթների և միջատների վերաբերյալ աշխատությունները։
- ↑ Օմալիուս դը Ալլուան Ժ.։
- ↑ 15,00 15,01 15,02 15,03 15,04 15,05 15,06 15,07 15,08 15,09 15,10 15,11 15,12 15,13 15,14 15,15 15,16 15,17 15,18 15,19 15,20 Օուեն։
- ↑ 16,0 16,1 16,2 16,3 16,4 16,5 16,6 Ժոֆֆրուա Սենտ-Իլեր Իսիդոր (Geoffrey Saint Hilaire Isidor)։— Կենդանաբան Ֆրանսիայում (1805—1861), Է. Ժոֆֆրուայի որդին։ Եղել է Փարիզի բուսաբանական այգու կաթնասունների բաժանմունքի վարիչ, 1854 թվականին հիմնել է Փարիզի կլիմայավարժեցման ընկերությունը։ Գրել է կենդանաբանության վերաբերյալ բազմաթիվ աշխատություններ, այդ թվում «Տրակտատ մարդու և կենդանիների տերատոլոգիայի մասին» (1832—1836)։ Հեղինակ «Տեսակների սահմանափակ փոփոխելիության» տեսության։
- ↑ Ֆրեկ (Freke John)։— Բժիշկ, անատոմ և գրող (1688—1756)։
- ↑ 18,0 18,1 18,2 18,3 18,4 Սպենսեր Հերբերտ (Spencer Herbert)։— Հայտնի փիլիսոփա, բուրժուական պոզիտիվիստ Անգլիայում (1820—1903)։ Իր փիլիսոփայական երկերում մինչև Դարվինը զարգացրել է ամենահարմարվածի գերապրելու միջոցով բնության մեջ էվոլյուցիայի միտքը։ Ողջունել է Դարվինի «Տեսակների ծագման» լույս տեսնելը և դարձել է նրա ջերմ կողմնակից։ Սոցիոլոգիայում փորձել է հասարակական կյանքի երևույթները քննարկել, հենվելով բնության երևույթների մալթուսական խեղաթյուրված մեկնաբանության վրա։ Այսպես կոչված «սոցիալ-դարվինիստներից» մեկն է, սոցիալիզմի թշնամի։
- ↑ 19,0 19,1 19,2 19,3 Նոդեն (Naudin Ch.)։— Բուսաբան Ֆրանսիայում (1858—1899)։ Սկզբում ասիստենտ Musée d’histoire naturalle-ու՝ Փարիզում, հետո դիրեկտոր Villa Thuret ակկլիմատիզացիայի այգու՝ Անտիբի մոտ, Ֆրանսիայի հարավում։ Հիմնական աշխատություններ ունի դդմազգիների ընտանիքի մասին։ Կողմնակից տեսակների ծագման ընտրության միջոցով, որը սակայն ներգործում է վերջնական պատճառների շնորհիվ։
- ↑ Կեյզերլինգ (Keyserling Alexander)։— Բնախույզ Էստոնիայում (1815—1891)։ Մասնակցել է Ռուսաստանում կատարված մի շարք գիտարշավների (Ուրալ, Պեչորա)։ Հրատարակել է իր ճանապարհորդությունների նկարագրությունները, մասնակցել է «Եվրոպայի ողնաշարավորները» կոլեկտիվ աշխատությունը կազմելուն։
- ↑ Շաֆհաուզեն (Schaffhausen Hermann)։—Մարդաբան Գերմանիայում (1816—1893)։
- ↑ Լեկոկ (Lecoq Henri)։— Բնախույզ Ֆրանսիայում (1802—1871)։ Բնապատմության պրոֆեսոր, Բուսաբանական այգու դիրեկտոր Կլերմոնում։ Գրել է բնագիտության զանազան ճյուղերի՝ բուսաբանության, քիմիայի, երկրաբանության և գյուղատնտեսության վերաբերյալ բազմաթիվ աշխատություններ։
- ↑ Բադեն-Պաուել (Baden-Powell)։— Գիտնական Անգլիայում (1796—1860)։ Հեղինակ «Փորձ աշխարհների միասնության մասին» աշխատության, որը հրատարակվել է 1855 թ։
- ↑ Հերշել (Herschel John-Frederic-William)։— Աստղաբաշխ և ֆիզիկոս Անգլիայում (1792—1871)։
- ↑ 25,0 25,1 25,2 25,3 Բեր Կարլ-Էրնեստ (Baer von Karl-Ernest)։— Հայտնի կենդանաբան և սաղմնաբան Ռուսաստանում (1792—1876)։ Իդեալիստ, օրգանիզմների ծագման միասնականության տեսության հակառակորդ։ Պետերբուրգի Գիտությունների ակադեմիայի իսկական անդամ։ Գրել է բազմաթիվ աշխատություններ սաղմնաբանության բնագավառից, առաջադրել է հայտնի «Բերի օրենքը»՝ ի հակակշիռ մատերիալիստ-էվոլյուցիոնիստների «զարգացման բիոգենետիկական օրենքին»։
- ↑ Վագներ Ռուդոլֆ (Wagner Rudolph)։— Ֆիզիոլոգ Գերմանիայում (1805—1864)։ Ֆիզիոլոգիայի պրոֆեսոր Գետտինգենում։ Իր կյանքի վերջին շրջանում զբաղվել է մարդաբանությամբ և հրատարակել է Բերի հետ միասին մարդկանց զանազան գանգերի առաջացման մասին մի աշխատություն։
- ↑ 27,0 27,1 27,2 27,3 27,4 27,5 Հյոքսլի (Huxley Thomas-Henry)։— Կենդանաբան Անգլիայում (1825—1895)։ Սկզբում բժիշկ անգլիական նավատորմումդ իսկ հետո կենդանաբանության պրոֆեսոր, Դարվինի «Տեսակների ծագումը» լույս տեսնելուց հետո դարձել է նրա ջերմ կողմնակիցը և դարվինիզմի ժողովրդականացնողն Անգլիայում, կատարելով այդ տեսակետից նույն դերը, ինչ որ Հեկկելը Գերմանիայում։
- ↑ 28,00 28,01 28,02 28,03 28,04 28,05 28,06 28,07 28,08 28,09 28,10 28,11 28,12 28,13 28,14 28,15 28,16 28,17 28,18 28,19 28,20 28,21 28,22 28,23 Հուկեր Ժոզեֆ-Դալտոն (Hooker Joseph-Dalton)։ Հայտնի բուսաբան Անգլիայում (1817—1911), Բուսաբանական այգու դիրեկտոր Լոնդոնի մոտ, Կյու քաղաքում։ Նկարագրել է երկրագնդի մեծ մարզերի բուսական աշխարհները և հիմք է դրել նրանց զարգացման պատմության ուսումնասիրության։ Եղել է Դարվինի բարեկամը, մշակել է նրա «Բիգլ» նավով կատարած ճանապարհորդության ժամանակ Գալապագոս կղզիների վրա հավաքած բույսերի հավաքածուները և երկրաբան Լյաելի հետ (տես) 1858 թվականին ներկայացրել է Լոնդոնի Լիննեյան ընկերությանը՝ «Դարվինի չհրատարակած աշխատությունից մի քաղվածք տեսակի մասին» և տեսակների ծագման մասին ամերիկացի բուսաբան Ազա-Գրեյին 1857 թ. Դարվինի գրած նամակից մի հատված, որն ապացուցում է նրա առաջնությունը այդ տեսության ձևակերպման մեջ։
- ↑ 29,00 29,01 29,02 29,03 29,04 29,05 29,06 29,07 29,08 29,09 29,10 29,11 29,12 29,13 29,14 29,15 29,16 29,17 29,18 29,19 29,20 29,21 Լյաել (Lyell Charles)։— Հռչակավոր երկրաբան Անգլիայում (1797—1875)։ Գրել է «Երկրաբանության հիմունքները» աշխատությունը (1833), որը հսկայական դեր է խաղացել ոչ միայն երկրաբանության, այլև կենդանաբանության, մասնավորապես էվոլուցիոն ուսմունքի հիմնավորման գործում։ Այդ գիրքը ուժեղ ազդեցություն է գործել Դարվինի տեսակետների վրա և հնարավորություն է տվել նրան հիմնավորելու տեսակների փոփոխությունը բնական ընտրության միջոցով։ Լյաելը Հուկերի հետ միասին 1838 թ. Լոնդոնի Լիննեյան ընկերությանն է ներկայացրել Դարվինի «Տեսակների ծագումը» աշխատությունից մի քաղվածք։
- ↑ 30,0 30,1 Մալթուս (Malthus Thomas-Robert)։— Հայտնի բուրժուական վուլգար տնտեսագետ Անգլիայում (1766—1834)։ Գրել է «Փորձ ազգաբնակչության սկզբունքի մասին» (1798 թ.)։ Աշխատավոր դասակարգի աղետները բացատրել է իբրև թե բնության հավիտենական օրենքներով, նրանով, որ մարդիկ իբրև թե բազմանում են երկրաչափական պրոգրեսիայով, մինչդեռ ապրուստի միջոցները աճում են միայն թվաբանական պրոգրեսիայով։ Ֆեոդալական և բուրժուական դասակարգերի պաշտպան և պրոլետարիատի թշնամի։ Իմպերիալիզմի և ռասիզմի ամենախաբեբա գաղափարախոսներից մեկը, ռասաների միջև տոտալ պատերազմների անխուսափելիության գաղափարի պրոպագանդող։ Դարվինի կողմից Մալթուսի սխեման կիրառելը կոպիտ սխալ է։
- ↑ 31,0 31,1 31,2 Նայտ Անդրյու (Knight Thomas-Andrew)։— Բուսաբան և այգեգործ Անգլիայում (1759—1838)։ Հիմնադիր այգեգործների ընկերության Լոնդոնում, որի նախագահն է եղել 1811 թվականից սկսած մինչև իր մահը։ Գրել է բազմաթիվ աշխատություններ այգեգործության և բույսերի ֆիզիոլոգիայի վերաբերյալ։
- ↑ 32,0 32,1 Վեյսման Ավգուստ (Weismann August)։— Կենդանաբանության պրոֆեսոր Ֆրեյբուրգում՝ Գերմանիայում (1834—1915)։ Գրել է մի շարք աշխատություններ կենդանաբանության և ընդհանուր կենսաբանության վերաբերյալ։ Հայտնի է որպես մետաֆիզիկ, մեխանիստական իդեալիստ, որը ի հակակշիռ դարվինյան բնական ընտրության, առաջադրել է «սաղմնային ընտրության տեսությունը» և «սաղմնային պլազմայի անմահությունը», որը կախված չէ օրգանիզմի մարմնի վրա արտաքին միջավայրի պայմանների ազդեցությունից։ Հիմնադիր, այսպես կոչված, «նեոդարվինիզմի», համառ հակառակորդ մատերիալիստական այն դրույթի, որի համաձայն արտաքին պայմանների ազդեցության տակ ձեռք բերված հատկությունները ժառանգվում են։ Ա. Վեյսմանի ժառանգականության ավտոգենետիկ ռեակցիոն իդեալիստական «տեսությունը» կազմում է այժմյան նեոդարվինիզմի-մորգանիզմի հիմքը։ Վեյսմանիզմը վերջնականապես մերկացվել ու դատապարտվել է Վ. Ի. Լենինի անվան Գյուղատնտեսական գիտությունների ակադեմիայի օգոստոսյան սեսիայում 1948 թվականին։
- ↑ 33,00 33,01 33,02 33,03 33,04 33,05 33,06 33,07 33,08 33,09 33,10 33,11 Կյոլռեյտեր Իոզեֆ-Գոտլիբ (Koelreuter Joseph-Gottlieb)։— Բնախույզ, առավելապես բուսաբան Գերմանիայում (1733—֊1806)։ 1756-1761 թ. Պետերբուրգում Գիտությունների ակադեմիայի բնապատմության ադյունկտ։ Իր ամբողջ կյանքում աշխատանքի նպատակն է եղել բույսերի հիբրիդային ժառանգականության օրինաչափությունների ուսումնասիրությունը։ Կատարել է էքսպերիմենտալ դրվածքի տեսակետից ուշագրավ փորձեր բույսերի հիբրիդներ ստանալու վերաբերյալ և մատնանշել է հիբրիդների օգտագործումը գործնական նպատակների համար։ Գրել է մի շարք աշխատություններ բուսաբանության և կենդանաբանության վերաբերյալ։
- ↑ Տետ (Tait)։
- ↑ Հոյզինգեր (Heusinger Karl-Friedrich)։— Բնախույզ Գերմանիայում (1792—1883)։ Համեմատական անատոմիայի հիմնադիրներից մեկը։ Անատոմիայի և ֆիզիոլոգիայի պրոֆեսոր Մարբուրգում։
- ↑ 36,0 36,1 Վիման (Wyman)։
- ↑ 37,0 37,1 Լյուկ (Lucas Prosper)։— Բժիշկ Ֆրանսիայում (ծնվ. 1805 թ.)։ Գրել է բժշկականության և ֆիզիոլոգիայի վերաբերյալ մի շարք աշխատություններ, որոնց մեջ իր ժամանակին հայտնի է եղել «Առողջ և հիվանդ վիճակում գտնվող նյարդային համակարգության բնական ժառանգականության փիլիսոփայության և ֆիզիոլոգիայի վերաբերյալ տրակտատ»։
- ↑ 38,0 38,1 Հեեր Օսվալդ (Heer Oswald)։— Հայտնի հնաբուսաբան Շվեյցարիայում (1809—1883)։ Բուսաբանության պրոֆեսոր Ցյուրիխում։ Նրա բազմաթիվ աշխատություններից ամենից հայտնի են արկտիկական մարզի երրորդային ժամանակաշրջանի բրածո բուսականության և Շվեյցարիայի ցցաշենքերում գտնված բույսերի ուսումնասիրությունները։ Հեղինակ տեսակների կրեացիոնիստական տեսության, որի էությունը հանգում է այն բանին, որ տեսակները երկարատև գոյություն ունեն անփոփոխ, մինչև որ վրա են հասնում հանկարծակի մուտացիոն ձևափոխության ժամանակաշրջանները և նրանք այլ տեսակներ են դառնում «ստեղծագործական ուժի» ներգործությամբ։
- ↑ 39,0 39,1 39,2 39,3 39,4 Բլիտ (Blyth Edward)։— Կենդանաբան Անգլիայում (1810—1873)։
- ↑ 40,0 40,1 Ռյուտիմեյեր (Rütimeyer Ludwig)։— Բնախույզ Շվեյցարիայում (1825—1895)։ Գրել է մի շարք աշխատություններ հնէաբանության, կենդանաբանության և զոոտեխնիկայի վերաբերյալ։
- ↑ Սեբրայտ (Sebright John)։— Սելեկցիոներ Անգլիայում։
- ↑ Էլլիոտ (Elliot Walter)։— Հնէաբան Հնդկաստանում (1803—1887)։
- ↑ 43,0 43,1 Մուրրեյ Կ. (Murray Ch.)։
- ↑ Ստրիկլենդ։
- ↑ Լեպսիուս (Lepsius Karl-Richard)։— Արևելագետ և եգիպտաբան Գերմանիայում (1810—1884)։ 1842 թվականին ճանապարհորդություն է կատարել դեպի Եգիպտոս, որը տևել է չորս տարի։ Նրա գլխավոր աշխատությունները նվիրված են Եգիպտոսին և պարունակում են պատմական, աշխարհագրական, ազգագրական և հնագիտական տվյալներ, որոնք նա հավաքել է իր ճանապարհորդության ժամանակ։
- ↑ Բիրչ Սամուիլ (Birch Samuel)։— Եգիպտաբան Անգլիայում (1813—1885)։ Վարել է Բրիտանական թանգարանում Եգիպտոսի և Ասորեստանի բաժինները։
- ↑ 47,0 47,1 47,2 Պլինիուս (Caius Plinius secundus)։— Հռչակավոր բնախույզ հին Հռոմում (մեր թվականության 23—79 թթ.)։ Գրել է 37 հատորից բաղկացած «Բնապատմություն», որտեղ հավաքել է բնագիտության բոլոր բնագավառներին վերաբերող իրեն ժամանակակից բոլոր տեղեկությունները։
- ↑ Ակբար-Խան (Akbar Khan)։— Մոնղոլների միապետ (մեռել է 1605 թ.), Լենկթեմուրի տոհմից։ Նրա թագավորությունն ընդգրկում էր Հնդկաստանի հսկայական տերիտորիան։
- ↑ Վան-Մոնս (Van Mons J. B.)։— Պտղաբույծ Բելգիայում (1765—1842)։ Ստեղծել է Բրյուսելում 50000 ծառից բաղկացած մի փորձնական պտղատու այգի։ Հեղինակ պտղաբուծության բարելավման տեսության, որը շարադրել է մի շարք աշխատություններում, որոնցից առանձին հռչակ է վայելել «Պտղատու ծառեր» աշխատությունը (1835)։
- ↑ 50,0 50,1 50,2 Յուատ (Youatt)։— Հայտնի անասնաբույծ Անգլիայում։
- ↑ Սոմերվիլ (Somerville)։
- ↑ Լիվինգստոն (Livingstone David)։— Հռչակավոր ճանապարհորդ և քարոզիչ Անգլիայում (1813-1883)։
- ↑ 53,0 53,1 Բաքուել Ռոբերտ (Bakewell Robert)։— Հայտնի անասնաբուծարանատեր Անգլիայում (1725—1795)։
- ↑ Կոլլինզ (Kollings Charles)։— Եղջերավոր անասունների սելեկցիոներ Անգլիայում (1750—1836)։ Իր եղբայր Ռոբերտի (1749—1820) հետ միասին առաջ է բերել տավարի կարճաեղջյուր շորտհորնների ցեղը, որը համաշխարհային հռչակ է ստացել։
- ↑ Բորրոու Գեորգ (Borrow George)։— Գրականագետ և ճանապարհորդ Անգլիայում (1803—1881)։ Կատարել է բազմաթիվ ճանապարհորդություններ Ասիայում, Աֆրիկայում և Հարավային Եվրոպայում, առանձնապես Իսպանիայում, որոնց նվիրել է նկարագրություններ մի շարք հատորներում։
- ↑ Սպենսեր (Spenser)։ — Լորդ Անգլիայում։
- ↑ Բեքլեյ (Buckley)։
- ↑ Բերգեսս (Burgess)։
- ↑ 59,0 59,1 Մարշալ (Marshall John)։— Ֆիզիոլոգ և անատոմ Անգլիայում (1818—1891)։ Անատոմիայի պրոֆեսոր Royal Academy-ում։ Գրել է իր ժամանակին հայտնի ֆիզիոլոգիայի երկհատոր ձեռնարկը։
- ↑ 60,0 60,1 60,2 60,3 60,4 60,5 60,6 Լեբբոկ Ջոն (Lubbock John Baron Avebury)։— Բնախույզ Անգլիայում (1834—1813)։ Հայտնի է բնագիտության բնագավառում իր բազմաթիվ գիտական և հանրամատչելի աշխատություններով, որոնց մեջ նա տվել է Դարվինի տեսության ճշտությունը հաստատող մի շարք հիմնավորումներ։ Դրանց թվում կան երկասիրություններ միջատների ծագման և կերպարանափոխության մասին, ծաղիկների, պտուղների և տերևների կենսաբանության մասին, կենդանիների բնազդների մասին և այլն։
- ↑ 61,00 61,01 61,02 61,03 61,04 61,05 61,06 61,07 61,08 61,09 61,10 61,11 61,12 61,13 61,14 61,15 61,16 Մյուլլեր Ֆրից (Müller Fritz)։— Հայտնի կենդանաբան Բրազիլիայում (1821—1897)։ Ծնվել է Գերմանիայում, կրեացիոնիզմի կատաղի հակառակորդ։ Վտարանդվել է Բրազիլիա, որտեղ հետո ընտրվել է պրոֆեսոր Դեստերրոյում։ Առաջին դարվինիստներից մեկը։ 1864 հրատարակել է «Դարվինի օգտին» գիրքը, որի մեջ առաջին անգամ հաստատել է անցյալի ֆիլոգենետիկական կրկնությունը խեցեմորթների սաղմնային զարգացման մեջ (կենսաբանական հիմնական օրենքը) և ռեկապիտուլացիայի խանգարման պատճառները։ Ուսումնասիրել է խեցգետինների, թիթեռների, մեղուների, մրջյունների զարգացումը, միջատների միջոցով ծաղիկների փոշոտման հարմարումները և հետազոտել է միմիկրիայի երևույթը։
- ↑ 62,0 62,1 62,2 62,3 62,4 62,5 62,6 62,7 62,8 62,9 Ուոտսոն (Watson Hewett-Cottrel)։— Բուսաբան Անգլիայոմ (1804—1881)։
- ↑ Բաբինգտոն Չարլզ (Babington Charles-Cardale)։— Բուսաբան Անգլիայում (1808—1895)։ Հեղինակ մի շարք բուսաբանական աշխատությունների, այդ թվում Անգլիական նեղուցի (Channel Islands) կղզիների բուսականության մասին։
- ↑ 64,0 64,1 Բենտամ Գեորգ (Bentham George)։— Հայտնի բուսաբան Անգլիայում (1800—1884)։ Հեղինակ է բույսերի սիստեմատիկային վերաբերող բազմաթիվ աշխատությունների, որոնցից գլխավորներն են «Ավստրալիայի բուսականությունը» 7 հատորում, և «Genera plantarum» աշխատությունը, որ նա գրել է Հուկեր Ժոզեֆի հետ միասին։
- ↑ 65,0 65,1 65,2 65,3 65,4 65,5 65,6 65,7 Ուոլլաստոն (Wollastone William Hyde)։— Քիմիկոս և բնափիլիսոփա Անգլիայում (1766—1828)։ Նրա աշխատությունները հիմնականում քիմիական բնույթ ունեն, բայց նրանց մեջ շոշափվում են նաև հարցեր, որոնք վերաբերում են այլ դիսցիպլինների շարքին՝ օպտիկային, ակուստիկային, հանքաբանության, աստղաբաշխության, ֆիզիոլոգիային, բուսաբանության և այլն։
- ↑ Ուելշ (Walch B. D.)։— Միջատաբան ԱՄՆ-ում (1808—1869)։
- ↑ 67,00 67,01 67,02 67,03 67,04 67,05 67,06 67,07 67,08 67,09 67,10 67,11 67,12 67,13 67,14 67,15 67,16 67,17 67,18 Դը-Կանդոլ Ալֆոնս (De Candolle Alphonse)։— Հռչակավոր բուսաբան Շվեյցարիայում (1810—1893)։ Բուսաբանության պրոֆեսոր Ժնևում։ Շարունակել է հրատարակել «Բուսական թագավորության պրոդրոմուսը», որ սկսել էր նրա հայրը՝ Ավգուստ Պիրամ դը Կանդոլը, որին Դարվինն անվանում է «Ավագ դը-Կանդոլ»։ Գլխավոր աշխատություններն են «Բուսաբանական աշխարհագրությունը», որի մեջ երկրաբանական անցյալի հիման վրա հաստատել է բույսերի տարածման օրենքները, և «Կուլտուրական բույսերի ծագումը»։
- ↑ 68,0 68,1 68,2 68,3 68,4 68,5 68,6 68,7 68,8 68,9 Գրեյ-Ազա (Gray Asa)։— Հայտնի բուսաբան ԱՄՆ-ում (1810-1888)։ 1842 թվականից բնապատմության պրոֆեսոր Քեմբրիջում (ԱՄՆ)։ Հեղինակ բազմաթիվ բուսաբանական աշխատությունների, այդ թվում «Հյուսիսային Ամերիկայի բուսականության»։ Հաստատել է Հյուսիսային Ամերիկայի և Արևելյան Ասիայի բուսականությունների միջև եղած կապը, ինչպես նաև այն ազդեցությունը, որ ունեցել է սառցադաշտային ժամանակաշրջանը բույսերի տարածման վրա։
- ↑ դե-Սապորտ Գաստոն
- ↑ 70,0 70,1 Ֆրիս Էլիաս (Fries Elljas)։—Հայտնի բուսաբան Շվեդիայում (ծնվ. 1794 թ.)։ 1819 թվականից սկսած բուսաբանության պրոֆեսոր Լունդում, իսկ 1853 թվականից բուսաբանանական այգու դիրեկտոր Ուպսալայում։ Գրել է բազմաթիվ բուսաբանական աշխատություններ։
- ↑ 71,0 71,1 71,2 Ուեստվուդ (Westwood John-Obadiah)։— Միջատաբան և հնաշխարհագրագետ Անգլիայում (1805—1893)։
- ↑ 72,0 72,1 72,2 72,3 Լիննեյ Կարլ (Lìnné Carl)։— Հռչակավոր բնագետ Շվեդիայում (1707—1778)։ Հեղինակ բույսերի ու կենդանիների տեսակների դասակարգման ամենակատարյալ արհեստական սիստեմի։ Հետևողականորեն կիրառել է բինար անվանակարգությունը, ըստ որի յուրաքանչյուր կենդանի և բույս ստանում է կրկնակի՝ սեռային և տեսակային լատիներեն անուն։ Առաջին անգամ բիոլոգիական դասակարգման մեջ մտցրել է նաև մարդուն։ Հեղինակ տեսակառաջացման, այսպես կոչված, «հիբրիդագեն» տեսության, որը հիմնված է այն գաղափարի վրա, որ այժմյան դասերը, սեռերը և տեսակները ծագել են ինքնակամ հիբրիդացման միջոցով։
- ↑ 73,0 73,1 73,2 73,3 73,4 73,5 73,6 Ֆոկոներ (Falconer Hugh)։— Բուսաբան, երկրաբան և հնէաբան Անգլիայում (1808—1865)։ 1830 թ. սկսած մինչև 1842 թ. և 1847 թվականից մինչև 1855 թվականն աշխատել է Հնդկաստանում։ Նշանակվել է բուսաբանական այգու դիրեկտոր 1832 թ. Սինգապուրում և 1848 թ. Կալկաթայում։ Նրա նախաձեռնությամբ Հնդկաստանում մշակության մեջ են մտցվել 1830 թ. թեյի թուփը Բենգալիայում, 1848 թ. խինինի ծառը։ Գրել է բազմաթիվ աշխատություններ, գլխավորապես հնէաբանության վերաբերյալ։
- ↑ 74,0 74,1 Ազարա (Asara Felix)։— Ճանապարհորդ և բնախույզ Իսպանիայում (1746—1811)։ Նրա գլխավոր աշխատությանն է «Ճանապարհորդություն դեպի Հարավային Ամերիկա» (1809)։
- ↑ Ռենգեր (Rengger Johann-Rudolf)։— Ճանապարհորդ և բնախույզ (1795—1832)։ Ճանապարհորդել է Հարավային Ամերիկայում, զբաղվել է կենդանիների՝ գլխավորապես կաթնասունների կյանքի վերաբերյալ դիտումներով։ Ուսումնասիրել է նաև ծովային կենդանիները Իտալիայում իր կատարած ճանապարհորդության ժամանակ (1831)։
- ↑ Նյուման (Newmann Edward)։— Միջատաբան Անգլիայում (ծնվ. 1801 թ.)։ Հատկապես ուսումնասիրել է գյուղատնտեսության համար օգտակար և վնասակար միջատները։
- ↑ Դոունինգ (Downing Andre Jackson)։— Այգեգործ ԱՄՆ-ում (1815—1852)։ Գրել է այգեգործության և պտղաբուծության վերաբերյալ մի շարք աշխատություններ, այդ թվում՝ «Ամերիկայի պտուղների և պտղատու ծառերի մասին»։
- ↑ 78,0 78,1 78,2 Ֆաբր (Fabre Henri)։— Հայտնի միջատաբան Ֆրանսիայում (1823—1915)։ Մասնագիտացած էր միջատների բնազդների և հակումների ուսումնասիրության գործում, հրատարակել է այդ բնագավառին վերաբերող մի շարք լայն հռչակված աշխատություններ։
- ↑ Հիրոն (Heron Robert)։— Գիտնական և գրող Շոտլանդիայում (1764—1807)։
- ↑ Պիրս (Pierce)։
- ↑ Գրաբա (Graba)։
- ↑ 82,0 82,1 82,2 Շպրենգել' (Sprengel Christian-Konrad)։— Մանկավարժ Գերմանիայում (1750—1816)։ Գրել է «Բնության բացված գաղտնիքը ծաղիկների կառուցվածքի և բեղմնավորության մեջ» հռչակավոր աշխատությունը, որի մեջ բերել է միջատների միջոցով փոշոտվող 300-ից ավելի բույսերի վրա կատարած դիտումները և դրանով ապացուցել է, որ միջատների միջոցով ծաղկափոշին տեղափոխվում է մի բույսից մյուսի վրա։ Այդ աշխատությունը նրա ժամանակակիցների կողմից բոլորովին աննկատելի էր մնացել և գնահատության է արժանացել միայն Դարվինի շնորհիվ։
- ↑ 83,00 83,01 83,02 83,03 83,04 83,05 83,06 83,07 83,08 83,09 83,10 83,11 83,12 83,13 83,14 83,15 83,16 83,17 83,18 83,19 83,20 Գերտներ (Gärtner Karl-Friedrich)։— Հայտնի բուսաբան Գերմանիայում (1772—1850)։ Գրել է «Փորձեր և դիտողություններ հիբրիդների առաջացման վրա» կապիտալ աշխատությունը, որը վերջնականապես հաստատել է սեռի գոյությունը բույսերի մոտ։
- ↑ 84,0 84,1 Հիլդեբրանդ (Hildebrand Friedridh)։— Բուսաբան Գերմանիայում (1835—1915)։ Պրոֆեսոր և Բուսաբանական այգու դիրեկտոր Ֆրեյբուրգում։ Գրել է մի շարք բուսաբանական աշխատություններ, մասնավորապես աշխատություններ, որոնց առարկան է եղել բույսերի սեռի և տարածման եղանակների ուսումնասիրությունը։
- ↑ 85,0 85,1 85,2 Վագներ Մորից (Wagner Moritz)։— Հայտնի բնախույզ Գերմանիայում (1813—1887)։ Կատարել է մի շարք գիտարշավներ դեպի Ալժիր, Հարավային Եվրոպա, այդ թվում նաև Ղրիմ, Կովկաս, Պարսկաստան, Կանադա, Կենտրոնական և Հարավային Ամերիկա։ Նրա աշխատություններից առանձնապես հայտնի է եղել «Դարվինի տեսությունը և օրգանիզմների միգրացիոն օրենքը», որը ներկայացնում է դարվինիզմի քննադատությունը «կյանքի հավիտենականություն» և «ստեղծագործական ուժի» տեսության դիրքերից (տե՛ս Էնգելս, «Բնության դիալեկտիկան»)։ Բնական ընտրության տեսության փոխարեն առաջադրել է «անջատման տեսությունը»։
- ↑ 86,0 86,1 86,2 86,3 86,4 86,5 Էդվարդս (Edwards)։
- ↑ 87,0 87,1 87,2 87,3 Ուոտերհաուզ (Waterhouse George-Robert)։— Բնախույզ Անգլիայում (1810—1888)։
- ↑ 88,00 88,01 88,02 88,03 88,04 88,05 88,06 88,07 88,08 88,09 88,10 88,11 88,12 Ֆորբս (Forbes Edward)։— Հռչակավոր բնախույզ Անգլիայում (1815—1854)։ Հեղինակ կապիտալ հետազոտությունների բնագիտության զանազան բնագավառներում, որոնցից բացառիկ նշանակություն ունի նրա «Բրիտանական կղզիների կենդանական և բուսական աշխարհների տարածման մեջ եղած փոխհարաբերությունների և այն փոփոխությունների մասին, որոնց ենթարկվել են նրանք երկրաբանական ժամանակաշրջանների և առանձնապես սառցադաշտային ժամանակաշրջանի ազդեցության տակ» հետազոտությունը։ Տրանսֆորմիստ է եղել։
- ↑ 89,0 89,1 89,2 Գուլդ (Gould John)։— Կենդանաբան Անգլիայում (1804—1881)։ Հեղինակ թռչունների և մասնավորապես Ավստրալիայի թռչունների կենդանաբանության վերաբերյալ մի շարք ուսումնասիրությունների։
- ↑ Մոկեն-Տանդոն (Moquin-Tandon Alfred-Horace-Benedict)։— Բուսաբան Ֆրանսիայում (1804—1863)։
- ↑ Կյոնինգհեմ (Cunningham Richard)։— Բուսաբան և ճանապարհորդ Անգլիայում (1793—1835)։ Բուսաբանական այգու դիրեկտոր Սիդնեյում։
- ↑ Կերբի Վիլիամ (Kirby William)։— Միջատաբան Անգլիայում (1759—1850)։ Նրա աշխատությունների թվում կա մի մենագրություն Անգլիայի մեղուների մասին և կենդանիների պատմության, սովորությունների և բնազդների մասին երկհատոր աշխատությունը։
- ↑ Բրոուն Սեկար (Brown-Séquard Charles-Edward)։— Հայտնի ֆիզիոլոգ և նյարդաբան Ֆրանսիայում (1817—1894)։ Աշխատել է նաև Ամերիկայում և Անգլիայում։ 1878 թ. նշանակվել է էքսպերիմենտալ բժշկականության պրոֆեսոր College de France-ում որպես Կլոդ Բերնարի հաջորդ։ Գրել է մոտ 500 աշխատություն ֆիզիոլոգիայի բնագավառից, այդ թվում նաև ներքին սեկրեցիայի մասին։ Հեղինակ լայնորեն հայտնի փորձերի, որոնք ապացուցում են օրգանիզմների ձեռք բերած հատկությունների ժառանգումը և նյարդային սիստեմի վճռական նշանակությունը ձեռք բերվածների ժառանգելիության գործում։
- ↑ Սիլլիման (Silliman Benjamin)։— Քիմիկոս և հանքաբան Ամերիկայում (1816—1885)։
- ↑ 95,0 95,1 95,2 Շիեդտե (Schiödte Jorgen-Christian)։— Նշանավոր միջատաբան Դանիայում (1815—1884)։ 1842 թվականին նշանակվել է միջատաբանական թանգարանի դիրեկտոր, 1854 թ. սկսած պրոֆեսոր։ Գրել է գերազանցապես հոդվածոտանիների մասին, առանձին ուշադրության է դարձրել կարծրաթև միջատների մորֆոլոգիային և կերպարանափոխություններին։
- ↑ 96,0 96,1 96,2 Դանա (Dana James-Dwight)։— Բնախույզ ԱՄՆ-ում (1822—1895)։ Բնագիտության պրոֆեսոր Իելի կոլեջում։ Մասնակցել է որպես երկրաբան և հանքաբան Վիլկիսի (Wilkes) դեպի Խաղաղ օվկիանոսը կատարած գիտարշավին, որի արդյունքները հրատարակել է երեք հատորով։
- ↑ 97,00 97,01 97,02 97,03 97,04 97,05 97,06 97,07 97,08 97,09 97,10 97,11 97,12 97,13 97,14 Ագասսիս (Agassis Louis)։— Հայտնի հնէաբան և երկրաբան, հակաէվոլյուցիոնիստ ԱՄՆ-ում (1807-1873)։ Ծագումով շվեյցարացի, բնագիտության պրոֆեսոր Նեվշատելում։ 1846 թ. տեղափոխվել է Ամերիկա, որտեղ եղել է երկրաբանության և կենդանաբանության պրոֆեսոր։ Կենդանաբանության, հնէաբանության և երկրաբանության (մասնավորապես սառցադաշտային ժամանակաշրջանի) վերաբերյալ բազմաթիվ աշխատությունների հեղինակ է։ Էվոլյուցիոն ուսմունքի և դարվինիզմի կատաղի հակառակորդ, տեսակների, սեռերի և այլնի աստվածային ծագման գաղափարի պաշտպան։
- ↑ Թվեյտս (Thwaites George-Henry-Hendrick)։— Բուսաբան և միջատաբան Անգլիայում (1811—1882)։
- ↑ Շլեգել (Schlegel Hermann)։— Հայտնի կենդանաբան Գերմանիայում (1804—1884)։
- ↑ 100,00 100,01 100,02 100,03 100,04 100,05 100,06 100,07 100,08 100,09 100,10 100,11 100,12 100,13 100,14 100,15 100,16 100,17 100,18 100,19 100,20 100,21 100,22 100,23 100,24 100,25 Մայվարտ (Mivart St. George-Jakson)։— Կենսաբան Անգլիայում (1827—1900)։ Գրել է մի շարք ինչպես մասնագիտական, այնպես էլ տեսական աշխատություններ կենսաբանության վերաբերյալ։ Հայտնի է որպես Դարվինի կատաղի հակառակորդ, կողմնակից տեսակառաջացման իդեալիստական «մուտացիոն» տեսության և ավտոգենեզի գաղափարի։ 1871 թվականին նրա լույս ընծայած «Տեսակի գենեզիսը» գիրքը առաջ է բերել կատաղի բանավեճ։ Այդ գրքում նա, ընդունելով տեսակի էվոլյուցիան, ժխտում էր այն մարդու ինտելեկտի նկատմամբ։ Ավելի ուշ իր «Մարդկային բանականության ծագումը» աշխատության մեջ նա մարդուն հակադրել է կենդանիներին։ Հետագայում բելգիական եպիսկոպոսարանի հրավերով նա մի շարք աշխատություններով դուրս է եկել ընդդեմ գիտության, որպես կրոնի դեմ կոնֆլիկտ առաջացնողի։
- ↑ Գյոթե (Goethe Iohann Wolfgang)։— Հռչակավոր բանաստեղծ և գրող Գերմանիայում (1749—1832)։ Հայտնի է նաև բույսերի մորֆոգիայի, համեմատական անատոմիայի և ֆիզիկայի բնագավառներում կատարած մի շարք գիտական ուսումնասիրություններով։ 18-րդ դարի վերջերին մտքեր է հայտնել հօգուտ օրգանիզմների մետամորֆոզի (կերպարանափոխության)։ Մետամորֆոզի ուսմունքի հիմքն է կազմում այն գաղափարը, որ կառուցվածքի իդեալականորեն գոյություն ունեցող նախասկզբնական տիպերը ձևափոխվում են նոր վարիանտների, որոնք չեն փոխում տիպի էությունը։
- ↑ Հունտեր (Hunter John)։— Վիրահատ և անատոմ Անգլիայում (1728—1794)։ Գրել է մի շարք աշխատություններ կենսաբանության և համեմատական անատոմիայի բնագավառին վերաբերող։ Իրենից հետո թողել է մի թանգարան 10000 էքսպոնատներից բաղկացած, որի վրա ծախսել է իր ամբողջ կարողությունը։
- ↑ Ուոլշ (Walsh)։
- ↑ 104,0 104,1 104,2 Պալլաս Պետր-Սիմոն (Pallas Peter-Simon)։— Հռչակավոր բնախույզ, էվոլյուցիոն տեսության ակտիվ հակառակորդ Ռուսաստանում (1741—1811), ծագումով գերմանացի։ Դուրս է եկել ընդդեմ Լիննեյի տեսության՝ հիբրիդացման միջոցով տեսակների ծագման մասին և ընդդեմ Բյուֆոնի տեսության՝ կլիմայի ազդեցության տակ տեսակների ձևափոխման մասին։ Ռուսաստան է եկել 1768 թ. որպես արդեն հայտնի կենդանաբան և նշանակվել է Պետերբուրգի Գիտությունների ակադեմիայի իսկական անդամ։ Իր կյանքի քառասուն տարին նվիրել է Ռուսաստանի հետազոտությանը։ Կատարել է երկու ճանապարհորդություն դեպի Սիբիր, որոնք տևել են 10 տարուց ավելի, ուսումնասիրել է Ղրիմը, որտեղ ապրել է 15 տարի։ Նրա ճանապարհորդությունների նկարագրությունները արժեքավոր տեղեկություններ են տվել Ռուսաստանի բնության մասին։ Պալլասի աշխատություններում եղած փաստական տվյալների հարստությունը վկայում է, որ նա բացառիկ գիտնական է իր գիտելիքներով և դիտողականությամբ։
- ↑ 105,0 105,1 105,2 Պուլ (Poole)։— Գնդապետ, ձիաբուծություն իմացող Անգլիայում։
- ↑ 106,0 106,1 Գրեյ Ջ. Է. (Gray John-Edward)։— Հայտնի կենդանաբան Անգլիայում (ծնվ. 1800 թ.)։ Գրել է կենդանաբանությունից բազմաթիվ աշխատություններ կաթնասունների, կակղամորթների և այլոց մասին։
- ↑ Սմիտ Համիլտոն (Smith Hamilton)։— Գնդապետ Անգլիայում։
- ↑ Ռոլլեն (Rollin)։— Բուսաբան Ֆրանսիայում։
- ↑ Գոսսե (Gosse Philip-Henry)։— Անգլիական բնախույզ (1810—1888)։ Ճանապարհորդ և հեղինակ մի շարք կենդանաբանական աշխատությունների, ողնաշարավորների, միջատների և ծովային պլանկտոնի մասին, «Օմֆալոս-երկրաբանական հանգույցը արձակելու մի փորձ» գրքում ծայրահեղ հետադիմական փորձով հանդես է եկել ապացուցելու, որ աշխարհի ստեղծագործության մասին աստվածաշնչի առասպելը ճիշտ է, այդ բոլորը պաշտպանելով ամենաանհեթեթ «փաստարկներով» ընդդեմ բնության վերաբերմամբ եղած պատմական տեսակետի։
- ↑ Մարտին Վիլիամ (Martin William)։— Բնախույզ Անգլիայում (1767—1810)։ Հեղինակ հնէաբանության վերաբերյալ մի շարք աշխատությունների։
- ↑ Մորտոն (Morton Samuel-Georg)։— Հռչակավոր բնախույզ Ամերիկայում (1799—1851)։ Բժիշկ։ Գրել է բազմաթիվ աշխատություններ բժշկականության, կենդանաբանության, հնէաբանության և մարդաբանության վերաբերյալ։
- ↑ 112,0 112,1 Ռիչարդսոն (Richardson John)։— Բնախույզ Անգլիայում (1787—1865)։ Որպես բժիշկ և բնախույզ մասնակցել է կապիտան Ֆրանկլինի դեպի Հյուսիսային բևեռը կատարած ճանապարհորդություններին։ 1848 թ. մասնակցել է անհետացած Ֆրանկլինին որոնելու համար կազմակերպված էքսպեդիցիային։ Նրան է պատկանում բևեռային ծովերում Ֆրանկլինի գիտարշավի գիտական արդյունքների նկարագրությունը։
- ↑ 113,0 113,1 Էյտոն (Eyton Thomas-Campbell)։— Բնախույզ Անգլիայում (1809—1880)։
- ↑ Հերն (Hearne Samuel)։— Ծովագնաց և ճանապարհորդ Անգլիայում (1745—1792)։ Հայտնի է Հյուսիսային Ամերիկայում՝ Հուդզոնի ծոցում կատարած իր հետազոտություններով։ Թողել է ճանապարհորդությունների նկարագրությունը։
- ↑ 115,0 115,1 Հուդզոն (Hudson William)։— Բնախույզ Անգլիայում (1730—1793)։
- ↑ Սոսյուր-դե (Saussure Horace-Benedict de)։— Հռչակավոր բնախույզ և ֆիզիկոս Շվեյցարիայում (1740—1799)։ Աշխատել է բնագիտության զանազան բնագավառներում, գլխավորապես ֆիզիկայի, օդերևութաբանության, բուսաբանության և երկրաբանության բնագավառներում։ Հիմնադիր բույսերի ֆիզիոլոգիայի։ Հետազոտել է Ալպերը, որոնց վրա կատարած ճանապարհորդություններին նվիրել է մի հատուկ աշխատություն։
- ↑ 117,0 117,1 117,2 Օդյուբուն' (Audubon)։
- ↑ Ժուրդեն (Jourdain Sylvain Hippolyte)։— Բնախույզ Ֆրանսիայում (ծնվ. 1832 թ.)։
- ↑ Վիրխով (Virchow Rudolph)։— Հայտնի բժիշկ և անատոմ, գիտության մեջ հետադիմական, քաղաքականության մեջ սոցիալիզմի թշնամի։ Հեղինակ տեսության օրգանիզմի մասին՝ որպես «ինքնուրույն բջիջների ֆեդերացիայի» և պրեֆորմիստական ուսմունքի, որ ամեն կենդանի բան միայն բջջից է»։ Այդ տեսությունը հերքված է Օ. Բ. Լեպեշինսկայայի աշխատանքներով։
- ↑ Լանդուա (Landois Leonard)։— Ֆիզիոլոգ Գերմանիայում (ծնվ. 1837 թ.)։ Գրել է ֆիզիոլոգիայի հայտնի դասագիրք, որը մի քանի անգամ է հրատարակվել և թարգմանված է շատ լեզուներով, այդ թվում նաև ռուսերեն։ Շատ է գրել զանազան պարազիտների անատոմիայի մասին։
- ↑ Կոպ (Cope Edvard-Drinker)։— Հնէաբան ԱՄՆ-ում (1840—1897)։ 1864—1867 թվականներին բնապատմության պրոֆեսոր Haverford College-ում, իսկ հետագայում երկրաբանության պրոֆեսոր Պենսիլվանիայի համալսարանում։ Նեոլամարկիստ իդեալիստական ուղղությամբ (այսպես կոչված «պսիխոլամարկիստ»), որը համարում է, որ կենդանի ձևերի առաջադիմական զարգացման հիմքը կազմում է «ճիգը», որը պայմանավորված է «գիտակցությամբ»։
Գրել է հնէաբանության և կենդանիների էվոլյուցիայի վերաբերյալ աշխատություններ։ - ↑ Մյուրի (Mühry Adalbert-Adolf)։— Կլիմայագետ Գերմանիայում (1810—1888)։ Ձեռնարկել է մի շարք ճանապարհորդություններ և հրատարակել է իր դիտումների արդյունքները բազմաթիվ հատուկ գրքերում։
- ↑ Մատտեուչչի (Matteucci Carlo)։— Ֆիզիկոս Իտալիայում (1811—1868)։ Հայտնի էր իր աշխատություններով կենդանիների մեջ կատարվող էլեկտրական երևույթների բնագավառում։
- ↑ Մակ-Դոննել (Мс-Donnell R.)։
- ↑ Ռադկլիֆ (Radcliffe John)։— Բժիշկ Անգլիայում (1650—1714)։
- ↑ Պաչինի (Pacini Philippo)— Անատոմ Իտալիայում (1812—1883), անատոմիայի և հյուսվածաբանության պրոֆեսոր Ֆլորենցիայում։ Գրել է այդ գիտությունների վերաբերյալ բազմաթիվ աշխատություններ։
- ↑ Հենզեն (Hensen Victor)։— Ֆիզիոլոգ Գերմանիայում (1835—1924)։ Ֆիզիողոգիայի պրոֆեսոր Քիլում։ Նրա հիմնական աշխատությունները վերաբերում են զգայարանների սաղմնաբանությանը, անատոմիային և ֆիզիոլոգիային։
- ↑ Կլապարեդ (Claparede Jean-Louis-Rene Antoine Edouard)։— Ականավոր կենդանաբան Շվեյցարիայում (1832—1870)։ Համեմատական անատոմիայի պրոֆեսոր Ժևում։ Դարվինի «Տեսակների ծագումը» լույս տեսնելուց հետո դարձել է նրա կողմնակիցներից մեկը և հրատարակել է մի շարք արժեքավոր աշխատություններ, որոնք հաստատում են նրա տեսակետները։
- ↑ Կրյուգեր (Cruger), Դոկտոր։
- ↑ Նատուզյուս (Nathusius Hermann Engelhardt)։— Բնախույզ Գերմանիայում (ծնվ. 1809 թ.)։ Զբաղվել է Անգլիայում եղջերավոր անասունների զանազան ցեղերի նատուրալիզացիայով։ Նրա տնտեսությունը դարձել էր Անգլիայում ցեղական անասունների ստեղծման կենտրոն։ Իր աշխատությունների և դիտումների արդյունքները նա շարադրել է մի շարք աշխատություններում, որոնց մեջ հանդես է եկել Դարվինի տված էվոլյուցիայի ընդհանուր սխեմայի կողմնակից։
- ↑ Պալեյ (Paley)։
- ↑ Հելմհոլց Հերման (Helmholtz Hermann)։— Ֆիզիոլոգ և ֆիզիկոս Գերմանիայում (1821—1894), ֆիզիոլոգիայի պրոֆեսոր Քյոնիկսբերգում, Բոննում և Հայդելբերգում։ Հայտնի է զգայարանների, հատկապես տեսողության զգայարանի իր հետազոտություններով։ Նրա երկասիրությունների մեջ գլխավորներից մեկն է «Ֆիզիոլոգիայի օպտիկան»։
- ↑ 133,0 133,1 133,2 133,3 133,4 133,5 Կյուվյե (Cuvier George Leopold Frederic-Dagobert)։— Հռչակավոր կենդանաբան և հնէաբան Ֆրանսիայում (1769—1832)։ Կրեացիոնիստ, կողմնակից «կատաստրոֆների տեսության» և այն գաղափարի, որի համաձայն տեսակները շատ անգամ են երևան եկել ոչ թե պատմական զարգացման մեջ, այլ «վերջնական պատճառների» հետևանքով։ Բացատրել է կենդանական աշխարհների հաջորդականությունը կատասարոֆաներից գերապրած «կյանքի անմահ սաղմերով»։ Օրգանիզմների ծագման միասնականության տեսության հակառակորդ։ Գրել է բազմաթիվ աշխատություններ կենդանաբանության և ողնաշարավորների, հատկապես կաթնասունների համեմատական անատոմիայի բնագավառի վերաբերյալ։ Գրել է «օրգանիզմների կառուցվածքի բացարձակ նպատակահարմարության և գոյության պայմանների» սկզբունքի մասին տեսությունը։
- ↑ 134,0 134,1 Լանկեստեր Ռեյ (Lankester Rau-Edwin)։— Կենդանաբան Անգլիայում (1847—1929)։ Բրիտանական թանգարանի բնապատմական բաժանմունքի դիրեկտոր, հիմնադիր Ծովային կենսաբանական միության Անգլիայում։ Գրել է մի շարք աշխատություններ համեմատական անատոմիայի վերաբերյալ և բազմաթիվ հանրամատչելի գրքեր ու հոդվածներ։
- ↑ 135,0 135,1 135,2 Լյուիս (Lewes George-Harris)։— Փիլիսոփա և գրականագետ Անգլիայում (1817—1878)։ Նպատակ ունենալով ժողովրդականացնել գիտության զանազան ճյուղերը, հրատարակել է մի շարք աշխատություններ, այդ թվում «Կենդանիների կյանքը», «Գիտության պատմությունը», «Կյանքի պրոբլեմները»։
- ↑ 136,0 136,1 136,2 136,3 136,4 136,5 Բրոնն Հենրիխ-Գեորգ (Bronn Hejnrich-Georg)։— Հնէաբան Գերմանիայում (1800—1862)։ Բնապատմության պրոֆեսոր Հայդելբերգում։ Հեղինակ է մի շարք հնէաբանական աշխատությունների։
- ↑ Բրոկա (Broca Paul)։— Վիրահատ Ֆրանսիայում (ծն. 1824 թ.)։ Իրեն ժամանակակից անտրոպոլոգների դպրոցի պարագլուխ։ Հեղինակ բժշկականության և անտրոպոլոգիայի վերաբերյալ մի շարք աշխատությունների։
- ↑ 138,0 138,1 138,2 Նեգելի (Naegeli Karl-Wilhelm)։— Հայտնի բուսաբան Գերմանիայում (1817—1891)։ Ծագումով շվեյցարացի։ Պրոֆեսոր Մյունխենում։ Հեղինակ «իդիոպլազմների» տեսության, որը հետագայում Վեյսմանը դրել է իր հակադարվինիստական տեսության հիմքում։ Կողմնակից «իմանենտ պրոգրեսի» (ավտոգենեզի) գաղափարի։ Նրա բազմաթիվ աշխատություններից ամենից ավելի հետաքրքրական է «Էվոլյուցիայի մեխանիկա-ֆիզիոլոգիական տեսությունը»։
- ↑ Շեբլ (Schoobl), դոկտոր։
- ↑ 140,0 140,1 Ժյուսյե (Jussien Antoine-Laurent de)։— Հռչակավոր բուսաբան Ֆրանսիայում (1748—1836)։ Բուսաբանության պրոֆեսոր և Բուսաբանական այգու դիրեկտոր Փարիզում։ Հեղինակ է հանդիսանում բուսական թագավորության առաջին բնական սիստեմի, որի հիմքը կազմում են այն սկզբունքները, որոնք մշակել է նրա հորեղբայր Բեռնար Ժյուսյեն։ Հիմնական աշխատությունն է «Cenera plantarum»։
- ↑ Շախտ (Schacht Hermann)։— Բուսաբան (1814—1864)։ Եղել է պրոֆեսոր Բոննում։ Գրել է շատ աշխատություններ, գլխավորապես բույսերի անատոմիայի վերաբերյալ։
- ↑ Տաուշ (Tausch Ignaz Friedrich)։— Բուսաբան-սիստեմատիկ (1792—1848)։ Եղել է բուսաբանության պրոֆեսոր Պրագայում։ Նրա հայտնի երկերից մեկում նկարագրված է Բոհեմիայի բուսականությունը։
- ↑ Բրաուն Ալեքսանդր (Braun Alexander)։— Հայտնի բուսաբան Գերմանիայում (1805—1877)։ Բուսաբանության պրոֆեսոր Գերմանիայի զանազան քաղաքներում, այդ թվում նաև Բեռլինում։ Հեղինակ է բազմաթիվ աշխատությունների, գլխավորապես բույսերի մորֆոլոգիայի և սիստեմատիկայի մասին, ստեղծել է բուսական աշխարհի բնական սիստեմներից մեկը։
- ↑ 144,0 144,1 Սենտ-Իլեր Օգյուստ (Saint-Hillare Auguste François César Prouvencal de)։— Բուսաբան Ֆրանսիայում (1799—1853)։ 1816—1822 և 1830 թվականներին կատարել է ճանապարհորդություններ Հարավային Ամերիկայում, Բրազիլիայում։ Իր ճանապարհորդությունների արդյունքները հրատարակել է բազմաթիվ գրքերով ու հոդվածներով, որոնցից առանձին հետաքրքրություն են ներկայացնում «Հարավային Բրազիլիայի բուսականությունը» (1825—1832) և «Բրազիլիայի օգտակար բույսերը» (1827—1828)։
- ↑ Մաստերս (Masters)։
- ↑ 146,0 146,1 Ռայտ Չոնսեյ (Wright Chauncey)։— Մաթեմատիկոս և փիլիսոփա ԱՄՆ-ում (1830—1875)։
- ↑ Բեկեր Սամուիլ (Beker Samuel-White)։— Ճանապարհորդ Անգլիայում (1821—1893)։ Հետազոտել է անհայտ շատ մարզեր, գլխավորապես Կենտրոնական Աֆրիկայում, այդ թվում և Նեղոսի ակունքները։ Թողել է իր ճանապարհորդությունների արժեքավոր նկարագրությունները։
- ↑ Դայակ (Դայակներ — ցեղի անուն)։
- ↑ Սկորեսբի (Scoresby William)։— Ծովագնաց և գիտնական Անգլիայում (1789—1857)։ Գրել է ձկնորսական նավերով Արկտիկական ծովերում իր ճանապարհորդությունների մի շարք նկարագրություններ, որոնց թվում նաև կետերի որսը Գրենլանդիայի ափերի մոտ։
- ↑ Սալվին (Salvin)։
- ↑ Բարտլետտ (Bartlett Elisha)։— Բժիշկ ԱՄՆ-ում (1804—1855)։ Բժշկականության պրոֆեսոր Նյու-Յորքում։
- ↑ Լասեպեդ (Lacopede)։— Ֆրանսիական կենդանաբան, Բյուֆֆոնի շարունակողը։ «Տեսակների սահմանափակ փոփոխելիության» տեսության կողմնակից։
- ↑ 153,0 153,1 Մալմ (Malm)։
- ↑ 154,0 154,1 154,2 154,3 154,4 154,5 Գյունտեր (Günther Albert-Karl-Ludwik-Gotthilf)։ Կենդանաբան Անգլիայում (1830—1914)։
- ↑ Տրակվեր (Traquair)։
- ↑ Յարրել (Yarrel)։— Բնախույզ Անգլիայում (1784—1856)։
- ↑ Պուշե (Pouchet Felix-Archimede)։— Գիտնական Ֆրանսիայում (1800—1872)։ Կրթությամբ բժիշկ, հետագայում աշխատել է բուսաբանության և կենդանաբանության բնագավառներում, հրատարակել է գիտության այդ ճյուղերի վերաբերյալ 80-ից ավելի աշխատություններ։ Բնագիտության պրոֆեսոր Ռուսաստանում։ Հայտնի է որպես Պասստյորի հակառակորդ օրգանիզմների ինքնածնության հարցերում և հեղինակ այն փորձերի, որոնք ապացուցում են ամենապարզ կենդանի մարմինների, այսպես կոչված, ինքնաբերական ծնունդն անկենդան օրգանական նյութերից։
- ↑ Բրեմ (Brehm Alfred)։— Կենդանաբան և ճանապարհորդ Գերմանիայում (1829—1884)։ Կենդանաբանական այգու դիրեկտոր Համբուրգում, Բեռլինի «Ակվարիումի» հիմնադիր։ Հեղինակ «Կենդանիների կյանքը» հայտնի աշխատության։
- ↑ Հենսլոու (Henslow John-Stevens)։— Բուսաբան և հանքաբան Անգլիայում (1796—1861), սկզբում հանքաբանության, իսկ հետո բուսաբանության պրոֆեսոր Քեմբրիջի համալսարանում։
- ↑ Լոկվուդ (Lockwood)։— Կենդանաբան ԱՄՆ-ում։
- ↑ 161,0 161,1 161,2 161,3 Ֆլոուեր (Flower William Henry)։— Կենսաբան Անգլիայում (1831—1899)։ Բնապատմական թանգարանի դիրեկտոր Հարավային Կենսինգտոնում։ Աշխատել է կաթնասունների համեմատական անատոմիայի բնագավառում և ֆիզիոլոգիայի բնագավառում։
- ↑ Պերյե (Perrier Jean-Octave-Edmond)։— Կենդանաբան Ֆրանսիայում (ծնվ. 1844 թ.)։ Գրել է բազմաթիվ աշխատություններ անողնաշարավոր կենդանիների կենդանաբանության վերաբերյալ։
- ↑ 163,0 163,1 163,2 163,3 Բյոսկ (Busk George)։— Վիրահատ, կենդանաբան և հնէաբան, Անգլիայում (1807—1886)։ Մի շարք կենդանաբանական հնէաբանական աշխատությունների հեղինակ է։
- ↑ Սմիտտ (Smitt), դոկտոր։
- ↑ Նիտշե (Nitsche Dr.)։— Կենդանաբան։
- ↑ Մորրեն (Morren Charles-Francois Antoine)։— Բնախույզ Բելգիայում (1807—1858)։ Բուսաբանության պրոֆեսոր և բուսաբանական այգու դիրեկտոր Լիեժում։ Գրել է մի շարք աշխատություններ բույսերի ֆիզիոլոգիայի, այգեգործության և գյուղատնտեսության վերաբերյալ։
- ↑ Հոֆմեյստեր (Hofmeister Wilhelm Friedrich Benedikt)։— Հայտնի բուսաբան Գերմանիայում (1824—1877)։ Բուսաբանության պրոֆեսոր Հեյդելբերգում, հետո Թյուրինգենում։ Կատարել է հետազոտություններ բույսերի համեմատական սաղմնաբանության բնագավառում, որոնց հիման վրա հաստատվել են բույսերի մոտ անսեռ ու սեռական սերունդների հերթափոխության մասին եղած պատկերացումները։
- ↑ 168,0 168,1 168,2 168,3 168,4 168,5 168,6 168,7 168,8 Հուբեր (Huber Pierre)։— Բնախույզ Շվեյցարիայում (1777—1840), որդին մեղուների կյանքը հետազոտող Ֆրանսուա Հուբերի, որին Դարվինն անվանում է Հուբեր ավագ։ Շարունակել է հոր աշխատանքը։ Գրել է մի շարք աշխատություններ միջատների՝ իշամեղուների, մրջյունների, թիթեռների կենսաբանության մասին։
- ↑ Մոցարտ (Mozart Wolfgang-Gottlieb)։— Հռչակավոր կոմպոզիտոր Ավստրիայում (1756—1791)։
- ↑ Ստ. Ջոն (St. John Charles-William George)։— Բնախույզ և սպորտսմեն Անգլիայում (1809—1856)։ Գրել է մի քանի աշխատություններ «Բնագիտություն և սպորտ» վերնագրով։
- ↑ Լերուա (Le Roy)։
- ↑ Բրենտ (Brent)։— Աղավնաբույծ։
- ↑ 173,0 173,1 Հյոտոն (Hutton)
- ↑ Մերրել (Merrell), դոկտոր։
- ↑ Մյուլլեր Ադոլֆ (Müller Adolf)։— Գերմանիայում (ծնվ. 1821 թ.)։ Իր եղբոր՝ Կարլ Մյուլլերի հետ միասին գրել է մի շարք աշխատություններ բնապատմության վերաբերյալ։
- ↑ 176,0 176,1 176,2 176,3 176,4 Ռեմզեյ (Ramsey Andrew Crombia)։— Երկրաբան Անգլիայում (1814—1891)։ Գրել է մի շարք աշխատություններ երկրաբանության վերաբերյալ
- ↑ 177,0 177,1 177,2 177,3 Սմիտ Ֆ. (Smith F.)։
- ↑ 178,0 178,1 Միլլեր (Miller)։— Երկրաչափ Անգլիայում։ Քեմբրիջի համալսարանի պրոֆեսոր։
- ↑ 179,0 179,1 Տեգետմեյեր (Tegetmeier William Bernhard)։— Բնախույզ Անգլիայում (1816— 1912)։
- ↑ Վերլո (Verlot Jean Baptiste)։— Բուսաբան Ֆրանսիայում, բուսաբանական այգու դիրեկտոր Գրենոբլեում։
- ↑ 181,0 181,1 181,2 181,3 181,4 Բետս (Bates Henry Walter)։— Բնախույզ և ճանապարհորդ Անգլիայում (ծն. 1825 թ.)։ Հետազոտել է Ամազոնը և նրա վտակները, հավաքել է արժեքավոր կենդանաբանական կոլեկցիաներ և գրել է «Բնասերը Ամազոն գետի վրա» հետաքրքրական գիրքը։ Կենդանաբանական մի քանի աշխատությունների հեղինակ է։
- ↑ 182,0 182,1 Սկոտտ (Scott Reynold)։— Բուսաբան Շոտլանդիայում, պրոֆեսոր Դուբլինում։
- ↑ Նոբլ (Noble C.)։— Այգեգործ Անգլիայում։
- ↑ Կատրֆաժ (de Quatrefages de Breau Jean Louis Armand)։— Կենդանաբան և մարդաբան, հակադարվինական, Ֆրանսիայում (1809—1891)։ Գրել է մի մեծ մենագրություն օղակավոր որդերի մասին։ Հայտնի է իր մարդաբանության վերաբերյալ աշխատություններով։ 1870 թ. լույս է տեսել նրա գիրքը «Չ. Դարվինը և նրա ֆրանսիական նախորդները»։ Լինելով «մարդու թագավորության» առանձնացածության պաշտպան, իրեն հայտարարել է Դարվինի և նրա տեսության հակառակորդ։
- ↑ 185,0 185,1 Տյուրե (Thuret Gustave-Adolphe)։— Բուսաբան Ֆրանսիայում (1817—1875)։ Գրել է մի շարք աշխատություններ ջրիմուռների և այլ ստորակարգ բույսերի մասին։ Հիմնել է կլիմայավարժեցման այգի — Villa Thuret Ֆրանսիական Ռիվերայում՝ Անտիբ քաղաքի մոտ։
- ↑ 186,0 186,1 Սաժերե (Sageret)։— Բնագետ, հիբրիդիզատոր Ֆրանսիայում։
- ↑ Տուեն (Thouin Andre)։ Բուսաբան Ֆրանսիայում (1747—1824)։
- ↑ Հյուետ'
- ↑ Սոլտեր (Salter John-William)։— Հնէաբան Անգլիայում (1820—1869)։ Հիմնական աշխատությունները վերաբերում են Բրիտանական կղզիների տրիլոբիտներին։
- ↑ 190,0 190,1 190,2 Վիխուրա Մաքս (Wichura Max)։— Բուսաբան և ճանապարհորդ Գերմանիայում (1817—1866)։
- ↑ Ժիրու-դը-Բյուզարենգը (Girou de Buzareingues)։
- ↑ Ջյուքս (Jukes Joseph-Beete)։— Երկրաբան Անգլիայում (1811—1869)։ Երկրաբանության պրոֆեսոր Դուբլինում։ Մասնակցել է դեպի Նոր Գվինեա և Ավստրալիայի արևելյան ափերը կազմակերպված գիտարշավին։
- ↑ Գեյկի (Geikie Archibald).— Հայտնի երկրաբան Շոտլանդիայում (1835—1924)։ Երկրաբանության պրոֆեսոր Էդինբուրգում։ Նրա հիմնական աշխատությունները վերաբերում են երկրաբանության բնագավառին և սառցադաշտային ժամանակաշրջանի երևույթների ազդեցության ուսումնասիրությանը Շոտլանդիայում, ինչպես նաև երրորդային ժամանակաշրջանի հրաբխային երևույթների հետազոտության բնագավառին։ Հեղինակ երկրաբանության լայնորեն տարածում ստացած դասագրքի։
- ↑ 194,0 194,1 194,2 194,3 194,4 194,5 Կրոլլ (Croll James)։— Գիտնական Շոտլանդիայում (1821—1890)։ Գրել է աշխատություններ կլիմայական պայմանների հերթափոխությունների մասին երկրաբանական ժամանակաշրջանների, մասնավորապես սառցադաշտային շրջանի ընթացքում և այդ հերթափոխությունները բացատրել է կոսմիկական պատճառներով։ Նրա տեսությունը, որ սկզբում մեծ հաջողություն է ունեցել, ներկայումս հերքված է։
- ↑ Ույտիկեր (Whitaker)։— Երկրաբան Անգլիայում։
- ↑ 196,0 196,1 196,2 Դոուսոն (Dawson John-William)։— Երկրաբան և հնէաբան Կանադայում (1820—1899)։ Նրա անունը կապված է Լավրենտյան նստվածքների բրածո կենդանիների նկարագրության հետ։ Իր աշխատություններում պաշտպանել է աստվածաբանական տեսակետները, համարելով, որ մարդը ծագել է վերջին ժամանակներում։ Հեղինակել է երկրաբանության և հնէաբանության վերաբերյալ բազմաթիվ աշխատություններ։
- ↑ 197,0 197,1 197,2 197,3 Մուրչիսոն (Murchison Roderick Impey)։— Հայտնի երկրաբան Անգլիայում (1792—1871)։ Նախագահ երկրաբանական և աշխարհագրական ընկերության Անգլիայում, իսկական անդամ Պետերբուրգի Գիտությունների ակադեմիայի (1845)։ Երկրաբանության վերաբերյալ բազմաթիվ աշխատություններից ամենից ավելի հայտնի է նրա «Սիլուրյան սիստեմը» աշխատությունը (1833)։ Ռուսաստանում կատարած ճանապարհորդության արդյունք են հանդիսանում նրա երկու աշխատությունները՝ նվիրված Ռուսաստանի հյուսիսի և Ուրալի երկրաբանության։
- ↑ 198,0 198,1 Հոպկինս (Hopkins William)։— Մաթեմատիկոս և երկրաբան Անգլիայում (1793—1866)։ Գրել է մի շարք աշխատություններ ընդհանուր երկրաբանության վերաբերյալ, այդ թվում լեռնակազմական պրոցեսների, Երկրի մակերևույթի վրա ջերմության պայմանների փոփոխությունների մասին և այլն, որոնք քննության է առել մաթեմատիկական հաշվումների հիման վրա։
- ↑ 199,0 199,1 Հումբոլդտ Ալեքսանդր (Humboldt Friedrich-Heinrich-Alexander)։— Հռչակավոր բնախույզ Գերմանիայում (1769—1859)։ 1799 թ. ձեռնարկել է մի ճանապարհորդություն դեպի Հարավային և Կենտրոնական Ամերիկա, որ տևել է հինգ տարի, հավաքած նյութերը մշակել է 20 տարում և հրատարակել է 30 հատորից բաղկացած մի աշխատություն։ Գրել է 300-ից ավելի աշխատություններ բնագիտության զանազան բնագավառների վերաբերյալ։ Հիմնադիր բույսերի աշխարհագրության՝ որպես գիտության։
- ↑ Բուե (Boue Ami)։— Երկրաբան Ֆրանսիայում (1794—1881)։ Հեղինակ մի շարք երկրաբանական աշխատությունների։
- ↑ Էշվեգ-ֆոն (Eshweg Wilhelm-Louis von)։— Լեռնային ինժեներ Գերմանիայում (1777—1855)։ 1809 թ. տեղափոխվել է Բրազիլիա, որտեղ եղել է հանքահորերի դիրեկտոր։ Հեղինակ բազմաթիվ աշխատությունների, որոնց թվումն են նաև Բրազիլիայում իր կատարած ճանապարհորդությունների նկարագրությունները։
- ↑ Ռոջերս (Н. D. Rogers)։
- ↑ Տրաուտշոլդ (Trautschold H.)։— Երկրաբան և հնէաբան Ռուսաստանում։ Հիմնական աշխատությունները վերաբերում են Մոսկվայի մարզի յուրայի և քարածխի ժամանակաշրջանների նստվածքների ստրատիգրաֆիային և հնէաբանությանը (1860)։
- ↑ Հիլգենդորֆ (Hilgendorf)։— Հնէաբան Շվեյցարիայում։
- ↑ 205,0 205,1 205,2 205,3 Վուդվորդ (Woodward John)։— Բժիշկ և բնախույզ Անգլիայում (1665—1722)։ Հրատարակել է մի շարք աշխատություններ հնէաբանության, երկրաբանության և բժշկականության բնագավառներում։
- ↑ Դ՚Օրբինի (D’Orbigny Acide Dessaline)։— Բնախույզ Ֆրանսիայում (1802—1857)։ Մասնակցել է դեպի Հարավային Ամերիկա կատարված գիտարշավին, որտեղից բերել է բազմազան հավաքածուներ։ Նրա գլխավոր աշխատությունները հնէաբանության բնագավառից են։ Կրեացիոնիստ, կողմնակից Ժ. Կյուվյեի դպրոցի, որը պաշտպանում էր այն գաղափարը, որ աստվածությունը բազմիցս ստեղծագործական փորձեր է արել տեսակառաջացման պլանը իրագործելիս։
- ↑ 207,0 207,1 207,2 207,3 207,4 207,5 207,6 207,7 207,8 Պիկտե (Pictet Franfois-Jules)։— Կենդանաբան և հնէաբան Շվեյցարիայում (1809—1872)։ Դարվինի «Տեսակների ծագումը» լույս տեսնելուց հետո դարձել է նրա տեսակետների կողմնակից, տալով իր աշխատություններում նրանց ճշտության մասին մի շարք ապացույցներ։ Գրել է բազմաթիվ աշխատություններ կենդանաբանության և հնէաբանության վերաբերյալ։
- ↑ Գոդվին-Աուստեն (Godwin-Austen Robert-Alfred-Cloyne)։ Երկրաբան Անգլիայում (1808—1884)։ Հեղինակ Անգլիայի երկրաբանության վերաբերյալ մի շարք աշխատությունների։
- ↑ 209,0 209,1 Սեջվիկ (Sedgwick Adam)։— Երկրաբան Անգլիայում (1786—1873)։ Հիմնադիր երկրաբանական ընկերության՝ Լոնդոնում, երկրաբանության պրոֆեսոր Քեմբրիջում։ Հիմնական աշխատանքները վերաբերում են Բրիտանական կղզիների պալեոզոիկ շրջանին։ Աստծու կողմից աշխարհի ստեղծման և տեսակների անփոփոխության գաղափարի կողմնակից։
- ↑ Բոսկե (Boskuet)։— Երկրաբան և հնէաբան։
- ↑ 211,0 211,1 Թոմսոն Վիլիամ (Thompson William)։— Բնախույզ Իրլանդիայում (1805—1852)։ Սկզբում զբաղվել է Իրլանդիայի թռչնաբանությամբ. հետագայում, մասնակցելով երկրաբան Ֆորբսի ճանապարհորդություններին, անցել է բնագիտության ընդհանուր հարցերի ուսումնասիրության։ Նրա հիմնական աշխատությունն է «Իրլանդիայի բնապատմությունը»։
- ↑ 212,0 212,1 212,2 212,3 212,4 212,5 212,6 212,7 Բարրանդ Իոախիմ (Barrande Ioachitn)։— Երկրաբան և հնէաբան Ավստրիայում (1799—1883)։ Հակաէվոլյուցիոնիստ, տեսակների անփոփոխելիության կողմնակից։ 1831 թ. ապրել է Պրագայում։ Գրել է Չեխիայի սիլուրի մասին կապիտալ աշխատություններ։
- ↑ Հիկս (Hicks Henry)։— Հնէաբան Անգլիայում (1837—1899)։ Կրթությամբ բժիշկ։ Գրել է մի շարք աշխատություններ Անգլիայի հնէաբանության վերաբերյալ։ Լոնդոնում երկրաբանական ընկերության նախագահ (1896—1898)։
- ↑ Լոգան (Logan William Edmond)։— Երկրաբան Անգլիայում (1798—1875)։ Աշխատել է քարածխի և Անգլիայում գտնվող քարածխային շերտերի առաջացման հարցերի ուսումնասիրությամբ։
- ↑ Էլի-դը-Բոմոն (Elie de Beaumont Jean Baptiste-Armand-Louis-Leonce)։— Երկրաբան Ֆրանսիայում, (1798—1874)։ Հեղինակ բազմաթիվ երկրաբանական ուսումնասիրությունների, այդ թվում լեռնակազմական պրոցեսների վերաբերյալ ղեկավար աշխատությունների։ Նրա ամենահայտնի երկասիրություններն են՝ «Երկրագնդի մակերևույթի վրա տեղի ունեցած մի քանի ռևոլյուցիաների ուսումնասիրությունները» (1829—1830) և «Դիտողություն լեռնային սիստեմների մասին» (1840—1852)։
- ↑ Բրուս (Bruce James)։— Ճանապարհորդ Շոտլանդիայում (1730—1799), նրա բազմաթիվ ճանապարհորդություններից գլխավոր տեղն է բռնում Հյուսիսային և Կենտրոնական Աֆրիկայում և առանձնապես Հաբեշստանում կատարած ճանապարհորդությունը։
- ↑ Վերնեյլ (Verneuil Philippe Edouard)։— Երկրաբան և հնէաբան Ֆրանսիայում (1805—1873)։ Կատարել է մի շարք ճանապարհորդություններ, այդ թվում նաև դեպի Ղրիմ և հարավային Ռուսաստան՝ զանազան երկրների երկրաբանության ծանոթանալու համար։
- ↑ Դ’Արշիակ (D’Archiac Etjenne-Jules-Adolphe-Desmier de Saint-Simon)։— Երկրաբան Ֆրանսիայում (1802—1868)։ Գրել է Ֆրանսիայի երկրաբանության վերաբերյալ բազմաթիվ աշխատություններ և «Երկրաբանության հաջողությունների պատմությունը 1834 թվականից սկսած մինչև 1851 թվականը»։
- ↑ Պրեստովիչ (Prestwich Joseph)։— Երկրաբան Անգլիայում (1812—1896)։ Հայտնի է գլխավորապես Անգլիայի էոցենյան նստվածքների ուսումնասիրությունների կապակցությամբ։ Աշխատել է մարդու երկրաբանական անցյալի վրա։
- ↑ Բոկլենդ (Buckland William)։— Հայտնի երկրաբան և հնէաբան Անգլիայում (1784—1856)։ Սկզբում Օքսֆորդի համալսարանի պրոֆեսոր, հետագայում Վեստմինստերյան աբբայության ավագ քահանա։ Մի շարք երկրաբանական և հնէաբանական աշխատությունների հեղինակ է։ Կրեացիոնիստ, փորձել է համաձայնեցնել երկրաբանական տվյալները աստվածաշնչի հետ։
- ↑ Գոդրի (Gaudry Albert)։— Հնէաբան Ֆրանսիայում (ծն. 1827 թ.)։ Հայտնի է իր կատարած հնէաբանական հետազոտություններով Հունաստանում (1855—1860) և ընդհանրապես Արևելքում։ Գրել է մի շարք աշխատություններ հնէաբանության վերաբերյալ, այդ թվում նաև Արկտիկայի բրածո կենդանիների մասին։ Իդեալիստ, վիտալիստ։
- ↑ Ժերվե (Gervais Paul)։— Կենդանաբան և հնէաբան Ֆրանսիայում (1816—1879)։ Կենդանաբանության և համեմատական անատոմիայի պրոֆեսոր սկզբում Մոնպելյեում և ապա Փարիզում՝ Սորբոնում։ Գրել է կենդանաբանության և հնէաբանության վերաբերյալ բազմաթիվ աշխատություններ, այդ թվում՝ «Կաթնասունների բնապատմությունը» երկհատոր աշխատությունը։
- ↑ Կարպենտեր (Carpenter William-Benjamin)։— Բնախույզ և բժիշկ Անգլիայում (1813—1885)։ Դատական բժշկականության պրոֆեսոր Լոնդոնում։ Գրել է կենդանաբանության և ընդհանուր ֆիզիոլոգիայի, ինչպես նաև մարդու ֆիզիոլոգիայի վերաբերյալ աշխատություններ։ Գերմանական և Միջերկրական ծովերում խորջրյա գիտարշավի ղեկավարներից մեկը։
- ↑ Կլիֆտ (Clift Williaim)։— Բնախույզ Անգլիայում (1775—1849)։ Աշխատել է ոսկրաբանության, հնէաբանության և այլ բնագավառներում։
- ↑ Լունդ (Lund P. W.)։— Հնէաբան Դանիայում։ Աշխատել է Բրազիլիայում, որտեղ գտել է հարուստ բրածո կենդանական աշխարհ, որը հիանալի կերպով նկարագրել է (1841—1845)։
- ↑ Կլաուզեն (Clausen)։— Հնէաբան։
- ↑ Կոտլի (Cautley Ptoby-Thomas)։— Երկրաբան-ինժեներ Անգլիայում (1802—1871)։ Աշխատել է Հնդկաստանում Գանգես գետի ջրանցքների կառուցման վրա, զբաղվել է Ասիայի հնէաբանությամբ։ Գրել է մի շարք աշխատություններ Հյուսիսային Հնդկաստանի բրածո կենդանական աշխարհի վերաբերյալ։
- ↑ Բերկլե (Berkeley)։
- ↑ Ջոնստոն (Johnston)։
- ↑ Մարտենս (Martens Eduard von)։— Կենդանաբան և ճանապարհորդ Գերմանիայում (1831—1904)։ Կենդանաբանական թանգարանի դիրեկտոր և պրոֆեսոր Բեռլինի համալսարանում։ Մասնակցել է դեպի Արևելյան Ասիա կատարված գիտարշավին։ Գրել է կակղամորթների և խեցեմորթների վերաբերյալ աշխատություններ։
- ↑ 231,0 231,1 Լոու (Lowe Richard-Thomas)։— Բնախույզ և ճանապարհորդ Անգլիայում։
- ↑ Ուիլ (Weale)։
- ↑ Սվեյսլանդ (Swaysland)։
- ↑ Նյուտոն (Newton Alfred)։— Կենդանաբան Անգլիայում (1829—1907)։
- ↑ 235,0 235,1 Հարտունգ Մ. (Hartung)։
- ↑ Գմելին Յոհանն-Գեորգ (Gmelin Iohann- Georg)։— Հայտնի բնախույզ և ճանապարհորդ (1709—1755)։ 1727 թ. Գերմանիայից տեղափոխվել է Ռուսաստան, որտեղ մնացել է մինչև 1747 թվականը, 1731 թ. ընտրվել է ակադեմիկոս։ 1733 թ, մասնակցել է դեպի Սիբիր կատարված գիտարշավին, որը տևել է 10 տարի։ Գիտարշավի արդյունքները հրատարակված են «Ճանապարհորդություն Սիբիրում» չորս հատորյակում, նույնպես և «Սիբիրի բուսականությունը» չորս հատորյակում։ 1747 թ. վերադարձել է Գերմանիա, որտեղ մինչև իր մահը եղել է բուսաբանության պրոֆեսոր Տյուբինգենում։
- ↑ Ռամոն (Ramond de Carbonnieres Louis François-Elisabeth)։— Քաղաքագետ և գիտնական Ֆրանսիայում (1755—1827)։ Կատարել է ճանապարհորդություն Պիրենյան լեռներում։ 1796 թ. եղել է բնապատմության պրոֆեսոր Տարբեում։ Որպես գիտնական մեծ ավանդ է մտցրել Պիրենյան լեռների երկրաբանության ուսումնասիրության տեսակետից։ Պիրենյան լեռների վերաբերմամբ կատարած դիտումները և այդ լեռների ու Ալպերի համեմատությունները նա հրատարակել է մի քանի աշխատությունների մեջ։
- ↑ Հաաստ (Haast Johann-Franz-Julius von)։— Երկրաբան Նոր Զելանդիայում (1827—1887)։ Ծագումով գերմանացի։ 1858 թ. տեղափոխվել է Նոր Զելանդիա, որտեղ մնացել է մինչև իր կյանքի վերջը։ Նելսոնում հայտնաբերել է ոսկի և քարածուխ, հետազոտել է Dinornis և Moas հանածո թռչունների բնակավայրը։ Ստեղծել է բնապատմական թանգարան Կրիստչյորչում։ Գրել է մի շարք աշխատություններ Նոր Զելանդիայի երկրաբանության վերաբերյալ։
- ↑ Հեկտոր (Hector), Դոկտոր։
- ↑ 240,0 240,1 Կլարկ Վ. Բ. (Clarke William-Branwhite)։— Երկրաբան Անգլիայում (1798—1878)։ 1839 թվականին տեղափոխվել է Ավստրալիա։ Նա դրել է Ավստրալիայի երկրաբանության ուսումնասիրության հիմքը, հայտնաբերել է ոսկու և ալմաստի հանքավայրեր։ Նրան է պատկանում բրածո թռչունների մնացորդների գյուտը։
- ↑ Գարդներ Gardner)։
- ↑ Մեն (Mann)։
- ↑ Զեեման (Seemann Berthold)։— Ճանապարհորդ և բնախույզ Գերմանիայում (1825—1871)։ 1846 թ. մասնակցել է որպես բնագետ Խաղաղ օվկիանոսում «Հերալդ» նավով կատարված գիտարշավին, որը տևել է չորս տարի։ Այցելել է Հարավային Ամերիկայի զանազան մասերը և ուսումնասիրել է Պանամայի պարանոցը, որն այն ժամանակ դեռ բոլորովին չէր հետազոտված։ Այդ ճանապարհորդության բուսաբանական արդյունքները հրատարակվեի են 1752—1757 թվականներին։
- ↑ Վալյանսեն (Valenciennes Achille)։— Կենդանաբան Ֆրանսիայում (1794—1865)։ Լամարկի օգնականը, խմբագրել է Լամարկի երկերը նրա կուրանալուց հետո։ Հետագայում պրոֆեսոր կենդանաբան։ Գրել է բազմաթիվ աշխատություններ, որոնցից ամենից ավելի նշանակալի է 11 հատորից բաղկացած «Ձկների պատմությունը»։
- ↑ Ջոնս Դ. Մ. (J. M. Jones)։
- ↑ Բորի Սեն-Վենսեն (Boryde Saint-Vincent)։— Բնախույզ, Ֆրանսիայում (1780—1846)։ Հետազոտել է Ատլանտյան և Հնդկական օվկիանոսների այն կղզիները, որոնք հարակից են Աֆրիկային։ Մասնակցել է բազմաթիվ գիտական հանդեսների, խմբագրել է «Բնապատմության դասական բառարանը»։
- ↑ Տոմս (Tomes)։
- ↑ Իրլ Ուինձոր (Windsor Earl)։
- ↑ Գուլդ Ա. Ա. (Gould Augustus A.)։— Ականավոր մալակոալոգ Ամերիկայում (մեռել է 1866 թ.)։ Շատ է աշխատել հյուսիս-ամերիկյան թերթախռիկավոր կակղամորթների սիստեմատիկայի վրա։
- ↑ Աուկեպտեն (Aucapitaine)։ Կակղամորթների մասնագետ։
- ↑ 251,0 251,1 Բրոուն Ռոբերտ (Brown Robert)։— Հռչակավոր բուսաբան Անգլիայում (1781—1858)։ 1801 թ. սկսած մինչև 1805 թվականը մասնակցել է կապիտան Ֆլինդերի դեպի Ավստրալիա կատարած գիտարշավին։ Հեղինակ «Նոր Հոլանդիայի բուսականությունը» աշխատության։ 1827 թ. նշանակվել է բրիտանական թանգարանի բուսաբանական բաժանմունքի դիրեկտոր։ 1849 թվականից Լիննեյան ընկերության նախագահ Լոնդոնում։ Հետազոտել է բուսական բջիջների կորիզը և բույսերի բեղմնավորման առաջին ստադիաներըp որոնց հետևել է նա մինչև այն մոմենտը, երբ ծաղկափոշու խողովակը թափանցում է սերմնաբողբոջի մեջ։
- ↑ Ռիշար (Richard Louis-CIaude-Marie)։— Բուսաբան Ֆրանսիայում (1754—1821)։ Կատարել է ճանապարհորդություններ ֆրանսիական Գվիանայում, Անտիլյան կղզիներում և Մեքսիկական ծովածոցի կղզիներում։ 1789 թվականին վերադարձել է Ֆրանսիա և իր հետ բերել է նոր և հազվագյուտ բույսերի մեծ հավաքածուներ։ Գրել է մի շարք բուսաբանական աշխատություններ։
- ↑ Տեմմինկ (Temminck C. I.)։— Կենդանաբան Հոլանդիայում (1770—1858)։ Հիմնական աշխատությունները վերաբերում են թռչնաբուծության։
- ↑ Ստենստրուպ (Steenstrup Johann Jabet Smitt)։— Կենդանաբան և հնագետ Դանիայում (1813—1897)։ Հայտնի է դարձել սերունդների հերթափոխության հարցի, ինչպես նաև գլխոտանի կակղամորթների և ստորակարգ մակաբույծ խեցգետինների վերաբերյալ աշխատություններով։
- ↑ Մեկլեյ (Macleay William Sharp)։— Բուսաբան և կենդանաբան Ավստրալիայում (1792—1865)։
- ↑ Տրայմեն (Trimen Henry)։— Բուսաբան Անգլիայում (1843—1896)։
- ↑ Հեկկել (Haeckel Ernst-Heinrich)։— Հռչակավոր կենդանաբան-դարվինիստ Գերմանիայում (1834—1919)։ Կենդանաբանության պրոֆեսոր Ենայում։ Գրել է կենդանաբանության և օրգանիզմների էվոլուցիայի վերաբերյալ աշխատություններ։ Դարվինի «Տեսակների ծագումը» լnւյu տեսնելուց հետո դարձել է նրա ջերմ երկրպագու և դարվինիզմի քարոզիչ Գերմանիայում և ընդհանրապես Եվրոպայում։ Նրա գլխավոր երկասիրությունն է «Աշխարհի ստեղծման բնապատմությունը», որ հաստատում է կենդանիների տոհմաբանական ծառը։ Նա գրել է նաև «Անտրոպոգենիան» և «Համաշխարհային հանելուկներ»։ Գիտության մեջ Հեկկելի ընդհանուր դերի բնութագիրը տես Վ. Ի. Լենինի «Մատերիալիզմ և էմպիրիոկրիտիցիզմ» գրքում և Էնգելսի «Բնության դիալեկտիկան» գրքում, Ի. Ի. Մեչնիկովի «Տեսակների ծագման հարցի պատմության ուրվագծում»։
- ↑ Գրիմմ (Grimm)։
- ↑ 259,0 259,1 Լեյբնից (Leibnitz Gottfroy-Wilhelm)։— Հռչակավոր փիլիսոփա իդեալիստ (մոնադների տեսությունը) և մաթեմատիկոս Գերմանիայում (1646—1716)։ Գտել է դիֆերենցիալ և ինտեգրալ հաշիվը։ Սկիզբ է դրել լայն տարածված այն գաղափարին, որ բնության մեջ թռիչքներ չկան։
- ↑ Նյուտոն Իսահակ (Newton Isaac)։— Մեծ մաթեմատիկոս, աստղագետ և ֆիզիկոս Անգլիայում (1642—1727)։ Հաստատել է տիեզերական ձգողականության օրենքը։ Նրա գլխավոր աշխատությունը՝ «Բնական փիլիսոփայության մաթեմատիկական հիմունքները», կազմել է դասական մեխանիկայի հիմքը, որը հետագայում ստացել է փայլուն տեսական զարգացում և գործնական կիրառություն։
- ↑ Մոպերտյուի (Maupertuis Pierre-Louis Moreau de)։— Երկրաչափ և փիլիսոփա Ֆրանսիայում (1698—1759)։